Энигмозавр
| Энигмозавр | |
|---|---|
| |
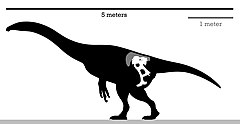
| Скелетная схема голотипа | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Клэйд : | Динозаврия |
| Клэйд : | Саурисхия |
| Клэйд : | Теропода |
| Клэйд : | † Теризинозаврия |
| Суперсемейство: | † Теризинозавроидеа |
| Род: | † Энигмозавр Барсболд, 1983 год. |
| Типовой вид | |
| † Энигмозавр монгольский Барсболд, 1983 год.
| |
Энигмозавр (что означает «загадочная ящерица» или «загадочная ящерица») — род теризинозавроидов , обитавших в Азии в позднемеловой период . Это было наземное двуногое травоядное животное среднего размера , которое представляет третий таксон теризинозавров из формации Баян Шире , хотя известно из нижней части. Род монотипичен , включая только типовой вид E. mongoliensis , известный по хорошо сохранившемуся тазу и другим предполагаемым остаткам тела.
Открытие и присвоение имени
[ редактировать ]
Голотип Монголия , IGM 100/84 , был обнаружен в местонахождении Хара Хутул в формации Баян Ширех (иногда называемой Байнширской свитой или Байнширской свитой), юго-восточная , датируемый позднемеловым периодом, и впервые сообщалось в 1979 году при сравнении тазовых органов. с другими динозаврами-тероподами. В то время о теризинозаврах было мало что известно. [ 1 ] В 1980 году он был упомянут еще раз, на этот раз в новом инфраотряде, созданном монгольскими палеонтологами Ринченом Барсболдом и Алтангерэлом Перле : Сегнозаврия . Прозванный «Динозавром из Хара Хутула», он был вскоре описан и включен в Сегнозаврию. [ 2 ]

Три года спустя типовой вид был Enigmosaurus mongoliensis назван и описан в 1983 году Барсболдом. Сохранившиеся элементы представляют собой частичный скелет без черепа, включающий хорошо сохранившийся тазовый пояс с другими посткраниями. Родовое название , Энигмозавр , было заявлено как производное от греческого αίνιγμα ( aínigma , что означает загадка) и σαῦρος ( sauros , что означает ящерица), что связано с аберрационной и необычной формой его таза. Видовое название mongoliensis . относится к стране его открытия — Монголии [ 3 ] Некоторые находки окаменелостей теризинозавров в формации Ирен-Дабасу сначала считались относящимися к энигмозаврам . [ 4 ] Хотя он широко известен по тазу, несколько останков, не упомянутых в первоначальном описании голотипа, были помечены тем же номером экземпляра (IGM 100/84). Эти элементы находятся в плохом состоянии по сравнению с тазом, не измерялись и не иллюстрировались: проксимальный конец бедренной кости ; большой стержень бедренной кости, возможно, стержень большеберцовой кости; несколько ребер ; дистальный конец плечевой кости ; ориентировочная лучевая кость и проксимальный конец локтевой кости . Однако бедренная кость не была обнаружена в связи с особью-голотипом, поэтому ее следует отнести к Therizinosauria incertae sedis ; остальные останки могли принадлежать голотипу. Очень большая левая бедренная кость длиной 105 см (1050 мм) также была отмечена тем же номером экземпляра, однако она не была связана с голотипом из-за ее больших размеров (больше самого таза). Тем не менее, похоже, он приближается к размеру более близкого сегнозавра . [ 5 ]
Возможная синонимия с эрликозавром.
[ редактировать ]Некоторые палеонтологи предположили, что энигмозавр, , был тем же животным, что и эрликозавр , поскольку оба были найдены в одной и той же геологической формации . Энигмозавр известен по останкам таза, тогда как таз эрликозавра вероятно неизвестен, и отличительные черты между ними отсутствовали; , это сделает Энигмозавра младшим синонимом Эрликозавра если это будет доказано . [ 6 ] [ 7 ] Однако таз энигмозавра не похож на таз сегнозавра , как можно было бы ожидать от связи с эрликозавром , и существует огромная разница в размерах. [ 2 ] [ 3 ] [ 8 ] Кроме того, оба рода известны из разных слоев (нижних и верхних), [ 3 ] [ 9 ] [ 10 ] и большинство авторов считают их отдельными животными. [ 11 ] [ 12 ]
Описание
[ редактировать ]
Энигмозавр был теризинозавром с относительно крупным телом, предполагаемая длина которого составляла 5 м (16 футов), а вес - от 454 до 907 кг (от 1001 до 2000 фунтов). [ 13 ] Как отметил Барсболд в оригинальном описании энигмозавра , его можно узнать по следующим признакам: лобковая и седалищная кости короткие; удлиненный край в передней пресимфизарной области дистальной части лобковой кости. [ 3 ] Однако в пересмотренном диагнозе Zanno et al. 2010 есть еще более специфические черты , у энигмозавра , которые ранее не были отмечены/проанализированы: выраженные краниальные и каудальные отростки на несколько уплощенном в дорсовентральном направлении лобковом ботинке; лобковые стопы сращены, удлинены медиально, медиальное расширение образует V-образную структуру; медиальное сращение запирательного отростка и лонного тела, образует тетралучевой отросток. [ 5 ]

Голотип таза находится в относительно хорошей сохранности, повреждены крестцовые позвонки, частичная подвздошная кость , правая и левая лобковые кости и левая седалищная кость . Таз в целом несколько большой, опистолобковый . Подвздошная кость широко расположена и повернута наружу; в постедорсальной области сохраняется крупный кубический отросток. удлиненные лобковые ножки В подвздошной кости имеются ; однако седалищные ножки более редуцированы. Дистальный конец лобка удлинен, загнут и коренаст. Седалищная кость немного короче лобковой кости и параллельна ей, с узким стержнем. Запирательный отросток на переднем крае седалищной кости горизонтально вытянутый и низкий. заметны открытые края На большой вертельной ямке . По-видимому, в крестце сохранилось шесть позвонков, с удлиненными поперечными отростками . [ 3 ] Например, его таз очень своеобразен по сравнению с другими родственниками теризинозавров: на подвздошной кости имеются участки резорбции и ремоделирования костей . Эти специфические черты могут указывать на пожилой возраст особи, если это правда, то слияние запирательного отростка и лобкового тела можно было бы отбросить как подлинную аутапоморфию для этого вида. Занно отметил, что для решения этой загадки необходимы дополнительные исследования. [ 5 ]
Классификация
[ редактировать ]Энигмозавр был отнесен авторами описания к отдельной группе Enigmosauridae (теперь устаревшей) из-за ее аномального таза, но позже считался членом группы Segnosauridae , которая сегодня называется Therizinosauridae . [ 3 ] Линдси Занно в 2010 году восстановила более базальную позицию среди Therizinosauroidea . [ 5 ]

Ниже приведены результаты недавно проведенного филогенетического анализа Therizinosauria Hartman et al. 2019 , в котором энигмозавр обнаружен как производное теризинозавроид. [ 12 ]
| Теризинозаврия |
| ||||||||||||||||||||||||||||||||||||
Палеоэкология
[ редактировать ]
Останки энигмозавра были обнаружены в отложениях, отложившихся в позднемеловой период на формации Баян Ширех , местонахождение Хара Хутул, примерно от 95,9 ± 6,0 млн до 89,6 ± 4,0 млн лет назад, сеноман - сантонский возраст. [ 3 ] [ 14 ] Будучи теризинозавром, он, вероятно, был медленным травоядным и/или всеядным , как предполагало большинство авторов того времени. [ 15 ] [ 16 ] [ 17 ] Биоразнообразие останках формации характеризовалось теризинозаврами, как видно на энигмозавра , эрликозавра и сегнозавра . [ 18 ] В местоположении голотипа, Хара Хутул, также был обнаружен современный сегнозавр. [ 2 ] и безымянный велоцирапторин . [ 19 ] [ 20 ] Большая часть оставшейся палеофауны этой формации известна из верхних слоев, тогда как энигмозавр известен из нижних слоев. [ 10 ]
В этом месте была обнаружена разнообразная палеофлора, например, Cornaceae, а также Bothrocarium gobience и Nyssoidea mongolica как прекрасные представители. [ 21 ] Многочисленные находки ископаемых фруктов на этом участке отражают большое присутствие в покрытосеменных растений формации . Раскопанные плоды имеют некоторое сходство с Abelmoschus esculentus , однако окончательное таксономическое сходство совершенно неясно. [ 22 ]
См. также
[ редактировать ]- Хронология исследований теризинозавров
- Ринчен Барсболд
- Глоссарий анатомии динозавров
- Анатомические условия расположения
Ссылки
[ редактировать ]- ^ Барсболд, Р. (1979). «Опистолобковый таз хищных динозавров». Природа . 279 (5716): 792–793. Бибкод : 1979Natur.279..792B . дои : 10.1038/279792a0 . S2CID 4348297 .
- ^ Jump up to: а б с Барсболд, Р.; Перл, А. (1980). «Сегнозаврия, новый подотряд хищных динозавров» (PDF) . Acta Palaeontologica Polonica . 25 (2): 190–192.
- ^ Jump up to: а б с д и ж г Barsbold, R. (1983). "Хищные динозавры мела Монголии" [Carnivorous dinosaurs from the Cretaceous of Mongolia] (PDF) . Transactions of the Joint Soviet-Mongolian Paleontological Expedition (in Russian). 19 : 89. Translated paper
- ^ Карри, Пи Джей; Эберт, Д.А. (1993). «Палеонтология, седиментология и палеоэкология формации Ирен-Дабасу (верхний мел), Внутренняя Монголия, Китайская Народная Республика». Меловые исследования . 14 (2): 138. doi : 10.1006/cres.1993.1011 .
- ^ Jump up to: а б с д Занно, Л.Е. (2010). «Таксономическая и филогенетическая переоценка теризинозаврии (Dinosauria: Maniraptora)» . Журнал систематической палеонтологии . 8 (4): 503–543. дои : 10.1080/14772019.2010.488045 . S2CID 53405097 .
- ^ Кларк, Дж. М.; Марьянская, Т.; Барсболд, Р. (2004). «Теризинозавроидеа» . Динозаврия . Издательство Калифорнийского университета. п. 159. ИСБН 9780520242098 .
- ^ Пол, GS (2010). Принстонский полевой справочник по динозаврам . Издательство Принстонского университета. п. 159. ИСБН 978-0-6911-3720-9 .
- ^ Додсон, П. (1993). «Эрликозавр». Эпоха динозавров . Публикации Интернешнл, ООО. п. 142. ИСБН 0-7853-0443-6 .
- ^ Кларк, Дж. М.; Перл, А.; Норелл, М. (1994). «Череп Erlicosaurus andrewsi, позднемелового сегнозавра (Theropoda, Therizinosauridae) из Монголии». Новитаты Американского музея (3115): 1–39. hdl : 2246/3712 .
- ^ Jump up to: а б Цогтбаатар, К.; Вейшампель, Д.Б.; Эванс, округ Колумбия; Ватабе, М. (2019). «Новый гадрозавроид (Dinosauria: Ornithopoda) из позднемеловой байнширской формации пустыни Гоби (Монголия)» . ПЛОС ОДИН . 14 (4): e0208480. Бибкод : 2019PLoSO..1408480T . дои : 10.1371/journal.pone.0208480 . ПМК 6469754 . ПМИД 30995236 .
- ^ Стефан, Л.; Эмили-младший; Перл, А.; Линдси, ЭЗ; Лоуренс, Миссури (2012). «Эндокраниальная анатомия теризинозаврии и ее влияние на сенсорные и когнитивные функции» . ПЛОС ОДИН . 7 (12): e52289. Бибкод : 2012PLoSO...752289L . дои : 10.1371/journal.pone.0052289 . ПМЦ 3526574 . ПМИД 23284972 .
- ^ Jump up to: а б Хартман, С.; Мортимер, М.; Валь, WR; Ломакс, ДР; Липпинкотт, Дж.; Лавлейс, DM (2019). «Новый паравианский динозавр из поздней юры Северной Америки подтверждает позднее приобретение птичьего полета» . ПерДж . 7 : е7247. дои : 10.7717/peerj.7247 . ПМК 6626525 . ПМИД 31333906 .
- ^ Хольц, ТР; Рей, Л.В. (2007). Динозавры: самая полная и современная энциклопедия для любителей динозавров всех возрастов . Случайный дом. Список родов Holtz 2012 Информация о весе
- ^ Курумада, Ю.; Аоки, С.; Аоки, К.; Като, Д.; Санеёси, М.; Цогтбаатар, К.; Уиндли, БФ; Исигаки, С. (2020). «Кальцитовый U-Pb возраст меловой формации Баин Шир, содержащей позвоночных, в восточной пустыне Гоби в Монголии: полезность калише для определения возраста». Терра Нова 32 (4): 246–252. Бибкод : 2020Ноябрь..32..246K дои : 10.1111/тер.12456 .
- ^ Пол, GS (1984). «Сегнозавры динозавры: реликты прозавропод-птицетазового перехода?». Журнал палеонтологии позвоночных . 4 (4): 507–515. дои : 10.1080/02724634.1984.10012026 . ISSN 0272-4634 . JSTOR 4523011 .
- ^ Занно, Ле; Цогтбаатар, К.; Чинзориг, Т.; Гейтс, Т.А. (2016). «Специализации анатомии нижней челюсти и зубочелюстного аппарата Segnosaurus galbinensis (Theropoda: Therizinosauria) » ПерДж . 4 : е1885. дои : 10.7717/peerj.1885 . ПМЦ 4824891 . ПМИД 27069815 .
- ^ Макалузо, Л.; Чопп, Э.; Маннион, П. (2018). «Эволюционные изменения в ориентации лобка у динозавров сильнее коррелируют с системой вентиляции, чем с травоядностью». Палеонтология . 61 (5): 703–719. дои : 10.1111/пала.12362 . S2CID 133643430 .
- ^ Ли, Ю.М.; Ли, HJ; Кобаяши, Ю.; Карабахал, АП; Барсболд, Р.; Фиорилло, Арканзас; Цогтбаатар, К. (2019). «Необычное поведение при передвижении, сохранившееся на крокодилообразных путях из верхнемеловой баянширской формации Монголии, и его палеобиологические последствия». Палеогеография, Палеоклиматология, Палеоэкология . 533 (109353): 2. Цифровой код : 2019PPP...533j9239L . дои : 10.1016/j.palaeo.2019.109239 . S2CID 197584839 .
- ^ Кубота, К.; Барсболд, Р. (2007). «Новый дромеозаврид (Dinosauria Theropoda) из верхнемеловой баянширской свиты Монголии». Журнал палеонтологии позвоночных . 27 (дополнение к 3): 102А. дои : 10.1080/02724634.2007.10010458 .
- ^ Тернер, А.Х.; Маковицкий, П.Дж.; Норелл, Массачусетс (2012). «Обзор систематики дромеозаврид и филогении паравианов» . Бюллетень Американского музея естественной истории . 371 (371): 1–206. дои : 10.1206/748.1 . hdl : 2246/6352 . S2CID 83572446 .
- ^ Кханд, Ю.; Бадамгарав, Д.; Ариунчимег, Ю.; Барсболд, Р. (2000). «Меловая система Монголии и условия ее отложения». Меловая среда Азии . Развитие палеонтологии и стратиграфии. Том. 17. С. 49–79. дои : 10.1016/s0920-5446(00)80024-2 . ISBN 9780444502766 .
- ^ Ксепка, Д.Т.; Норелл, Массачусетс (2006). «Эркету эллисони, длинношеий зауропод из Бор Гуве (Дорноговский аймак, Монголия)» (PDF) . Американский музей Novitates (3508): 1–16. doi : 10.1206/0003-0082(2006)3508[1:EEALSF]2.0.CO;2 . S2CID 86032547 .









