Эрликозавр
| Эрликозавр Временной диапазон: поздний мел ,
~ | |
|---|---|
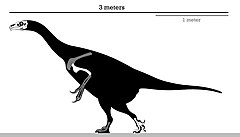
| |
| Скелетная схема голотипа | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Клэйд : | Динозаврия |
| Клэйд : | Саурисхия |
| Клэйд : | Теропода |
| Суперсемейство: | † Теризинозавроидеа |
| Семья: | † Теризинозавриды |
| Род: | † Эрликозавр Барсболд и Перл , 1980 г. |
| Типовой вид | |
| † Эрликозавр Эндрюси Барсболд и Перл, 1980 г.
| |
| Синонимы | |
| |
Эрликозавр (что означает « Эрлика ящерица ») — род теризинозавридов , обитавших в Азии в позднемеловой период. Окаменелости, череп и некоторые посткраниальные фрагменты, были найдены в формации Баян Шире в Монголии в 1972 году и датируются примерно 96 и 89 миллионами лет назад. Эти останки были позже описаны Альтангерелем Перлом и Ринченом Барсболдом в 1980 году, назвав новый род и вид Erlikosaurus andrewsi . Он представляет собой второй таксон теризинозавров из этой формации (наряду с энигмозавром и сегнозавром ) с наиболее полным черепом среди представителей этого своеобразного семейства динозавров.
В отличие от большинства теризинозавридов, эрликозавр был небольшим представителем, достигавшим почти 3,4 м (11 футов) в длину и от 150 до 250 кг (от 330 до 550 фунтов) в массе. У него был хорошо развитый клюв на кончике морды и зубчатые челюсти, которые использовались для травоядного питания. Ступни заканчивались четырьмя пальцами , первый из которых сочленялся с лодыжкой — в отличие от рудиментарного первого пальца у большинства теропод. Как и другие теризинозавриды, эрликозавр имел большую кишку для переработки пищи, сильные руки, оканчивающиеся удлиненными когтями , и направленный назад таз .
Эрликозавр классифицируется как теризинозавр в отряде Therizinosauridae . Теризинозавры были загадочными динозаврами с неясными взаимоотношениями в первые годы исследований. Последующие исследования доказали их истинную природу как динозавров-тероподов и систематическое положение среди манирапторов . Клюв и челюсти эрликозавра указывают на метод кормления с отрывом листьев , характеризующийся активным использованием клюва при помощи шеи. Некоторые различия с симпатрическим сегнозавром показывают, что эти родственные роды были разделены по нишам .
Открытие и присвоение имени
[ редактировать ]
Образец голотипа , MPC-D 100/111 , был найден в слоях из местонахождения Байшин-Цав формации Баян-Шире и состоит из исключительно хорошо сохранившегося черепа , практически полной правой стопы , в которой отсутствуют только проксимальные концы плюсневых костей II, III и IV и почти полная левая плечевая кость . Другие останки включают фрагментарные шейные позвонки , однако их количество не указано и они не иллюстрированы. Эти находки были сделаны во время советско-монгольской экспедиции в провинции Омногови в 1972 году. [ 1 ] [ 2 ] Восемь лет спустя род и типовой вид , Erlikosaurus andrewsi , был назван и описан (хотя и очень кратко) палеонтологами Ринченом Барсболдом и Алтангерелем Перле в 1980 году , однако Барсболд не был указан как названитель этого конкретного вида. Родовое имя , Erlikosaurus , было взято от названия короля демонов Эрлика , из тюрко-монгольской мифологии и греческого σαῦρος ( sauros , что означает ящерица). Видовое название andrewsi . дано в честь американского палеонтолога Роя Чепмена Эндрюса , который был руководителем американских азиатских экспедиций с 1922 по 1930 год [ 1 ] Судя по всему, в первоначальном описании частью голотипа была указана левая стопа. [ 1 ] однако это заявление больше не упоминалось. [ 2 ] [ 3 ] [ 4 ] [ 5 ]

Как ни странно, в 1981 году Перл снова назвал и описал этот вид, как если бы он был новым, но на этот раз более подробно и написав родовое название как латинизированное «Эрликозавр». [ 2 ] Сегодня большинство авторов широко признают, что первоначальное название Erlikosaurus действительно. На момент открытия это был единственный известный теризинозавр (тогда его называли сегнозаврами). [ 6 ] ), у которого был обнаружен целый череп, это помогло пролить свет на загадочную и малоизученную группу динозавров. Он до сих пор представляет собой наиболее известный череп теризинозавра. [ 4 ] [ 5 ]
В 2010 году Грегори С. Пол оспорил обоснованность этого таксона, утверждая, что эрликозавр может быть синонимом энигмозавра (названного в 1983 году) . [ 7 ] ), поскольку останки последнего были найдены в той же геологической формации и известны только по останкам таза, тогда как таз у эрликозавра неизвестен; сделало бы Энигмозавра младшим синонимом Эрликозавра это . [ 8 ] Однако, поскольку голотип бедра энигмозавра не очень напоминал бедро экземпляра сегнозавра , как можно было бы ожидать от сегнозавроподобных останков эрликозавра , и существует значительная разница в размерах, палеонтолог Ринчен Барсболд оспорил предполагаемую синонимию. [ 9 ] Кроме того, останки эрликозавра и энигмозавра известны с верхней и нижней границы соответственно. [ 3 ] [ 7 ] [ 10 ] Следовательно, энигмозавр и эрликозавр обычно считаются отдельными родами. [ 11 ]
Описание
[ редактировать ]
было проблематично Поскольку род известен только по очень фрагментарному материалу, определить размер эрликозавра , тем более что большая часть позвоночного столба голотипа отсутствует. Длина черепа экземпляра-голотипа составляет примерно 25 см (250 мм), что указывает на очень маленькую особь. В целом, эрликозавр был теризинозавром небольшого размера, его рост, по оценкам, составлял около 3,4 м (11 футов), а телосложение было более легким, чем у тяжеловесного сегнозавра . [ 12 ] В 2012 году Стефан Лаутеншлагер и его коллеги использовали уравнения, специфичные для теропод, для оценки массы тела эрликозавра и других теризинозавров. Однако, поскольку бедренная кость неизвестна, они использовали двумерный регрессионный анализ лог-преобразованных данных для Erlikosaurus . В результате длина бедра составила 44,33 см (443,3 мм), а вес - 173,7 кг (383 фунта). Учитывая неопределенность этих оценок, они установили общий диапазон массы от 150 до 250 кг (от 330 до 550 фунтов). [ 5 ] Альтернативные оценки предполагают, что максимальная длина составляет 6 м (20 футов). [ 9 ] и более консервативная длина 4,5 метра и вес 500 кг (1100 фунтов). [ 13 ] Хотя у эрликозавра в основном отсутствуют останки тела, как у теризинозаврид он имел сильное строение рук с большими когтями, широкое и громоздкое туловище и опистолобковый (направленный назад) таз. [ 14 ] , известно, что теризинозавры были пернатыми Судя по сохранившимся отпечаткам перьев у экземпляров Beipiaosaurus и Jianchangosaurus животными , поэтому вполне вероятно , что эрликозавр также был пернатым. [ 15 ] [ 16 ]
Череп
[ редактировать ]

Морда умеренно удлиненная, предчелюстная кость имеет удлиненные носовые отростки . Тонкая вертикальная костная пластинка рострально соединена с медиальным краем предчелюстной кости, что указывает на то, что когда животное было живо, у него хрящевая межносовая перегородка присутствовала . В дополнение к этому, в предчелюстной кости есть латеральные и медиальные отверстия , которые соединены сложной системой сосудистых каналов , которая пронизывает структуру предчелюстной кости и, вероятно, связана с сенсорными ветвями нейрососудистой сети и глазным нервом, поддерживающими рамфотеку (клюв). Верхняя челюсть треугольной формы, сохраняет 24 , зубы гомодонтные с . грубыми зазубринами альвеолы Зубная кость имеет клиновидно-вытянутую форму, сохраняет 31 альвеолу. При взгляде сверху он имеет U-образную форму, уплощен сзади с лежащим поперек расширением. На латеральной и вентральной поверхностях симфизарной области имеется ряд отверстий диаметром от 2 до 5 мм (от 0,20 до 0,50 см). Изолированные отверстия соединены внутри сложным сосудисто-нервным каналом. После восстановления длина черепа составляет 26 см (260 мм), а длина нижней челюсти — около 24 см (240 мм). [ 17 ]
Хорошо сохранившаяся черепная коробка практически полная, отсутствует только сфенетмоид-мезетмоидный комплекс, тогда как латеросфеноиды и орбитосфеноиды сохранились не полностью при медиальной проекции. Кости вокруг черепной коробки сильно окостеневшие , но швы между отдельными элементами поверхностно не видны, за исключением нескольких участков. [ 3 ] Однако эти внутренние швы можно проследить на компьютерной томографии и, следовательно, элементы черепной коробки можно отличить друг от друга. [ 17 ] Восстановленный мозг экземпляра несколько удлинен. Обонятельный аппарат и полушария головного мозга очень известны: обонятельный тракт намного больше, чем настоящий мозг. Полушария головного мозга большие и широкие. На поверхности мозга можно обнаружить сложные сосудистые бороздки, которые обычно встречаются у птиц и млекопитающих , а также у других динозавров. Наконец, мозжечок не столь печально известен, как предыдущие элементы, он удлиненный и коренастый. [ 5 ]

Кератиновые клювы , или рамфотеки, хорошо известны среди различных групп динозавров . [ 17 ] У орнитомимозавров есть веские доказательства этого. [ 18 ] [ 19 ] Однако это не является показателем того, что можно предположить отсутствие данной анатомической особенности у других групп. Некоторые характеристики указывают на рамфотеку, такие как беззубая предчелюстная кость с тонким, сужающимся нижним краем, последовательная потеря верхних и зубных зубов, вогнутость нижней челюсти на нижней стороне, смещение нижней поверхности в зубной кости и ростральная проекция нижнечелюстного симфиза. [ 17 ]
У эрликозавра о наличии ороговевшего клюва на верхней и предчелюстной кости можно судить по наличию многочисленных сосудисто-нервных отверстий на ростральной и латеральной поверхностях черепа, кроме того, он несет все указанные выше признаки, однако неясно, расширение клюва. [ 17 ] Сохранившаяся рамфотека у экземпляров Gallimimus и Ornithomimus свидетельствует о том, что кератиновый чехол покрывал предчелюстную кость и перекрывал ее с нижней стороны на несколько миллиметров. [ 18 ] У некоторых современных птиц рамфотека обычно ограничивается предчелюстной и верхней челюстями, хотя в некоторых случаях у некоторых птиц она частично закрывает носовой отросток. [ 20 ] По-видимому, у эрликозавра рамфотека прикрывала носовой отросток предчелюстной кости. [ 17 ]
Посткраниальный скелет
[ редактировать ]
Останки тела эрликозавра очень редки по сравнению с элементами черепа и состоят из плечевой кости , правой стопы и некоторых шейных позвонков . Отдельные шейки матки не были изображены и подсчитаны, а лишь кратко описаны. Шейные позвонки платикоелусные (слегка вогнутые на обоих концах) с низкими невральными дугами . [ 1 ] Будучи относительно крепкими, они имеют толстые презигапофизы и крупные парапофизы. Кроме того, шейные позвонки имеют некоторое сходство с шейками сегнозавра , однако они намного меньше. [ 2 ]
Сохранившаяся правая стопа практически цела, отсутствует только проксимальный конец плюсневых костей II, III и IV . Она укорочена в длину, имеет крепкие плюсневые кости с расширенными суставными конечностями и образует некомпактную плюсну. Плюсневая кость I самая короткая по сравнению с ней, ее длина составляет 7 см (70 мм) и она расширяет расширенную латерально проксимальную суставную поверхность плюсны. Все остальные плюсневые кости примерно одинакового размера, длина II плюсневой кости составляет 11 см (110 мм). пальцы Педальные имеют весьма своеобразное строение; первая цифра уменьшена в длине, при этом все остальные цифры почти равны по длине, однако четвертая цифра очень тонкая по сравнению с остальными. Фаланги . трех первых пальцев укорочены, крепкие, сопоставимого строения Вторая и третья фаланги четвертого пальца дискоидальные, коренастые. Наконец, когти загнуты назад, исключительно велики и сильно уплощены с боков. [ 1 ] [ 2 ] Грегори С. Пол предположил, что длинные тонкие когти ног использовались в качестве механизма самозащиты. [ 13 ]
Левая плечевая кость — единственный сохранившийся остаток грудной области. Плечевая кость имеет удлиненные эпифизы и относительно крупный дельтовидный отросток. [ 1 ] Он прочный, его расчетная длина составляет 30 см (300 мм). Имеет уменьшенный вал. Проксимальный конец плечевых костей очень широкий. Головка плечевой кости имеет суставную поверхность выпуклую и широкую, посередине она редуцирована к краям. Имеется выступающий дельтопекторальный гребень, вершина которого расположена на 1/3 длины плечевой кости от проксимального конца. сочленения Мыщелки лучевой дифференцированы и разделены и локтевой костей укороченной бороздкообразной ямкой и в целом сильно уменьшены в размерах. Ямка для локтевого отростка умеренно глубокая и широкая. Заметна внутренняя шероховатость головки. [ 2 ] как и у неродственных Dromaeosauridae . [ 21 ]
Классификация
[ редактировать ]
Эрликозавр был отнесен Перлем к семейству сегнозавров . [ 1 ] группа, сегодня известная как Therizinosauridae , что подтверждено более поздними кладистическими анализами. [ 4 ] Теризинозавры были странной группой теропод , которые питались растениями вместо мяса и имели обращенный назад лобок , как и птицетазовые животные . Как и у птицетазовых, их челюсти имели на конце широкий округлый костный клюв, который можно было использовать для обрезки растений. [ 9 ] [ 4 ]
Отношения теризинозавров были довольно сложными, когда были обнаружены первые представители. Например, первый известный таксон теризинозавров, Therizinosaurus , интерпретировался как черепахоподобное животное, которое использовало удлиненные когти для питания морскими водорослями. [ 22 ] Однако в 1970 году Рождественский выдвинул идею о том, что теризинозавры (тогда известные как сегнозавры) вместо того, чтобы быть существами, не являющимися динозаврами, на самом деле были тероподами . [ 23 ] Позже, в 1980 году, сегнозавров считали медленными полуводными животными, при этом Грегори С. Пол утверждал, что эти спорные животные не имели характеристик теропод и были прозауроподами с птицетазовыми адаптациями, а также имели общие эволюционные отношения. [ 24 ] Однако с описанием большего количества родов, таких как Alxasaurus , [ 25 ] Наньшиунгозавр , [ 26 ] и переописание черепа эрликозавра стали подтверждаться новыми доказательствами теропод. [ 3 ] С открытием и описанием пернатого бейпиозавра теризинозавры были полностью признаны тероподами, и их начали реконструировать в точной двуногой позе. [ 15 ]
Следовательно, теризинозавры теперь классифицируются как тероподы в составе целурозаврии . Линдси Занно была одним из первых авторов, подробно изучивших взаимоотношения и родство теризинозавров. Ее работа оказалась полезной во многих филогенетических анализах. [ 4 ] Кладограмма ниже является результатом филогенетического анализа, проведенного Hartman et al. 2019 с использованием данных, предоставленных Занно в 2010 году. Эрликозавр занимал весьма производное положение в кладе с двумя видами Nothronychus : [ 11 ]
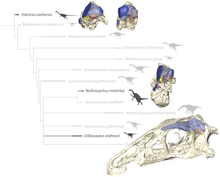
| Теризинозавриды |
| |||||||||||||||||||||||||||||||||||||||||||||||||||
Палеобиология
[ редактировать ]Чувства
[ редактировать ]
Эрликозавр малоизвестен по посткраниальному материалу, но череп-голотип стал предметом изучения в сканах компьютерной томографии (КТ) , которые были опубликованы еще в 2012 году палеонтологом Стефаном Лаутеншлагером и доктором Эмили Рэйфилд из Школы наук о Земле Бристольского университета, профессором Линдси Занно. из Музея естественной истории Северной Каролины и Университета штата Северная Каролина , а также Лоуренс Уитмер, профессор палеонтологии Чанга в Колледже остеопатической медицины Университета Огайо. Анализ полости мозга показал, что эрликозавр и, вполне вероятно, большинство других теризинозавридов имели хорошо развитые чувства обоняния, слуха и равновесия — черты, лучше присущие плотоядным тероподам . Увеличенный передний мозг эрликозавра , возможно, также был полезен в сложном социальном поведении и уклонении от хищников . Эти чувства были также хорошо развиты у более ранних целурозавров и других теропод, что указывает на то, что теризинозавры, возможно, унаследовали многие из этих черт от своих плотоядных предков и использовали их для различных и специализированных диетических целей. [ 5 ]
В 2019 году Грэм М. Хьюз и Джон А. Финарелли проанализировали соотношение обонятельных луковиц у современных птиц и сохранили черепа нескольких вымерших видов динозавров, чтобы предсказать, сколько генов могло быть задействовано в обонятельной силе этих вымерших видов. Их анализ показал, что у эрликозавра было около 477 генов, кодирующих его обонятельные рецепторы, и соотношение обонятельных луковиц, равное 40, что указывает на умеренное обоняние. Оценки эрликозавра были выше, чем у большинства дромеозаврид, несмотря на травоядный образ жизни этого таксона, и могут отражать переход к сложной социальности и/или снижение зрительных способностей. Хьюз и Финарелли отметили, что по мере того, как линии динозавров становились крупнее, размер обонятельной луковицы увеличивался, что может указывать на то, что обоняние является основной сенсорной модальностью у крупных нептичьих динозавров. [ 27 ]
Кормление и сила укуса
[ редактировать ]
В 2013 году реконструкцию черепной мускулатуры эрликозавра Лаутеншлагер выполнил цифровую и обнаружил относительно слабую силу укуса по сравнению с другими тероподами. В целом приводящая мускулатура челюстей, которая в первую очередь выполняет функцию смыкания челюстей, генерирует общую силу 374 и 570 Н, но фактически при накусывании используется лишь небольшая часть, поскольку сила укуса начинает уменьшаться по мере увеличения расстояния. точка укуса – челюстной сустав. Лаутеншлагер обнаружил наименьшую силу на кончике рыла (43–65 Н) и самую высокую в области последнего зуба верхней челюсти (90–134 Н). Такие факторы, как наличие большой кишки для обработки растительности и отсутствие повреждений на Зубы предполагают, что эрликозавр использовал только кончик морды и предчелюстную область, чтобы дотянуться до мягкой листвы или фруктов, а меньшая сила укуса эрликозавра лучше использовалась в механизме кормления с отрывом листьев и обрезкой растений, а не в активном жевании . В этом исследовании Лаутеншлагер также предположил, что Эрликозавр , возможно, был способен перерабатывать в основном тонкие ветки и растительные вещества, основываясь на Стегозавре . Более того, сравнительно узкая ширина морды могла указывать на избирательное питание у этого теризинозаврид. с обрезанием ветвей Наконец, поведение эрликозавра могло быть компенсировано посткраниальной мускулатурой. [ 28 ] В том же году Лаутеншлагер и его команда создали цифровые модели черепа эрликозавра , чтобы проверить функцию рамфотеки ( ороговевшего клюва), обнаружив, что эта структура челюстей действует как структура, смягчающая стресс. Они пришли к выводу, что кератиновые клювы полезны для повышения устойчивости черепа, делая его менее восприимчивым к изгибу и/или деформации во время кормления. [ 29 ]

Хорошо сохранившиеся челюсти также позволили исследованию Бристольского университета определить, как стиль кормления и диетические предпочтения связаны с тем, насколько широко они могут открывать рот. В исследовании, проведенном Лаутеншлагером и его коллегами в 2015 году , было обнаружено, что эрликозавр мог открывать рот максимум на угол 43 градуса. Также в исследование для сравнения были включены хищные тероподы Аллозавр и Тираннозавр . В результате сравнений было показано, что хищные динозавры имели более широкие челюстные щели, чем травоядные, так же, как современные плотоядные животные сегодня. [ 30 ]
В 2016 году с использованием анализа конечных элементов (FEA) и анализа динамики многих тел (MDA) были протестированы силы укуса эрликозавра , платеозавра и стегозавра , чтобы оценить пищевые привычки. Результирующая сила укуса эрликозавра составляла от 50 до 121 Н , при этом череп характеризовался высокой восприимчивостью к стрессу и деформации, что указывает на пищевое поведение, специализирующееся на активном использовании клюва. Результаты еще раз подтверждают, что эрликозавр полагался на посткраниальную мускулатуру, чтобы компенсировать низкую силу укуса и снять нагрузку на структуру черепа. [ 31 ]
Палеосреда
[ редактировать ]
Голотип эрликозавра обнаружен в местонахождении Байшин Цав на верхней границе Баянширехской свиты , в карьере, сложенном серыми песками с конгломератами , гравием и серыми аргиллитами . Считается, что Байши Цав образовался в результате извилистых рек . [ 32 ] [ 3 ] [ 33 ] Исследование магнитостратиграфии формации , по-видимому, подтверждает, что весь Баян Шире лежит в пределах меловой длинной нормали , просуществовавшей лишь до конца сантонского этапа. Более того, кальцита U-Pb-измерения оценивают возраст формации Баян-Шире от 95,9 ± 6,0 миллиона до 89,6 ± 4,0 миллиона лет назад, от сеномана до сантона. [ 33 ] [ 34 ]
Речные , озерные и калишеобразующие отложения указывают на менее засушливый климат с наличием влажной среды, состоящей из крупных меандров и озер . Крупномасштабная перекрестная стратификация во многих слоях песчаника в местонахождениях Байн Шире и Бурхант, по-видимому, указывает на наличие крупных извилистых рек, и эти крупные водоемы, возможно, осушали восточную часть пустыни Гоби . [ 35 ] [ 33 ] Многочисленные окаменелости фруктов были обнаружены в местонахождениях Бор-Гуве и Хара-Хутул. [ 36 ] В формации известно огромное разнообразие фауны , включая роды динозавров и нединозавров. К тероподам относятся большой Ахиллобатор , [ 37 ] и дейнохейрид Garudimimus . [ 38 ] Другие растительноядные динозавры представлены анкилозаврами Таларурусом и Цагантегией , [ 39 ] мелкие маргиноцефалы Amtocephale и Graciliceratops , [ 40 ] [ 41 ] гадрозавроид Гобихадрос , [ 10 ] и зауропод Эркету . [ 36 ] Другая фауна включает полуводных рептилий, таких как крокодиломорфы и черепахи наньсюнчелид . [ 42 ] [ 43 ]
Сосуществование с сегнозавром
[ редактировать ]
Эрликозавр жил рядом с более крупным видом теризинозавридов в формации Баян Шире, Сегнозавром . В 2016 году Занно и его коллеги повторно исследовали нижнюю челюсть и зубной ряд сегнозавра , прямое сравнение с таковыми у эрликозавра проведя при этом довольно сложные особенности . Они выявили в зубных зубах сегнозавра , которые представлены наличием многочисленных килей (режущих кромок) и складчатых килей с зубчатыми передними краями, а также увеличением зубчиков (зазубринами). Эти черты вместе создают шероховатую, измельчающую поверхность у основания коронок зубов, которая была уникальной для сегнозавра , и позволяют предположить, что он потреблял уникальные пищевые ресурсы или использовал узкоспециализированные стратегии кормления с добавлением более высокой степени пероральной обработки пищи, чем симпатрическая . родственный вид, обитавший на той же территории в одно и то же время — Erlikosaurus . Напротив, у последнего очень симметричные зубы с умеренными зубчиками. Соответствующий нечеткий и специализированный зубной ряд Эрликозавр и Сегнозавр указывают на то, что эти два теризинозавриды были разделены дифференциацией ниш в добывании, переработке или ресурсах пищи. Этот вывод подкрепляется большой разницей в расчетных массах тела, которая составляет до 500%. [ 44 ]

В исследовании разделения ниш у теризинозавров в 2017 году с помощью цифрового моделирования Лаутеншлагер обнаружил, что более прямые и удлиненные зубные кости примитивных теризинозавров испытывали самые высокие величины стресса и напряжения во время сценариев внешнего питания. Напротив, эрликозавру и сегнозавру помогали загнутые вниз кончики нижних челюстей и симфизарные (костные соединения) области, а также, вероятно, клювы, смягчающие стресс и напряжение. Результаты также показали разницу в силе укуса между сегнозавром и эрликозавром , что указывает на то, что первый мог питаться более жесткой растительностью, в то время как общая надежность последнего предполагает большую гибкость в способе питания. Лаутеншлагер отметил, что эти два таксона были адаптированы к различным способам добывания пищи, и что разница в размерах и высоте между двумя теризинозавридами еще больше разделила их ниши . Хотя сегнозавр был приспособлен использовать свой специализированный зубной ряд для добычи или переработки пищи, Эрликозавр в основном полагался на мускулатуру своего клюва и шеи при сборе урожая во время добывания пищи . [ 45 ]
Помимо этих черепных различий, в 2019 году Баттон и Занно отмечают, что травоядные динозавры придерживались двух основных способов питания. Одним из них была переработка пищи в кишечнике, для которого характерны изящные черепа и относительно низкая сила укуса, а вторым — пероральная обработка пищи, характеризующаяся особенностями, связанными с обширной обработкой, например, нижней челюстью или зубными рядами. Было обнаружено, что сегнозавр принадлежал к первому типу, тогда как эрликозавр с большей вероятностью попадал во вторую группу, что еще раз подтверждает, что эти два теризинозавриды были разделены четко определенной дифференциацией ниш. [ 46 ]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а б с д и ж г Барсболд, Р.; Перл, А. (1980). «Сегнозаврия, новый подотряд хищных динозавров» (PDF) . Acta Palaeontologica Polonica . 25 (2): 190–192.
- ^ Jump up to: а б с д и ж Перл, А. (1981). «Новый сегнозаврид из верхнего мела Монголии». Труды совместной советско-монгольской палеонтологической экспедиции (на русском языке). 15 :50–59. Переведенная статья
- ^ Jump up to: а б с д и Кларк, Дж. М.; Перл, А.; Норелл, М. (1994). «Череп Erlicosaurus andrewsi, позднемелового сегнозавра (Theropoda, Therizinosauridae) из Монголии». Новитаты Американского музея (3115): 1–39. hdl : 2246/3712 .
- ^ Jump up to: а б с д и Занно, Л.Е. (2010). «Таксономическая и филогенетическая переоценка теризинозаврии (Dinosauria: Maniraptora)» . Журнал систематической палеонтологии . 8 (4): 503–543. дои : 10.1080/14772019.2010.488045 . S2CID 53405097 .
- ^ Jump up to: а б с д и Лаутеншлагер, С.; Рэйфилд, Э.Дж.; Перл, А.; Занно, Ле; Витмер, LM (2012). «Эндокраниальная анатомия теризинозаврии и ее влияние на сенсорные и когнитивные функции» . ПЛОС ОДИН . 7 (12): e52289. Бибкод : 2012PLoSO...752289L . дои : 10.1371/journal.pone.0052289 . ПМЦ 3526574 . ПМИД 23284972 .
- ^ Перл, А. (1979). «Сегнозавриды — новое семейство теропод из позднего мела Монголии». Труды совместной советско-монгольской палеонтологической экспедиции (на русском языке). 8 : 45–55. Переведенная статья
- ^ Jump up to: а б Barsbold, R. (1983). "Хищные динозавры мела Монголии" [Carnivorous dinosaurs from the Cretaceous of Mongolia] (PDF) . Transactions of the Joint Soviet-Mongolian Paleontological Expedition (in Russian). 19 : 107. Translated paper
- ^ Пол, GS (2010). Принстонский полевой справочник по динозаврам . Издательство Принстонского университета. п. 159 . ISBN 978-0-6911-3720-9 .
- ^ Jump up to: а б с Додсон, П. (1993). «Эрликозавр». Эпоха динозавров . Публикации Интернешнл, ООО. п. 142. ИСБН 0-7853-0443-6 .
- ^ Jump up to: а б Цогтбаатар, К.; Вейшампель, Д.Б.; Эванс, округ Колумбия; Ватабе, М. (2019). «Новый гадрозавроид (Dinosauria: Ornithopoda) из позднемеловой байнширской формации пустыни Гоби (Монголия)» . ПЛОС ОДИН . 14 (4): e0208480. Бибкод : 2019PLoSO..1408480T . дои : 10.1371/journal.pone.0208480 . ПМК 6469754 . ПМИД 30995236 .
- ^ Jump up to: а б Хартман, С.; Мортимер, М.; Валь, WR; Ломакс, ДР; Липпинкотт, Дж.; Лавлейс, DM (2019). «Новый паравианский динозавр из поздней юры Северной Америки подтверждает позднее приобретение птичьего полета» . ПерДж . 7 : е7247. дои : 10.7717/peerj.7247 . ПМК 6626525 . ПМИД 31333906 .
- ^ Хольц, ТР; Рей, Л.В. (2007). Динозавры: самая полная и современная энциклопедия для любителей динозавров всех возрастов . Случайный дом. ISBN 9780375824197 . Список родов Holtz 2012 Информация о весе
- ^ Jump up to: а б Пол, GS (2016). Принстонский полевой справочник по динозаврам (2-е изд.). Принстон, Нью-Джерси: Издательство Принстонского университета. стр. 162−168. ISBN 9780691167664 .
- ^ Хедрик, BP; Занно, Ле; Вульф, генеральный директор; Додсон, П. (2015). «Ленивый коготь: остеология и тафономия Nothronychus mckinleyi и N. graffami (Dinosauria: Theropoda) и анатомические соображения для производных теризинозавридов» . ПЛОС ОДИН . 10 (6): e0129449. Бибкод : 2015PLoSO..1029449H . дои : 10.1371/journal.pone.0129449 . ПМЦ 4465624 . ПМИД 26061728 .
- ^ Jump up to: а б Сюй, Х.; Тан, З.; Ван, XA (1999). «Теризинозавроидный динозавр с покровными структурами из Китая». Природа . 339 (6734): 350–354. Бибкод : 1999Natur.399..350X . дои : 10.1038/20670 . S2CID 204993327 .
- ^ Пу, Х.; Кобаяши, Ю.; Лю, Дж.; Сюй, Л.; Ву, Ю.; Чанг, Х.; Чжан, Дж.; Цзя, С. (2013). «Необычный базальный динозавр-теризинозавр с орнитисхическим расположением зубов из северо-восточного Китая» . ПЛОС ОДИН . 8 (5): e63423. Бибкод : 2013PLoSO...863423P . дои : 10.1371/journal.pone.0063423 . ПМЦ 3667168 . ПМИД 23734177 .
- ^ Jump up to: а б с д и ж Лаутеншлагер, С.; Уитмер, Л.М.; Перл, А.; Занно, Ле; Рэйфилд, Э.Дж. (2014). «Краниальная анатомия Erlikosaurus andrewsi (Dinosauria, Therizinosauria): новые идеи, основанные на цифровой реконструкции». Журнал палеонтологии позвоночных . 34 (6): 1263–1291. дои : 10.1080/02724634.2014.874529 .
- ^ Jump up to: а б Норелл, Массачусетс; Маковицкий, П.Дж.; Карри, Пи Джей (2001). «Клюв страусиных динозавров». Природа . 412 (6850): 873–874. Бибкод : 2001Natur.412..873N . дои : 10.1038/35091139 . ПМИД 11528466 . S2CID 4313779 .
- ^ Барретт, премьер-министр (2005). «Питание динозавров-страусов» . Палеонтология . 48 (2): 347–358. дои : 10.1111/j.1475-4983.2005.00448.x .
- ^ Кнутсен, Э.М. (2007). Морфология клюва современных птиц и влияние на морфологию клюва орнитомимид (PDF) . Университет Осло. п. 44.
- ^ Остром, Дж. Х. (1969). «Остеология Deinonychus antirhopus, необычного теропода из нижнего мела Монтаны» (PDF) . Бюллетень Музея естественной истории Пибоди . 30 :1–165. Архивировано из оригинала (PDF) 4 мая 2020 г. Проверено 26 января 2020 г.
- ^ Maleev, E. A. (1954). "Noviy cherepachoobrazhniy yashcher v Mongolii" [New turtle−like reptile in Mongolia]. Priroda (3): 106–108. Translated paper
- ^ Рождественский, А.К. (1970). «На гигантских когтях загадочных мезозойских рептилий». Палеонтологический журнал (1): 131–141.
- ^ Пол, GS (1984). «Сегнозавры динозавры: реликты прозавропод-птицетазового перехода?». Журнал палеонтологии позвоночных . 4 (4): 507–515. дои : 10.1080/02724634.1984.10012026 . ISSN 0272-4634 . JSTOR 4523011 .
- ^ Рассел, Д.А.; Донг, З. (1993). «Сродство нового теропода из пустыни Альха, Внутренняя Монголия, Китайская Народная Республика». Канадский журнал наук о Земле . 30 (10): 2107–2127. Бибкод : 1993CaJES..30.2107R . дои : 10.1139/e93-183 .
- ^ Донг, З. (1979). Динозавры мелового периода Южного Китая (PDF) (на китайском языке). Красные пласты мезозоя и кайнозоя Южного Китая, Science Press. стр. 342–350.
- ^ Хьюз, генеральный менеджер; Финарелли, Дж. А. (2019). «Размер репертуара обонятельных рецепторов динозавров» . Труды Королевского общества Б. 286 (1904): 20190909. doi : 10.1098/rspb.2019.0909 . ПМК 6571463 . PMID 31185870 .
- ^ Лаутеншлагер, С. (2013). «Краниальная миология и сила укуса Erlikosaurus andrewsi: новый подход к цифровой реконструкции мышц» . Журнал анатомии . 222 (2): 260–272. дои : 10.1111/joa.12000 . ПМЦ 3632231 . ПМИД 23061752 .
- ^ Лаутеншлагер, С.; Уитмер, Л.М.; Перл, А.; Рэйфилд, Э.Дж. (2013). «Эдентулизм, клювы и биомеханические инновации в эволюции тероподных динозавров» . ПНАС . 110 (51): 20657–20662. Бибкод : 2013PNAS..11020657L . дои : 10.1073/pnas.1310711110 . ПМК 3870693 . ПМИД 24297877 .
- ^ Лаутеншлагер, С. (2015). «Оценка краниальных скелетно-мышечных ограничений у тероподовых динозавров» . Королевское общество открытой науки . 2 (11): 150495. Бибкод : 2015RSOS....250495L . дои : 10.1098/rsos.150495 . ПМК 4680622 . ПМИД 26716007 .
- ^ Лаутеншлагер, С.; Брасси, Калифорния; Баттон, диджей; Барретт, премьер-министр (2016). «Раздельные формы и функции у разных клад травоядных динозавров» . Природа . 6 (26495): 26495. Бибкод : 2016NatSR...626495L . дои : 10.1038/srep26495 . ПМЦ 4873811 . ПМИД 27199098 .
- ^ Цыбин Ю.И.; Курзанов, С.М. (1979). «Новые данные о верхнемеловых местонахождениях позвоночных Байшин-Цавского района». Труды совместной советско-монгольской палеонтологической экспедиции . 8 : 108–112.
- ^ Jump up to: а б с Хикс, Дж. Ф.; Бринкман, Д.Л.; Николс, диджей; Ватабе, М. (1999). «Палеомагнитный и палинологический анализ отложений от альба до сантона в Байн Шире, Бурхант и Хурен Дух, восточная часть пустыни Гоби, Монголия» . Меловые исследования . 20 (6): 829–850. дои : 10.1006/cres.1999.0188 .
- ^ Курумада, Ю.; Аоки, С.; Аоки, К.; Като, Д.; Санеёси, М.; Цогтбаатар, К.; Уиндли, БФ; Исигаки, С. (2020). «Кальцитовый U-Pb возраст меловой формации Баин Шир, содержащей позвоночных, в восточной пустыне Гоби в Монголии: полезность калише для определения возраста» . Терра Нова 32 (4): 246–252. Бибкод : 2020Ноябрь..32..246K дои : 10.1111/тер.12456 .
- ^ Самойлов В.С.; Бенджамини, К. (1996). «Геохимические особенности останков динозавров из пустыни Гоби, Южная Монголия». ПАЛЕОС . 11 (6): 519–531. Бибкод : 1996Палай..11..519S . дои : 10.2307/3515188 . JSTOR 3515188 .
- ^ Jump up to: а б Ксепка, Д.Т.; Норелл, Массачусетс (2006). «Эркету эллисони, длинношеий зауропод из Бор Гуве (Дорноговский аймак, Монголия)» (PDF) . Американский музей Novitates (3508): 1–16. doi : 10.1206/0003-0082(2006)3508[1:EEALSF]2.0.CO;2 . S2CID 86032547 .
- ^ Перл, А.; Норелл, Массачусетс; Кларк, Дж. (1999). «Новый теропод-манираптор — Achillobator giganticus (Dromaeosauridae) — из верхнего мела Бурханта, Монголия» . Материалы кафедры геологии и минералогии Национального музея Монголии (101): 1–105. OCLC 69865262 .
- ^ Кобаяши, Ю.; Барсболд, Р. (2005). «Повторное исследование примитивного орнитомимозавра Garudimimus brevipes Barsbold, 1981 (Dinosauria: Theropoda) из позднего мела Монголии». Канадский журнал наук о Земле . 42 (9): 1501–1521. Бибкод : 2005CaJES..42.1501K . дои : 10.1139/e05-044 . hdl : 2115/14579 .
- ^ Парк, Дж. (2020). «Дополнительные черепа Talarurus plicatospineus (Dinosauria: Ankylosauridae) и значение для палеобиогеографии и палеоэкологии панцирных динозавров». Меловые исследования . 108 : е104340. дои : 10.1016/j.cretres.2019.104340 . S2CID 212423361 .
- ^ Серено, ПК (2000). «Летопись окаменелостей, систематика и эволюция пахицефалозавров и цератопсов Азии» (PDF) . Эпоха динозавров в России и Монголии . Издательство Кембриджского университета. стр. 489–491.
- ^ Ватабе, М.; Цогтбаатар, К.; Салливан, РМ (2011). «Новый пахицефалозаврид из формации Байншир (сеноман-поздний сантон), пустыня Гоби, Монголия» (PDF) . Ископаемая запись 3. Музей естественной истории и науки Нью-Мексико, Бюллетень . 53 : 489–497.
- ^ Данилов И.Г.; Хираяма, Р.; Суханов В.Б.; Сузуки, С.; Ватабе, М.; Витек, Н.С. (2014). «Меловые мягкопанцирные черепахи (Trionychidae) Монголии: новое разнообразие, записи и пересмотр» . Журнал систематической палеонтологии . 12 (7): 799–832. дои : 10.1080/14772019.2013.847870 . S2CID 86304259 .
- ^ Тернер, А.Х. (2015). «Обзор шамозуха и параллигатора (Crocodyliformes, Neosuria) из мелового периода Азии» . ПЛОС ОДИН . 10 (2): e0118116. Бибкод : 2015PLoSO..1018116T . дои : 10.1371/journal.pone.0118116 . ПМК 4340866 . ПМИД 25714338 .
- ^ Занно, Ле; Цогтбаатар, К.; Чинзориг, Т.; Гейтс, Т.А. (2016). «Специализации анатомии нижней челюсти и зубочелюстного аппарата Segnosaurus galbinensis (Theropoda: Therizinosauria) » ПерДж . 4 : е1885. дои : 10.7717/peerj.1885 . ПМЦ 4824891 . ПМИД 27069815 .
- ^ Лаутеншлагер, С. (2017). «Функциональное разделение ниш у Therizinosauria дает новое представление об эволюции травоядных теропод» (PDF) . Палеонтология . 60 (3): 375–387. дои : 10.1111/пала.12289 . S2CID 90965431 .
- ^ Баттон, диджей; Занно, LE (2019). «Повторяющаяся эволюция различных способов травоядности у нептичьих динозавров» . Современная биология . 30 (1): 158–168.e4. дои : 10.1016/j.cub.2019.10.050 . ПМИД 31813611 . S2CID 208652510 .









