млекопитающее
| Млекопитающие Временной диапазон: | |
|---|---|
 | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Клэйд : | Амниота |
| Клэйд : | Сторона синапса |
| Клэйд : | Млекопитающие |
| Сорт: | Млекопитающие Линней , 1758 г. |
| Живые подгруппы | |
Млекопитающее mamma (от латинского « грудь») [1] — позвоночное животное класса млекопитающих ( / m ə ˈ m eɪ l i . ə / ) . Млекопитающие характеризуются наличием молоко вырабатывающих молочных желез, для кормления детенышей, неокортексной области мозга, шерсти или волос и трех косточек среднего уха . Эти характеристики отличают их от рептилий и птиц , от которых их предки отделились в каменноугольном периоде более 300 миллионов лет назад. около 6400 современных видов Описано млекопитающих, разделенных на 29 отрядов .
Крупнейшими отрядами млекопитающих по числу видов являются грызуны , летучие мыши и эвлипотифлы (в том числе ежи , кроты и землеройки ). Следующие три — это приматы (включая людей , обезьян и лемуров ), парнокопытные (включая свиней , верблюдов и китов ) и хищные животные (включая кошек , собак и тюленей ).
Млекопитающие — единственные живые члены Synapsida ; эта клада вместе с Sauropsida (рептилиями и птицами) составляет более крупную кладу Amniota . Ранние синапсиды называются « пеликозаврами ». Более продвинутые терапсиды стали доминировать в средней перми . Млекопитающие произошли от цинодонтов , развитой группы терапсидов, в период от позднего триаса до ранней юры . Млекопитающие достигли своего современного разнообразия в палеогеновом и неогеновом периодах кайнозойской эры, после вымирания нептичьих динозавров , и были доминирующей группой наземных животных с 66 миллионов лет назад до настоящего времени.
Основной тип тела млекопитающих — четвероногий , и большинство млекопитающих используют свои четыре конечности для передвижения по земле ; но у некоторых конечности приспособлены к жизни на море , в воздухе , на деревьях , под землей или на двух ногах . Размеры млекопитающих варьируются от шмелиной летучей мыши длиной 30 м (98 футов) размером 30–40 мм (1,2–1,6 дюйма) до синего кита — возможно, самого крупного животного, когда-либо жившего на свете. Максимальная продолжительность жизни варьируется от двух лет у землероек до 211 лет у гренландского кита . Все современные млекопитающие рождают живых детенышей, за исключением пяти видов однопроходных , которые являются яйцекладущими млекопитающими. Самая богатая видами группа млекопитающих, инфракласс , называемый плацентами , имеет плаценту , которая обеспечивает питание плода во время беременности .
Большинство млекопитающих разумны , некоторые обладают большим мозгом, самосознанием и умением пользоваться инструментами . Млекопитающие могут общаться и издавать звуки несколькими способами, включая производство ультразвука , маркировку запахами , сигналы тревоги , пение , эхолокацию ; и, в случае с людьми, сложный язык . Млекопитающие могут организовываться в общества деления-слияния , гаремы и иерархии , но также могут быть одиночными и территориальными . Большинство млекопитающих полигинны , но некоторые могут быть моногамными или полиандрическими .
Одомашнивание многими видами млекопитающих людьми сыграло важную роль в неолитической революции и привело к тому, что сельское хозяйство заменило охоту и собирательство в качестве основного источника пищи для людей. Это привело к серьезной реструктуризации человеческого общества с кочевого образа жизни на оседлый, с усилением сотрудничества между все большими и большими группами и, в конечном итоге, с развитием первых цивилизаций . Одомашненные млекопитающие обеспечивали и продолжают обеспечивать энергию для транспорта и сельского хозяйства, а также продукты питания ( мясо и молочные продукты ), мех и кожу . На млекопитающих также охотятся и гоняют в спортивных целях, их держат в качестве домашних и рабочих животных различных типов, а также используют в качестве модельных организмов в науке. Млекопитающие изображались в искусстве со времен палеолита и появляются в литературе, кино, мифологии и религии. Снижение численности и исчезновение многих млекопитающих вызваны, прежде всего, браконьерством и разрушением среды обитания , прежде всего вырубкой лесов .
Классификация

Классификация млекопитающих претерпела несколько изменений с тех пор, как Карл Линней первоначально определил этот класс, и в настоящее время ни одна система классификации не является общепринятой. McKenna & Bell (1997) и Wilson & Reeder (2005) предоставили полезные недавние сборники. [2] Симпсон (1945) [3] обеспечивает систематику происхождения и взаимоотношений млекопитающих, которой учили повсеместно до конца 20 века.Однако с 1945 года постепенно было обнаружено большое количество новой и более подробной информации: палеонтологические данные были перекалиброваны, и за прошедшие годы наблюдалось много споров и прогресса относительно теоретических основ самой систематизации, отчасти благодаря новой концепции кладистика . Хотя полевые и лабораторные исследования постепенно устарели классификацию Симпсона, она остается наиболее близкой к официальной классификации млекопитающих, несмотря на известные проблемы. [4]
Большинство млекопитающих, включая шесть наиболее богатых видами отрядов , относятся к плацентарной группе. Три крупнейших по числу видов отряда — Rodentia : мыши , крысы , дикобразы , бобры , капибары и другие грызущие млекопитающие; Рукокрылые : летучие мыши; и Soricomorpha : землеройки , кроты и соленодоны . Следующие три крупнейших отряда, в зависимости от биологической классификации используемой схемы , — это приматы : человекообразные обезьяны , мартышки и лемуры ; Cetartiodactyla : киты и парнокопытные ; и Хищники , куда входят кошки , собаки , ласки , медведи , тюлени и их союзники. [5] По данным Mammal Species of the World , в 2006 году было выявлено 5416 видов. Они были сгруппированы в 1229 родов , 153 семейства и 29 отрядов. [5] В 2008 году Международный союз охраны природы (МСОП) завершил пятилетнюю глобальную оценку млекопитающих для своего Красного списка МСОП , в котором насчитывалось 5488 видов. [6] Согласно исследованию, опубликованному в Журнале маммологии в 2018 году, количество признанных видов млекопитающих составляет 6495, включая 96 недавно вымерших. [7]
Определения
Слово « млекопитающее » происходит от научного названия Mammalia, придуманного Карлом Линнеем в 1758 году и происходящего от латинского mamma («соска, папочка»). В влиятельной статье 1988 года Тимоти Роу филогенетически определил Mammalia как коронную группу млекопитающих, кладу, состоящую из самого недавнего общего предка ныне живущих однопроходных ( ехидны и утконосы ) и терианских млекопитающих ( сумчатые и плацентарные ) и всех потомков этого предка. [8] Поскольку этот предок жил в юрском периоде, определение Роу исключает всех животных из более раннего триаса , несмотря на то, что триасовые окаменелости в харамиидах с середины 19 века относят к млекопитающим. [9] Если Mammalia рассматривать как коронную группу, ее происхождение можно грубо датировать первым известным появлением животных, более тесно связанных с некоторыми современными млекопитающими, чем с другими. Амбондро более тесно связан с однопроходными, чем с териальными млекопитающими, в то время как амфилест и амфитерий более тесно связаны с терианами; поскольку окаменелости всех трех родов датируются примерно 167 миллионами лет назад в средней юре , это разумная оценка внешнего вида кроновой группы. [10]
Т. С. Кемп дал более традиционное определение: « Синапсиды , обладающие зубно - чешуйчатым сочленением челюстей и окклюзией между верхними и нижними коренными зубами с поперечным компонентом движения» или, что то же самое, по мнению Кемпа, клада, происходящая от последнего общего предка Синоконодон и современные млекопитающие. [11] Самым ранним известным синапсидом, удовлетворяющим определениям Кемпа, является Tikitherium , датированный 225 млн лет назад , поэтому появление млекопитающих в этом более широком смысле можно отнести к этой дате позднего триаса . [12] [13] Однако это животное, возможно, действительно эволюционировало в неогене. [14]
Классификация Маккенны / Белла
В 1997 году млекопитающие были тщательно пересмотрены Малкольмом К. Маккенной и Сьюзен К. Белл, в результате чего была создана классификация Маккенна/Белл. Авторы вместе работали палеонтологами в Американском музее естественной истории . Маккенна унаследовал проект от Симпсона и вместе с Беллом построил полностью обновленную иерархическую систему, охватывающую живые и вымершие таксоны, которая отражает историческую генеалогию млекопитающих. [4] Их книга 1997 года «Классификация млекопитающих выше уровня вида » [15] представляет собой комплексную работу по систематике, взаимоотношениям и распространению всех таксонов млекопитающих, ныне живущих и вымерших, вплоть до рода, хотя молекулярно-генетические данные ставят под сомнение некоторые из группировок.
В следующем списке вымершие группы отмечены кинжалом ( †).
Класс Млекопитающие
- Подкласс Prototheria : однопроходные: ехидны и утконосы.
- Подкласс Theriiformes : живородящие млекопитающие и их доисторические родственники.
- Инфракласс † Аллотерия : мультитуберкулезные.
- Инфракласс † Eutriconodonta : эвтриконодонты.
- Инфракласс голотерии : современные живородящие млекопитающие и их доисторические родственники.
- Суперлегион † Кюнеотерия
- Суперкогорта Theria : живородящие млекопитающие.
- Когорта Marsupialia : сумчатые.
- Магнодер Австралидельфия : австралийские сумчатые и монито-дель-монте.
- Магнодер Америдельфия : сумчатые Нового Света. Сейчас считается парафилетическим , при этом землеройки-опоссумы ближе к австралидельфиям. [16]
- Когорта Плаценталия : плаценты.
- Магнодер Ксенартра : ксенартран
- Магнодер Эпитерия : эпитерии
- Надотряд † Лептицида
- Надотряд Прептотерии
- Grandorder Anagalida : зайцеобразные , грызуны и землеройки-слоны.
- Grandorder Ferae : хищники , ящеры , † креодонты и родственники.
- Grandorder Lipotyphla : насекомоядные.
- Grandorder Archonta : летучие мыши , приматы , колуго и землеройки (теперь считаются парафилетическими, при этом летучие мыши ближе к другим группам)
- Grandorder Euungulata : копытные.
- Отряд Tubulidentata incertae sedis : трубкозуб.
- Mirorder Eparctocyona : † мыщелки , киты и парнокопытные (парнокопытные)
- Мирордёр † Meridiungulata : южноамериканские копытные.
- Миротряд Altungulata : непарнокопытные (непарнокопытные), слоны , ламантины и даманы.
- Когорта Marsupialia : сумчатые.
Молекулярная классификация плаценты

В начале 21 века молекулярные исследования, основанные на анализе ДНК , позволили предположить новые взаимоотношения между семействами млекопитающих. Большинство этих результатов были независимо подтверждены ретротранспозонов данными о присутствии/отсутствии . [18] Системы классификации, основанные на молекулярных исследованиях, выявляют три основные группы или линии плацентарных млекопитающих — Afrotheria , Xenarthra и Boreoeutheria , — которые разошлись в меловом периоде . Отношения между этими тремя линиями являются спорными, и были предложены все три возможные гипотезы относительно того, какая группа является базовой . Этими гипотезами являются Atlantogenata (базальная Boreoeutheria), Epitheria (базальная Xenarthra) и Exafroplacentalia (базальная Afrotheria). [19] Boreoeutheria, в свою очередь, включает две основные линии — Euarchontoglires и Laurasiatheria .
По оценкам, время расхождения между этими тремя плацентарными группами варьируется от 105 до 120 миллионов лет назад, в зависимости от типа используемой ДНК (например, ядерной или митохондриальной ). [20] и различные интерпретации палеогеографических данных. [19]
| Тарвер и др. 2016 год [21] | Сандра Альварес-Карретеро и др. 2022 год [22] [23] | |||
|---|---|---|---|---|
|

Эволюция
Происхождение
Synapsida , клада, включающая млекопитающих и их вымерших родственников, возникла во время пенсильванского субпериода (от ~323 до ~300 миллионов лет назад), когда они отделились от линии рептилий. Млекопитающие кроновой группы произошли от более ранних млекопитающих в ранней юре . На кладограмме Mammalia считается кронной группой. [24]
| Млекопитающие |
| ||||||||||||||||||||||||||||||||||||||||||
Эволюция от старых амниот

Первыми полностью наземными позвоночными были амниоты . Как и их ранние четвероногие предшественники-амфибии, у них были легкие и конечности. Однако амниотические яйца имеют внутренние мембраны, которые позволяют развивающемуся эмбриону дышать, но удерживают воду. Следовательно, амниоты могут откладывать яйца на суше, тогда как земноводным обычно приходится откладывать яйца в воде.
Первые амниоты, по-видимому, возникли в пенсильванском подпериоде каменноугольного периода . Они произошли от более ранних рептилийоморфных четвероногих амфибий. [25] которые жили на земле, уже заселенной насекомыми и другими беспозвоночными, а также папоротниками , мхами и другими растениями. В течение нескольких миллионов лет стали различаться две важные линии амниот: синапсиды , которые позже включали общего предка млекопитающих; и зауропсиды , к которым сейчас относятся черепахи , ящерицы , змеи , крокодилы и динозавры (включая птиц ). [26] Синапсиды имеют по одному отверстию ( височному оконному стеклу ) внизу с каждой стороны черепа. Примитивные синапсиды включали самых крупных и свирепых животных ранней перми , таких как диметродон . [27] Синапсиды немлекопитающих традиционно (и неправильно) назывались «рептилиями, подобными млекопитающим» или пеликозаврами ; теперь мы знаем, что они не были ни рептилиями, ни частью линии рептилий. [28] [29]
Терапсиды , группа синапсид, развились в средней перми , около 265 миллионов лет назад, и стали доминирующими наземными позвоночными. [28] Они отличаются от базальных эвпеликозавров некоторыми особенностями черепа и челюстей, в том числе: более крупными черепами и резцами , которые у терапсидов одинаковы по размеру, но не у эвпеликозавров. [28] Линия терапсидов, ведущая к млекопитающим, прошла ряд стадий, начиная с животных, которые были очень похожи на своих ранних синапсидных предков, и заканчивая пробайногнатическими цинодонтами , некоторых из которых можно было легко принять за млекопитающих. Эти этапы характеризовались: [30]
- Постепенное развитие костного вторичного неба .
- Резкое приобретение эндотермии среди Mammaliamorpha , то есть до возникновения млекопитающих на 30–50 миллионов лет. [31] .
- Переход к прямому положению конечностей, что повысит выносливость животных за счет избежания ограничений Кэрриера . Но этот процесс был медленным и беспорядочным: например, все травоядные терапсиды, не относящиеся к млекопитающим, сохранили раскидистые конечности (некоторые поздние формы могли иметь полупрямые задние конечности); Пермские хищные терапсиды имели раскидистые передние конечности, а некоторые позднепермские - полураскидистые задние конечности. Фактически, современные однопроходные все еще имеют полураскидистые конечности.
- постепенно Зубная кость стала основной костью нижней челюсти, которая к триасу превратилась в полностью челюсть млекопитающих (нижняя часть состоит только из зубной кости) и среднего уха (которое построено из костей, которые ранее использовались для построения челюстей). рептилий).
Первые млекопитающие
Пермско -триасовое вымирание около 252 миллионов лет назад, которое было длительным событием из-за накопления нескольких импульсов вымирания, положило конец доминированию хищных терапсидов. [32] В раннем триасе большинство ниш наземных хищников среднего и крупного размера были заняты архозаврами. [33] который в течение длительного периода (35 миллионов лет) стал включать крокодиломорфов , [34] птерозавры ; и динозавры [35] однако крупные цинодонты, такие как Trucidocynodon и traversodontids, все еще занимали крупные ниши плотоядных и травоядных соответственно. К юрскому периоду динозавры стали доминировать и в крупных нишах наземных травоядных. [36]
Первые млекопитающие (в понимании Кемпа) появились в эпоху позднего триаса (около 225 миллионов лет назад), через 40 миллионов лет после первых терапсидов. они вышли из своей ночной ниши насекомоядных животных ; Начиная с середины юрского периода, [37] Юрский касторокауда , например, был близким родственником настоящих млекопитающих, у которых были приспособления для плавания, рытья и ловли рыбы. [38] Считается, что большинство, если не все, оставались ночными ( ночное узкое место ), что объясняет большую часть типичных черт млекопитающих. [39] Большинство видов млекопитающих, существовавших в мезозойскую эру, были мультитуберкулятами, эвтриконодонтами и спалакотериидами . [40] Самый ранний известный метатерий — Sinodelphys возрастом 125 миллионов лет раннего мела , найденный в сланцах в северо-восточной китайской провинции Ляонин . Окаменелость почти полная и включает в себя пучки меха и отпечатки мягких тканей. [41]

Самая старая известная окаменелость среди Eutheria («настоящие звери») — это маленькая землероекка Juramaia sinensis , или «юрская мать из Китая», датируемая 160 миллионами лет назад, в позднем юрском периоде. [42] Более поздний родственник эвтерий, Eomaia , датируемый 125 миллионами лет назад в раннем меловом периоде, обладал некоторыми чертами, общими с сумчатыми, но не с плацентарными, что свидетельствует о том, что эти черты присутствовали у последнего общего предка двух групп, но были позже обнаружены. потерян в плацентарной линии. [43] В частности, надлобковые кости выступают вперед от таза. Они не обнаружены ни в одной современной плаценте, но они обнаружены у сумчатых, однопроходных, других нетерийных млекопитающих и Ukhaatherium , раннего мелового животного из эвтерийского отряда Asioryctitheria . Это относится и к мультитуберкулезам. [44] По-видимому, они являются наследственной особенностью, которая впоследствии исчезла в плацентарной линии. Эти надлобковые кости, по-видимому, функционируют, напрягая мышцы во время передвижения, уменьшая объем представляемого пространства, которое требуется плаценте для содержания плода во время беременности. Узкое выходное отверстие таза указывает на то, что детеныши при рождении были очень маленькими и поэтому беременность была короткой, как у современных сумчатых. Это говорит о том, что плацента возникла позднее. [45]
Одним из самых ранних известных однопроходных был тейнолофос , живший около 120 миллионов лет назад в Австралии. [46] У однопроходных есть некоторые особенности, которые могут быть унаследованы от первоначальных амниот, например, одно и то же отверстие для мочеиспускания, дефекации и размножения ( клоака ), что также есть у ящериц и птиц. [47] и они откладывают яйца , кожистые и некальцинированные. [48]
Самые ранние появления функций
Hadrocodium , чьи окаменелости датируются примерно 195 миллионами лет назад, в ранней юре , представляет собой первое явное свидетельство существования челюстного сустава, образованного исключительно чешуйчатой и зубной костями; в челюсти нет места для суставной кости, входящей в состав челюстей всех ранних синапсид. [49]

Самые ранние явные свидетельства наличия волос или меха встречаются в окаменелостях Castorocauda и Megaconus , датируемых 164 миллионами лет назад в середине юрского периода. В 1950-х годах было высказано предположение, что отверстия (ходы) в верхних челюстях и предчелюстных костях (костях передней части верхней челюсти) цинодонтов представляют собой каналы, которые снабжают кровеносные сосуды и нервы вибриссами ( усами ) и, таким образом, свидетельствуют о наличии волос или шерсть; [50] [51] Однако вскоре было отмечено, что отверстия не обязательно указывают на то, что у животного были вибриссы, поскольку у современной ящерицы Tupinambis есть отверстия, которые почти идентичны тем, которые обнаружены у немлекопитающих цинодонтов Thrinaxodon . [29] [52] Популярные источники, тем не менее, продолжают приписывать усы Тринаксодону . [53] Исследования пермских копролитов немлекопитающих показывают, что синапсиды той эпохи уже имели мех, что указывает на то, что эволюция волос, возможно, началась еще у дицинодонтов . [54]
Когда эндотермия впервые появилась в эволюции млекопитающих, неизвестно, хотя общепринято считать, что она впервые возникла у терапсидов, не относящихся к млекопитающим . [54] [55] Современные однопроходные имеют более низкую температуру тела и более изменчивую скорость метаболизма, чем сумчатые и плацентарные. [56] но есть свидетельства того, что некоторые из их предков, возможно, включая предков терианцев, могли иметь температуру тела, подобную температуре тела современных терианцев. [57] Точно так же у некоторых современных терианцев, таких как афротеры и ксенартраны, вторично развилась более низкая температура тела. [58]
Эволюция прямостоячих конечностей у млекопитающих не завершена: у живых и ископаемых однопроходных конечности раскидистые. Парасагиттальное (нераскинутое) положение конечностей появилось где-то в поздней юре или раннем мелу; он обнаружен у эвтерийных Eomaia и метатерийных Sinodelphys , возраст которых датируется 125 миллионами лет назад. [59] Надлобковые кости, особенность, которая сильно повлияла на размножение большинства клад млекопитающих, впервые обнаружены у Tritylodontidae , что позволяет предположить, что это синапоморфия между ними и млекопитающими . Они повсеместно присутствуют у неплацентарных млекопитающих, хотя у Megazostrodon и Erythrotherium они, по-видимому, отсутствуют. [60]
Было высказано предположение, что первоначальная функция лактации ( производства молока ) заключалась в поддержании яиц влажными. Большая часть аргументов основана на однопроходных, млекопитающих, откладывающих яйца. [61] [62] У женщин молочные железы полностью развиваются в период полового созревания, независимо от беременности. [63]
Восстание млекопитающих

Терианские млекопитающие заняли средние и крупные экологические ниши в кайнозое после того, как мел-палеогеновое вымирание примерно 66 миллионов лет назад опустошило экологическое пространство, когда-то заполненное нептичьими динозаврами и другими группами рептилий, а также различными другими видами. группы млекопитающих, [65] и претерпел экспоненциальное увеличение размеров тела ( мегафауна ). [66] Затем млекопитающие очень быстро диверсифицировались; и птицы, и млекопитающие демонстрируют экспоненциальный рост разнообразия. [65] Например, самая ранняя из известных летучих мышей появилась примерно 50 миллионов лет назад, всего через 16 миллионов лет после вымирания нептичьих динозавров. [67]
Молекулярно-филогенетические исследования первоначально предполагали, что большинство плацентарных отрядов разошлись примерно от 100 до 85 миллионов лет назад и что современные семьи появились в период с позднего эоцена по миоцен . [68] Однако до конца мелового периода не было обнаружено никаких окаменелостей плаценты. [69] Самые ранние бесспорные окаменелости плаценты относятся к раннему палеоцену , после вымирания нептичьих динозавров. [69] (Ученые идентифицировали животное раннего палеоцена по имени Protungulatum donnae как одно из первых плацентарных млекопитающих. [70] но с тех пор его переклассифицировали как неплацентарный евтериальный.) [71] Перекалибровка показателей генетического и морфологического разнообразия позволила предположить позднемеловое происхождение плацент и палеоценовое происхождение большинства современных клад. [72]
Самый ранний известный предок приматов — Архицебус Ахиллес. [73] примерно 55 миллионов лет назад. [73] Этот крошечный примат весил 20–30 граммов (0,7–1,1 унции) и мог поместиться на человеческой ладони. [73]
Анатомия
Отличительные особенности
Живые виды млекопитающих можно идентифицировать по наличию потовых желез , в том числе тех, которые специализируются на выработке молока для питания детенышей. [74] Однако при классификации окаменелостей необходимо использовать и другие признаки, поскольку в окаменелостях не видны железы мягких тканей и многие другие признаки. [75]
Многие черты, общие для всех ныне живущих млекопитающих, появились у самых ранних членов группы:
- Челюстной сустав . Зубная кость (кость нижней челюсти, на которой расположены зубы) и чешуйчатая кость (небольшая черепная кость) встречаются, образуя сустав. У большинства челюстноротых , включая ранних терапсидов , сустав состоит из суставного (небольшая кость в задней части нижней челюсти) и квадратного (небольшая кость в задней части верхней челюсти). [49]
- Среднее ухо . У млекопитающих коронной группы звук переносится от барабанной перепонки цепочкой из трех костей: молоточка , наковальни и стремени . Наследственно молоточек и наковальня произошли от суставной и квадратной костей, которые составляли челюстной сустав ранних терапсидов. [76]
- Замена зубов . Зубы могут быть заменены один раз ( дифиодонтия ) или (как у зубатых китов и мюридовых грызунов) не сменяться вообще ( монофиодонтия ). [77] У слонов, ламантинов и кенгуру на протяжении всей жизни постоянно вырастают новые зубы ( полифиодонтия ). [78]
- Призматическая эмаль . Эмалевое покрытие на поверхности зуба состоит из призм, твердых стержнеобразных структур, идущих от дентина к поверхности зуба. [79]
- Затылочные мыщелки . Две выступы у основания черепа подходят к самому верхнему шейному позвонку ; у большинства других четвероногих , напротив, есть только одна такая ручка. [80]
По большей части эти характеристики отсутствовали у триасовых предков млекопитающих. [81] Почти все млекопитающие имеют надлобковую кость, за исключением современных плацентарных. [82]
Половой диморфизм

В среднем самцы млекопитающих крупнее самок, при этом самцы как минимум на 10% крупнее самок более чем у 45% исследованных видов. , ориентированный на самцов Большинство отрядов млекопитающих также демонстрируют половой диморфизм , хотя некоторые отряды не демонстрируют какой-либо предвзятости или имеют значительную предвзятость к самкам ( Lagomorpha ). Половой диморфизм размеров увеличивается с размером тела у млекопитающих ( правило Ренша ), что позволяет предположить, что существует параллельное давление отбора как на размер самцов, так и на самок. Диморфизм, ориентированный на самцов, связан с половым отбором самцов посредством конкуренции между самцами за самок, поскольку существует положительная корреляция между степенью полового отбора, на что указывают системы спаривания , и степенью диморфизма размера, ориентированного на самцов. Степень полового отбора также положительно коррелирует с размером самцов и самок млекопитающих. Кроме того, параллельное давление отбора на массу самок определяется тем, что возраст отъема от груди значительно выше у более полигинных видов, даже с поправкой на массу тела. Кроме того, скорость размножения ниже у более крупных самок, что указывает на то, что отбор по плодовитости у млекопитающих осуществляется в пользу более мелких самок. Хотя эти закономерности сохраняются для млекопитающих в целом, между отрядами существуют значительные различия. [83]
Биологические системы
У большинства млекопитающих семь шейных позвонков (костей шеи). Исключение составляют ламантин и двупалый ленивец , у которых их шесть, а также трехпалый ленивец , у которого их девять. [84] Мозг всех млекопитающих имеет неокортекс — область мозга, уникальную для млекопитающих. [85] Плацентарный мозг имеет мозолистое тело , в отличие от однопроходных и сумчатых. [86]




Кровеносные системы
млекопитающих Сердце состоит из четырех камер: двух верхних предсердий (приемных камер) и двух нижних желудочков (выбрасывающих камер). [87] Сердце имеет четыре клапана, которые разделяют его камеры и обеспечивают ток крови через сердце в правильном направлении (предотвращая обратный ток). После газообмена в легочных капиллярах (кровеносных сосудах легких) обогащенная кислородом кровь возвращается в левое предсердие через одну из четырех легочных вен . Кровь почти непрерывно течет обратно в предсердие, которое действует как приемная камера, и отсюда через отверстие в левый желудочек. Большая часть крови пассивно поступает в сердце, пока предсердия и желудочки расслаблены, но к концу периода релаксации желудочков левое предсердие сокращается, перекачивая кровь в желудочек. Сердцу также необходимы питательные вещества и кислород, которые содержатся в крови, как и другие мышцы, и снабжаются ими через коронарные артерии . [88]
Дыхательные системы
Легкие млекопитающих губчатые и сотовые. Дыхание осуществляется преимущественно с помощью диафрагмы , которая отделяет грудную клетку от брюшной полости, образуя выпуклый к грудной клетке купол. Сокращение диафрагмы уплощает купол, увеличивая объем полости легкого. Воздух поступает через ротовую и носовую полости, проходит через гортань, трахею и бронхи , расширяет альвеолы . Расслабление диафрагмы имеет противоположный эффект: объем полости легких уменьшается, что приводит к вытеснению воздуха из легких. Во время тренировки брюшная стенка сокращается , увеличивая давление на диафрагму, которая выталкивает воздух быстрее и сильнее. Грудная клетка способна расширять и сжимать грудную полость под действием других дыхательных мышц. Следовательно, воздух всасывается в легкие или выбрасывается из них, всегда перемещаясь по градиенту давления. [89] [90] Этот тип легких известен как сильфонное легкое из-за его сходства с кузнечными мехами . [90]
Покровные системы
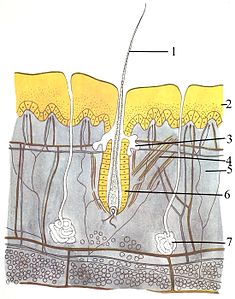
Покровная система (кожа) состоит из трех слоев: внешнего эпидермиса , дермы и гиподермы . Эпидермис обычно имеет толщину от 10 до 30 клеток; его основная функция — создание водонепроницаемого слоя. Его внешние клетки постоянно теряются; его самые нижние клетки постоянно делятся и продвигаются вверх. Средний слой, дерма, в 15–40 раз толще эпидермиса. Дерма состоит из многих компонентов, таких как костные структуры и кровеносные сосуды. Гиподерма состоит из жировой ткани , которая хранит липиды и обеспечивает амортизацию и изоляцию. Толщина этого слоя широко варьируется от вида к виду; [91] : 97 морским млекопитающим требуется толстая гиподерма ( жир ) для изоляции, а у китов самый толстый жир — 20 дюймов (51 см). [92] Хотя у других животных есть такие особенности, как усы, перья , щетинки или реснички , которые внешне напоминают его, ни у одного животного, кроме млекопитающих, нет волос . Это определяющая характеристика класса, хотя у некоторых млекопитающих ее очень мало. [91] : 61
Пищеварительная система


Травоядные животные развили разнообразные физические структуры, облегчающие потребление растительного материала . Чтобы разрушить неповрежденные растительные ткани, млекопитающие развили структуру зубов , отражающую их пищевые предпочтения. Например, плодоядные (животные, питающиеся в основном фруктами) и травоядные, питающиеся мягкой листвой, имеют зубы с низкой коронкой, предназначенные для измельчения листвы и семян . Пасущиеся животные, которые питаются твердой, богатой кремнеземом травой, имеют зубы с высокой коронкой, которые способны перемалывать твердые ткани растений и не изнашиваются так быстро, как зубы с низкой коронкой. [93] У большинства плотоядных млекопитающих зубы карнассиальной формы (разной длины в зависимости от рациона), длинные клыки и схожие закономерности замены зубов. [94]
Желудок парнокопытных (Artiodactyla) разделен на четыре отдела: рубец , сетку , книжку и сычуг (рубец есть только у жвачных ). После употребления растительного материала он смешивается со слюной в рубце и сетке и разделяется на твердый и жидкий материал. Твердые вещества комкаются вместе, образуя комок (или жвачку ), и отрыгиваются. При попадании комка в рот жидкость выдавливается языком и снова проглатывается. Поглощенная пища попадает в рубец и ретикулум, где целлюлолитические микробы ( бактерии , простейшие и грибы ) производят целлюлазу , которая необходима для расщепления целлюлозы в растениях. [95] Непарнокопытные , в отличие от жвачных, запасают переваренную пищу, покинувшую желудок, в расширенной слепой кишке , где она ферментируется бактериями. [96] У хищных животных простой желудок, приспособленный для переваривания в основном мяса, в отличие от сложной пищеварительной системы травоядных животных, которая необходима для расщепления жестких и сложных растительных волокон. Слепая кишка либо отсутствует, либо короткая и простая, а толстая кишка не мешотчатая или значительно шире тонкой кишки. [97]
Выделительная и мочеполовая системы


млекопитающих Выделительная система включает в себя множество компонентов. Как и большинство других наземных животных, млекопитающие являются уреотеликами и преобразуют аммиак в мочевину , что осуществляется печенью как часть цикла мочевины . [98] Билирубин , продукт жизнедеятельности клеток крови , выводится через желчь и мочу с помощью ферментов, выделяемых печенью. [99] Прохождение билирубина с желчью через кишечный тракт придает фекалиям млекопитающих характерную коричневую окраску. [100] К отличительным особенностям почек млекопитающих относится наличие почечной лоханки и почечных пирамид , четко различимых коркового и мозгового слоя , что обусловлено наличием удлиненных петель Генле . Только почка млекопитающих имеет бобовидную форму, хотя есть и некоторые исключения, например многолопастные ретикуляционные почки ластоногих, китообразных и медведей. [101] [102] У большинства взрослых плацентарных млекопитающих не осталось следов клоаки . У эмбриона эмбриональная клоака разделяется на задний отдел, который становится частью ануса, и передний отдел, судьба которого различна в зависимости от пола особи: у самок она развивается в преддверие или мочеполовой синус , куда попадает уретра. и влагалище , тогда как у мужчин он образует всю уретру полового члена . [102] [103] Однако тенреки , золотые кроты и некоторые землероек сохраняют клоаку и во взрослом возрасте. [104] У сумчатых половые пути отделены от ануса, но следы исходной клоаки все же остаются снаружи. [102] Monotremes, что в переводе с греческого означает «единственное отверстие», имеет настоящую клоаку. [105] Моча течет из мочеточников в клоаку у однопроходных и в мочевой пузырь у плацентарных млекопитающих. [102]
Звуковое производство

Как и у всех других четвероногих, у млекопитающих есть гортань , которая может быстро открываться и закрываться для воспроизведения звуков, а также надгортанный голосовой тракт , который фильтрует этот звук. Легкие и окружающая мускулатура обеспечивают поток воздуха и давление, необходимые для фонации . Гортань контролирует высоту и громкость звука, но сила, с которой легкие выдыхают, также способствует увеличению громкости. Более примитивные млекопитающие, например ехидна, могут только шипеть, поскольку звук достигается исключительно за счет выдоха через частично закрытую гортань. Другие млекопитающие голосуют с помощью голосовых связок . Движение или напряжение голосовых связок может привести к появлению множества звуков, таких как мурлыканье и крик . Млекопитающие могут менять положение гортани, позволяя им дышать через нос, глотать через рот, а также формировать как ротовые, так и носовые звуки; носовые звуки, такие как собачье вой, обычно тихие, а оральные звуки, такие как лай собаки, обычно громкие. [106]
У некоторых млекопитающих большая гортань и, следовательно, низкий голос, а именно у летучей мыши-молота ( Hypsignathus monstrosus ), у которой гортань может занимать всю грудную полость , одновременно выталкивая легкие, сердце и трахею в брюшную полость . [107] Большие голосовые подушечки также могут понижать высоту звука, как в случае с низким рыком больших кошек . [108] Производство инфразвука возможно у некоторых млекопитающих, таких как африканский слон ( виды Loxodonta ) и усатые киты . [109] [110] Мелкие млекопитающие с маленькой гортанью обладают способностью производить ультразвук , который можно обнаружить с помощью изменений среднего уха и улитки . Ультразвук не слышен птицам и рептилиям, что могло быть важно во времена мезозоя, когда птицы и рептилии были доминирующими хищниками. Этот частный канал используется некоторыми грызунами, например, при общении между матерью и щенком, а также летучими мышами при эхолокации. Зубатые киты также используют эхолокацию, но, в отличие от голосовой перепонки, простирающейся вверх от голосовых складок, у них есть дыня для манипулирования звуками. У некоторых млекопитающих, а именно приматов, к гортани прикреплены воздушные мешки, которые могут снижать резонансы или увеличивать громкость звука. [106]
Система голосовой продукции контролируется ядрами черепных нервов головного мозга и снабжается возвратным гортанным нервом и верхним гортанным нервом , ветвями блуждающего нерва . Голосовой тракт иннервируется подъязычным нервом и лицевыми нервами . Электрическая стимуляция околоводопроводной серой (ПЭГ) области среднего мозга млекопитающих вызывает вокализацию. Способность изучать новые вокализации наблюдается только у людей, тюленей, китообразных, слонов и, возможно, летучих мышей; у человека это результат прямой связи между моторной корой , управляющей движением, и мотонейронами спинного мозга. [106]
Шерсть

Основная функция меха млекопитающих — терморегуляция . Другие включают защиту, сенсорные цели, гидроизоляцию и камуфляж. [111] Разные виды меха служат разным целям: [91] : 99
- Окончательный - который может потеряться после достижения определенной длины.
- Вибриссы - чувствующие волоски, чаще всего усы.
- Шерсть – остевые волосы, подшерсток и ость.
- Шипы – жесткие остевые волосы, используемые для защиты (например, у дикобразов ).
- Щетина – длинные волоски, обычно используемые для визуальных сигналов. (например, львиная грива )
- Велли - часто называемый «пуховым мехом», который изолирует новорожденных млекопитающих.
- Шерсть – длинная, мягкая и часто вьющаяся.
Терморегуляция
Длина волос не является фактором терморегуляции: например, у некоторых тропических млекопитающих, таких как ленивцы, длина меха такая же, как у некоторых арктических млекопитающих, но с меньшей изоляцией; и, наоборот, другие тропические млекопитающие с короткой шерстью обладают такой же изолирующей способностью, как и арктические млекопитающие. Густота меха может повысить изоляционные свойства животного, а у арктических млекопитающих особенно густой мех; например, у овцебыка есть остевые волосы длиной 30 см (12 дюймов), а также густой подшерсток, образующий воздухонепроницаемую шерсть, позволяющую им выживать при температуре -40 ° C (-40 ° F). [91] : 162–163 Некоторые млекопитающие пустыни, такие как верблюды, используют густой мех, чтобы предотвратить попадание солнечного тепла на кожу, что позволяет животному сохранять прохладу; Летом температура верблюжьего меха может достигать 70 ° C (158 ° F), но температура кожи остается 40 ° C (104 ° F). [91] : 188 Водные млекопитающие , наоборот, задерживают воздух в своем мехе, чтобы сохранить тепло, сохраняя кожу сухой. [91] : 162–163

Окраска
Шерсть млекопитающих окрашена по разным причинам, основные факторы отбора включают камуфляж , половой отбор , общение и терморегуляцию. Окраска как волос, так и кожи млекопитающих определяется главным образом типом и количеством меланина ; эумеланин для коричневого и черного цвета и феомеланин для диапазона от желтоватого до красноватого цвета, придающего млекопитающим землистый оттенок . [112] [113] У некоторых млекопитающих более яркие цвета; некоторые обезьяны, такие как мандрилы и верветки , а также опоссумы, такие как мексиканские опоссумы-мыши и шерстистые опоссумы Дерби , имеют синюю кожу из-за дифракции света в коллагеновых волокнах. [114] Многие ленивцы кажутся зелеными, потому что в их мехе обитают зеленые водоросли ; это может быть симбиотическая связь, позволяющая маскироваться . ленивцам [115]
Камуфляж оказывает сильное влияние на большое количество млекопитающих, поскольку помогает скрыть особей от хищников или добычи. [116] У арктических и субарктических млекопитающих, таких как песец ( Alopex lagopus ), лемминг с ошейником ( Dicrostonyx groenlandicus ), горностай ( Mustela erminea ) и заяц-беляк ( Lepus americanus ), сезонное изменение цвета между коричневым летом и белым зимой обусловлено главным образом по камуфляжу. [117] Некоторые древесные млекопитающие, особенно приматы и сумчатые, имеют оттенки фиолетовой, зеленой или синей кожи на частях тела, что указывает на определенное преимущество в их преимущественно древесной среде обитания благодаря конвергентной эволюции . [114]
Апосематизм , предупреждающий возможных хищников, является наиболее вероятным объяснением черно-белой шерсти многих млекопитающих, способных защитить себя, например, у дурно пахнущего скунса и мощного и агрессивного медоеда . [118] Цвет шерсти иногда имеет половой диморфизм , как у многих видов приматов . [119] Различия в окрасе шерсти самок и самцов могут указывать на питание и уровень гормонов, что важно при выборе партнера. [120] Цвет шерсти может влиять на способность сохранять тепло в зависимости от того, сколько света отражается. Млекопитающие с более темной шерстью могут поглощать больше тепла от солнечной радиации и сохранять тепло, а у некоторых более мелких млекопитающих, таких как полевки , зимой мех темнее. Белый беспигментированный мех арктических млекопитающих, таких как белый медведь, может отражать больше солнечной радиации непосредственно на кожу. [91] : 166–167 [111] Ослепительные черно-белые полосы зебр , по-видимому, обеспечивают некоторую защиту от укусов мух. [121]
Репродуктивная система

Млекопитающие исключительно гонохорны (животное рождается либо с мужскими, либо с женскими гениталиями, в отличие от гермафродитов , у которых такого раскола нет). [122] Пенис совокупления используется как для мочеиспускания, так и для у мужских плацент. [123] и сумчатые. [124] В зависимости от вида эрекция может вызываться притоком крови в сосуды, губчатую ткань или мышечным действием. В не эрегированном состоянии половой член может находиться в крайней плоти , а в некоторых плацентах также имеется кость полового члена ( бакулюм ). [125] У сумчатых обычно раздвоенные пенисы. [126] тогда как пенис ехидны обычно имеет четыре головки, из которых функционируют только две. [127] Яички . большинства млекопитающих опускаются в мошонку , которая обычно находится позади полового члена, но у сумчатых часто находится спереди У самок млекопитающих обычно имеется вульва ( клитор и половые губы ) снаружи, тогда как внутренняя система содержит парные яйцеводы , 1–2 матки , 1–2 шейки матки и влагалище . [128] [129] У сумчатых есть два боковых влагалища и одно медиальное влагалище. «Влагалище» однопроходных лучше понимать как «мочеполовой синус». Маточные системы плацентарных млекопитающих могут варьироваться от дуплексной, когда имеются две матки и шейки матки, открывающиеся во влагалище, и двудольной, когда два рога матки имеют одну шейку, которая соединяется с влагалищем, и двурогой , состоящей из двух маточных систем. рога, которые соединены дистально, но разделены медиально, образуя Y-образную форму, и симплекс, имеющий единственную матку. [130] [131] [91] : 220–221, 247

Исходным условием размножения млекопитающих является рождение относительно неразвитых детенышей либо путем прямого живорождения , либо в течение короткого периода в виде яиц с мягкой скорлупой. Вероятно, это связано с тем, что туловище не могло расширяться из-за наличия надлобковых костей . Самая старая демонстрация этого репродуктивного стиля - Kayentatherium , который произвел неразвитых перинатов , но в помете гораздо большего размера, чем у любого современного млекопитающего, - 38 особей. [132] Большинство современных млекопитающих являются живородящими , рожающими живых детенышей. Однако пять видов однопроходных, утконос и четыре вида ехидны откладывают яйца. У однопроходных система определения пола отличается от системы большинства других млекопитающих. [133] В частности, половые хромосомы утконоса больше похожи на хромосомы курицы, чем на хромосомы терийского млекопитающего. [134]
Живородящие млекопитающие относятся к подклассу Theria; живущие сегодня относятся к сумчатым и плацентарным инфраклассам. короткий У сумчатых период беременности , обычно короче эстрального цикла , и обычно они рождают несколько неразвитых новорожденных, которые затем подвергаются дальнейшему развитию; у многих видов это происходит внутри мешочка, сумчатого матери , расположенного в передней части живота . Это плезиоморфное состояние среди живородящих млекопитающих; наличие надлобковых костей у всех неплацентарных млекопитающих препятствует расширению туловища, необходимому для полноценной беременности. [82] Даже неплацентарные особи, вероятно, размножались таким образом. [44] Плаценты рождают относительно полноценных и развитых детенышей, обычно после длительной беременности. [135] Свое название они получили от плаценты , которая соединяет развивающийся плод со стенкой матки, обеспечивая поглощение питательных веществ. [136] У плацентарных млекопитающих надлобковая кость либо полностью утрачена, либо преобразована в бакулюм; позволяя туловищу расширяться и, таким образом, рождать развитое потомство. [132]
Молочные железы млекопитающих специализируются на выработке молока, основного источника питания новорожденных. Однопроходные рано ответвились от других млекопитающих и не имеют сосков, характерных для большинства млекопитающих, но у них есть молочные железы. Детеныши слизывают молоко с участка молочной железы на животе матери. [137] По сравнению с плацентарными млекопитающими молоко сумчатых сильно меняется как по продуктивности, так и по составу питательных веществ из-за недоразвитости молодняка. Кроме того, молочные железы обладают большей автономией, что позволяет им поставлять отдельное молоко детенышам на разных стадиях развития. [138] Лактоза является основным сахаром в молоке плацентарных млекопитающих, тогда как в молоке однопроходных и сумчатых преобладают олигосахариды . [139] Отлучение от груди — это процесс, при котором млекопитающее становится менее зависимым от материнского молока и больше от твердой пищи. [140]
Эндотермия
Почти все млекопитающие эндотермичны («теплокровны»). У большинства млекопитающих также есть волосы, которые помогают им сохранять тепло. Как и птицы, млекопитающие могут добывать пищу или охотиться в погоде и климате, слишком холодных для экзотермических («хладнокровных») рептилий и насекомых. Эндотермия требует большого количества пищевой энергии, поэтому млекопитающие съедают больше пищи на единицу массы тела, чем большинство рептилий. [141] Мелкие насекомоядные млекопитающие едят огромные количества пищи для своего размера. Редкое исключение — голый землекоп выделяет мало метаболического тепла, поэтому его считают оперативным пойкилотермом . [142] Птицы также эндотермичны, поэтому эндотермия присуща не только млекопитающим. [143]
Продолжительность жизни вида
Среди млекопитающих максимальная продолжительность жизни видов значительно варьируется (например, продолжительность жизни землеройки составляет два года, тогда как зарегистрированный возраст самого старого гренландского кита составляет 211 лет). [144] Хотя основная причина этих различий в продолжительности жизни все еще неясна, многочисленные исследования показывают, что способность восстанавливать повреждения ДНК является важным фактором, определяющим продолжительность жизни млекопитающих. В исследовании Харта и Сетлоу, проведенном в 1974 году, [145] Было обнаружено, что способность к эксцизионному восстановлению ДНК систематически увеличивается с увеличением продолжительности жизни семи видов млекопитающих. Было обнаружено, что продолжительность жизни видов тесно коррелирует со способностью распознавать двухцепочечные разрывы ДНК, а также с уровнем белка репарации ДНК Ku80 . [144] В исследовании клеток шестнадцати видов млекопитающих было обнаружено, что гены, участвующие в репарации ДНК, активируются у долгоживущих видов. [146] клеточный уровень фермента репарации ДНК поли-АДФ-рибозо-полимеразы коррелирует с продолжительностью жизни видов. При исследовании 13 видов млекопитающих было обнаружено, что [147] Три дополнительных исследования различных видов млекопитающих также сообщили о корреляции между продолжительностью жизни видов и способностью к восстановлению ДНК. [148] [149] [150]
Передвижение
Земной

Большинство позвоночных животных — земноводные, рептилии и некоторые млекопитающие, такие как человек и медведи — стопоходящие , ходят на всей нижней стороне стопы. Многие млекопитающие, такие как кошки и собаки, ходят на пальцах ног , причем большая длина шага обеспечивает большую скорость. Некоторые животные, например лошади , негулиградны и ходят на кончиках пальцев. Это еще больше увеличивает длину шага и, следовательно, скорость. [151] Известно, что некоторые млекопитающие, а именно человекообразные обезьяны, ходят на костяшках пальцев , по крайней мере, на передних ногах. Гигантские муравьеды [152] и утконосы [153] также ходят на костяшках пальцев. Некоторые млекопитающие являются двуногими и используют для передвижения только две конечности, что можно увидеть, например, у людей и человекообразных обезьян. Двуногие виды имеют большее поле зрения, чем четвероногие, сохраняют больше энергии и способны манипулировать объектами руками, что помогает в поиске пищи. Вместо того, чтобы идти, некоторые двуногие прыгают, например, кенгуру и кенгуру-крысы . [154] [155]
Животные будут использовать разные походки в зависимости от скорости, местности и ситуаций. Например, лошади демонстрируют четыре естественных аллюра: самый медленный аллюр — это шаг , затем есть три более быстрых аллюра, от самого медленного к самому быстрому: рысь , галоп и галоп . Животные также могут иметь необычную походку, которую иногда используют, например, для движения вбок или назад. Например, основными человеческими походками являются ходьба и бег на четырех ногах на двух ногах, но иногда они используют и многие другие походки, включая ползание в ограниченном пространстве. [156] Млекопитающие демонстрируют широкий диапазон походок , порядок расположения и подъема придатков при передвижении. Походки можно сгруппировать по категориям в соответствии с их последовательностью поддержки. Четвероногих животных можно разделить на три основные категории: ходьба, бег и прыжки . [157] Ходьба — наиболее распространенная походка, при которой некоторые ступни в любой момент времени стоят на земле, и встречается почти у всех четвероногих животных. Считается, что бег происходит, когда в некоторые моменты шага все ступни отрываются от земли в момент зависания. [156]
Древесный

Древесные животные часто имеют удлиненные конечности, которые помогают им преодолевать промежутки, достигать фруктов или других ресурсов, проверять твердость опоры впереди и, в некоторых случаях, раскачиваться ( качаться между деревьями). [158] Многие древесные виды, такие как древесные дикобразы, шелковистые муравьеды , паукообразные обезьяны и опоссумы , используют цепкие хвосты , чтобы хвататься за ветки. У обезьяны-паука кончик хвоста имеет либо оголенный участок, либо липкую подушечку, обеспечивающую повышенное трение. Когти можно использовать для взаимодействия с грубым субстратом и изменения направления сил, прилагаемых животным. Именно это позволяет белкам взбираться на стволы деревьев, которые настолько велики, что с точки зрения такого маленького животного они кажутся практически плоскими. Однако когти могут помешать животному хватать очень маленькие ветки, поскольку они могут слишком сильно обхватить и уколоть собственную лапу животного. Приматы используют фрикционный захват, полагаясь на безволосые кончики пальцев. Сжатие ветки между кончиками пальцев создает силу трения, которая удерживает руку животного на ветке. Однако этот тип захвата зависит от угла силы трения, то есть от диаметра ветки, причем более крупные ветви приводят к снижению способности захвата. Чтобы контролировать спуск, особенно по ветвям большого диаметра, у некоторых древесных животных, таких как белки, развились очень подвижные голеностопные суставы, которые позволяют поворачивать ступню в «перевернутое» положение. Это позволяет когтям цепляться за шероховатую поверхность коры, противодействуя силе тяжести. Небольшой размер дает много преимуществ древесным видам: например, увеличение относительного размера ветвей для животного, более низкий центр массы, повышенную устойчивость, меньшую массу (что позволяет передвигаться по меньшим ветвям) и способность перемещаться по более загроможденной среде обитания. [158] Размер, связанный с весом, влияет на планирующих животных, таких как сахарный планер . [159] Некоторые виды приматов, летучих мышей и все виды ленивцев достигают пассивной устойчивости, зависая под веткой. И подача, и опрокидывание становятся неуместными, поскольку единственным способом провала будет потеря хватки. [158]
Воздушная
Летучие мыши — единственные млекопитающие, которые по-настоящему умеют летать. Они летают по воздуху с постоянной скоростью, перемещая крылья вверх и вниз (обычно с некоторым движением вперед-назад). Поскольку животное находится в движении, относительно его тела существует некоторый поток воздуха, который в сочетании со скоростью крыльев создает более быстрый поток воздуха, движущийся по крылу. Это создает вектор подъемной силы, направленный вперед и вверх, и вектор силы сопротивления, направленный назад и вверх. Направленные вверх компоненты противодействуют гравитации, удерживая тело в воздухе, в то время как передний компонент обеспечивает тягу, противодействующую сопротивлению как крыла, так и тела в целом. [160]
Крылья летучих мышей намного тоньше и состоят из большего количества костей, чем у птиц, что позволяет летучим мышам более точно маневрировать и летать с большей подъемной силой и меньшим сопротивлением. [161] [162] Складывая крылья внутрь по направлению к телу при взмахе вверх, они тратят на 35% меньше энергии во время полета, чем птицы. [163] Мембраны нежные, легко рвутся; однако ткань мембраны летучей мыши способна отрастать заново, поэтому небольшие разрывы быстро заживают. [164] Поверхность их крыльев оснащена сенсорными рецепторами на небольших выпуклостях, называемых клетками Меркеля , которые также встречаются на кончиках пальцев человека. Эти чувствительные области у летучих мышей различны, так как каждая шишка имеет крошечный волосок в центре, что делает ее еще более чувствительной и позволяет летучей мыши обнаруживать и собирать информацию о воздухе, проходящем через ее крылья, а также летать более эффективно, изменяя форму. своих крыльев в ответ. [165]
Ископаемые и подземные


Ископаемое (от латинского fossor , что означает «копатель») — животное, приспособленное к рытью, которое живет в основном, но не исключительно, под землей. Некоторые примеры — барсуки и голые землекопы . Многие виды грызунов также считаются ископаемыми, поскольку они живут в норах большую часть дня, но не весь день. Виды, живущие исключительно под землей, являются подземными, а виды с ограниченной адаптацией к роющему образу жизни — субфоссорными. Некоторые организмы являются ископаемыми, чтобы помочь в регулировании температуры , в то время как другие используют подземную среду обитания для защиты от хищников или для хранения пищи . [166]
Ископаемые млекопитающие имеют веретенообразное тело, наиболее толстое в плечах и суживающееся к хвосту и носу. Неспособные видеть в темных норах, у большинства деградировали глаза, но дегенерация варьируется у разных видов; Карманные суслики , например, имеют только полуфоссорный вид и имеют очень маленькие, но функциональные глаза, у полностью окаменелого сумчатого крота глаза деградировали и бесполезны, тальповых кротов у рудиментарные глаза, а у плащового золотого крота глаза покрыты слоем кожи. . Наружные ушные раковины также очень малы или отсутствуют. У истинно окаменелых млекопитающих короткие и толстые ноги, поскольку для роющих млекопитающих сила важнее скорости, а у полуфоссорных млекопитающих ноги бегающие . Передние лапы широкие и имеют сильные когти, которые помогают разрыхлять землю при рытье нор, а на задних лапах есть перепонки и когти, которые помогают отбрасывать разрыхленную землю назад. У большинства из них большие резцы, чтобы грязь не попадала в рот. [167]
Многие ископаемые млекопитающие, такие как землеройки, ежи и кроты, были отнесены к ныне устаревшему отряду Insectivora . [168]
Водный
Полностью водные млекопитающие, китообразные и сирены , лишились ног и имеют хвостовой плавник, позволяющий передвигаться по воде. Движение ласт непрерывное. Киты плавают, перемещая хвостовой плавник и нижнюю часть тела вверх и вниз, двигаясь вертикально, а их ласты в основном используются для управления. Их скелетная анатомия позволяет им быть быстрыми пловцами. У большинства видов есть спинной плавник , который не позволяет им переворачиваться в воде. [169] [170] Лапы сирен поднимаются вверх и вниз длинными взмахами, чтобы переместить животное вперед, и могут поворачиваться для поворота. Передние конечности представляют собой ласты, похожие на весла, которые помогают поворачиваться и замедляться. [171]
Полуводные млекопитающие, такие как ластоногие, имеют две пары ласт спереди и сзади: передние и задние. Локти и лодыжки заключены внутри тела. [172] [173] Ластоногие имеют несколько приспособлений для уменьшения сопротивления . В дополнение к обтекаемому телу, у них есть гладкая сеть мышечных пучков на коже, которая может увеличивать ламинарный поток и облегчать им скольжение в воде. У них также отсутствуют выпрямляющие пили , поэтому их мех может быть обтекаемым во время плавания. [174] Для передвижения они полагаются на передние ласты, подобно крыльям, подобно пингвинам и морским черепахам . [175] Движение передних ласт не является непрерывным, и животное скользит между каждым гребком. [173] По сравнению с наземными хищниками, передние конечности уменьшены в длине, что дает локомоторным мышцам плечевых и локтевых суставов большее механическое преимущество; [172] задние ласты служат стабилизаторами. [174] Другие полуводные млекопитающие включают бобров, бегемотов , выдр и утконосов. [176] Бегемоты — очень крупные полуводные млекопитающие, их бочкообразные тела имеют гравитационно- портальную структуру скелета. [177] приспособлены выдерживать свой огромный вес, а их удельный вес позволяет им тонуть и передвигаться по дну реки. [178]
Поведение
Общение и вокализация

Многие млекопитающие общаются посредством вокала. Голосовое общение служит многим целям, в том числе в брачных ритуалах, в качестве предупреждающих сигналов , [180] для указания источников пищи и для социальных целей. Самцы часто кричат во время брачных ритуалов, чтобы отогнать других самцов и привлечь самок, например, львов и рев благородных оленей . [181] Песни ; горбатого кита могут быть сигналами для самок [182] в разных регионах океана у них разные диалекты. [183] Социальные вокализации включают территориальные крики гиббонов и использование частоты у больших остроносых летучих мышей для различения групп. [184] Верветка подает отчетливый сигнал тревоги для каждого из как минимум четырех различных хищников, а реакция других обезьян варьируется в зависимости от сигнала. Например, если сигнал тревоги сигнализирует питону, обезьяны забираются на деревья, тогда как сигнал тревоги орла заставляет обезьян искать укрытие на земле. [179] У луговых собачек также есть сложные крики, которые сигнализируют о типе, размере и скорости приближающегося хищника. [185] Слоны социально общаются с помощью различных звуков, включая фырканье, крик, трубный звук, рев и урчание. Некоторые из грохочущих криков являются инфразвуковыми , ниже диапазона слуха человека, и их могут услышать другие слоны на расстоянии до 6 миль (9,7 км) в неподвижное время перед восходом и закатом солнца. [186]
Млекопитающие подают сигналы различными способами. Многие подают визуальные сигналы против хищников , например, когда олени и газели останавливаются , честно указывая на свою физическую форму и способность убежать. [187] [188] или когда белохвостый олень и другие млекопитающие-жертвы при тревоге отмечают заметные отметки на хвосте, информируя хищника о том, что он был обнаружен. [189] Многие млекопитающие используют запаховую маркировку , иногда, возможно, для защиты территории, но, вероятно, с рядом функций как внутри вида, так и между видами. [190] [191] [192] Летучие мыши и зубатые киты, включая океанических дельфинов, издают звуки как социально, так и с помощью эхолокации . [193] [194] [195]
Кормление
Поддержание высокой постоянной температуры тела требует больших затрат энергии, поэтому млекопитающие нуждаются в питательном и обильном рационе. Хотя самые ранние млекопитающие, вероятно, были хищниками, с тех пор различные виды адаптировались к удовлетворению своих пищевых потребностей различными способами. Некоторые едят других животных — это плотоядная диета (включая насекомоядную). Другие млекопитающие, называемые травоядными , питаются растениями, которые содержат сложные углеводы, такие как целлюлоза. Травоядная диета включает такие подтипы, как зерноядные (поедание семян), фоливорные (поедание листьев), плодоядные (поедание фруктов), нектариворные (поедание нектара), мармеладные (поедание жевательной резинки) и микофагия (поедание грибов). Пищеварительный тракт травоядных животных является домом для бактерий, которые ферментируют эти сложные вещества и делают их доступными для пищеварения, которые находятся либо в многокамерном желудке , либо в большой слепой кишке. [95] Некоторые млекопитающие являются копрофагами и потребляют фекалии для поглощения питательных веществ, не переваренных при первом приеме пищи. [91] : 131–137 Всеядное животное питается как добычей, так и растениями. У плотоядных млекопитающих простой пищеварительный тракт , поскольку белки , липиды и минералы, содержащиеся в мясе, практически не требуют специального пищеварения. Исключением являются усатые киты также находится , у которых кишечная флора в многокамерном желудке, как и у наземных травоядных. [196]
Размер животного также является фактором, определяющим тип диеты ( правило Аллена ). Поскольку у мелких млекопитающих высокое соотношение площади теплоотдающей поверхности к теплогенерирующему объему, у них, как правило, высокие потребности в энергии и высокая скорость метаболизма . Млекопитающие весом менее 18 унций (510 г; 1,1 фунта) в основном насекомоядны, поскольку не переносят медленный и сложный процесс пищеварения травоядных. С другой стороны, более крупные животные выделяют больше тепла и теряют меньше этого тепла. Поэтому они могут переносить либо более медленный процесс сбора (хищники, питающиеся более крупными позвоночными), либо более медленный процесс пищеварения (травоядные). [197] Более того, млекопитающие весом более 18 унций (510 г; 1,1 фунта) обычно не могут собрать достаточно насекомых в часы бодрствования, чтобы поддерживать себя. Единственными крупными насекомоядными млекопитающими являются те, которые питаются огромными колониями насекомых ( муравьев или термитов ). [198]


Некоторые млекопитающие всеядны и демонстрируют различную степень плотоядности и травоядности, обычно склоняясь к одному больше, чем к другому. Поскольку растения и мясо перевариваются по-разному, одно предпочтение отдается другому, как у медведей, у которых некоторые виды могут быть в основном плотоядными, а другие - преимущественно травоядными. [200] Они сгруппированы в три категории: мезокарниворные (50–70% мяса), гиперплотоядные (70% и более мяса) и гипохищные (50% и менее мяса). Зубной ряд гипоплотоядных животных состоит из тупых треугольных хищных зубов, предназначенных для пережевывания пищи. Гиперплотоядные животные, однако, имеют конические зубы и острые хищники, предназначенные для рубящих ударов, а в некоторых случаях сильные челюсти для дробления костей, как в случае с гиенами , что позволяет им поедать кости; у некоторых вымерших групп, особенно Machairodontinae саблевидной формы , были клыки . [199]
Некоторые физиологические плотоядные животные потребляют растительные вещества, а некоторые физиологические травоядные животные потребляют мясо. С поведенческой точки зрения это сделало бы их всеядными, но с физиологической точки зрения это может быть связано с зоофармакогнозией . Физиологически, чтобы считаться всеядными, животные должны иметь возможность получать энергию и питательные вещества из растительных и животных материалов. Таким образом, таких животных по-прежнему можно классифицировать как плотоядных и травоядных, хотя они только получают питательные вещества из материалов, происходящих из источников, которые, казалось бы, не дополняют их классификацию. [201] Например, хорошо известно, что некоторые копытные, такие как жирафы, верблюды и крупный рогатый скот, грызут кости, чтобы потреблять определенные минералы и питательные вещества. [202] Кроме того, кошки, которых обычно считают облигатными хищниками, иногда едят траву, чтобы отрыгнуть неперевариваемый материал (например, комки шерсти ), помочь в выработке гемоглобина и в качестве слабительного. [203]
Многие млекопитающие при отсутствии достаточной потребности в пище в окружающей среде подавляют свой метаболизм и сохраняют энергию в процессе, известном как гибернация . [204] В период, предшествующий спячке, более крупные млекопитающие, такие как медведи, становятся полифагами , чтобы увеличить запасы жира, тогда как более мелкие млекопитающие предпочитают собирать и хранить пищу. [205] Замедление обмена веществ сопровождается учащением пульса и дыхания, а также падением внутренней температуры, которая в некоторых случаях может приближаться к температуре окружающей среды. Например, внутренняя температура зимующих арктических сусликов может упасть до -2,9 ° C (26,8 ° F); однако температура головы и шеи всегда остается выше 0 ° C (32 ° F). [206] Некоторые млекопитающие в жарких регионах впадают в спячку во время засухи или сильной жары, например, толстохвостый карликовый лемур ( Chirogaleus medius ). [207]
Питьевой

По необходимости наземные животные в неволе привыкают к питьевой воде, но большинство свободно перемещающихся животных сохраняют гидратацию за счет жидкости и влаги в свежей пище. [208] и научитесь активно искать продукты с высоким содержанием жидкости. [209] Когда условия вынуждают их пить из водоемов, методы и движения у разных видов сильно различаются. [210]
Кошки , собаки и жвачные животные опускают шею и плещутся в воде своими мощными языками. [210] Кошки и собаки лакают воду языком, напоминающим ложку. [211] Собаки лакают воду, зачерпывая ее в рот языком, имеющим форму ковша. Однако у кошек воды касается только кончик языка (гладкий), после чего кошка быстро втягивает язык обратно в рот, который вскоре закрывается; в результате в рот кошки втягивается столб жидкости, который затем фиксируется закрытием рта. [212] Жвачные и большинство других травоядных животных частично погружают кончик рта в воду, чтобы втянуть воду посредством ныряния с выпрямленным языком. [213] Кошки пьют значительно медленнее, чем жвачные животные, которые сталкиваются с большей опасностью естественных хищников. [210]
Многие животные пустыни не пьют, даже если есть вода, а питаются сочными растениями . [210] В холодных и морозных условиях некоторые животные, такие как зайцы , древесные белки и снежные бараны , прибегают к поеданию снега и сосулек. [214] В саваннах способ питья жирафов был источником спекуляций из-за его явного игнорирования гравитации; Самая последняя теория предполагает, что длинная шея животного функционирует как плунжерный насос . [215] Уникально то, что слоны набирают воду в хобот и впрыскивают ее в рот. [210]Интеллект
У разумных млекопитающих, таких как приматы , головной мозг крупнее остального мозга. Интеллект сам по себе нелегко определить, но признаки интеллекта включают способность к обучению в сочетании с поведенческой гибкостью. Крысы , например, считаются очень умными, поскольку они могут учиться и выполнять новые задачи, и эта способность может быть важна, когда они впервые колонизируют новую среду обитания . У некоторых млекопитающих сбор пищи, по-видимому, связан с интеллектом: у оленя, питающегося растениями, мозг меньше, чем у кошки, и ей приходится думать, чтобы перехитрить свою добычу. [198]

Использование инструментов животными может указывать на разные уровни обучения и познания . Морская выдра использует камни как неотъемлемую и регулярную часть своего добывающего поведения (разбивая морское ушко о камни или разбивая раковины), при этом некоторые популяции тратят 21% своего времени на изготовление инструментов. [216] Другое использование инструментов, например, когда шимпанзе используют ветки для «ловли» термитов, может быть развито путем наблюдения за тем, как другие используют инструменты , и может даже стать настоящим примером обучения животных. [217] Инструменты можно даже использовать для решения головоломок, в которых животное, кажется, переживает «момент Эврики» . [218] Другие млекопитающие, которые не используют инструменты, например собаки, также могут пережить момент Эврики. [219]
Размер мозга ранее считался основным показателем интеллекта животного. Поскольку большая часть мозга используется для поддержания функций организма, большее соотношение массы мозга к массе тела может увеличить количество массы мозга, доступной для более сложных когнитивных задач. Аллометрический анализ показывает, что размер мозга млекопитающих масштабируется примерно так же, как и у млекопитающих. 2 ⁄ 3 или 3/4 показателя . массы тела Сравнение размера мозга конкретного животного с ожидаемым размером мозга на основе такого аллометрического анализа дает коэффициент энцефализации , который можно использовать как еще один показатель интеллекта животного. [220] Кашалоты имеют самую большую массу мозга среди всех животных на земле: в среднем 8000 кубических сантиметров (490 куб. Дюймов) и 7,8 кг (17 фунтов) у половозрелых самцов. [221]
Самосознание представляется признаком абстрактного мышления. Самосознание, хотя и не имеет четкого определения, считается предшественником более продвинутых процессов, таких как метакогнитивное мышление . Традиционным методом измерения этого показателя является зеркальный тест , который определяет, обладает ли животное способностью самопознания. [222] К млекопитающим, прошедшим зеркальный тест, относятся азиатские слоны (некоторые проходят, некоторые нет); [223] шимпанзе; [224] бонобо ; [225] орангутанги ; [226] человек, от 18 месяцев ( стадия зеркала ); [227] обыкновенные афалины ; [а] [228] косатки ; [229] и ложные косатки . [229]
Социальная структура

Эусоциальность – это высший уровень социальной организации. В этих обществах наблюдается перекрытие взрослых поколений, разделение репродуктивного труда и совместная забота о молодёжи. Обычно насекомые, такие как пчелы , муравьи и термиты, обладают эусоциальным поведением, но оно продемонстрировано у двух видов грызунов: голого землекопа. [230] и дамаралендский землекоп . [231]
Пресоциальность — это когда животные демонстрируют больше, чем просто сексуальные взаимодействия с представителями того же вида, но не соответствуют критериям эусоциальности. То есть досоциальные животные могут демонстрировать коллективный образ жизни, совместную заботу о молодняке или примитивное разделение репродуктивного труда, но они не проявляют всех трех основных черт эусоциальных животных. Человек и некоторые виды Callitrichidae ( игрунки и тамарины ) уникальны среди приматов по степени совместной заботы о потомстве. [232] Гарри Харлоу поставил эксперимент с макаками-резусами , досоциальными приматами, в 1958 году; Результаты этого исследования показали, что социальные контакты необходимы для развития молодых обезьян как умственно, так и сексуально. [233]
Общество разделения-слияния — это общество, которое часто меняет свой размер и состав, образуя постоянную социальную группу, называемую «родительской группой». Постоянные социальные сети состоят из всех отдельных членов сообщества и часто меняются, чтобы отслеживать изменения в их среде. В обществе разделения-слияния основная родительская группа может распадаться (расщепляться) на более мелкие стабильные подгруппы или отдельных лиц, чтобы адаптироваться к экологическим или социальным обстоятельствам. Например, несколько самцов могут отделиться от основной группы, чтобы охотиться или добывать пищу в течение дня, но ночью они могут вернуться, чтобы присоединиться (слиться) с основной группой, чтобы поделиться едой и принять участие в других занятиях. Это наблюдается у многих млекопитающих, например у приматов (например, орангутанов и паукообразных обезьян ). [234] слоны, [235] пятнистые гиены , [236] львы, [237] и дельфины. [238]
Одиночные животные защищают территорию и избегают социальных взаимодействий с представителями своего вида, за исключением периода размножения. Это сделано для того, чтобы избежать конкуренции за ресурсы, поскольку две особи одного и того же вида будут занимать одну и ту же нишу, а также для предотвращения истощения запасов пищи. [239] Одиночное животное во время кормления также может быть менее заметным для хищников или добычи. [240]

В иерархии люди либо доминируют, либо подчиняются. Деспотическая иерархия – это когда один человек доминирует, а остальные подчиняются, как у волков и лемуров. [241] иерархия — это линейный ранжирование людей, в котором есть человек высшего и низшего уровня. Иерархия также может быть ранжирована по полу: самая низкая особь одного пола имеет более высокий рейтинг, чем высшая особь другого пола, как у гиен. [242] Доминирующие особи, или альфы, имеют высокие шансы на репродуктивный успех, особенно в гаремах , где один или несколько самцов (постоянных самцов) имеют исключительные права на размножение самок в группе. [243] Иногородних самцов также можно принимать в гаремы, но некоторые виды, например обыкновенная летучая мышь-вампир ( Desmodus rotundus ), могут быть более строгими. [244]
Некоторые млекопитающие совершенно моногамны , то есть они спариваются на всю жизнь и не принимают других партнеров (даже после смерти первоначального партнера), как, например, волки, евразийские бобры и выдры. [245] [246] Существует три типа полигамии: один или несколько доминирующих самцов имеют право на размножение ( полигиния ), несколько самцов, с которыми спариваются самки (полиандрия), или несколько самцов имеют исключительные отношения с несколькими самками ( полигинандрия ). Гораздо чаще случается полигинное спаривание, которое, за исключением токов , по оценкам, встречается у 90% млекопитающих. [247] Спаривание леков происходит, когда самцы собираются вокруг самок и пытаются привлечь их различными проявлениями ухаживания и вокалами, как у обыкновенных тюленей. [248]
У всех высших млекопитающих (за исключением однопроходных) есть два основных приспособления для ухода за детенышами: живорождение и лактация. Они подразумевают общегрупповой выбор степени родительской заботы . Они могут строить гнезда и рыть норы, чтобы выращивать в них птенцов, или часто кормить и охранять их в течение длительного периода времени. Многие млекопитающие прошли K-отбор и вкладывают больше времени и энергии в развитие своего потомства, чем отобранные r животные, . Когда два животных спариваются, они оба заинтересованы в успехе потомства, хотя часто и в разных крайностях. Самки млекопитающих проявляют некоторую степень материнской агрессии, что является еще одним примером родительской заботы, которая может быть направлена против других самок этого вида или детенышей других самок; однако некоторые млекопитающие могут «теть» детенышей других самок и заботиться о них. Самцы млекопитающих могут играть роль в воспитании детей, как и в случае с тенреками, однако это варьируется от вида к виду, даже в пределах одного рода. Например, самцы южной косичьей макаки ( Macaca nemestrina ) не участвуют в уходе за детьми, тогда как самцы японской макаки ( M. fuscata ) участвуют. [249]
Люди и другие млекопитающие
В человеческой культуре

Нечеловеческие млекопитающие играют самые разнообразные роли в человеческой культуре. Они являются самыми популярными домашними животными : десятки миллионов собак, кошек и других животных, включая кроликов и мышей, содержатся семьями по всему миру. [250] [251] [252] Млекопитающие, такие как мамонты , лошади и олени, являются одними из самых ранних предметов искусства, их можно найти в верхнего палеолита, наскальных рисунках таких как в Ласко . [253] Крупные художники, такие как Альбрехт Дюрер , Джордж Стаббс и Эдвин Ландсир, известны своими портретами млекопитающих. [254] На многие виды млекопитающих охотились ради развлечения и ради еды; Олени и кабаны особенно популярны в качестве охотничьих животных . [255] [256] [257] Млекопитающие, такие как лошади и собаки, широко участвуют в спортивных гонках, часто в сочетании со ставками на результат . [258] [259] Существует противоречие между ролью животных как товарищей по отношению к человеку и их существованием как личностей с собственными правами . [260] Млекопитающие также играют самые разнообразные роли в литературе. [261] [262] [263] фильм, [264] мифология и религия. [265] [266] [267]
Использование и важность

Одомашнивание млекопитающих сыграло важную роль в развитии сельского хозяйства и цивилизации в эпоху неолита , заставив фермеров заменить охотников-собирателей по всему миру. [б] [269] Этот переход от охоты и собирательства к выпасу стад и выращиванию сельскохозяйственных культур стал важным шагом в истории человечества. Новая сельскохозяйственная экономика, основанная на одомашненных млекопитающих, вызвала «радикальную реструктуризацию человеческого общества, глобальные изменения в биоразнообразии и значительные изменения в формах рельефа Земли и ее атмосфере... важные результаты». [270]
Домашние млекопитающие составляют большую часть поголовья скота , выращиваемого на мясо во всем мире. В их число входят (2009 г.) около 1,4 миллиарда крупного рогатого скота , 1 миллиард овец , 1 миллиард домашних свиней , [271] [272] и (1985) более 700 миллионов кроликов. [273] Рабочие домашние животные, включая крупный рогатый скот и лошадей, использовались для работы и транспорта с самого начала сельского хозяйства, их численность сокращается с появлением механизированного транспорта и сельскохозяйственных машин . В 2004 году они по-прежнему обеспечивали около 80% электроэнергии преимущественно мелких ферм в странах третьего мира и около 20% мирового транспорта, опять же в основном в сельских районах. В горных районах, непригодных для колесного транспорта, вьючные животные . грузы продолжают перевозить [274] Из шкур млекопитающих делают кожу для обуви , одежды и обивки . Шерсть млекопитающих, включая овец, коз и альпак, веками использовалась для изготовления одежды. [275] [276]
Млекопитающие играют важную роль в науке в качестве экспериментальных животных , как в фундаментальных биологических исследованиях, таких как генетика, так и в области генетики. [278] и в разработке новых лекарств, которые должны быть тщательно протестированы, чтобы продемонстрировать их безопасность . [279] используются миллионы млекопитающих, особенно мышей и крыс. в экспериментах Ежегодно [280] — Нокаутная мышь это генетически модифицированная мышь с инактивированным геном , замененным или разрушенным искусственным участком ДНК. Они позволяют изучать секвенированные гены, функции которых неизвестны. [281] Небольшой процент млекопитающих - это приматы, не относящиеся к человеку, которые используются в исследованиях из-за их сходства с людьми. [282] [283] [284]
Несмотря на пользу, которую одомашненные млекопитающие принесли для развития человечества, люди оказывают все более пагубное воздействие на диких млекопитающих во всем мире. Подсчитано, что масса всех диких млекопитающих сократилась до 4% от всех млекопитающих, при этом 96% млекопитающих сейчас составляют люди и их домашний скот (см. рисунок). Фактически наземные дикие млекопитающие составляют лишь 2% всех млекопитающих. [285] [286]
Гибриды


Гибриды — это потомство, полученное в результате скрещивания двух генетически различных особей, что обычно приводит к высокой степени гетерозиготности, хотя гибрид и гетерозигота не являются синонимами. Преднамеренная или случайная гибридизация двух или более видов близкородственных животных посредством разведения в неволе — это человеческая деятельность, которая существует на протяжении тысячелетий и развивается в экономических целях. [287] Гибриды между различными подвидами внутри вида (например, между бенгальским тигром и сибирским тигром ) известны как внутривидовые гибриды. Гибриды между разными видами внутри одного рода (например, между львами и тиграми) известны как межвидовые гибриды или скрещивания. Гибриды между разными родами (например, между овцами и козами) известны как межродовые гибриды. [288] Естественные гибриды будут возникать в гибридных зонах , где две популяции видов одного и того же рода или виды, живущие в одних и тех же или соседних областях, будут скрещиваться друг с другом. Некоторые гибриды были признаны видами, например красный волк (хотя это спорно). [289]
Искусственный отбор , преднамеренное селекционное разведение домашних животных, используется для восстановления недавно вымерших животных в попытке создать породу животных с фенотипом , напоминающим вымершего предка дикого типа . Размножающийся (внутривидовой) гибрид может быть очень похож на вымерший дикий тип по внешнему виду, экологической нише и в некоторой степени генетике, но исходный генофонд этого дикого типа навсегда утрачен с его исчезновением . В результате выведенные породы являются в лучшем случае расплывчатыми двойниками вымерших диких типов, так же как крупный рогатый скот Хека имеет сходство с зубром . [290]
Чистокровные дикие виды, эволюционировавшие в соответствии с определенной экологией, могут оказаться под угрозой исчезновения [291] посредством процесса генетического загрязнения , неконтролируемой гибридизации, интрогрессии генетического затопления, что приводит к гомогенизации или вытеснению гибридных видов из гетерозисных конкуренции . [292] Когда новые популяции завозятся или селективно разводятся людьми, или когда изменение среды обитания приводит к контакту ранее изолированных видов, возможно исчезновение некоторых видов, особенно редких разновидностей. [293] Скрещивание может поглотить более редкий генофонд и создать гибриды, истощая чистокровный генофонд. Например, находящийся под угрозой исчезновения дикий водяной буйвол находится под наибольшей угрозой исчезновения из-за генетического загрязнения домашним водяным буйволом . Такие вымирания не всегда очевидны с морфологической точки зрения. Некоторая степень потока генов является нормальным эволюционным процессом, тем не менее, гибридизация ставит под угрозу существование редких видов. [294] [295]
Угрозы

Утрата видов из экологических сообществ, дефаунация , в первую очередь обусловлена деятельностью человека. [296] Это привело к опустошению лесов , экологическим сообществам, лишенным крупных позвоночных. [297] [298] Во время четвертичного вымирания массовое вымирание представителей мегафауны совпало с появлением людей, что свидетельствует о человеческом влиянии. Одна из гипотез заключается в том, что люди охотились на крупных млекопитающих, таких как шерстистый мамонт , и привели к их исчезновению. [299] [300] 2019 год о глобальной оценке биоразнообразия и экосистемных услуг за В докладе IPBES говорится, что общая биомасса диких млекопитающих сократилась на 82 процента с момента зарождения человеческой цивилизации. [301] [302] Дикие животные составляют всего 4% биомассы млекопитающих на Земле, тогда как люди и их одомашненные животные составляют 96%. [286]
По прогнозам, в ближайшем будущем различные виды вымрут . [303] среди них носорог , [304] жирафы , [305] и виды приматов [306] и панголины . [307] Согласно отчету WWF « Живая планета» за 2020 год , популяции позвоночных диких животных сократились на 68% с 1970 года в результате деятельности человека, особенно чрезмерного потребления , роста населения и интенсивного земледелия , что является свидетельством того, что люди спровоцировали шестое массовое вымирание . [308] [309] Одна только охота угрожает сотням видов млекопитающих по всему миру. [310] [311] Ученые утверждают, что растущий спрос на мясо способствует утрате биоразнообразия , поскольку это является важным фактором вырубки лесов и разрушения среды обитания ; богатые видами места обитания, такие как значительная часть тропических лесов Амазонки , превращаются в сельскохозяйственные угодья для производства мяса. [312] [313] [314] Еще одним фактором влияния является чрезмерная охота и браконьерство , которые могут сократить общую популяцию охотничьих животных. [315] особенно те, что расположены вблизи деревень, [316] как в случае с пекари . [317] Последствия браконьерства особенно заметны в торговле слоновой костью африканских слонов. [318] , подвергаются риску запутывания в рыболовных снастях, Морские млекопитающие, особенно китообразные при этом смертность от выброса колеблется от 65 000 до 86 000 особей в год. [319]
Внимание уделяется исчезающим видам во всем мире, в частности, через Конвенцию о биологическом разнообразии , также известную как Соглашение Рио-де-Жанейро, в которую входят 189 подписавших стран, которые сосредоточены на выявлении исчезающих видов и мест обитания. [320] Еще одна известная природоохранная организация — МСОП, членами которого являются более 1200 правительственных и неправительственных организаций. [321]
Недавние вымирания можно напрямую объяснить влиянием человека. [322] [296] МСОП характеризует «недавнее» вымирание как вымирание, произошедшее после отсечки в 1500 лет. [323] и около 80 видов млекопитающих вымерли с тех пор и в 2015 году. [324] Некоторые виды, такие как олень Пер-Давида. [325] вымерли в дикой природе и выживают исключительно в популяциях, содержащихся в неволе. Другие виды, такие как флоридская пантера , экологически вымерли , выживая в таких небольших количествах, что практически не оказывают никакого влияния на экосистему. [326] : 318 Другие популяции вымерли лишь локально (искоренены), все еще существуют в других местах, но их распространение сократилось. [326] : 75–77 как и с исчезновением серых китов в Атлантике . [327]
См. также
- Список родов млекопитающих - современные млекопитающие
- Список маммологов
- Список однопроходных и сумчатых
- Список плацентарных млекопитающих
- Список доисторических млекопитающих
- Список млекопитающих, находящихся под угрозой исчезновения
- Списки млекопитающих по численности популяции
- Списки млекопитающих по регионам
- Млекопитающие, описанные в 2000-х годах.
- Млекопитающие в культуре
- Мелкое млекопитающее
Примечания
- ^ Уменьшение задержки подхода к зеркалу, повторяющиеся кружения головой и пристальное рассмотрение отмеченных областей считались признаками самоузнания, поскольку у них нет рук и они не могут коснуться отмеченных областей. [228]
- ↑ Даймонд подробно обсудил этот вопрос в своей книге 1997 года « Ружья, микробы и сталь» . [268]
Ссылки
- ^ Льюис, Чарльтон Т.; Короткий, Чарльз (1879). «мама» . Латинский словарь . Цифровая библиотека Персея. Архивировано из оригинала 29 сентября 2022 года . Проверено 29 сентября 2022 г.
- ^ Воган Т.А., Райан Дж.М., Чаплевски, Нью-Джерси (2013). «Классификация млекопитающих». Маммология (6-е изд.). Джонс и Бартлетт Обучение. ISBN 978-1-284-03209-3 .
- ^ Симпсон Г.Г. (1945). «Принципы классификации и классификация млекопитающих». Американский музей естественной истории . 85 .
- ^ Jump up to: Перейти обратно: а б Салай Ф.С. (1999). «Классификация млекопитающих выше видового уровня: Обзор». Журнал палеонтологии позвоночных . 19 (1): 191–195. дои : 10.1080/02724634.1999.10011133 . JSTOR 4523980 .
- ^ Jump up to: Перейти обратно: а б Уилсон Д.Е. , Ридер Д.М., ред. (2005). «Предисловие и вводный материал» . Виды млекопитающих мира: таксономический и географический справочник (3-е изд.). Издательство Университета Джонса Хопкинса. п. XXVI. ISBN 978-0-8018-8221-0 . OCLC 62265494 .
- ^ «Млекопитающие» . Красный список исчезающих видов МСОП . Международный союз охраны природы (МСОП). Апрель 2010 г. Архивировано из оригинала 3 сентября 2016 г. Проверено 23 августа 2016 г. .
- ^ Бургин С.Дж., Колелла Дж.П., Кан П.Л., Апхэм Н.С. (1 февраля 2018 г.). «Сколько существует видов млекопитающих?» . Журнал маммологии . 99 (1): 1–14. дои : 10.1093/jmammal/gyx147 .
- ^ Роу Т. (1988). «Определение, диагностика и происхождение млекопитающих» (PDF) . Журнал палеонтологии позвоночных . 8 (3): 241–264. Бибкод : 1988JVPal...8..241R . дои : 10.1080/02724634.1988.10011708 . Архивировано (PDF) из оригинала 18 января 2024 года . Проверено 25 января 2024 г.
- ^ Лайель С. (1871 г.). Студенческие элементы геологии . Лондон: Джон Мюррей. п. 347. ИСБН 978-1-345-18248-4 .
- ^ Чифелли Р.Л., Дэвис Б.М. (декабрь 2003 г.). «Палеонтология. Происхождение сумчатых». Наука . 302 (5652): 1899–1900. дои : 10.1126/science.1092272 . ПМИД 14671280 . S2CID 83973542 .
- ^ Кемп Т.С. (2005). Происхождение и эволюция млекопитающих (PDF) . Соединенное Королевство: Издательство Оксфордского университета. п. 3. ISBN 978-0-19-850760-4 . OCLC 232311794 . Архивировано (PDF) из оригинала 26 сентября 2023 года . Проверено 25 января 2024 г.
- ^ Датта ПМ (2005). «Самое раннее млекопитающее с поперечно расширенными верхними коренными зубами из формации Тики позднего триаса (карния), бассейн Южной Ревы Гондваны, Индия». Журнал палеонтологии позвоночных . 25 (1): 200–207. doi : 10.1671/0272-4634(2005)025[0200:EMWTEU]2.0.CO;2 . S2CID 131236175 .
- ^ Луо ZX, Мартин Т (2007). «Анализ молярной структуры и филогении родов докодонтов» (PDF) . Бюллетень Музея естественной истории Карнеги . 39 : 27–47. doi : 10.2992/0145-9058(2007)39[27:AOMSAP]2.0.CO;2 . S2CID 29846648 . Архивировано из оригинала (PDF) 3 марта 2016 года . Проверено 8 апреля 2013 г.
- ^ Аверьянов Александр О.; Войта, Леонид Л. (март 2024 г.). «Предполагаемое триасовое стволовое млекопитающее Tikitherium copyi — неогеновая землеройка» . Журнал эволюции млекопитающих . 31 (1). дои : 10.1007/s10914-024-09703-w . ISSN 1064-7554 .
- ^ Маккенна MC, Bell SG (1997). Классификация млекопитающих выше видового уровня . Нью-Йорк: Издательство Колумбийского университета. ISBN 978-0-231-11013-6 . OCLC 37345734 .
- ^ Нильссон М.А., Чураков Г., Зоммер М., Тран Н.В., Земанн А., Брозиус Дж., Шмитц Дж. (июль 2010 г.). «Отслеживание эволюции сумчатых с использованием архаичных геномных вставок ретропозонов» . ПЛОС Биология . 8 (7): e1000436. дои : 10.1371/journal.pbio.1000436 . ПМК 2910653 . ПМИД 20668664 .
- ^ Кригс Дж.О., Чураков Г., Кифманн М., Джордан У., Брозиус Дж., Шмитц Дж. (апрель 2006 г.). «Ретропозированные элементы как архивы эволюционной истории плацентарных млекопитающих» . ПЛОС Биология . 4 (4): е91. doi : 10.1371/journal.pbio.0040091 . ПМЦ 1395351 . ПМИД 16515367 .
- ^ Jump up to: Перейти обратно: а б Нишихара Х., Маруяма С., Окада Н. (март 2009 г.). «Анализ ретропозонов и недавние геологические данные предполагают почти одновременное расхождение трех надотрядов млекопитающих» . Труды Национальной академии наук Соединенных Штатов Америки . 106 (13): 5235–5240. Бибкод : 2009PNAS..106.5235N . дои : 10.1073/pnas.0809297106 . ПМЦ 2655268 . ПМИД 19286970 .
- ^ Спрингер М.С., Мерфи В.Дж., Эйзирик Э., О'Брайен С.Дж. (февраль 2003 г.). «Диверсификация плацентарных млекопитающих и граница мелового и третичного периодов» . Труды Национальной академии наук Соединенных Штатов Америки . 100 (3): 1056–1061. Бибкод : 2003PNAS..100.1056S . дои : 10.1073/pnas.0334222100 . ПМК 298725 . ПМИД 12552136 .
- ^ Тарвер Дж.Э., Дос Рейс М., Мирараб С., Моран Р.Дж., Паркер С., О'Рейли Дж.Э. и др. (январь 2016 г.). «Взаимоотношения плацентарных млекопитающих и пределы филогенетических выводов» . Геномная биология и эволюция . 8 (2): 330–344. дои : 10.1093/gbe/evv261 . hdl : 1983/64d6e437-3320-480d-a16c-2e5b2e6b61d4 . ПМЦ 4779606 . ПМИД 26733575 .
- ^ Альварес-Карретеро С., Тамури А.У., Баттини М., Насименто Ф.Ф., Карлайл Э., Ашер Р.Дж., Ян З., Донохью ПК и др. (2022). «Хронология эволюции млекопитающих на уровне вида, объединяющая филогеномные данные» . Природа . 602 (7896): 263–267. Бибкод : 2022Natur.602..263A . дои : 10.1038/s41586-021-04341-1 . hdl : 1983/de841853-d57b-40d9-876f-9bfcf7253f12 . ПМИД 34937052 . S2CID 245438816 .
- ^ Альварес-Карретеро, Сандра; Тамури, Асиф; Баттини, Маттео; Насименто, Фабрисия Ф.; Карлайл, Эмили; Ашер, Роберт; Ян, Цзыхэн; Донохью, Филип; дос Рейс, Марио (2021). «Данные для временной шкалы эволюции млекопитающих на уровне видов с интеграцией филогеномных данных» . Фиговая доля . doi : 10.6084/m9.figshare.14885691.v1 . Архивировано из оригинала 16 декабря 2023 года . Проверено 11 ноября 2023 г.
- ^ Мэн Дж, Ван Ю, Ли С (апрель 2011 г.). «Переходное среднее ухо млекопитающих от нового эвтриконодонта Джехол мелового периода». Природа . 472 (7342): 181–185. Бибкод : 2011Natur.472..181M . дои : 10.1038/nature09921 . ПМИД 21490668 . S2CID 4428972 .
- ^ Альберг П.Е., Милнер А.Р. (апрель 1994 г.). «Происхождение и ранняя диверсификация четвероногих». Природа . 368 (6471): 507–514. Бибкод : 1994Natur.368..507A . дои : 10.1038/368507a0 . S2CID 4369342 .
- ^ «Амниота-Палеос» . Архивировано из оригинала 20 декабря 2010 года.
- ^ «Обзор Synapsida – Палеос» . Архивировано из оригинала 20 декабря 2010 года.
- ^ Jump up to: Перейти обратно: а б с Кемп Т.С. (июль 2006 г.). «Происхождение и раннее распространение терапсидных млекопитающих рептилий: палеобиологическая гипотеза» (PDF) . Журнал эволюционной биологии . 19 (4): 1231–1247. дои : 10.1111/j.1420-9101.2005.01076.x . ПМИД 16780524 . S2CID 3184629 . Архивировано из оригинала (PDF) 8 марта 2021 года . Проверено 14 января 2012 г.
- ^ Jump up to: Перейти обратно: а б Беннетт А.Ф., Рубен Дж.А. (1986). «Метаболический и терморегуляторный статус терапсидов». В Hotton III N, MacLean JJ, Roth J, Roth EC (ред.). Экология и биология млекопитающих рептилий . Вашингтон, округ Колумбия: Издательство Смитсоновского института. стр. 207–218. ISBN 978-0-87474-524-5 .
- ^ Кермак Д.М., Кермак К.А. (1984). Эволюция характеров млекопитающих . Вашингтон, округ Колумбия: Крум Хелм. ISBN 978-0-7099-1534-8 . ОСЛК 10710687 .
- ^ Араужо; и др. (28 июля 2022 г.). «Биомеханика внутреннего уха показывает позднетриасовое происхождение эндотермии млекопитающих». Природа . 607 (7920): 726–731. Бибкод : 2022Natur.607..726A . дои : 10.1038/s41586-022-04963-z . ПМИД 35859179 . S2CID 236245230 .
- ^ Таннер Л.Х., Лукас С.Г., Чепмен М.Г. (2004). «Оценка данных и причин вымираний в позднем триасе» (PDF) . Обзоры наук о Земле . 65 (1–2): 103–139. Бибкод : 2004ESRv...65..103T . дои : 10.1016/S0012-8252(03)00082-5 . Архивировано из оригинала (PDF) 25 октября 2007 года.
- ^ Брусатте С.Л., Бентон М.Дж., Рута М., Ллойд Г.Т. (сентябрь 2008 г.). «Превосходство, конкуренция и оппортунизм в эволюционном развитии динозавров» (PDF) . Наука . 321 (5895): 1485–1488. Бибкод : 2008Sci...321.1485B . дои : 10.1126/science.1161833 . hdl : 20.500.11820/00556baf-6575-44d9-af39-bdd0b072ad2b . ПМИД 18787166 . S2CID 13393888 . Архивировано (PDF) из оригинала 19 июля 2018 года . Проверено 12 октября 2019 г.
- ^ Готье Ж.А. (1986). «Монофилия ящеров и происхождение птиц». В Падиане К. (ред.). Происхождение птиц и эволюция полета. Мемуары Калифорнийской академии наук . Том. 8. Сан-Франциско: Калифорнийская академия наук. стр. 1–55.
- ^ Серено ПК (1991). «Базальные архозавры: филогенетические связи и функциональные последствия». Мемуары Общества палеонтологии позвоночных . 2 :1–53. дои : 10.2307/3889336 . JSTOR 3889336 .
- ^ МакЛауд Н., Роусон П.Ф., Фори П.Л., Баннер Ф.Т., Будагер-Фадель М.К., Баун П.Р. и др. (1997). «Мел-третичный биотический переход». Журнал Геологического общества . 154 (2): 265–292. Бибкод : 1997JGSoc.154..265M . дои : 10.1144/gsjgs.154.2.0265 . S2CID 129654916 .
- ^ Хант Д.М., Ханкинс М.В., Коллин С.П., Маршалл, Нью-Джерси (2014). Эволюция визуальных и невизуальных пигментов . Лондон: Спрингер. п. 73. ИСБН 978-1-4614-4354-4 . OCLC 892735337 .
- ^ Бакалар Н. (2006). «Найден юрский бобр; переписывает историю млекопитающих» . Национальные географические новости . Архивировано из оригинала 3 марта 2006 года . Проверено 28 мая 2016 г.
- ^ Холл М.И., Камилар Дж.М., Кирк Э.К. (декабрь 2012 г.). «Форма глаз и ночное узкое место млекопитающих» . Труды Королевского общества B: Биологические науки . 279 (1749): 4962–4968. дои : 10.1098/rspb.2012.2258 . ПМЦ 3497252 . ПМИД 23097513 .
- ^ Луо ZX (декабрь 2007 г.). «Трансформация и диверсификация ранней эволюции млекопитающих». Природа . 450 (7172): 1011–1019. Бибкод : 2007Natur.450.1011L . дои : 10.1038/nature06277 . ПМИД 18075580 . S2CID 4317817 .
- ^ Пикрелл Дж (2003). «Самые древние окаменелости сумчатых, найденные в Китае» . Национальные географические новости. Архивировано из оригинала 17 декабря 2003 года . Проверено 28 мая 2016 г.
- ^ Jump up to: Перейти обратно: а б Луо ZX, Юань CX, Мэн QJ, Цзи Q (август 2011 г.). «Юрское плацентарное млекопитающее и расхождение сумчатых и плацентарных». Природа . 476 (7361): 442–5. Бибкод : 2011Natur.476..442L . дои : 10.1038/nature10291 . ПМИД 21866158 . S2CID 205225806 .
- ^ Цзи Q, Луо ZX, Юань CX, Вибл-младший, Чжан Дж. П., Георгий Дж. А. (апрель 2002 г.). «Самое раннее известное эвтерийное млекопитающее». Природа . 416 (6883): 816–822. Бибкод : 2002Natur.416..816J . дои : 10.1038/416816a . ПМИД 11976675 . S2CID 4330626 .
- ^ Jump up to: Перейти обратно: а б Новачек М.Дж., Ружье Г.В., Уибл-младший, Маккенна М.К., Дашевег Д., Горовиц I (октябрь 1997 г.). «Надлобковые кости плацентарных млекопитающих позднего мела Монголии». Природа . 389 (6650): 483–486. Бибкод : 1997Natur.389..483N . дои : 10.1038/39020 . ПМИД 9333234 . S2CID 205026882 .
- ^ Power ML, Schulkin J (2012). «Эволюция живорождения у млекопитающих» . Эволюция плаценты человека . Балтимор: Издательство Университета Джонса Хопкинса. п. 68. ИСБН 978-1-4214-0643-5 .
- ^ Роу Т., Рич Т.Х., Викерс-Рич П., Спрингер М., Вудберн М.О. (январь 2008 г.). «Самый старый утконос и его влияние на время расхождения клад утконоса и ехидны» . Труды Национальной академии наук Соединенных Штатов Америки . 105 (4): 1238–1242. Бибкод : 2008PNAS..105.1238R . дои : 10.1073/pnas.0706385105 . ПМК 2234122 . ПМИД 18216270 .
- ^ Грант Т. (1995). «Репродукция» . Утконос: уникальное млекопитающее . Сидней: Университет Нового Южного Уэльса. п. 55. ИСБН 978-0-86840-143-0 . OCLC 33842474 .
- ^ Goldman AS (июнь 2012 г.). «Эволюция иммунных функций молочной железы и защита ребенка». Медицина грудного вскармливания . 7 (3): 132–142. дои : 10.1089/bfm.2012.0025 . ПМИД 22577734 .
- ^ Jump up to: Перейти обратно: а б Роуз К.Д. (2006). Начало эпохи млекопитающих . Балтимор: Издательство Университета Джонса Хопкинса. стр. 82–83. ISBN 978-0-8018-8472-6 . OCLC 646769601 .
- ^ Бринк А.С. (1955). «Исследование скелета Диадемодона ». Палеонтология Африканская . 3 :3–39.
- ^ Кемп Т.С. (1982). Млекообразные рептилии и происхождение млекопитающих . Лондон: Академическая пресса. п. 363. ИСБН 978-0-12-404120-2 . ОСЛК 8613180 .
- ^ Эстес Р. (1961). «Краниальная анатомия рептилии-цинодонта Thrinaxodon liorhinus ». Бюллетень Музея сравнительной зоологии (1253 г.): 165–180.
- ^ « Тринаксодон: новое млекопитающее» . Ежедневные новости National Geographic. 11 февраля 2009 года. Архивировано из оригинала 14 февраля 2009 года . Проверено 26 августа 2012 г.
- ^ Jump up to: Перейти обратно: а б Байдек П., Кварнстрем М., Овоцкий К., Сулей Т., Сенников А.Г., Голубев В.К., Недзведски Г. (2015). «Микробиота и остатки пищи, включая возможные свидетельства присутствия волос до млекопитающих в копролитах верхней перми из России». Летайя . 49 (4): 455–477. дои : 10.1111/лет.12156 .
- ^ Бота-Бринк Дж., Ангельчик К.Д. (2010). «Объясняют ли необычайно высокие темпы роста пермско-триасовых дицинодонтов (Therapsida, Anomodontia) их успех до и после вымирания в конце пермского периода?» . Зоологический журнал Линнеевского общества . 160 (2): 341–365. дои : 10.1111/j.1096-3642.2009.00601.x .
- ^ Пол Г.С. (1988). Хищные динозавры мира . Нью-Йорк: Саймон и Шустер. п. 464 . ISBN 978-0-671-61946-6 . OCLC 18350868 .
- ^ Уотсон Дж. М., Грейвс Дж. А. (1988). «Монотремные клеточные циклы и эволюция гомеотермии». Австралийский журнал зоологии . 36 (5): 573–584. дои : 10.1071/ZO9880573 .
- ^ Макнаб Б.К. (1980). «Энергетика и пределы умеренного распространения броненосцев». Журнал маммологии . 61 (4): 606–627. дои : 10.2307/1380307 . JSTOR 1380307 .
- ^ Килан-Яворовска З., Хурум Дж.Х. (2006). «Положение конечностей у ранних млекопитающих: раскидистое или парасагиттальное» (PDF) . Acta Palaeontologica Polonica . 51 (3): 10237–10239. Архивировано (PDF) из оригинала 25 января 2024 года . Проверено 25 января 2024 г.
- ^ Лиллегравен Дж.А., Килан-Яворовска З., Клеменс В.А. (1979). Мезозойские млекопитающие: первые две трети истории млекопитающих . Издательство Калифорнийского университета. п. 321. ИСБН 978-0-520-03951-3 . OCLC 5910695 .
- ^ Офтедал ОТ (июль 2002 г.). «Молочная железа и ее происхождение в ходе эволюции синапсид». Журнал биологии молочной железы и неоплазии . 7 (3): 225–252. дои : 10.1023/А:1022896515287 . ПМИД 12751889 . S2CID 25806501 .
- ^ Офтедал ОТ (июль 2002 г.). «Происхождение лактации как источника воды для яиц в пергаментной скорлупе». Журнал биологии молочной железы и неоплазии . 7 (3): 253–266. дои : 10.1023/А:1022848632125 . ПМИД 12751890 . S2CID 8319185 .
- ^ «Развитие груди» . Техасская детская больница . Архивировано из оригинала 13 января 2021 года . Проверено 13 января 2021 г.
- ^ Пфафф, Кэтрин; Нагель, Дорис; Ганнелл, Грегг; Вебер, Герхард В.; Жажда, Юрген; Блэк, Майкл; Бастл, Кэтрин (2017). «Палеобиология Hyaenodon exiguus (Hyaenodonta, Mammalia) на основе морфометрического анализа костного лабиринта » Журнал анатомии . 230 (2): 282–289. дои : 10.1111/joa.12545 . ПМК 5244453 . ПМИД 27666133 .
- ^ Jump up to: Перейти обратно: а б Сахни С., Бентон М.Дж., Ферри, Пенсильвания (август 2010 г.). «Связи между глобальным таксономическим разнообразием, экологическим разнообразием и распространением позвоночных на суше» . Письма по биологии . 6 (4): 544–547. дои : 10.1098/rsbl.2009.1024 . ПМК 2936204 . ПМИД 20106856 .
- ^ Смит Ф.А., Бойер А.Г., Браун Дж.Х., Коста Д.П., Даян Т., Эрнест С.К. и др. (ноябрь 2010 г.). «Эволюция максимального размера тела наземных млекопитающих». Наука . 330 (6008): 1216–1219. Бибкод : 2010Sci...330.1216S . CiteSeerX 10.1.1.383.8581 . дои : 10.1126/science.1194830 . ПМИД 21109666 . S2CID 17272200 .
- ^ Симмонс Н.Б., Сеймур К.Л., Хаберсетцер Дж., Ганнелл Г.Ф. (февраль 2008 г.). «Примитивная летучая мышь раннего эоцена из Вайоминга и эволюция полета и эхолокации» . Природа . 451 (7180): 818–821. Бибкод : 2008Natur.451..818S . дои : 10.1038/nature06549 . hdl : 2027.42/62816 . ПМИД 18270539 . S2CID 4356708 . Архивировано из оригинала 25 января 2024 года . Проверено 25 января 2024 г.
- ^ Бининда-Эмондс О.Р., Кардилло М., Джонс К.Е., Макфи Р.Д., Бек Р.М., Греньер Р. и др. (март 2007 г.). «Замедленный рост современных млекопитающих» (PDF) . Природа . 446 (7135): 507–512. Бибкод : 2007Natur.446..507B . дои : 10.1038/nature05634 . ПМИД 17392779 . S2CID 4314965 . Архивировано (PDF) из оригинала 25 января 2024 года . Проверено 25 января 2024 г.
- ^ Jump up to: Перейти обратно: а б Уибл-младший, Ружье Г.В., Новачек М.Дж., Ашер Р.Дж. (июнь 2007 г.). «Меловые эвтерийцы и лавразийское происхождение плацентарных млекопитающих вблизи границы K/T». Природа . 447 (7147): 1003–1006. Бибкод : 2007Natur.447.1003W . дои : 10.1038/nature05854 . ПМИД 17581585 . S2CID 4334424 .
- ^ О'Лири М.А., Блох Дж.И., Флинн Дж.Дж., Годен Т.Дж., Джалломбардо А., Джаннини Н.П. и др. (февраль 2013 г.). «Предок плацентарных млекопитающих и пост-K-Pg-излучение плаценты» . Наука . 339 (6120): 662–667. Бибкод : 2013Sci...339..662O . дои : 10.1126/science.1229237 . hdl : 11336/7302 . ПМИД 23393258 . S2CID 206544776 . Архивировано из оригинала 10 ноября 2021 года . Проверено 30 июня 2022 г.
- ^ Холлидей Т.Дж., Апчерч П., Госвами А. (февраль 2017 г.). «Разрешение взаимоотношений плацентарных млекопитающих палеоцена» . Биологические обзоры Кембриджского философского общества . 92 (1): 521–550. дои : 10.1111/brv.12242 . ПМК 6849585 . ПМИД 28075073 .
- ^ Холлидей Т.Дж., Апчерч П., Госвами А. (июнь 2016 г.). «Эвтерийцы испытали повышенные темпы эволюции сразу после массового вымирания в мел-палеогеновом периоде» . Слушания. Биологические науки . 283 (1833): 20153026. doi : 10.1098/rspb.2015.3026 . ПМЦ 4936024 . ПМИД 27358361 .
- ^ Jump up to: Перейти обратно: а б с Ни Х, Гебо Д.Л., Дагосто М., Мэн Дж., Таффоро П., Флинн Дж.Дж., Бирд К.С. (июнь 2013 г.). «Самый старый известный скелет примата и ранняя эволюция гаплорина». Природа . 498 (7452): 60–64. Бибкод : 2013Natur.498...60N . дои : 10.1038/nature12200 . ПМИД 23739424 . S2CID 4321956 .
- ^ Ромер С.А., Парсонс Т.С. (1977). Тело позвоночного . Филадельфия: Холт-Сондерс Интернэшнл. стр. 129–145. ISBN 978-0-03-910284-5 . OCLC 60007175 .
- ^ Пурвес В.К., Садава Д.Е., Орианс Г.Х., Хелле Х.К. (2001). Жизнь: наука биологии (6-е изд.). Нью-Йорк: Sinauer Associates, Inc., с. 593. ИСБН 978-0-7167-3873-2 . OCLC 874883911 .
- ^ Антвал Н., Джоши Л., Такер А.С. (январь 2013 г.). «Эволюция среднего уха и челюсти млекопитающих: адаптации и новые структуры» . Журнал анатомии . 222 (1): 147–160. дои : 10.1111/j.1469-7580.2012.01526.x . ПМЦ 3552421 . ПМИД 22686855 .
- ^ ван Нивельт А.Ф., Смит К.К. (2005). «Заменять или не заменять: значение снижения функциональной замены зубов у сумчатых и плацентарных млекопитающих». Палеобиология . 31 (2): 324–346. doi : 10.1666/0094-8373(2005)031[0324:trontr]2.0.co;2 . S2CID 37750062 .
- ^ Либертини Дж., Феррара Н. (апрель 2016 г.). «Старение многолетних клеток и частей органов согласно парадигме программируемого старения» . Возраст . 38 (2): 35. дои : 10.1007/s11357-016-9895-0 . ПМЦ 5005898 . ПМИД 26957493 .
- ^ Мао Ф., Ван Ю, Мэн Дж (2015). «Систематическое исследование микроструктур зубной эмали Lambdopsalis Bulla (Multituberculate, Mammalia) – значение для мультитуберкулезной биологии и филогении» . ПЛОС ОДИН . 10 (5): e0128243. Бибкод : 2015PLoSO..1028243M . дои : 10.1371/journal.pone.0128243 . ПМЦ 4447277 . ПМИД 26020958 .
- ^ Осборн Х.Ф. (1900). «Происхождение млекопитающих, III. Затылочные мыщелки рептильного трехраздельного типа» . Американский натуралист . 34 (408): 943–947. дои : 10.1086/277821 . JSTOR 2453526 .
- ^ Кромптон А.В., Дженкинс-младший Ф.А. (1973). «Млекопитающие от рептилий: обзор происхождения млекопитающих». Ежегодный обзор наук о Земле и планетах . 1 : 131–155. Бибкод : 1973AREPS...1..131C . doi : 10.1146/annurev.ea.01.050173.001023 .
- ^ Jump up to: Перейти обратно: а б Power ML, Schulkin J (2013). Эволюция плаценты человека . Балтимор: Издательство Университета Джонса Хопкинса. стр. 1890–1891. ISBN 978-1-4214-0643-5 . OCLC 940749490 .
- ^ Линденфорс П., Гиттлман Дж.Л., Джонс К.Е. (2007). «Половой размерный диморфизм млекопитающих». Пол, размер и гендерные роли: эволюционные исследования диморфизма полового размера . Оксфорд: Издательство Оксфордского университета. стр. 16–26. ISBN 978-0-19-920878-4 . Архивировано из оригинала 25 января 2024 года . Проверено 25 января 2024 г.
- ^ Диерауф Л.А., Гулланд FM (2001). Справочник CRC по медицине морских млекопитающих: здоровье, болезни и реабилитация (2-е изд.). Бока-Ратон: CRC Press. п. 154. ИСБН 978-1-4200-4163-7 . OCLC 166505919 .
- ^ Луи Дж. Х., Хансен Д. В., Кригштейн А. Р. (июль 2011 г.). «Развитие и эволюция неокортекса человека» . Клетка . 146 (1): 18–36. дои : 10.1016/j.cell.2011.06.030 . ПМК 3610574 . ПМИД 21729779 .
- ^ Киллер CE (июнь 1933 г.). «Отсутствие мозолистого тела как менделизирующего признака у домовой мыши» . Труды Национальной академии наук Соединенных Штатов Америки . 19 (6): 609–611. Бибкод : 1933PNAS...19..609K . дои : 10.1073/pnas.19.6.609 . JSTOR 86284 . ПМК 1086100 . ПМИД 16587795 .
- ^ Стэндринг С., Борли Н.Р. (2008). Анатомия Грея: анатомические основы клинической практики (40-е изд.). Лондон: Черчилль Ливингстон. стр. 960–962. ISBN 978-0-8089-2371-8 . OCLC 213447727 .
- ^ Беттс Дж. Ф., Дезе П., Джонсон Э., Джонсон Дж. Э., Корол О., Круз Д. и др. (2013). Анатомия и физиология . Хьюстон: Издательство Университета Райса. стр. 787–846. ISBN 978-1-938168-13-0 . OCLC 898069394 . Архивировано из оригинала 23 февраля 2022 года . Проверено 25 января 2024 г.
- ^ Левицкий М.Г. (2013). «Механика дыхания». Легочная физиология (8-е изд.). Нью-Йорк: McGraw-Hill Medical. ISBN 978-0-07-179313-1 . OCLC 940633137 .
- ^ Jump up to: Перейти обратно: а б Умеш КБ (2011). «Легочная анатомия и физиология» . Справочник по механической вентиляции . Нью-Дели: Медицинское издательство Jaypee Brothers. п. 12. ISBN 978-93-80704-74-6 . OCLC 945076700 .
- ^ Jump up to: Перейти обратно: а б с д и ж г час я Фельдхамер Г.А., Дрикамер Л.С., Весси С.Х., Мерритт Дж.Ф., Краевски С. (2007). Маммология: адаптация, разнообразие, экология (3-е изд.). Балтимор: Издательство Университета Джонса Хопкинса. ISBN 978-0-8018-8695-9 . OCLC 124031907 .
- ^ Тинкер СВ (1988). Киты мира . Архив Брилла. п. 51. ИСБН 978-0-935848-47-2 .
- ^ Ромер А.С. (1959). История позвоночных (4-е изд.). Чикаго: Издательство Чикагского университета. ISBN 978-0-226-72490-4 .
- ^ де Мюзон К., Ланге-Бадре Б. (1997). «Плотоядные зубные адаптации трибосфенических млекопитающих и филогенетическая реконструкция». Летайя . 30 (4): 353–366. Бибкод : 1997Лета..30..353Д . дои : 10.1111/j.1502-3931.1997.tb00481.x .
- ^ Jump up to: Перейти обратно: а б Лангер П. (июль 1984 г.). «Сравнительная анатомия желудка травоядных млекопитающих». Ежеквартальный журнал экспериментальной физиологии . 69 (3): 615–625. doi : 10.1113/expphysicalol.1984.sp002848 . ПМИД 6473699 . S2CID 30816018 .
- ^ Воан Т.А., Райан Дж.М., Чаплевски, Нью-Джерси (2011). «Периссодактилия» . Маммология (5-е изд.). Джонс и Бартлетт. п. 322. ИСБН 978-0-7637-6299-5 . OCLC 437300511 .
- ^ Флауэр WH, Лидеккер Р. (1946). Введение в изучение ныне живущих и вымерших млекопитающих . Лондон: Адам и Чарльз Блэк. п. 496. ИСБН 978-1-110-76857-8 .
- ^ Шрикумар С (2010). Базовая физиология . PHI Learning Pvt. ООО, стр. 180–181. ISBN 978-81-203-4107-4 .
- ^ Чейфец А.С. (2010). Оксфордский американский справочник по гастроэнтерологии и гепатологии . Оксфорд: Издательство Оксфордского университета, США. п. 165. ИСБН 978-0-19-983012-1 .
- ^ Кунц Э (2008). Гепатология: Учебник и атлас . Германия: Шпрингер. п. 38. ISBN 978-3-540-76838-8 .
- ^ Ортис Р.М. (июнь 2001 г.). «Осморегуляция у морских млекопитающих» . Журнал экспериментальной биологии . 204 (Часть 11): 1831–1844. дои : 10.1242/jeb.204.11.1831 . ПМИД 11441026 . Архивировано из оригинала 25 января 2024 года . Проверено 25 января 2024 г.
- ^ Jump up to: Перейти обратно: а б с д Роман А.С., Парсонс Т.С. (1977). Тело позвоночного . Филадельфия: Холт-Сондерс Интернэшнл. стр. 396–399. ISBN 978-0-03-910284-5 .
- ^ Линзи, Дональд В. (2020). Биология позвоночных: систематика, таксономия, естественная история и охрана . Издательство Университета Джонса Хопкинса. п. 306. ИСБН 978-1-42143-733-0 . Архивировано из оригинала 22 января 2024 года . Проверено 22 января 2024 г.
- ^ «Биологические обзоры – Кембриджские журналы» . Архивировано из оригинала 22 ноября 2015 года . Проверено 21 января 2017 г.
- ^ Докинз Р., Вонг Ю. (2016). Рассказ предка: Паломничество на заре эволюции (2-е изд.). Бостон: Mariner Books. п. 281. ИСБН 978-0-544-85993-7 .
- ^ Jump up to: Перейти обратно: а б с Фитч WT (2006). «Производство вокализаций у млекопитающих» (PDF) . В Брауне К. (ред.). Энциклопедия языка и лингвистики . Оксфорд: Эльзевир. стр. 115–121. Архивировано из оригинала 1 июня 2024 года . Проверено 25 января 2024 г.
- ^ Ланжевен П., Барклай Р.М. (1990). « Гипсигнатус чудовищный » . Виды млекопитающих (357): 1–4. дои : 10.2307/3504110 . JSTOR 3504110 .
- ^ Вайсенгрубер Г.Е., Форстенпойнтнер Г., Петерс Г., Кюббер-Хейсс А., Fitch WT (сентябрь 2002 г.). «Подъязычный аппарат и глотка у льва (Panthera leo), ягуара (Panthera onca), тигра (Panthera tigris), гепарда (Acinonyxjubatus) и домашней кошки (Felis silvestris f. catus)» . Журнал анатомии . 201 (3): 195–209. дои : 10.1046/j.1469-7580.2002.00088.x . ПМК 1570911 . ПМИД 12363272 .
- ^ Штегер А.С., Хайльманн Г., Цеппельзауер М., Гансвиндт А., Хенсман С., Чарльтон Б.Д. (2012). «Визуализация звукового излучения слонов: свидетельства двух типов грохота» . ПЛОС ОДИН . 7 (11): е48907. Бибкод : 2012PLoSO...748907S . дои : 10.1371/journal.pone.0048907 . ПМЦ 3498347 . ПМИД 23155427 .
- ^ Кларк К.В. (2004). «Инфразвуковые звуки усатых китов: естественная изменчивость и функция». Журнал Акустического общества Америки . 115 (5): 2554. Бибкод : 2004ASAJ..115.2554C . дои : 10.1121/1.4783845 .
- ^ Jump up to: Перейти обратно: а б Доусон Т.Дж., Вебстер К.Н., Мэлони С.К. (февраль 2014 г.). «Мех млекопитающих в открытой среде; обязательно ли конфликтуют крипсис и тепловые потребности? Сравнение белого медведя и сумчатой коалы». Журнал сравнительной физиологии Б. 184 (2): 273–284. дои : 10.1007/s00360-013-0794-8 . ПМИД 24366474 . S2CID 9481486 .
- ^ Сломински А., Тобин Д.Д., Шибахара С., Вортсман Дж. (октябрь 2004 г.). «Меланиновая пигментация в коже млекопитающих и ее гормональная регуляция». Физиологические обзоры . 84 (4): 1155–1228. doi : 10.1152/physrev.00044.2003 . ПМИД 15383650 . S2CID 21168932 .
- ^ Хилтон-младший Б (1996). «Дикая природа Южной Каролины» . Цвета животных . 43 (4). Хилтон Понд Центр: 10–15. Архивировано из оригинала 25 января 2024 года . Проверено 26 ноября 2011 г.
- ^ Jump up to: Перейти обратно: а б Прум Р.О., Торрес Р.Х. (май 2004 г.). «Структурная окраска кожи млекопитающих: конвергентная эволюция когерентно рассеивающихся массивов дермального коллагена» (PDF) . Журнал экспериментальной биологии . 207 (Часть 12): 2157–2172. дои : 10.1242/jeb.00989 . hdl : 1808/1599 . ПМИД 15143148 . S2CID 8268610 . Архивировано (PDF) из оригинала 5 июня 2024 года . Проверено 25 января 2024 г.
- ^ Суутари М., Маянева М., Фьюер Д.П., Войрин Б., Айелло А., Фридл Т. и др. (март 2010 г.). «Молекулярные доказательства разнообразного сообщества зеленых водорослей, растущих в волосах ленивцев, и специфической связи с Trichophilus welckeri (Chlorophyta, Ulvophyceae)» . Эволюционная биология BMC . 10 (86): 86. Бибкод : 2010BMCEE..10...86S . дои : 10.1186/1471-2148-10-86 . ПМЦ 2858742 . ПМИД 20353556 .
- ^ Каро Т (2005). «Адаптационное значение окраски млекопитающих» . Бионаука . 55 (2): 125–136. doi : 10.1641/0006-3568(2005)055[0125:tasoci]2.0.co;2 .
- ^ Миллс Л.С., Зимова М., Ойлер Дж., Бегущий С., Абацоглу Дж.Т., Лукач П.М. (апрель 2013 г.). «Несоответствие камуфляжа сезонной окраски шерсти из-за уменьшения продолжительности снега» . Труды Национальной академии наук Соединенных Штатов Америки . 110 (18): 7360–7365. Бибкод : 2013PNAS..110.7360M . дои : 10.1073/pnas.1222724110 . ПМЦ 3645584 . ПМИД 23589881 .
- ^ Каро Т. (февраль 2009 г.). «Контрастная окраска у наземных млекопитающих» . Философские труды Лондонского королевского общества. Серия Б, Биологические науки . 364 (1516): 537–548. дои : 10.1098/rstb.2008.0221 . ПМК 2674080 . ПМИД 18990666 .
- ^ Плавкан Дж.М. (2001). «Половой диморфизм в эволюции приматов» . Американский журнал физической антропологии . Приложение 33 (33): 25–53. дои : 10.1002/ajpa.10011 . ПМИД 11786990 . S2CID 31722173 .
- ^ Брэдли Б.Дж., Джеральд М.С., Виддиг А., Манди Н.И. (2012). «Изменение цвета шерсти и экспрессия генов пигментации у макак-резус ( Macaca Mulatta )» (PDF) . Журнал эволюции млекопитающих . 20 (3): 263–270. дои : 10.1007/s10914-012-9212-3 . S2CID 13916535 . Архивировано из оригинала (PDF) 24 сентября 2015 года.
- ^ Каро Т. , Иззо А., Райнер Р.К., Уокер Х., Станкович Т. (апрель 2014 г.). «Функция полосок зебры» . Природные коммуникации . 5 : 3535. Бибкод : 2014NatCo...5.3535C . дои : 10.1038/ncomms4535 . ПМИД 24691390 . S2CID 9849814 .
- ^ Кобаяши К., Китано Т., Ивао Ю., Кондо М. (2018). Репродуктивные стратегии и стратегии развития: непрерывность жизни . Спрингер. п. 290. ИСБН 978-4-431-56609-0 .
- ^ Либби Генриетта Хайман (15 сентября 1992 г.). Сравнительная анатомия позвоночных Хаймана . Издательство Чикагского университета. стр. 583–. ISBN 978-0-226-87013-7 .
- ^ Леонард, Джанет; Кордова-Агилар, Алекс (19 июля 2010 г.). Эволюция первичных половых признаков у животных . Издательство Оксфордского университета, США. ISBN 978-0-19-532555-3 .
- ^ Ломбарди Дж. (1998). Сравнительное размножение позвоночных . Springer Science & Business Media. ISBN 978-0-7923-8336-9 .
- ^ Тиндейл-Биско Х., Ренфри М. (1987). Репродуктивная физиология сумчатых . Издательство Кембриджского университета. ISBN 978-0-521-33792-2 .
- ^ Джонстон С.Д., Смит Б., Пайн М., Стензель Д., Холт В.В. (2007). «Односторонняя эякуляция пучков сперматозоидов ехидны» (PDF) . Американский натуралист . 170 (6): E162–E164. дои : 10.1086/522847 . ПМИД 18171162 . S2CID 40632746 .
- ^ Бача-младший, Уильям Дж.; Бача, Линда М. (2012). Цветной атлас ветеринарной гистологии . Уайли. п. 308. ИСБН 978-1-11824-364-0 . Проверено 28 ноября 2023 г.
- ^ Кук, Фред; Брюс, Дженни (2004). Энциклопедия животных: полное наглядное руководство . Издательство Калифорнийского университета. п. 79. ИСБН 978-0-52024-406-1 . Проверено 28 ноября 2023 г.
- ^ Максвелл К.Е. (2013). Секс-императив: эволюционная история сексуального выживания . Спрингер. стр. 112–113. ISBN 978-1-4899-5988-1 .
- ^ Воган Т.А., Райан Дж.П., Чаплевски, Нью-Джерси (2011). Маммология . Издательство Джонс и Бартлетт. п. 387. ИСБН 978-0-03-025034-7 .
- ^ Jump up to: Перейти обратно: а б Хоффман Э.А., Роу ТБ (сентябрь 2018 г.). «Юрские перинаты стволовых млекопитающих и происхождение размножения и роста млекопитающих». Природа . 561 (7721): 104–108. Бибкод : 2018Natur.561..104H . дои : 10.1038/s41586-018-0441-3 . ПМИД 30158701 . S2CID 205570021 .
- ^ Уоллис М.К., Уотерс П.Д., Делбридж М.Л., Кирби П.Дж., Паск А.Дж., Грюцнер Ф. и др. (2007). «Определение пола у утконоса и ехидны: аутосомное расположение SOX3 подтверждает отсутствие SRY у однопроходных». Хромосомные исследования . 15 (8): 949–959. дои : 10.1007/s10577-007-1185-3 . ПМИД 18185981 . S2CID 812974 .
- ^ Маршалл Грейвс, Дж. А. (2008). «Странные геномы животных и эволюция пола и половых хромосом позвоночных» (PDF) . Ежегодный обзор генетики . 42 : 565–586. дои : 10.1146/annurev.genet.42.110807.091714 . ПМИД 18983263 . Архивировано из оригинала (PDF) 4 сентября 2012 года . Проверено 25 января 2024 г.
- ^ Салли М (2005). «Поведение и образ жизни млекопитающих» . Млекопитающие . Чикаго: Рейнтри. п. 6. ISBN 978-1-4109-1050-9 . OCLC 53476660 .
- ^ Верма П.С., Панди Б.П. (2013). Книга ISC по биологии ISC для XI класса . Нью-Дели: С. Чанд и компания. п. 288. ИСБН 978-81-219-2557-0 .
- ^ Офтедал ОТ (июль 2002 г.). «Молочная железа и ее происхождение в ходе эволюции синапсид». Журнал биологии молочной железы и неоплазии . 7 (3): 225–252. дои : 10.1023/а:1022896515287 . ПМИД 12751889 . S2CID 25806501 .
- ^ Крокенбергер А (2006). «Лактация». Дикман Ч.Р., Армати П.Дж., Хьюм И.Д. (ред.). Сумчатые . Издательство Кембриджского университета. п. 109. ИСБН 978-1-139-45742-2 .
- ^ Щулкин Дж. Power ML (2016). Молоко: биология лактации . Издательство Университета Джонса Хопкинса. п. 66. ИСБН 978-1-4214-2042-4 .
- ^ Томпсон К.В., Бейкер А.Дж., Бейкер А.М. (2010). «Отцовская забота и развитие поведения млекопитающих в неволе». Клейман Д.Г., Томпсон К.В., Баер К.К. (ред.). Дикие млекопитающие в неволе. Принципы и методы управления зоопарком (2-е изд.). Издательство Чикагского университета. п. 374. ИСБН 978-0-226-44011-8 .
- ^ Кэмпбелл Н.А., Рис Дж.Б. (2002). Биология (6-е изд.). Бенджамин Каммингс. п. 845 . ISBN 978-0-8053-6624-2 . OCLC 47521441 .
- ^ Буффенштейн Р., Яхав С. (1991). «Является ли голый землекоп Hererocephalus glaber эндотермическим, но пойкилотермным млекопитающим?». Журнал термической биологии . 16 (4): 227–232. Бибкод : 1991JTBio..16..227B . дои : 10.1016/0306-4565(91)90030-6 .
- ^ Шмидт-Нильсен К., Дюк Дж.Б. (1997). «Температурные эффекты» . Физиология животных: адаптация и окружающая среда (5-е изд.). Кембридж: Издательство Кембриджского университета. п. 218. ИСБН 978-0-521-57098-5 . OCLC 35744403 .
- ^ Jump up to: Перейти обратно: а б Лоренцини А., Джонсон Ф.Б., Оливер А., Трезини М., Смит Дж.С., Хдейб М. и др. (2009). «Значительная корреляция продолжительности жизни видов с распознаванием двухцепочечных разрывов ДНК, но не с длиной теломер» . Механизмы старения и развития . 130 (11–12): 784–792. дои : 10.1016/j.mad.2009.10.004 . ПМК 2799038 . ПМИД 19896964 .
- ^ Харт Р.В., Сетлоу Р.Б. (июнь 1974 г.). «Корреляция между эксцизионным восстановлением дезоксирибонуклеиновой кислоты и продолжительностью жизни у ряда видов млекопитающих» . Труды Национальной академии наук Соединенных Штатов Америки . 71 (6): 2169–2173. Бибкод : 1974PNAS...71.2169H . дои : 10.1073/pnas.71.6.2169 . ПМЦ 388412 . ПМИД 4526202 .
- ^ Ма С., Упнея А., Галецкий А., Цай Ю.М., Бурант К.Ф., Раскинд С. и др. (ноябрь 2016 г.). «Профилирование млекопитающих на основе клеточных культур показывает, что восстановление ДНК и метаболизм являются определяющими факторами долголетия вида» . электронная жизнь . 5 . doi : 10.7554/eLife.19130 . ПМК 5148604 . ПМИД 27874830 .
- ^ Грубе К., Бюркле А. (декабрь 1992 г.). «Активность поли(АДФ-рибозы)-полимеразы в мононуклеарных лейкоцитах 13 видов млекопитающих коррелирует с видоспецифичной продолжительностью жизни» . Труды Национальной академии наук Соединенных Штатов Америки . 89 (24): 11759–11763. Бибкод : 1992PNAS...8911759G . дои : 10.1073/pnas.89.24.11759 . ПМК 50636 . ПМИД 1465394 .
- ^ Фрэнсис А.А., Ли В.Х., Риган Дж.Д. (июнь 1981 г.). «Связь эксцизионной репарации ДНК поражений, вызванных ультрафиолетом, с максимальной продолжительностью жизни млекопитающих». Механизмы старения и развития . 16 (2): 181–189. дои : 10.1016/0047-6374(81)90094-4 . ПМИД 7266079 . S2CID 19830165 .
- ^ Третон Дж. А., Куртуа Ю. (март 1982 г.). «Корреляция между эксцизионной репарацией ДНК и продолжительностью жизни млекопитающих в эпителиальных клетках хрусталика». Международные отчеты по клеточной биологии . 6 (3): 253–260. doi : 10.1016/0309-1651(82)90077-7 (неактивен 31 января 2024 г.). ПМИД 7060140 .
{{cite journal}}: CS1 maint: DOI неактивен по состоянию на январь 2024 г. ( ссылка ) - ^ Маслански CJ, Williams GM (февраль 1985 г.). «Синтез репарации ДНК, индуцированный ультрафиолетовым светом, в гепатоцитах видов с разной продолжительностью жизни». Механизмы старения и развития . 29 (2): 191–203. дои : 10.1016/0047-6374(85)90018-1 . ПМИД 3974310 . S2CID 23988416 .
- ^ Уокер В.Ф., Хомбергер Д.Г. (1998). Анатомия и вскрытие плода свиньи (5-е изд.). Нью-Йорк: WH Freeman and Company. п. 3. ISBN 978-0-7167-2637-1 . OCLC 40576267 .
- ^ Орр CM (ноябрь 2005 г.). «Муравьед, ходящий на костяшках пальцев: тест конвергенции адаптации предполагаемых особенностей ходьбы на костяшках пальцев африканских гоминид». Американский журнал физической антропологии . 128 (3): 639–658. дои : 10.1002/ajpa.20192 . ПМИД 15861420 .
- ^ Фиш Ф.Э., Фраппелл П.Б., Бодинетт Р.В., Макфарлейн П.М. (февраль 2001 г.). «Энергетика наземного передвижения утконоса Ornithorhynchus anatinus» (PDF) . Журнал экспериментальной биологии . 204 (Часть 4): 797–803. дои : 10.1242/jeb.204.4.797 . hdl : 2440/12192 . ПМИД 11171362 . Архивировано (PDF) из оригинала 14 марта 2024 года . Проверено 25 января 2024 г.
- ^ Дхингра П. (2004). «Сравнительное прямохождение – как остальные представители животного мира ходят на двух ногах» . Антропологическая наука . 131 (231). Архивировано из оригинала 21 апреля 2021 года . Проверено 11 марта 2017 г.
- ^ Александр Р.М. (май 2004 г.). «Двуногие животные и их отличия от человека» . Журнал анатомии . 204 (5): 321–330. дои : 10.1111/j.0021-8782.2004.00289.x . ПМК 1571302 . ПМИД 15198697 .
- ^ Jump up to: Перейти обратно: а б Дагг А.И. (1973). «Походка млекопитающих». Обзор млекопитающих . 3 (4): 135–154. дои : 10.1111/j.1365-2907.1973.tb00179.x .
- ^ Робертс ТД (1995). Понимание баланса: механика позы и передвижения . Сан-Диего: Нельсон Торнс. п. 211. ИСБН 978-1-56593-416-0 . OCLC 33167785 .
- ^ Jump up to: Перейти обратно: а б с Картмилл М (1985). «Восхождение». Хильдебранд М., Брамбл Д.М., Лием К.Ф., Wake DB (ред.). Функциональная морфология позвоночных . Кембридж: Белкнап Пресс. стр. 73–88. ISBN 978-0-674-32775-7 . ОСЛК 11114191 .
- ^ Верн К. (2001). «Скольжение северной летяги ( Glaucomys sabrinus ) в зрелом смешанном лесу восточной Канады» . Журнал маммологии . 82 (4): 1026–1033. doi : 10.1644/1545-1542(2001)082<1026:GPOTNF>2.0.CO;2 . S2CID 78090049 .
- ^ Барба Л.А. (октябрь 2011 г.). «Летучие мыши – единственные летающие млекопитающие» . Биовоздушное передвижение . Архивировано из оригинала 14 мая 2016 года . Проверено 20 мая 2016 г.
- ^ «Летучие мыши в полете демонстрируют неожиданную аэродинамику» . ScienceDaily . 2007. Архивировано из оригинала 19 декабря 2019 года . Проверено 12 июля 2016 г.
- ^ Хеденстрем А., Йоханссон LC (март 2015 г.). «Полет летучей мыши: аэродинамика, кинематика и морфология полета» (PDF) . Журнал экспериментальной биологии . 218 (Часть 5): 653–663. дои : 10.1242/jeb.031203 . ПМИД 25740899 . S2CID 21295393 . Архивировано (PDF) из оригинала 25 января 2024 года . Проверено 25 января 2024 г.
- ^ «Летучие мыши экономят энергию, сжимая крылья при взмахе вверх» . ScienceDaily . 2012. Архивировано из оригинала 31 мая 2021 года . Проверено 12 июля 2016 г.
- ^ Карен Т. (2008). Встреча с летучими мышами: экобаты, вампиры и кинозвезды . Альбукерке: Издательство Университета Нью-Мексико. п. 14. ISBN 978-0-8263-4403-8 . OCLC 191258477 .
- ^ Стербинг-Д'Анджело С., Чадха М., Чиу С., Фальк Б., Сиань В., Барсело Дж. и др. (июль 2011 г.). «Датчики крыльев летучей мыши поддерживают управление полетом» . Труды Национальной академии наук Соединенных Штатов Америки . 108 (27): 11291–11296. Бибкод : 2011PNAS..10811291S . дои : 10.1073/pnas.1018740108 . ПМК 3131348 . ПМИД 21690408 .
- ^ Дамиани, Р., 2003, Самые ранние свидетельства рытья нор цинодонтов, Издательство Королевского общества, том 270, выпуск 1525.
- ^ Шимер Х.В. (1903). «Адаптации к водным, древесным, ископаемым и бегущим привычкам млекопитающих. III. Ископаемые адаптации» . Американский натуралист . 37 (444): 819–825. дои : 10.1086/278368 . JSTOR 2455381 . S2CID 83519668 . Архивировано из оригинала 9 апреля 2023 года . Проверено 23 августа 2020 г. .
- ^ Стэнхоуп М.Дж., Уодделл В.Г., Мэдсен О., де Йонг В., Хеджес С.Б., Клевен Г.К. и др. (август 1998 г.). «Молекулярные доказательства множественного происхождения насекомоядных и нового порядка эндемичных африканских насекомоядных млекопитающих» . Труды Национальной академии наук Соединенных Штатов Америки . 95 (17): 9967–9972. Бибкод : 1998PNAS...95.9967S . дои : 10.1073/pnas.95.17.9967 . ПМК 21445 . ПМИД 9707584 .
- ^ Перри Д.А. (1949). «Анатомические основы плавания китов». Журнал зоологии . 119 (1): 49–60. дои : 10.1111/j.1096-3642.1949.tb00866.x .
- ^ Фиш Ф.Е., Хуэй, Калифорния (1991). «Плавание дельфинов – обзор» (PDF) . Обзор млекопитающих . 21 (4): 181–195. дои : 10.1111/j.1365-2907.1991.tb00292.x . Архивировано из оригинала (PDF) 29 августа 2006 года.
- ^ Марш Х (1989). «Глава 57: Дюгонгиды» (PDF) . Фауна Австралии . Том. 1. Канберра: Публикации правительства Австралии. ISBN 978-0-644-06056-1 . ОСЛК 27492815 . Архивировано из оригинала (PDF) 11 мая 2013 года.
- ^ Jump up to: Перейти обратно: а б Берта А (2012). «Разнообразие ластоногих: эволюция и адаптация». Возвращение в море: жизнь и эпоха эволюции морских млекопитающих . Издательство Калифорнийского университета. стр. 62–64. ISBN 978-0-520-27057-2 .
- ^ Jump up to: Перейти обратно: а б Фиш Ф.Е., Херли Дж., Коста Д.П. (февраль 2003 г.). «Маневренность морского льва Zalophus Californianus: поворотные качества нестабильной конструкции тела» . Журнал экспериментальной биологии . 206 (Часть 4): 667–674. дои : 10.1242/jeb.00144 . ПМИД 12517984 .
- ^ Jump up to: Перейти обратно: а б Ридман М (1990). Ластоногие: тюлени, морские львы и моржи . Издательство Калифорнийского университета. ISBN 978-0-520-06497-3 . OCLC 19511610 .
- ^ Фиш ФЭ (1996). «Переход от движения на основе сопротивления к подъему в плавании млекопитающих» . Интегративная и сравнительная биология . 36 (6): 628–641. дои : 10.1093/icb/36.6.628 .
- ^ Рыба ФЭ (2000). «Биомеханика и энергетика водных и околоводных млекопитающих: от утконоса до кита» (PDF) . Физиологическая и биохимическая зоология . 73 (6): 683–698. CiteSeerX 10.1.1.734.1217 . дои : 10.1086/318108 . ПМИД 11121343 . S2CID 49732160 . Архивировано из оригинала (PDF) 4 августа 2016 года.
- ^ Элтрингем СК (1999). «Анатомия и физиология». Бегемоты . Лондон: T & AD Poyser Ltd., с. 8. ISBN 978-0-85661-131-5 . OCLC 42274422 .
- ^ «Гиппопотам Бегемот-амфибия » . Нэшнл Географик . Архивировано из оригинала 25 ноября 2014 года . Проверено 30 апреля 2016 г.
- ^ Jump up to: Перейти обратно: а б Сейфарт Р.М., Чейни Д.Л., Марлер П. (1980). «Тревожные звонки верветок: семантическая коммуникация у свободно живущего примата» . Поведение животных . 28 (4): 1070–1094. дои : 10.1016/S0003-3472(80)80097-2 . S2CID 53165940 . Архивировано из оригинала 12 сентября 2019 года . Проверено 22 сентября 2018 г.
- ^ Зубербюлер К (2001). «Тревожные сигналы, характерные для хищников, у обезьян Кэмпбелла, Cercopithecus Campbelli ». Поведенческая экология и социобиология . 50 (5): 414–442. дои : 10.1007/s002650100383 . JSTOR 4601985 . S2CID 21374702 .
- ^ Слаббекоорн Х., Смит Т.Б. (апрель 2002 г.). «Пение птиц, экология и видообразование» . Философские труды Лондонского королевского общества. Серия Б, Биологические науки . 357 (1420): 493–503. дои : 10.1098/rstb.2001.1056 . ПМК 1692962 . ПМИД 12028787 .
- ^ Баннистер Дж.Л. (2008). «Усатые киты (Mysticetes)» . В Perrin WF, Würsig B, Thewissen JG (ред.). Энциклопедия морских млекопитающих (2-е изд.). Академическая пресса. стр. 80–89. ISBN 978-0-12-373553-9 .
- ^ Скотт Н. (2002). «Существа культуры? Обоснование культурных систем у китов и дельфинов» . Бионаука . 52 (1): 9–14. doi : 10.1641/0006-3568(2002)052[0009:COCMTC]2.0.CO;2 . S2CID 86121405 .
- ^ Бофман Дж.В. (февраль 1998 г.). «Вокальное обучение больших остроносых летучих мышей» . Слушания. Биологические науки . 265 (1392): 227–233. дои : 10.1098/rspb.1998.0286 . ПМК 1688873 . ПМИД 9493408 .
- ^ «Язык луговых собачек расшифрован учеными» . Новости Си-Би-Си. 21 июня 2013 года. Архивировано из оригинала 22 мая 2015 года . Проверено 20 мая 2015 г.
- ^ Мэйелл Х. (3 марта 2004 г.). «Слоны звонят по междугородной связи в нерабочее время» . Нэшнл Географик . Архивировано из оригинала 5 марта 2004 года . Проверено 15 ноября 2016 г.
- ^ Смит Дж. М. , Харпер Д. (2003). Сигналы животных . Оксфордская серия по экологии и эволюции. Издательство Оксфордского университета. стр. 61–63. ISBN 978-0-19-852684-1 . OCLC 54460090 .
- ^ Компакт-диск ФитцГиббона, Фэншоу Дж.Х. (1988). «Стоттинг в газелях Томсона: честный сигнал о состоянии» (PDF) . Поведенческая экология и социобиология . 23 (2): 69–74. дои : 10.1007/bf00299889 . S2CID 2809268 . Архивировано из оригинала (PDF) 25 февраля 2014 года.
- ^ Бильдштейн К.Л. (май 1983 г.). «Почему белохвостые олени машут хвостами». Американский натуралист . 121 (5): 709–715. дои : 10.1086/284096 . JSTOR 2460873 . S2CID 83504795 .
- ^ Гослинг Л.М. (январь 1982 г.). «Переоценка функции ароматической маркировки территорий» . Zeitschrift für Tierpsychologie . 60 (2): 89–118. дои : 10.1111/j.1439-0310.1982.tb00492.x . Архивировано (PDF) из оригинала 27 марта 2018 года . Проверено 12 октября 2019 г.
- ^ Зала С.М., Поттс В.К., Пенн Диджей (март 2004 г.). «Дисплеи с ароматической маркировкой дают честные сигналы о здоровье и инфекции». Поведенческая экология . 15 (2): 338–344. дои : 10.1093/beheco/arh022 . hdl : 10.1093/beheco/arh022 .
- ^ Джонсон Р.П. (август 1973 г.). «Ароматная маркировка млекопитающих». Поведение животных . 21 (3): 521–535. дои : 10.1016/S0003-3472(73)80012-0 .
- ^ Шевилл В.Е., Макбрайд А.Ф. (1956). «Доказательства эхолокации китообразных». Глубоководные исследования . 3 (2): 153–154. Бибкод : 1956DSR.....3..153S . дои : 10.1016/0146-6313(56)90096-х .
- ^ Уилсон В., Мосс С. (2004). Томас Дж. (ред.). Эхолокация у летучих мышей и дельфинов . Издательство Чикагского университета. п. 22. ISBN 978-0-226-79599-7 . OCLC 50143737 .
- ^ Au WW (1993). Сонар дельфинов . Спрингер-Верлаг. ISBN 978-3-540-97835-0 . OCLC 26158593 .
- ^ Сандерс Дж.Г., Бейхман А.С., Роман Дж., Скотт Дж.Дж., Эмерсон Д., Маккарти Дж.Дж., Гиргис П.Р. (сентябрь 2015 г.). «У усатых китов имеется уникальный кишечный микробиом, сходный как с плотоядными, так и с травоядными» . Природные коммуникации . 6 : 8285. Бибкод : 2015NatCo...6.8285S . дои : 10.1038/ncomms9285 . ПМЦ 4595633 . ПМИД 26393325 .
- ^ Представитель JR (1996). «Энергетика и эволюция размеров тела мелких наземных млекопитающих» (PDF) . Симпозиумы Лондонского зоологического общества (69): 69–81. Архивировано из оригинала (PDF) 2 июня 2021 года . Проверено 31 мая 2016 г.
- ^ Jump up to: Перейти обратно: а б Уилсон Д.Е., Берни Д., ред. (2001). Животные: Полный визуальный путеводитель по дикой природе мира . Издательство ДК. стр. 86–89. ISBN 978-0-7894-7764-4 . OCLC 46422124 .
- ^ Jump up to: Перейти обратно: а б Ван Валкенбург Б. (июль 2007 г.). «Дежавю: эволюция морфологии питания хищных животных» . Интегративная и сравнительная биология . 47 (1): 147–163. дои : 10.1093/icb/icm016 . ПМИД 21672827 .
- ^ Сакко Т., ван Валкенбург Б. (2004). «Экоморфологические показатели пищевого поведения медведей (Carnivora: Ursidae)». Журнал зоологии . 263 (1): 41–54. дои : 10.1017/S0952836904004856 .
- ^ Певица М.С., Бернейс Е.А. (2003). «Понимание всеядности требует поведенческой точки зрения» . Экология . 84 (10): 2532–2537. Бибкод : 2003Ecol...84.2532S . дои : 10.1890/02-0397 .
- ^ Хатсон Дж. М., Берк К.С., Хейнс Дж. (1 декабря 2013 г.). «Остеофагия и модификации костей жирафа и других крупных копытных». Журнал археологической науки . 40 (12): 4139–4149. Бибкод : 2013JArSc..40.4139H . дои : 10.1016/j.jas.2013.06.004 .
- ^ «Почему кошки едят траву?» . Пет доктор медицины. Архивировано из оригинала 10 декабря 2016 года . Проверено 13 января 2017 г.
- ^ Гейзер Ф (2004). «Скорость обмена веществ и снижение температуры тела во время спячки и ежедневного оцепенения». Ежегодный обзор физиологии . 66 : 239–274. doi : 10.1146/annurev.phyol.66.032102.115105 . ПМИД 14977403 . S2CID 22397415 .
- ^ Хамфрис М.М., Томас Д.В., Крамер Д.Л. (2003). «Роль доступности энергии в спячке млекопитающих: подход с точки зрения затрат и выгод». Физиологическая и биохимическая зоология . 76 (2): 165–179. дои : 10.1086/367950 . ПМИД 12794670 . S2CID 14675451 .
- ^ Барнс Б.М. (июнь 1989 г.). «Избежание замерзания у млекопитающих: температура тела ниже 0 градусов Цельсия в арктической спячке». Наука . 244 (4912): 1593–1595. Бибкод : 1989Sci...244.1593B . дои : 10.1126/science.2740905 . ПМИД 2740905 .
- ^ Фриц Г. (2010). «Праздник млекопитающих и птиц» . В Навасе, Калифорния, Карвалью Дж.Э. (ред.). Летний период: молекулярные и физиологические аспекты . Прогресс молекулярной и субклеточной биологии. Том. 49. Шпрингер-Верлаг. стр. 95–113. дои : 10.1007/978-3-642-02421-4 . ISBN 978-3-642-02420-7 .
- ^ Майер, с. 59.
- ^ Гроув Дж.С., Грей Л.А., Ла Санта Медина Н., Сивакумар Н., Ан Дж.С., Корпус ТВ, Берке Дж.Д., Крейцер AC, Найт З.А. (июль 2022 г.). «Дофаминовые подсистемы, отслеживающие внутренние состояния» . Природа . 608 (7922): 374–380. Бибкод : 2022Natur.608..374G . дои : 10.1038/s41586-022-04954-0 . ПМЦ 9365689 . ПМИД 35831501 .
- ^ Jump up to: Перейти обратно: а б с д и Веник, с. 105.
- ^ Смит, с. 238.
- ^ «Кошачьи языки используют сложную физику» . 12 ноября 2010 г.
- ^ Смит, с. 237.
- ^ Майер, с. 54.
- ^ «Как жирафы пьют воду?» . Февраль 2016.
- ^ Манн Дж., Паттерсон Э.М. (ноябрь 2013 г.). «Использование орудий водными животными» . Философские труды Лондонского королевского общества. Серия Б, Биологические науки . 368 (1630): 20120424. doi : 10.1098/rstb.2012.0424 . ПМК 4027413 . ПМИД 24101631 .
- ^ Раффаэле П. (2011). Среди человекообразных обезьян: Приключения по следам наших ближайших сородичей . Нью-Йорк: Харпер. п. 83. ИСБН 978-0-06-167184-5 . OCLC 674694369 .
- ^ Келер В. (1925). Менталитет обезьян . Ливерайт. ISBN 978-0-87140-108-3 . OCLC 2000769 .
- ^ Макгоуэн Р.Т., Рен Т., Норлинг Ю., Килинг Л.Дж. (май 2014 г.). «Позитивный эффект и обучение: изучение «эффекта Эврики» у собак». Познание животных . 17 (3): 577–587. дои : 10.1007/s10071-013-0688-x . ПМИД 24096703 . S2CID 15216926 .
- ^ Карбовски Дж. (май 2007 г.). «Глобальное и региональное масштабирование метаболизма мозга и его функциональные последствия» . БМК Биология . 5 (18): 18. arXiv : 0705.2913 . Бибкод : 2007arXiv0705.2913K . дои : 10.1186/1741-7007-5-18 . ПМК 1884139 . ПМИД 17488526 .
- ^ Марино Л. (июнь 2007 г.). «Мозг китообразных: насколько они водные?» . Анатомическая запись . 290 (6): 694–700. дои : 10.1002/ar.20530 . ПМИД 17516433 . S2CID 27074107 . Архивировано из оригинала 20 марта 2020 года . Проверено 5 октября 2019 г.
- ^ Галоп Г.Г. (январь 1970 г.). «Шимпанзе: самопознание». Наука . 167 (3914): 86–87. Бибкод : 1970Sci...167...86G . дои : 10.1126/science.167.3914.86 . ПМИД 4982211 . S2CID 145295899 .
- ^ Плотник Дж. М., де Ваал Ф. Б., Рейсс Д. (ноябрь 2006 г.). «Самоузнавание в азиатском слоне» (PDF) . Труды Национальной академии наук Соединенных Штатов Америки . 103 (45): 17053–17057. Бибкод : 2006PNAS..10317053P . дои : 10.1073/pnas.0608062103 . ПМЦ 1636577 . ПМИД 17075063 . Архивировано (PDF) из оригинала 25 января 2024 года . Проверено 25 января 2024 г.
- ^ Роберт С. (1986). «Онтогенез зеркального поведения у двух видов человекообразных обезьян». Американский журнал приматологии . 10 (2): 109–117. дои : 10.1002/ajp.1350100202 . ПМИД 31979488 . S2CID 85330986 .
- ^ Вальравен В., ван Эльсакер Л., Верхейен Р. (1995). «Реакция группы карликовых шимпанзе (Pan paniscus) на свои зеркальные изображения: свидетельства самопознания». Приматы . 36 : 145–150. дои : 10.1007/bf02381922 . S2CID 38985498 .
- ^ Лики Р. (1994). «Происхождение разума» . Происхождение человечества . Нью-Йорк: BasicBooks. п. 150. ИСБН 978-0-465-05313-1 . ОСЛК 30739453 .
- ^ Арчер Дж (1992). Этология и развитие человека . Роуман и Литтлфилд. стр. 215–218. ISBN 978-0-389-20996-6 . OCLC 25874476 .
- ^ Jump up to: Перейти обратно: а б Мартен К., Псаракос С. (1995). «Свидетельства самосознания у афалин ( Tursiops truncatus )». В Паркер С.Т., Митчелл Р., Бочча М. (ред.). Самосознание у животных и людей: перспективы развития . Кембридж: Издательство Кембриджского университета. стр. 361–379. ISBN 978-0-521-44108-7 . ОСЛК 28180680 .
- ^ Jump up to: Перейти обратно: а б Дельфур Ф., Мартен К. (апрель 2001 г.). «Обработка зеркальных изображений трех видов морских млекопитающих: косаток (Orcinus orca), ложных косаток (Pseudorca crassidens) и калифорнийских морских львов (Zalophus Californianus)». Поведенческие процессы . 53 (3): 181–190. дои : 10.1016/s0376-6357(01)00134-6 . ПМИД 11334706 . S2CID 31124804 .
- ^ Джарвис Дж. Ю. (май 1981 г.). «Эусоциальность млекопитающих: совместное размножение в колониях голых землекопов». Наука . 212 (4494): 571–573. Бибкод : 1981Sci...212..571J . дои : 10.1126/science.7209555 . JSTOR 1686202 . ПМИД 7209555 . S2CID 880054 .
- ^ Джейкобс Д.С., Беннетт, Северная Каролина, Джарвис Дж., Кроу Т.М. (1991). «Структура колонии и иерархия доминирования дамаралендского землекопа Cryptomys damarensis (Rodentia: Bathyergidae) из Намибии». Журнал зоологии . 224 (4): 553–576. дои : 10.1111/j.1469-7998.1991.tb03785.x .
- ^ Харди С.Б. (2009). Матери и другие: эволюционные истоки взаимопонимания . Бостон: Belknap Press издательства Гарвардского университета. стр. 92–93.
- ^ Харлоу HF, Суоми SJ (июль 1971 г.). «Социальное восстановление обезьян, выращенных в изоляции» . Труды Национальной академии наук Соединенных Штатов Америки . 68 (7): 1534–1538. Бибкод : 1971PNAS...68.1534H . дои : 10.1073/pnas.68.7.1534 . ПМЦ 389234 . ПМИД 5283943 .
- ^ ван Шайк CP (январь 1999 г.). «Социоэкология социальности деления-слияния у орангутанов». Приматы; Журнал приматологии . 40 (1): 69–86. дои : 10.1007/BF02557703 . ПМИД 23179533 . S2CID 13366732 .
- ^ Арчи Э.А., Мосс СиДжей, Альбертс СК (март 2006 г.). «Связующие узы: генетическое родство предсказывает деление и слияние социальных групп у диких африканских слонов» . Слушания. Биологические науки . 273 (1586): 513–522. дои : 10.1098/rspb.2005.3361 . ПМК 1560064 . ПМИД 16537121 .
- ^ Смит Дж. Э., Меминис С. К., Холекамп К. Э. (2007). «Выбор партнера в зависимости от ранга в обществе деления-слияния пятнистой гиены ( Crocuta crocuta )» (PDF) . Поведенческая экология и социобиология . 61 (5): 753–765. дои : 10.1007/s00265-006-0305-y . S2CID 24927919 . Архивировано из оригинала (PDF) 25 апреля 2014 года.
- ^ Матоба Т., Куцукаке Н., Хасегава Т. (2013). Хейворд М. (ред.). «Потирание и облизывание головы укрепляют социальные связи в группе содержащихся в неволе африканских львов Panthera leo» . ПЛОС ОДИН . 8 (9): e73044. Бибкод : 2013PLoSO...873044M . дои : 10.1371/journal.pone.0073044 . ПМЦ 3762833 . ПМИД 24023806 .
- ^ Круцен М., Барре Л.М., Коннор Р.К., Манн Дж., Шервин В.Б. (июль 2004 г.). « О отец, где ты?» – Оценка отцовства в открытом обществе деления-слияния диких афалин (Tursiops sp.) в заливе Шарк, Западная Австралия». Молекулярная экология . 13 (7): 1975–1990. Бибкод : 2004МолЭк..13.1975К . дои : 10.1111/j.1365-294X.2004.02192.x . ПМИД 15189218 . S2CID 4510393 .
- ^ Мартин С (1991). Тропические леса Западной Африки: экология – угрозы – сохранение . Спрингер. дои : 10.1007/978-3-0348-7726-8 . ISBN 978-3-0348-7726-8 .
- ^ ле Ру А., Черри М.И., Гайгакс Л. (5 мая 2009 г.). «Поведение бдительности и последствия фитнеса: сравнение одиночного добывания пищи и млекопитающего, обязательного группового добывания пищи». Поведенческая экология и социобиология . 63 (8): 1097–1107. дои : 10.1007/s00265-009-0762-1 . S2CID 21961356 .
- ^ Палаги Э, Норша I (2015). Сэмондс К.Э. (ред.). «Сезон мира: примирение деспотического вида (Lemur catta)» . ПЛОС ОДИН . 10 (11): e0142150. Бибкод : 2015PLoSO..1042150P . дои : 10.1371/journal.pone.0142150 . ПМЦ 4646466 . ПМИД 26569400 .
- ^ Ист МЛ, Хофер Х (2000). «Самцы пятнистых гиен ( Crocuta crocuta ) стоят в очереди за статусом в социальных группах, где доминируют самки» . Поведенческая экология . 12 (15): 558–568. дои : 10.1093/beheco/12.5.558 .
- ^ Сэмюэлс А., Силк Дж.Б. , Родман П. (1984). «Изменения в ранге доминирования и репродуктивном поведении самцов шляпных макак ( Macaca radiate )». Поведение животных . 32 (4): 994–1003. дои : 10.1016/s0003-3472(84)80212-2 . S2CID 53186523 .
- ^ Дельпьетро Х.А., Руссо Р.Г. (2002). «Наблюдения за обыкновенной летучей мышью-вампиром ( Desmodus rotundus ) и волосатоногой летучей мышью-вампиром ( Diphylla ecaudata ) в неволе». Биология млекопитающих . 67 (2): 65–78. Бибкод : 2002МамБи..67...65D . дои : 10.1078/1616-5047-00011 .
- ^ Клейман Д.Г. (март 1977 г.). «Моногамия у млекопитающих». Ежеквартальный обзор биологии . 52 (1): 39–69. дои : 10.1086/409721 . ПМИД 857268 . S2CID 25675086 .
- ^ Холланд Б., Райс В.Р. (февраль 1998 г.). «Перспектива: преследование сексуального отбора: антагонистическое соблазнение против сопротивления» (PDF) . Эволюция; Международный журнал органической эволюции . 52 (1): 1–7. дои : 10.2307/2410914 . JSTOR 2410914 . ПМИД 28568154 . Архивировано из оригинала (PDF) 8 июня 2019 года . Проверено 8 июля 2016 г.
- ^ Клаттон-Брок, TH (май 1989 г.). «Системы спаривания млекопитающих» . Труды Лондонского королевского общества. Серия Б, Биологические науки . 236 (1285): 339–372. Бибкод : 1989РСПСБ.236..339С . дои : 10.1098/rspb.1989.0027 . ПМИД 2567517 . S2CID 84780662 .
- ^ Бонесс DJ, Боуэн Д., Бюлейер Б.М., Маршалл Дж.Дж. (2006). «Тактика спаривания и система спаривания водно-спаривающихся ластоногих: обыкновенный тюлень Phoca vitulina » . Поведенческая экология и социобиология . 61 : 119–130. дои : 10.1007/s00265-006-0242-9 . S2CID 25266746 .
- ^ Клопфер П.Х. (1981). «Истоки родительской заботы» . В Губернике DJ (ред.). Родительская забота у млекопитающих . Нью-Йорк: Пленум Пресс. ISBN 978-1-4613-3150-6 . OCLC 913709574 .
- ^ Мерти Р., Бирман Г., Браун С., Брайант К., Чинн Р., Хьюлетт А. и др. (май 2015 г.). «Животные в медицинских учреждениях: рекомендации по минимизации потенциальных рисков» (PDF) . Инфекционный контроль и госпитальная эпидемиология . 36 (5): 495–516. дои : 10.1017/ice.2015.15 . ПМИД 25998315 . S2CID 541760 . Архивировано (PDF) из оригинала 3 ноября 2023 года.
- ^ Гуманное общество США. «Статистика владения домашними животными в США» . Архивировано из оригинала 7 апреля 2012 года . Проверено 27 апреля 2012 г.
- ^ Министерство сельского хозяйства США. «Профиль кролиководческой отрасли США» (PDF) . Архивировано из оригинала (PDF) 7 августа 2019 года . Проверено 10 июля 2013 г.
- ^ Маккай Р. (26 мая 2013 г.). «Доисторическое наскальное искусство в Дордони» . Хранитель . Архивировано из оригинала 31 мая 2021 года . Проверено 9 ноября 2016 г.
- ^ Джонс Дж. (27 июня 2014 г.). «10 лучших портретов животных в искусстве» . Хранитель . Архивировано из оригинала 18 мая 2016 года . Проверено 24 июня 2016 г.
- ^ «Охота на оленей в Соединенных Штатах: анализ демографии и поведения охотников, дополнение к отчету о национальном обследовании рыболовства, охоты и отдыха, связанного с дикой природой, 2001–2006 гг.» . Служба рыболовства и дикой природы (США). Архивировано из оригинала 13 августа 2016 года . Проверено 24 июня 2016 г.
- ^ Шелтон Л. (5 апреля 2014 г.). «Рост популярности рекреационной охоты на свиней» . Натчезский демократ . Гранд-Вью на открытом воздухе. Архивировано из оригинала 12 декабря 2017 года . Проверено 24 июня 2016 г.
- ^ Нгуен Дж., Уитли Р. (2015). Охота за едой: Руководство по сбору урожая, полевой обработке и приготовлению дичи . Ф+В Медиа. стр. 6–77. ISBN 978-1-4403-3856-4 . Главы об охоте на оленя, кабана (кабана), кролика и белку.
- ^ «Скачки» . Британская энциклопедия . Архивировано из оригинала 21 декабря 2013 года . Проверено 6 мая 2014 г.
- ^ Гендеры Р (1981). Энциклопедия собачьих бегов . Книги Пелхэма. ISBN 978-0-7207-1106-6 . OCLC 9324926 .
- ^ Плюс С (1993). «Роль животных в человеческом обществе». Журнал социальных проблем . 49 (1): 1–9. дои : 10.1111/j.1540-4560.1993.tb00906.x .
- ^ Фаулер К.Дж. (26 марта 2014 г.). «10 лучших книг об умных животных» . Хранитель . Архивировано из оригинала 28 мая 2021 года . Проверено 9 ноября 2016 г.
- ^ Гэмбл Н., Йейтс С. (2008). Изучение детской литературы (2-е изд.). Лос-Анджелес: Сейдж. ISBN 978-1-4129-3013-0 . OCLC 71285210 .
- ^ «Книги для взрослых» . Натурщики тюленей . Архивировано из оригинала 11 июля 2016 года . Проверено 9 ноября 2016 г.
- ^ Патерсон Дж (2013). «Животные в кино и медиа». Оксфордские библиографии . дои : 10.1093/обо/9780199791286-0044 .
- ^ Джонс С. (2011). Крупный рогатый скот: история, миф, искусство . Лондон: Издательство Британского музея. ISBN 978-0-7141-5084-0 . OCLC 665137673 .
- ^ ван Гулик Р.Х. Хаягрива: Мантраянский аспект культа лошади в Китае и Японии . Архив Брилла. п. 9.
- ^ Грейнджер Р. (24 июня 2012 г.). «Изображение льва в древних и современных религиях» . ТРЕВОГА. Архивировано из оригинала 23 сентября 2016 года . Проверено 6 ноября 2016 г.
- ^ Даймонд Дж. М. (1997). «Часть 2: Возникновение и распространение производства продуктов питания» . Оружие, микробы и сталь: судьбы человеческих обществ . Нью-Йорк: WW Norton & Company. ISBN 978-0-393-03891-0 . ОСЛК 35792200 .
- ^ Ларсон Дж., Бургер Дж. (апрель 2013 г.). «Взгляд на приручение животных с точки зрения популяционной генетики» (PDF) . Тенденции в генетике . 29 (4): 197–205. дои : 10.1016/j.tig.2013.01.003 . ПМИД 23415592 . Архивировано из оригинала (PDF) 8 июня 2019 года . Проверено 9 ноября 2016 г.
- ^ Зедер М.А. (август 2008 г.). «Одомашнивание и раннее сельское хозяйство в Средиземноморском бассейне: происхождение, распространение и влияние» . Труды Национальной академии наук Соединенных Штатов Америки . 105 (33): 11597–11604. Бибкод : 2008PNAS..10511597Z . дои : 10.1073/pnas.0801317105 . ПМЦ 2575338 . ПМИД 18697943 .
- ^ «Графическая деталь. Графики, карты и инфографика. Подсчет кур» . Экономист . 27 июля 2011 года. Архивировано из оригинала 15 июля 2016 года . Проверено 6 ноября 2016 г.
- ^ «Породы крупного рогатого скота в КРС СЕГОДНЯ» . Крупный рогатый скот сегодня . Крупный рогатый скот-сегодня.com. Архивировано из оригинала 15 июля 2011 года . Проверено 6 ноября 2016 г.
- ^ Люкфар С.Д., Чик PR. «Стратегии развития кролиководческих проектов в системах натурального хозяйства» . Продовольственная и сельскохозяйственная организация . Архивировано из оригинала 6 мая 2016 года . Проверено 6 ноября 2016 г.
- ^ Пруд РГ (2004). Энциклопедия зоотехники . ЦРК Пресс. стр. 248–250. ISBN 978-0-8247-5496-9 . OCLC 57033325 . Архивировано из оригинала 23 января 2023 года . Проверено 5 октября 2018 г.
- ^ Бротен А.В. (2005). "Шерсть" . В Стиле V (ред.). Энциклопедия одежды и моды . Том. 3. Томсон Гейл . стр. 441–443 . ISBN 978-0-684-31394-8 . ОСЛК 963977000 .
- ^ Quiggle C (осень 2000 г.). «Альпака: древняя роскошь». Переплетение лиц : 74–76.
- ^ «Дикие млекопитающие составляют лишь несколько процентов млекопитающих мира» . Наш мир в данных . Проверено 8 августа 2023 г.
- ^ «Генетические исследования» . Фонд здоровья животных. Архивировано из оригинала 12 декабря 2017 года . Проверено 6 ноября 2016 г.
- ^ «Разработка лекарств» . Исследования животных.info. Архивировано из оригинала 8 июня 2016 года . Проверено 6 ноября 2016 г.
- ^ «Статистика ЕС показывает снижение количества исследований на животных» . Кстати об исследованиях. 2013. Архивировано из оригинала 24 апреля 2019 года . Проверено 6 ноября 2016 г.
- ^ Пилчер Х.Р. (2003). «Это нокаут» . Природа . дои : 10.1038/news030512-17 . Архивировано из оригинала 10 ноября 2016 года . Проверено 6 ноября 2016 г.
- ^ «Поставки и использование приматов в ЕС» . Европейская ассоциация биомедицинских исследований. 1996. Архивировано из оригинала 17 января 2012 года.
- ^ Карлссон Х.Э., Шапиро С.Дж., Фарах И., Хау Дж. (август 2004 г.). «Использование приматов в исследованиях: глобальный обзор». Американский журнал приматологии . 63 (4): 225–237. дои : 10.1002/ajp.20054 . ПМИД 15300710 . S2CID 41368228 .
- ^ Уэзералл Д. и др. (2006). Использование приматов в исследованиях (PDF) (Отчет). Лондон: Академия медицинских наук. Архивировано из оригинала (PDF) 23 марта 2013 года.
- ^ Ричи Х., Розер М. (15 апреля 2021 г.). «Биоразнообразие» . Наш мир в данных . Архивировано из оригинала 11 декабря 2022 года . Проверено 29 августа 2021 г.
- ^ Jump up to: Перейти обратно: а б Бар-Он Ю.М., Филлипс Р., Майло Р. (июнь 2018 г.). «Распределение биомассы на Земле» . Труды Национальной академии наук Соединенных Штатов Америки . 115 (25): 6506–6511. Бибкод : 2018PNAS..115.6506B . дои : 10.1073/pnas.1711842115 . ПМК 6016768 . ПМИД 29784790 .
- ^ Прайс Е (2008). Принципы и применение поведения домашних животных: вводный текст . Сакраменто: Издательство Кембриджского университета. ISBN 978-1-84593-398-2 . ОСЛК 226038028 .
- ^ Таупитц Дж., Весшка М. (2009). Химбриды – химеры и гибриды в сравнительных европейских и международных исследованиях . Гейдельберг: Спрингер. п. 13. ISBN 978-3-540-93869-9 . OCLC 495479133 .
- ^ Чемберс С.М., Фейн С.Р., Фацио Б., Амарал М. (2012). «Отчет о систематике североамериканских волков на основе морфологического и генетического анализа» . Североамериканская фауна . 77 :2. дои : 10.3996/нафа.77.0001 . Архивировано из оригинала 31 мая 2021 года . Проверено 12 октября 2019 г.
- ^ ван Вюр Т (2005). По следам зубра – история, морфология и экология вымершего дикого быка . Издательство Пенсофт. ISBN 978-954-642-235-4 . OCLC 940879282 .
- ^ Муни Х.А., Клеланд Э.Э. (май 2001 г.). «Эволюционное влияние инвазивных видов» . Труды Национальной академии наук Соединенных Штатов Америки . 98 (10): 5446–5451. Бибкод : 2001PNAS...98.5446M . дои : 10.1073/pnas.091093398 . ПМЦ 33232 . ПМИД 11344292 .
- ^ Ле Ру Дж.Дж., Фокскрофт Л.К., Хербст М., Макфадьен С. (январь 2015 г.). «Генетический анализ показывает низкий уровень гибридизации между африканскими дикими кошками (Felis silvestris lybica) и домашними кошками (F. s. catus) в Южной Африке» . Экология и эволюция . 5 (2): 288–299. Бибкод : 2015EcoEv...5..288L . дои : 10.1002/ece3.1275 . ПМК 4314262 . ПМИД 25691958 .
- ^ Уилсон А. (2003). Отчет о состоянии лесов Австралии . п. 107.
- ^ Раймер Дж. М., Симберлофф Д. (ноябрь 1996 г.). «Вымирание путем гибридизации и интрогрессии». Ежегодный обзор экологии и систематики . 27 : 83–109. doi : 10.1146/annurev.ecolsys.27.1.83 .
- ^ Поттс Б.М. (2001). Барбур Р.К., Хингстон AB (ред.). Генетическое загрязнение в результате фермерского лесного хозяйства с использованием видов и гибридов эвкалипта: отчет для программы совместного предприятия RIRDC/L&WA/FWPRDC по агролесомелиорации . Корпорация сельских промышленных исследований и развития Австралии. ISBN 978-0-642-58336-9 . OCLC 48794104 .
- ^ Jump up to: Перейти обратно: а б Дирзо Р., Янг Х.С., Галетти М., Себальос Г., Исаак Н.Дж., Коллен Б. (июль 2014 г.). «Дефаунация в антропоцене» (PDF) . Наука . 345 (6195): 401–406. Бибкод : 2014Sci...345..401D . дои : 10.1126/science.1251817 . ПМИД 25061202 . S2CID 206555761 . Архивировано (PDF) из оригинала 7 августа 2019 года . Проверено 25 января 2024 г.
- ^ Примак Р. (2014). Основы биологии сохранения (6-е изд.). Сандерленд, Массачусетс: Издательство Sinauer Associates, Inc. стр. 217–245. ISBN 978-1-60535-289-3 . OCLC 876140621 .
- ^ Виньери С (июль 2014 г.). «Исчезающая фауна. Введение» . Наука . 345 (6195): 392–395. Бибкод : 2014Sci...345..392V . дои : 10.1126/science.345.6195.392 . ПМИД 25061199 .
- ^ Берни Д.А., Фланнери Т.Ф. (июль 2005 г.). «Пятьдесят тысячелетий катастрофических вымираний после контакта с человеком» (PDF) . Тенденции в экологии и эволюции . 20 (7): 395–401. дои : 10.1016/j.tree.2005.04.022 . ПМИД 16701402 . Архивировано из оригинала (PDF) 10 июня 2010 года.
- ^ Даймонд Дж (1984). «Исторические вымирания: Розеттский камень для понимания доисторических вымираний». Мартин П.С., Кляйн Р.Г. (ред.). Четвертичные вымирания: доисторическая революция . Тусон: Издательство Университета Аризоны. стр. 824–862. ISBN 978-0-8165-1100-6 . ОСЛК 10301944 .
- ^ Уоттс Дж. (6 мая 2019 г.). «Человеческое общество находится под серьезной угрозой утраты естественной жизни на Земле» . Хранитель . Архивировано из оригинала 14 июня 2019 года . Проверено 1 июля 2019 г.
- ^ МакГрат М. (6 мая 2019 г.). «Природный кризис: люди угрожают исчезновением миллиону видов » . Би-би-си . Архивировано из оригинала 30 июня 2019 года . Проверено 1 июля 2019 г.
- ^ Главный D (22 ноября 2013 г.). «7 знаковых животных, которых люди ведут к вымиранию» . Живая наука . Архивировано из оригинала 6 января 2023 года . Проверено 25 января 2024 г.
- ^ Платт-младший (25 октября 2011 г.). «Браконьеры привели к вымиранию яванского носорога во Вьетнаме» . Научный американец . Архивировано из оригинала 6 апреля 2015 года.
- ^ Кэррингтон Д. (8 декабря 2016 г.). «Жирафам грозит исчезновение после разрушительного сокращения, предупреждают эксперты» . Хранитель . Архивировано из оригинала 13 августа 2021 года . Проверено 4 февраля 2017 г.
- ^ Эстрада А., Гарбер П.А., Райландс А.Б., Роос С., Фернандес-Дюке Е., Ди Фиоре А. и др. (январь 2017 г.). «Надвигающийся кризис вымирания приматов в мире: почему приматы имеют значение» . Достижения науки . 3 (1): e1600946. Бибкод : 2017SciA....3E0946E . дои : 10.1126/sciadv.1600946 . ПМЦ 5242557 . ПМИД 28116351 .
- ^ Флетчер М. (31 января 2015 г.). «Панголины: почему это милое доисторическое млекопитающее находится на грани исчезновения» . Телеграф . Архивировано из оригинала 10 января 2022 года.
- ^ Гринфилд П. (9 сентября 2020 г.). «Люди эксплуатируют и уничтожают природу в беспрецедентных масштабах – доклад» . Хранитель . Архивировано из оригинала 21 октября 2021 года . Проверено 13 октября 2020 г.
- ^ Маккарти Д. (1 октября 2020 г.). «Ужасающие потери дикой природы показывают, что конец игры по вымиранию начался, но еще не поздно измениться» . Независимый . Архивировано из оригинала 7 апреля 2023 года . Проверено 13 октября 2020 г.
- ^ Пенниси Э (18 октября 2016 г.). «Люди истребляют приматов, летучих мышей и других млекопитающих» . Наука . Архивировано из оригинала 20 октября 2021 года . Проверено 3 февраля 2017 г.
- ^ Риппл У.Дж., Абернети К., Беттс М.Г., Чапрон Г., Дирзо Р., Галетти М. и др. (октябрь 2016 г.). «Охота на мясо диких животных и риск исчезновения млекопитающих мира» . Королевское общество открытой науки . 3 (10): 160498. Бибкод : 2016RSOS....360498R . дои : 10.1098/rsos.160498 . hdl : 1893/24446 . ПМК 5098989 . ПМИД 27853564 .
- ^ Уильямс М., Заласевич Дж., Хафф П.К., Швегерль С., Барноски А.Д. , Эллис ЕС (2015). «Антропоценовая биосфера». Обзор антропоцена . 2 (3): 196–219. Бибкод : 2015AntRv...2..196W . дои : 10.1177/2053019615591020 . S2CID 7771527 .
- ^ Морелл В. (11 августа 2015 г.). «Мясоеды могут ускорить исчезновение видов во всем мире, предупреждает исследование» . Наука . Архивировано из оригинала 20 декабря 2016 года . Проверено 3 февраля 2017 г.
- ^ Маховина Б., Фили К.Дж., Риппл У.Дж. (декабрь 2015 г.). «Сохранение биоразнообразия: главное – сокращение потребления мяса». Наука об общей окружающей среде . 536 : 419–431. Бибкод : 2015ScTEn.536..419M . doi : 10.1016/j.scitotenv.2015.07.022 . ПМИД 26231772 .
- ^ Редфорд К.Х. (1992). «Пустой лес» (PDF) . Бионаука . 42 (6): 412–422. дои : 10.2307/1311860 . JSTOR 1311860 . Архивировано (PDF) оригинала 28 февраля 2021 года . Проверено 4 февраля 2017 г.
- ^ Перес К.А., Насименто Х.С. (2006). «Влияние охоты на дичь каяпо на юго-востоке Амазонии: последствия для сохранения дикой природы в местных заповедниках тропических лесов». Человеческая эксплуатация и сохранение биоразнообразия . Том. 3. Спрингер. стр. 287–313. ISBN 978-1-4020-5283-5 . OCLC 207259298 .
- ^ Альтрихтер М., Боальо Г. (2004). «Распространение и относительная численность пекари в аргентинском Чако: связь с человеческим фактором». Биологическая консервация . 116 (2): 217–225. Бибкод : 2004BCons.116..217A . дои : 10.1016/S0006-3207(03)00192-7 .
- ^ Гобуш К. «Влияние браконьерства на африканских слонов» . Центр биологии сохранения . Университет Вашингтона. Архивировано из оригинала 8 декабря 2021 года . Проверено 12 мая 2021 г.
- ^ Алверсон Д.Л., Фрибург М.Х., Муравски С.А., Поуп Дж.Г. (1996) [1994]. «Прилов морских млекопитающих» . Глобальная оценка рыболовного прилова и выбросов . Рим: Продовольственная и сельскохозяйственная организация Объединенных Наций. ISBN 978-92-5-103555-9 . ОСЛК 31424005 . Архивировано из оригинала 17 февраля 2019 года . Проверено 25 января 2024 г.
- ^ Гловка Л., Бурхенне-Гильмин Ф., Synge HM, Макнили Дж.А., Гюндлинг Л. (1994). Документ по экологической политике и законодательству МСОП . Путеводитель по Конвенции о биоразнообразии. Международный союз охраны природы. ISBN 978-2-8317-0222-3 . ОСЛК 32201845 .
- ^ «О МСОП» . Международный союз охраны природы. 3 декабря 2014 г. Архивировано из оригинала 15 апреля 2020 г. Проверено 3 февраля 2017 г.
- ^ Себальос Г., Эрлих П.Р., Барноски А.Д., Гарсиа А., Прингл Р.М., Палмер Т.М. (июнь 2015 г.). «Ускоренная антропогенная гибель видов в современном мире: на пороге шестого массового вымирания» . Достижения науки . 1 (5): e1400253. Бибкод : 2015SciA....1E0253C . дои : 10.1126/sciadv.1400253 . ПМК 4640606 . ПМИД 26601195 .
- ^ Фишер Д.О., Бломберг С.П. (апрель 2011 г.). «Корреляты повторного открытия и возможность обнаружения вымирания млекопитающих» . Слушания. Биологические науки . 278 (1708): 1090–1097. дои : 10.1098/rspb.2010.1579 . ПМК 3049027 . ПМИД 20880890 .
- ^ Себальос Г., Эрлих А.Х., Эрлих П.Р. (2015). Уничтожение природы: вымирание птиц и млекопитающих человеком . Балтимор: Издательство Университета Джонса Хопкинса. п. 69. ИСБН 978-1-4214-1718-9 .
- ^ Цзян, З.; Харрис, РБ (2016). « Элафурус Давидианус » . Красный список исчезающих видов МСОП . 2016 : e.T7121A22159785. doi : 10.2305/IUCN.UK.2016-2.RLTS.T7121A22159785.en . Проверено 12 ноября 2021 г.
- ^ Jump up to: Перейти обратно: а б МакКинни М.Л., Шох Р., Йонавьяк Л. (2013). «Сохранение биологических ресурсов» . Наука об окружающей среде: системы и решения (5-е изд.). Джонс и Бартлетт Обучение. ISBN 978-1-4496-6139-7 . OCLC 777948078 .
- ^ Перрин В.Ф., Вюрсиг Б.Ф., Тьювиссен Дж.Г. (2009). Энциклопедия морских млекопитающих . Академическая пресса. п. 404. ИСБН 978-0-12-373553-9 . OCLC 455328678 .
- На момент редактирования в этой статье используется контент из раздела «Анатомия и физиология» , который лицензируется таким образом, чтобы его можно было повторно использовать по лицензии Creative Commons Attribution-ShareAlike 3.0 Unported License , но не по GFDL . Все соответствующие условия должны быть соблюдены.
Дальнейшее чтение
- Браун ВМ (2001). «Естественный отбор компонентов мозга млекопитающих». Тенденции экологии и эволюции . 16 (9): 471–473. Бибкод : 2001TEcoE..16..471B . дои : 10.1016/S0169-5347(01)02246-7 .
- Маккенна MC, Bell SK (1997). Классификация млекопитающих выше видового уровня . Нью-Йорк: Издательство Колумбийского университета. ISBN 978-0-231-11013-6 . OCLC 37345734 . [ постоянная мертвая ссылка ]
- Новак Р.М. (1999). Млекопитающие мира Уокера (6-е изд.). Балтимор: Издательство Университета Джонса Хопкинса. ISBN 978-0-8018-5789-8 . OCLC 937619124 .
- Симпсон Г.Г. (1945). «Принципы классификации и классификация млекопитающих». Бюллетень Американского музея естественной истории . 85 : 1–350.
- Мерфи В.Дж., Эйзирик Э., О'Брайен С.Дж., Мэдсен О., Скалли М., Дуади С.Дж. и др. (декабрь 2001 г.). «Разрешение ранней плацентарной радиации млекопитающих с использованием байесовской филогенетики». Наука . 294 (5550): 2348–2351. Бибкод : 2001Sci...294.2348M . дои : 10.1126/science.1067179 . ПМИД 11743200 . S2CID 34367609 .
- Спрингер М.С., Стэнхоуп М.Дж., Мэдсен О., де Йонг В.В. (август 2004 г.). «Молекулы консолидируют плацентарное дерево млекопитающих» (PDF) . Тенденции в экологии и эволюции . 19 (8): 430–438. дои : 10.1016/j.tree.2004.05.006 . ПМИД 16701301 . S2CID 1508898 . Архивировано из оригинала (PDF) 29 июля 2016 года . Проверено 21 января 2005 г.
- Воган Т.А., Райан Дж.М., Капзаплевски, Нью-Джерси (2000). Маммология (4-е изд.). Форт-Уэрт, Техас: Издательство Saunders College. ISBN 978-0-03-025034-7 . OCLC 42285340 .
- Кригс Дж.О., Чураков Г., Кифманн М., Джордан У., Брозиус Дж., Шмитц Дж. (апрель 2006 г.). «Ретропозированные элементы как архивы эволюционной истории плацентарных млекопитающих» . ПЛОС Биология . 4 (4): е91. doi : 10.1371/journal.pbio.0040091 . ПМЦ 1395351 . ПМИД 16515367 .
- Макдональд Д.В., Норрис С. (2006). Энциклопедия млекопитающих (3-е изд.). Лондон: Справочная группа Брауна. ISBN 978-0-681-45659-4 . OCLC 74900519 .
Внешние ссылки
- База данных ASM по разнообразию млекопитающих. Архивировано 25 декабря 2022 года на Wayback Machine.
- Biodiversitymapping.org - Все отряды млекопитающих в мире с картами распространения. Архивировано 26 сентября 2016 г. в Wayback Machine.
- Палеоценовые млекопитающие. Архивировано 3 февраля 2024 года на Wayback Machine , сайте, освещающем возникновение млекопитающих, paleocene-mammals.de.
- Эволюция млекопитающих. Архивировано 25 января 2024 года в Wayback Machine , краткое введение в ранних млекопитающих, enchantedlearning.com.
- Европейский атлас млекопитающих EMMA. Архивировано 25 января 2024 г. в Wayback Machine от Societas Europaea Mammalogica, European-mammals.org.
- Морские млекопитающие мира. Архивировано 8 июня 2019 года в Wayback Machine . Обзор всех морских млекопитающих, включая описания, как полностью водных, так и полуводных, noaa.gov.
- Mammalogy.org. Архивировано 1 марта 2020 года в Wayback Machine. Американское общество маммологов было основано в 1919 году с целью содействия изучению млекопитающих, и этот веб-сайт включает библиотеку изображений млекопитающих.










