Анхиорнис
| Анхиорнис Временной диапазон: поздняя юра ( оксфорд ),
| |
|---|---|

| |
| Ископаемый образец выставлен в Пекинском музее естественной истории. | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Клэйд : | Динозаврия |
| Клэйд : | Саурисхия |
| Клэйд : | Теропода |
| Клэйд : | Паравес |
| Семья: | † Анхиорнитиды |
| Род: | † Анхиорнис Сюй и др. , 2009 г. |
| Разновидность: | † А. Хаксли
|
| Биномиальное имя | |
| † Anchiornis huxleyi Сюй и др. , 2009 г.
| |
| Синонимы | |
| |
Anchiornis — род небольших четырёхкрылых паравианских динозавров , известный только один вид — типовой вид Anchiornis huxleyi , названный в честь сходства с современными птицами. Латинское название Anchiornis происходит от греческого слова, означающего «около птицы», а huxleyi относится к Томасу Генри Хаксли , современнику Чарльза Дарвина .
Окаменелости Anchiornis были найдены только в формации Тяоцзишань в провинции Ляонин , Китай, в породах, датируемых поздней юрой , около 160 миллионов лет назад. Он известен по сотням экземпляров, и, учитывая превосходную сохранность некоторых из этих окаменелостей, он стал первым мезозойским видом динозавров, для которого можно было определить почти весь жизненный облик, и важным источником информации о ранней эволюции птиц. [2]
Открытие и история
[ редактировать ]
Первая известная окаменелость Anchiornis (его типовой образец ) была раскопана в районе Яолугоу округа Цзяньчан , Ляонин , Китай . Эти породы трудно датировать, но большинство исследований пришли к выводу, что они принадлежат к формации Тяоцзишань , состоящей из пород, датируемых поздним юрским периодом ( оксфордский возраст), возрастом от 160,89 до 160,25 миллионов лет. [1] [3] [4] Anchiornis был изучен и описан палеонтологом Сюй Сином и его коллегами в статье, принятой в Китайский научный бюллетень в 2009 году. В настоящее время образец находится в коллекции Института палеонтологии и палеоантропологии позвоночных под каталожным номером IVPP V14378. Это сочлененный скелет, в котором отсутствуют череп, часть хвоста и правая передняя конечность. Название Anchiornis huxleyi было выбрано Сюй и его коллегами в честь Томаса Генри Хаксли , одного из первых сторонников биологической эволюции и одного из первых, кто предположил тесную эволюционную связь между птицами и динозаврами . Родовое название Anchiornis происходит от сочетания древнегреческих слов, означающих «близость» и «птица», поскольку оно интерпретировалось как важное для заполнения пробела в переходе между строениями тела птиц и динозавров. [5]
Второй экземпляр был обнаружен примерно в то же время и передан группе ученых из Шэньянского педагогического университета местным фермером, когда они проводили раскопки на месте раскопок Яолугоу. По словам фермера, этот второй экземпляр был найден неподалеку в районе Даксишань, также в породах формации Тяоцзишань примерно того же возраста, что и первый Anchiornis . Два учёных посетили это место, чтобы сравнить новое ископаемое с найденными там типами горных пород, и смогли подтвердить, что новый образец, вероятно, действительно прибыл из района, описанного фермером. Им удалось выкопать несколько окаменелостей рыб и третье ископаемое Anchiornis . Окаменелость фермера подверглась исследованию, которое было опубликовано 24 сентября 2009 года в журнале Nature . Ему был присвоен каталожный номер LPM — B00 169 в Ляонинском палеонтологическом музее . Он крупнее и гораздо более полный, чем первый экземпляр, и сохранил длинные перья на кистях, руках, ногах и ступнях, что указывает на то, что это был четырехкрылый динозавр, похожий на Микрораптор . [6]

Хотя подробно описано лишь несколько экземпляров, многие другие были идентифицированы и хранятся как в частных коллекциях, так и в музеях. Об одном из них, почти полном скелете без хвоста, а также сохранивших обширные остатки перьев, сообщалось в 2010 году. Эта окаменелость также свидетельствует о том, что у Анхиорниса был оперенный гребень на голове, и она использовалась для определения жизненной окраски животного. Он хранится в Пекинском музее естественной истории под номером экземпляра BMNHC PH828. [2] Другой экземпляр из того же карьера окаменелостей, что и типовой образец, был найден местным дилером по ископаемым и продан Парку ископаемых и геологии Ичжоу и внесен в каталог как YTGP-T5199. Это ископаемое, почти полный скелет, было подготовлено и изучено учеными Геологического парка и идентифицировано как Anchiornis . Затем его использовали для с помощью сканирующего электронного микроскопа исследования микроструктуры перьев Anchiornis . В ходе исследования также были изучены хорошо сохранившиеся меланосомы перьев, чтобы определить их цвет. Ученые, участвовавшие в исследовании, обнаружили, что окраска, обнаруженная для этого образца, отличалась от цвета, зарегистрированного для BMNHC PH828, и отметили, что образец BMNHC на самом деле не может быть Anchiornis , поскольку он был описан до того, как аналогичные виды из той же формации были обнаружены. был обнаружен. [7]
в Шаньдунском музее природы Тяньюй в уезде Пинъи , Китай, было 255 экземпляров Anchiornis . Например, в 2010 году [8] Среди их коллекции есть очень хорошо сохранившаяся окаменелость с видимыми цветными узорами, занесенная в каталог как STM 0-214. Хотя этот экземпляр еще не полностью описан, он был сфотографирован для статьи в National Geographic использовался при исследовании кроющих перьев и анатомии крыльев Anchiornis . в 2011 году и в следующем году [9]
Описание
[ редактировать ]
Anchiornis huxleyi был небольшим двуногим тероподом динозавром- с треугольным черепом, имеющим несколько общих деталей с дромеозавридами , троодонтидами и примитивными авиаланами . Как и другие ранние паравианы, Анхиорнис был небольшим, размером с ворону . [10] У него были длинные руки с крыльями, длинные ноги и длинный хвост. Как и все паравианы, он был покрыт перьями, хотя на некоторых частях тела имел чешую. Крылья, ноги и хвост поддерживали длинные, но относительно узкие перья с лопастями. два типа более простых ( перистых Остальную часть тела покрывали пушистых ) перьев : пуховые перья, состоящие из нитей, прикрепленных к их основаниям, и более сложные пуховые перья с зазубринами, прикрепленными вдоль центрального пера. Длинные простые перья покрывали почти всю голову и шею, туловище, верхнюю часть ног и первую половину хвоста. Остальная часть хвоста имела перистые рулевые перья (прямоугольники). [3] Длинные перья на голове (макушке) могли образовывать гребень. [2] Хотя у первого экземпляра Anchiornis сохранились лишь слабые следы перьев вокруг сохранившейся части тела, с тех пор было обнаружено гораздо больше хорошо сохранившихся окаменелостей. Исследования образцов Anchiornis с использованием лазерной флуоресценции выявили не только более подробную информацию о перьях, но также о коже и мышечной ткани. В совокупности эти данные дали ученым почти полную картину анатомии Anchiornis . Дополнительные исследования показывают, что у Anchiornis оперение тела состояло из коротких игл с длинными независимыми гибкими зазубринами. Эти зазубрины торчали из игл под низким углом на двух противоположных лезвиях. Это также придало каждому перу общую раздвоенную форму и привело к тому, что теропод имел более мягкую текстуру и «лохматое» оперение, чем у современных птиц. Контурные перья «лохматого цвета», вероятно, влияли на терморегуляторные и водоотталкивающие способности и в сочетании с перьями крыльев с открытыми лопастями снижали аэродинамическую эффективность. [11] [12]
Голотип, принадлежащий полувзрослому или молодому человеку, имел длину 34 сантиметра (1,12 фута) и весил 110 граммов (3,9 унции). [5] Общая длина тела самых крупных экземпляров составляла 60–62 сантиметра (1,97–2,03 фута). [13] размах крыльев почти 57,4 сантиметра (1,88 фута) и вес около 0,6–1 килограмма (1,3–2,2 фунта). [14] [а]
Крылья
[ редактировать ]
Как и у других ранних паравианцев , у Анхиорниса были большие крылья, состоящие из перистых перьев, прикрепленных к руке и кисти. Крыло Anchiornis состояло из 11 первичных и 10 второстепенных перьев . Первичные перья Anchiornis были примерно такой же длины, как и второстепенные, и образовывали крыло округлой формы. Перья крыльев имели изогнутые, но симметричные центральные перья, небольшие и тонкие относительные размеры и закругленные кончики, что указывает на плохую аэродинамическую способность. У родственных динозавров Microraptor и Archeopteryx самые длинные перья крыльев располагались ближе всего к кончику крыла, из-за чего крылья казались относительно длинными и заостренными. Однако у Anchiornis самые длинные перья крыла находились ближе всего к запястью, что делало крыло самым широким посередине и сужалось к кончику, создавая более округлый и менее приспособленный к полету профиль. [3]
Как и у других манирапторов , Анхиорниса был пропатагиум у — лоскут кожи, соединяющий запястье с плечом и закругляющий передний край крыла. У Anchiornis эта часть крыла была покрыта кроющими перьями , которые сглаживали крыло и закрывали промежутки между более крупными первичными и вторичными перьями. Однако, в отличие от современных птиц , кроющие перья Anchiornis не располагались полосками или рядами. [6] Расположение кроющих перьев у Anchiornis также было более примитивным , чем у птиц и более продвинутых паравиан. У современных птиц кроющие обычно покрывают только верхнюю часть крыла, при этом большая часть поверхности крыла состоит из непокрытых маховых перьев. С другой стороны, у некоторых окаменелостей Anchiornis несколько слоев кроющих перьев, по-видимому, простираются вниз и покрывают большую часть поверхности крыла, так что крыло по существу состоит из нескольких слоев перьев, а не из слоя широких перьев, состоящих только из их частей. основания скрыты слоями кроющих покровов. Такое многослойное расположение крыльев могло помочь укрепить крыло, учитывая, что сами первичные и вторичные перья были узкими и слабыми. [9]
На крыле было три когтистых пальца; однако, в отличие от некоторых более примитивных теропод, два самых длинных пальца не были отдельными, а были связаны кожей и другими тканями, образующими крыло, поэтому Anchiornis был функционально двупалым. [6] Эти связанные пальцы были включены в постпатагий, или лоскут кожи и других тканей, которые помогали поддерживать основания основных перьев крыла. Как и пальцы ног, кожа вокруг нижней части пальцев была покрыта крошечными округлыми чешуйками. В отличие от пальцев ног, плоть вокруг нижней стороны костей пальцев была вдвое толще самих костей и не имела отчетливых подушечек; вместо этого пальцы были прямыми и гладкими, без каких-либо серьезных складок в суставах. Чешуя и кожа вокруг пальцев очень редко сохраняются в окаменелостях ранних пеннарапторанов , единственными заметными исключениями являются Anchiornis и Caudipteryx , у которых были похожие толстые чешуйчатые пальцы, связанные с крыльями. [6]
Ноги
[ редактировать ]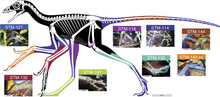
Помимо передних крыльев, у Анхиорниса были длинные перья на задних лапах. Это побудило многих учёных назвать Анхиорниса четырёхкрылым динозавром , наряду с такими животными, как Микрораптор и Сапеорнис . [16] Однако перья на задних лапах Anchiornis не имели формы и расположения, ожидаемых от маховых перьев, и вполне вероятно, что их основная роль заключалась в демонстрации, а не в полете. [17]
У Anchiornis были очень длинные ноги, что обычно указывает на то, что животное хорошо бегает. Однако обширные перья на ногах указывают на то, что это может быть рудиментарной чертой, поскольку у бегающих животных волосы или перья на ногах, как правило, уменьшаются, а не увеличиваются. [3] Как и у большинства паравианцев, у Анхиорниса было четыре пальца на ноге, причем третий и четвертый пальцы были самыми длинными. Первый палец, или Hallux , не был перевернут, как у сидячих видов. [3] Задние крылья Anchiornis также были короче, чем у Microraptor , и состояли из 12–13 маховых перьев, прикрепленных к голени (голень) и от 10 до 11 к предплюсне (верхняя часть стопы). Также в отличие от Microraptor , перья задних крыльев были самыми длинными ближе к телу, а перья на лапах были короткими и направлены вниз, почти перпендикулярно костям стопы. [3]
В отличие от многих других паравианов, ноги Анхиорниса (за исключением когтей) были полностью покрыты перьями, хотя они были намного короче, чем те, что составляют заднее крыло. [3] У некоторых экземпляров сохранились чешуи на пальцах ног, лапках и даже голени (голенях), что позволяет предположить, что чешуя существовала под перьями. Нижняя сторона пальцев представляла собой мясистые подушечки с отчетливыми складками в суставах. Подушечки лап были покрыты мелкими, похожими на гальку чешуйками. Чешуя также присутствовала на верхней части ступней, но ее очень трудно увидеть во всех известных окаменелостях. [6]
Цвет
[ редактировать ]
В 2010 году группа ученых исследовала многочисленные точки среди перьев чрезвычайно хорошо сохранившегося экземпляра Anchiornis в Пекинском музее естественной истории, чтобы изучить распределение меланосом — пигментных клеток, которые придают перьям их цвет. Изучая типы меланосом и сравнивая их с таковыми у современных птиц, ученые смогли составить карту конкретных цветов и рисунков, присутствующих у этого Anchiornis , когда он был жив. Хотя этот метод использовался и описывался для отдельных перьев птиц и частей других динозавров (таких как хвост Sinosauropteryx ), Anchiornis стал первым мезозойским динозавром, для которого была известна почти вся прижизненная окраска (обратите внимание, что хвост этого экземпляра был не сохранилось). [2] Исследование показало, что большинство перьев на теле этого экземпляра Anchiornis были серыми и черными. Перья на макушке были в основном рыжими с серым основанием и передней частью, а на лице были рыжие крапинки среди преимущественно черных перьев на голове. Перья передних и задних крыльев белые с черными кончиками. Кроющие (более короткие перья, покрывающие основания длинных перьев крыльев) были серыми, контрастируя с преимущественно белыми основными крыльями. Более крупные кроющие крыла также были белыми с серыми или черными кончиками, образующими ряды более темных точек вдоль середины крыла. Они имели форму темных полос или даже рядов точек на внешнем крыле (первичные кроющие перья), а также в виде более неравномерного набора крапинок на внутреннем крыле (вторичные кроющие перья). Голяшки ног были серыми, за исключением длинных перьев задних крыльев, а ступни и пальцы ног были черными. [2]
В 2015 году вторая окаменелость Anchiornis в Парке ископаемых и геологии Ичжоу подверглась аналогичному исследованию, которое включало изучение форм меланосом во всех перьях. В отличие от исследования 2010 года были обнаружены только меланосомы серо-черного типа. Даже когда были исследованы коронные перья, ни одной из более круглых меланосом рыжего типа не было обнаружено. Ученые, проводившие это второе исследование, предложили несколько возможных объяснений этого несоответствия. Во-первых, различная сохранность меланосом или разные методы исследования могли повлиять на результаты первоначального исследования. Во-вторых, поскольку экземпляр из Пекинского музея был меньше, вполне возможно, что рыжий цвет сменился по мере старения этих животных. В-третьих, возможно, существовали региональные различия или даже разные виды Anchiornis , которые имели разную окраску оперения. [7]
Классификация
[ редактировать ]
| |||||||||||||||||||||||||||||||||
| Кладограмма по результатам филогенетического исследования Lefèvre et al. , 2017. [18] |
Когда он был впервые обнаружен, учёные, изучавшие Anchiornis, провели филогенетический анализ и пришли к выводу, что он был ранним членом группы Avialae , наряду с археоптериксом . [5] Представители Avialae, называемые авиаланами, все более тесно связаны с современными птицами, чем с динозаврами -дромеозавридами и троодонтидами , хотя самые ранние и наиболее примитивные представители всех трех групп чрезвычайно похожи друг на друга, что затрудняет точное разделение. к какой из этих трех основных паравианских ветвей они принадлежат. [10]
Второй экземпляр Anchiornis был более полным, чем первый, и сохранил несколько особенностей, которые побудили Ху Дунъюя и его коллег реклассифицировать Anchiornis как троодонтида. [3] Еще несколько исследований с использованием аналогичного анализа также показали, что Anchiornis является троодонтидом, хотя были и исключения. Одно исследование показало, что Anchiornis является членом Archaeopterygidae и вместе с археоптериксом считался более примитивным, чем дромеозавриды, троодонтиды или авиаланы. В 2015 году Санкар Чаттерджи поместил Anchiornis вместе с Microraptor и другими четырехкрылыми паравианами в группу, которую он назвал «Tetrapterygidae», недалеко от Avialae, хотя это не было подтверждено филогенетическим анализом. [19] Более комплексные исследования показали, что Анхиорнис все-таки мог быть авиаланом. [20] хотя новые находки и обновленные версии того же исследования позже изменили этот вывод, сделав вывод, что Anchiornis, скорее всего, был базальным членом клады Paraves, сразу за пределами клады, в которую входят дромеозавриды, троодонтиды и авиаланы. [18]
В ходе повторной оценки археоптерикса образца из Харлема в 2017 году было обнаружено, что Anchiornis находится в группе с другими родами, такими как Eosinopteryx , Xiaotingia , и был помещен в семейство Anchiornithidae вместе с другими родственниками. [21] [22]
Палеобиология
[ редактировать ]
Anchiornis примечателен своими пропорционально длинными передними конечностями, которые составляют 80% от общей длины задних конечностей. Это похоже на состояние ранних птиц, таких как археоптерикс , и авторы отметили, что для полета необходимы длинные передние конечности. У Anchiornis также было более птичье запястье, чем у других нептичьих теропод. мог Первоначально авторы предполагали, что Анхиорнис летать или планировать. Однако дальнейшие находки показали, что крылья Anchiornis , хотя и были хорошо развиты, были короткими по сравнению с более поздними видами, такими как Microraptor , с относительно короткими первичными перьями с закругленными, симметричными кончиками, в отличие от заостренных перьев аэродинамических пропорций Microraptor . [3] Исследование потенциальных летных качеств ранних паравианцев, проведенное в 2016 году, пришло к выводу, что, хотя молодые экземпляры Anchiornis, возможно, были способны использовать свои крылья для подъема по склону и, возможно, могли достичь машущего полета, если использовался взмах крыльев под очень большим углом, более крупные взрослые особи не получили бы никакой аэродинамической выгоды от своих крыльев — они были просто слишком тяжелыми по сравнению с общей площадью их крыльев. То же исследование показало, что взмахи крыльями во время бега привели бы к небольшому (10%) увеличению скорости бега. Аналогично, использование крыльев во время прыжка привело бы к увеличению высоты и расстояния на 15–20%. [15] Примечательно, что у Anchiornis , по-видимому, отсутствовала грудина ( грудина ), которая могла состоять из хряща, а не из кости, как у более примитивных теропод. [23] [24]

Пропорции задних ног Anchiornis больше напоминают пропорции более примитивных динозавров-теропод, чем птиц, а длинные ноги указывают на быстрый образ жизни. Однако, хотя длинные ноги обычно указывают на быстрого бега, ноги и даже ступни и пальцы Анхиорниса были покрыты перьями, в том числе длинными перьями на ногах, похожими на перья на задних крыльях Микрораптора . [3] Длинные перья на голенях ног, возможно, замедляли скорость бега Анхиорниса . У современных птиц, особенно у тех, которые живут на земле, на голенях наблюдается уменьшение или даже потеря перьев. [3] Задние крылья Anchiornis были меньше и состояли из более изогнутых и симметричных перьев, чем у Microraptor , что позволяет предположить, что они использовались в основном для демонстрации, а не для полета. [17] Тем не менее, они все равно могли предоставить животному какое-то аэродинамическое преимущество, даже если их основной целью была демонстрация или какая-то другая функция. [10] [25]
Скелетная структура Anchiornis похожа на Eosinopteryx , который, судя по всему, был способным бегуном из-за неизогнутых задних когтей и отсутствия маховых перьев на хвосте и голенях. Anchiornis имел схожее строение тела и ту же экосистему, что и Eosinopteryx , что предполагает разные ниши и сложную картину происхождения полета. [26] [27]
Как и многие современные птицы, Anchiornis демонстрировал сложную окраску с пестрыми узорами разного цвета по телу и крыльям, или «окраску оперения внутри и между перьями». У современных птиц такой цветовой рисунок используется для общения и демонстрации либо представителям одного и того же вида (например, для спаривания или демонстрации территориальной угрозы), либо для угрозы и предупреждения конкурирующих или хищных видов. [2]
Кормление
[ редактировать ]В исследовании 2018 года сообщалось, что желудочные гранулы связаны с Anchiornis образцами ; некоторые из анхиорнисов даже сохранились с гранулами внутри тел. Anchiornis — самый ранний теропод, который, как известно, производил погадки. Гранулы содержали кости ящерицы и чешую птихолепидных рыб. [28]
См. также
[ редактировать ]Примечания
[ редактировать ]- ^ Оценка размаха крыльев основана на уравнении, представленном в исследовании Dececchi et al. (2016), в котором говорится, что размах крыльев паравианцев, участвовавших в исследовании, в 2,1 раза превышал длину крыла. [15] В случае Anchiornis один из крупнейших экземпляров PKUP V1068 имел длину крыла более 27,3 сантиметра (0,90 фута), в результате чего размах крыльев составлял почти 57,4 сантиметра (1,88 фута). [14]
Ссылки
[ редактировать ]- ^ Jump up to: а б Чу, З.; Он, Х.; Рамезани Дж.; Боуринг, ЮАР; Ху, Д.; Чжан, Л.; Чжэн, С.; Ван, X.; Чжоу, З.; Дэн, К.; Го, Дж. (2016). «Высокоточная U-Pb геохронология юрской биоты Янляо из Цзяньчана (западная провинция Ляонин, Китай): возрастные ограничения на появление пернатых динозавров и млекопитающих» . Геохимия, геофизика, геосистемы . 17 (10): 3983. Бибкод : 2016GGG....17.3983C . дои : 10.1002/2016GC006529 .
- ^ Jump up to: а б с д и ж Ли, К.; Гао, К.-К.; Винтер, Дж.; Шоки, доктор медицины; Кларк, Дж.А.; д'Альба, Л.; Мэн, К.; Бриггс, DEG; Прум, Р.О. (2010). «Цвет оперения вымершего динозавра» (PDF) . Наука . 327 (5971): 1369–1372. Бибкод : 2010Sci...327.1369L . дои : 10.1126/science.1186290 . ПМИД 20133521 . S2CID 206525132 .
- ^ Jump up to: а б с д и ж г час я дж к Ху, Д.; Хоу, Л.; Чжан, Л.; Сюй, X. (2009). «Троодонтидный теропод до археоптерикса из Китая с длинными перьями на плюсне». Природа . 461 (7264): 640–643. Бибкод : 2009Natur.461..640H . дои : 10.1038/nature08322 . ПМИД 19794491 . S2CID 205218015 .
- ^ Лю, Ю.-К.; Куанг, Х.-В.; Цзян, X.-J.; Пэн, Н.; Сюй, Х.; Сунь, Х.-Ю. (2012). «Время появления самых ранних известных пернатых динозавров и переходных птерозавров старше биоты Джехол». Палеогеография, Палеоклиматология, Палеоэкология . 323–325: 1–12. Бибкод : 2012PPP...323....1L . дои : 10.1016/j.palaeo.2012.01.017 .
- ^ Jump up to: а б с Сюй, Х; Чжао, К.; Норелл, М.; Салливан, К.; Хон, Д.; Эриксон, Г.; Ван, X.; Хан, Ф.; Го, Ю. (2009). «Новая окаменелость пернатого динозавра-манираптора, которая заполняет морфологический пробел в птичьем происхождении». Китайский научный бюллетень . 54 (3): 430–435. Бибкод : 2009SciBu..54..430X . дои : 10.1007/s11434-009-0009-6 .
- ^ Jump up to: а б с д и Ван, X.; Питтман, М.; Чжэн, X.; Кэй, Т.Г.; Фальк, Арканзас; Хартман, SA; Сюй, X. (2017). «Базальная паравиальная функциональная анатомия, освещенная высокодетализированными контурами тела» . Природные коммуникации . 8 : 14576. Бибкод : 2017NatCo...814576W . дои : 10.1038/ncomms14576 . ПМЦ 5339877 . ПМИД 28248287 .
- ^ Jump up to: а б Линдгрен, Дж.; Шёвалл, П.; Карни, РМ; Чинкотта, А.; Раньше П.; Хатчесон, Юго-Запад; Густафссон, О.; Улисс Лефевр, Ю.; Франсуа Эскуилье, Ф.; Хеймдал, Дж.; Андерс Энгдаль, А.; Грен, Дж.А.; Кир, BP; Вакамацу, К.; Янс, Дж.; Годфруа, П. (2015). «Молекулярный состав и ультраструктура перьев юрских паравианцев» . Научные отчеты . 5 : 13520. Бибкод : 2015NatSR...513520L . дои : 10.1038/srep13520 . ПМК 4550916 . ПМИД 26311035 .
- ^ «В Китае находится самый большой в мире музей динозавров» . Рассвет . 4 августа 2010 г. Архивировано из оригинала 28 октября 2010 г. Проверено 6 ноября 2010 г.
- ^ Jump up to: а б Лонгрич, Северная Каролина; Винтер, Дж.; Мэн, К.; Ли, К.; Рассел, AP (2012). «Примитивное расположение перьев крыльев у Archeopteryx Lithographica и Anchiornis huxleyi » . Современная биология . 22 (23): 2262–2267. дои : 10.1016/j.cub.2012.09.052 . ПМИД 23177480 .
- ^ Jump up to: а б с Витмер, LM (2009). «Палеонтология: Пернатые динозавры в клубке» . Природа . 461 (7264): 601–602. Бибкод : 2009Natur.461..601W . дои : 10.1038/461601a . ПМИД 19794481 . S2CID 205049989 .
- ^ «Пернатые динозавры оказались даже пушистее, чем мы думали» . sciencedaily.com .
- ^ Сайтта, Эван Т.; Гелернтер, Ребекка; Винтер, Якоб (2018). «Дополнительные сведения о примитивном контуре и оперении крыльев паравианских динозавров» . Палеонтология . 61 (2): 273–288. Бибкод : 2018Palgy..61..273S . дои : 10.1111/пала.12342 . hdl : 1983/61351c6d-1517-4101-bac8-50cbb733761d . S2CID 56104335 .
- ^ Пей, Р.; Ли, К.; Мэн, К.; Норелл, Марк А.; Гао, К.-К. (2017). «Новые экземпляры Anchiornis huxleyi (Theropoda: Paraves) из поздней юры Северо-Восточного Китая» . Бюллетень Американского музея естественной истории . 411 (411): 1–67. дои : 10.1206/0003-0090-411.1.1 . S2CID 90650697 .
- ^ Jump up to: а б Дечекки, штат Калифорния; Ларссон, HCE; Питтман, М.; Хабиб, МБ (2020). «11 — Высокий полет или высокая мода? Сравнение летного потенциала среди паравианцев с маленьким телом» . В Питтмане, М.; Син, X. (ред.). Пеннарапторские динозавры-тероподы: прошлый прогресс и новые рубежи . Бюллетень Американского музея естественной истории. стр. 295–320. дои : 10.1206/0003-0090.440.1.1 . S2CID 221692656 – через Библиотеку наследия биоразнообразия.
- ^ Jump up to: а б Дечекки, штат Калифорния; Ларссон, HCE; Хабиб, МБ (2016). «Крылья перед птицей: оценка локомоторных гипотез, основанных на взмахах рук, у предшественников птиц» . ПерДж . 4 : е2159. дои : 10.7717/peerj.2159 . ПМЦ 4941780 . ПМИД 27441115 .
- ^ Сюй, Х.; Чжоу, З.; Дадли, Р.; Макем, С.; Чуонг, CM; Эриксон, генеральный директор; Варриккио, диджей (2014). «Интегративный подход к пониманию происхождения птиц» . Наука . 346 (6215): 1253293. doi : 10.1126/science.1253293 . ПМИД 25504729 . S2CID 24228777 .
- ^ Jump up to: а б О'Коннор, Дж. К.; Чанг, Х. (2015). «Перья задних конечностей у паравианов: прежде всего «крылья» или украшения?». Биологический вестник . 42 (7): 616–621. дои : 10.1134/s1062359015070079 . S2CID 15637177 .
- ^ Jump up to: а б Лефевр, Улисс; Кау, Андреа; Чинкотта, Од; Ху, Дунъюй; Чинсами, Анусуя; Эскуийе, Франсуа; Годфруа, Паскаль (2017). «Новый юрский теропод из Китая документирует переходный этап в макроструктуре перьев». Наука о природе . 104 (9–10): 74. Бибкод : 2017SciNa.104...74L . дои : 10.1007/s00114-017-1496-y . ПМИД 28831510 . S2CID 32780661 .
- ^ Чаттерджи, С. (2015). Возвышение птиц: 225 миллионов лет эволюции (2-е изд.). Издательство Университета Джонса Хопкинса. ISBN 978-1-4214-1590-1 .
- ^ Годфруа, П.; Кау, А.; Донг-Ю, Х.; Эскуилье, Ф.; Вэньхао, В.; Дайк, Г. (2013). «Юрский птичий динозавр из Китая раскрывает раннюю филогенетическую историю птиц». Природа . 498 (7454): 359–362. Бибкод : 2013Natur.498..359G . дои : 10.1038/nature12168 . ПМИД 23719374 . S2CID 4364892 .
- ^ Фот, К.; Раухут, OWM (2017). «Переоценка харлемского археоптерикса и радиации динозавров-теропод манирапторанов» . Эволюционная биология BMC . 17 (1): 236. doi : 10.1186/s12862-017-1076-y . ПМК 5712154 . ПМИД 29197327 .
- ^ Кау, А.; Бейранд, В.; Воетен, Д.; Фернандес, В.; Таффоро, П.; Штейн, К.; Барсболд, Р.; Цогтбаатар, К.; Карри, П.; Годфруа, П. (2017). «Синхротронное сканирование выявило экоморфологию земноводных в новой кладе птицеподобных динозавров». Природа . 552 (7685): 395–399. Бибкод : 2017Natur.552..395C . дои : 10.1038/nature24679 . ПМИД 29211712 . S2CID 4471941 .
- ^ Чжэн, X.; О'Коннор, Дж.; Ван, X.; Ван, М.; Чжан, X.; Чжоу, З. (2014). «Об отсутствии стернальных элементов у Anchiornis (Paraves) и Sapeornis (Aves) и сложной ранней эволюции грудины птиц» . Труды Национальной академии наук . 111 (38): 13900–13905. Бибкод : 2014PNAS..11113900Z . дои : 10.1073/pnas.1411070111 . ПМЦ 4183337 . ПМИД 25201982 .
- ^ Фот, К. (2014). «Комментарий по поводу отсутствия окостеневших стернальных элементов у базальных паравианских динозавров» . Труды Национальной академии наук . 111 (50): Е5334. Бибкод : 2014PNAS..111E5334F . дои : 10.1073/pnas.1419023111 . ПМЦ 4273336 . ПМИД 25480553 .
- ^ Чжэн, X.; Чжоу, З.; Ван, X.; Чжан, Ф.; Чжан, X.; Ван, Ю.; Вэй, Г.; Ван, С.; Сюй, X. (2013). «Задние крылья базальных птиц и эволюция перьев ног». Наука . 339 (6125): 1309–1312. Бибкод : 2013Sci...339.1309Z . CiteSeerX 10.1.1.1031.5732 . дои : 10.1126/science.1228753 . ПМИД 23493711 . S2CID 206544531 .
- ^ Годфруа, П.; Демуйнк, Х.; Дайк, Г.; Ху, Д.; Эскуилье, Ф.; Клейс, П. (2012). «Уменьшенное оперение и летные способности нового юрского паравианского теропода из Китая» . Природные коммуникации . 4 : 1394. Бибкод : 2013NatCo...4.1394G . дои : 10.1038/ncomms2389 . ПМИД 23340434 .
- ^ Фрейзер, Г. (2014). « Причудливые структуры» указывают на то, что дромеозавры являются паразитами, и на новую теорию происхождения птичьего полета» (Автоматическая загрузка в формате PDF) . Журнал палеонтологических наук . 6 :1–27.
- ^ Сюй, Х.; Ли, Ф.; Ван, Ю.; Салливан, К.; Чжан, Ф.; Чжан, X.; Салливан, К.; Ван, X.; Чжэн, X. (2018). «Исключительные окаменелости динозавров указывают на раннее происхождение птичьего пищеварения» . Научные отчеты . 8 (1): 14217. Бибкод : 2018NatSR...814217Z . дои : 10.1038/s41598-018-32202-x . ISSN 2045-2322 . ПМК 6155034 . ПМИД 30242170 .









