Слон
| Слоны Временной диапазон: | |
|---|---|
 | |
| A female African bush elephant in Mikumi National Park, Tanzania | |
| Scientific classification | |
| Domain: | Eukaryota |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Class: | Mammalia |
| Order: | Proboscidea |
| Superfamily: | Elephantoidea |
| Family: | Elephantidae |
| Groups included | |
| |
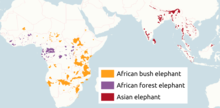 | |
| Distribution of living elephant species | |
| Cladistically included but traditionally excluded taxa | |
| |
Слоны — самые крупные из ныне живущих наземных животных. три живых вида В настоящее время известно : африканский кустарниковый слон ( Loxodonta africana ), африканский лесной слон ( L. cycltis ) и азиатский слон ( Elephas maximus ). Это единственные сохранившиеся представители семейства Elephantidae и отряда Proboscidea ; вымершие родственники включают мамонтов и мастодонтов . Отличительные особенности слонов включают длинный хобот , называемый хоботом, бивни , большие ушные раковины, столбообразные ноги и жесткую, но чувствительную серую кожу. Хобот цепкий , принося ко рту пищу и воду и хватая предметы. Бивни, происходящие от резцов, служат одновременно оружием и инструментами для перемещения предметов и рытья. Большие ушные раковины помогают поддерживать постоянную температуру тела, а также общаться. У африканских слонов уши больше и вогнутая спина, тогда как у азиатских слонов уши меньше, а спина выпуклая или ровная.
Слоны разбросаны по всей Африке к югу от Сахары , Южной Азии и Юго-Восточной Азии и встречаются в разных местах обитания, включая саванны , леса, пустыни и болота . Они травоядны и держатся возле воды, когда она доступна. Они считаются ключевыми видами из-за их воздействия на окружающую среду. У слонов существует общество деления-слияния , в котором несколько семейных групп собираются вместе для общения. Самки (коровы), как правило, живут семейными группами, которые могут состоять из одной самки с телятами или нескольких родственных самок с потомством. Лидер женской группы, обычно самая старая корова, известен как матриарх .
Males (bulls) leave their family groups when they reach puberty and may live alone or with other males. Adult bulls mostly interact with family groups when looking for a mate. They enter a state of increased testosterone and aggression known as musth, which helps them gain dominance over other males as well as reproductive success. Calves are the centre of attention in their family groups and rely on their mothers for as long as three years. Elephants can live up to 70 years in the wild. They communicate by touch, sight, smell, and sound; elephants use infrasound and seismic communication over long distances. Elephant intelligence has been compared with that of primates and cetaceans. They appear to have self-awareness, and possibly show concern for dying and dead individuals of their kind.
African bush elephants and Asian elephants are listed as endangered and African forest elephants as critically endangered by the International Union for Conservation of Nature (IUCN). One of the biggest threats to elephant populations is the ivory trade, as the animals are poached for their ivory tusks. Other threats to wild elephants include habitat destruction and conflicts with local people. Elephants are used as working animals in Asia. In the past, they were used in war; today, they are often controversially put on display in zoos, or employed for entertainment in circuses. Elephants have an iconic status in human culture and have been widely featured in art, folklore, religion, literature, and popular culture.
Etymology
The word elephant is derived from the Latin word elephas (genitive elephantis) 'elephant', which is the Latinised form of the ancient Greek ἐλέφας (elephas) (genitive ἐλέφαντος (elephantos[1])), probably from a non-Indo-European language, likely Phoenician.[2] It is attested in Mycenaean Greek as e-re-pa (genitive e-re-pa-to) in Linear B syllabic script.[3][4] As in Mycenaean Greek, Homer used the Greek word to mean ivory, but after the time of Herodotus, it also referred to the animal.[1] The word elephant appears in Middle English as olyfaunt (c. 1300) and was borrowed from Old French oliphant (12th century).[2]
Taxonomy
| A cladogram of the elephants within Afrotheria based on molecular evidence[5] |
Elephants belong to the family Elephantidae, the sole remaining family within the order Proboscidea. Their closest extant relatives are the sirenians (dugongs and manatees) and the hyraxes, with which they share the clade Paenungulata within the superorder Afrotheria.[6] Elephants and sirenians are further grouped in the clade Tethytheria.[7]
Three species of living elephants are recognised; the African bush elephant (Loxodonta africana), forest elephant (Loxodonta cyclotis), and Asian elephant (Elephas maximus).[8] African elephants were traditionally considered a single species, Loxodonta africana, but molecular studies have affirmed their status as separate species.[9][10][11] Mammoths (Mammuthus) are nested within living elephants as they are more closely related to Asian elephants than to African elephants.[12] Another extinct genus of elephant, Palaeoloxodon, is also recognised, which appears to have close affinities with African elephants and to have hybridised with African forest elephants.[13]
Evolution
Over 180 extinct members of order Proboscidea have been described.[14] The earliest proboscideans, the African Eritherium and Phosphatherium are known from the late Paleocene.[15] The Eocene included Numidotherium, Moeritherium, and Barytherium from Africa. These animals were relatively small and, some, like Moeritherium and Barytherium were probably amphibious.[16][17] Later on, genera such as Phiomia and Palaeomastodon arose; the latter likely inhabited more forested areas. Proboscidean diversification changed little during the Oligocene.[16] One notable species of this epoch was Eritreum melakeghebrekristosi of the Horn of Africa, which may have been an ancestor to several later species.[18]
| Proboscidea phylogeny based on morphological and DNA evidence[19][20][13] |



A major event in proboscidean evolution was the collision of Afro-Arabia with Eurasia, during the Early Miocene, around 18–19 million years ago, allowing proboscideans to disperse from their African homeland across Eurasia and later, around 16–15 million years ago into North America across the Bering Land Bridge. Proboscidean groups prominent during the Miocene include the deinotheres, along with the more advanced elephantimorphs, including mammutids (mastodons), gomphotheres, amebelodontids (which includes the "shovel tuskers" like Platybelodon), choerolophodontids and stegodontids.[21] Around 10 million years ago, the earliest members of the family Elephantidae emerged in Africa, having originated from gomphotheres.[22]
Elephantids are distinguished from earlier proboscideans by a major shift in the molar morphology to parallel lophs rather than the cusps of earlier proboscideans, allowing them to become higher-crowned (hypsodont) and more efficient in consuming grass.[23] The Late Miocene saw major climactic changes, which resulted in the decline and extinction of many proboscidean groups.[21] The earliest members of the modern genera of Elephantidae appeared during the latest Miocene–early Pliocene around 5 million years ago. The elephantid genera Elephas (which includes the living Asian elephant) and Mammuthus (mammoths) migrated out of Africa during the late Pliocene, around 3.6 to 3.2 million years ago.[24]
Over the course of the Early Pleistocene, all non-elephantid probobscidean genera outside of the Americas became extinct with the exception of Stegodon,[21] with gomphotheres dispersing into South America as part of the Great American interchange,[25] and mammoths migrating into North America around 1.5 million years ago.[26] At the end of the Early Pleistocene, around 800,000 years ago the elephantid genus Palaeoloxodon dispersed outside of Africa, becoming widely distributed in Eurasia.[27] Proboscideans were represented by around 23 species at the beginning of the Late Pleistocene. Proboscideans underwent a dramatic decline during the Late Pleistocene as part of the Late Pleistocene extinctions of most large mammals globally, with all remaining non-elephantid proboscideans (including Stegodon, mastodons, and the American gomphotheres Cuvieronius and Notiomastodon) and Palaeoloxodon becoming extinct, with mammoths only surviving in relict populations on islands around the Bering Strait into the Holocene, with their latest survival being on Wrangel Island, where they persisted until around 4,000 years ago.[21][28]
Over the course of their evolution, probobscideans grew in size. With that came longer limbs and wider feet with a more digitigrade stance, along with a larger head and shorter neck. The trunk evolved and grew longer to provide reach. The number of premolars, incisors, and canines decreased, and the cheek teeth (molars and premolars) became longer and more specialised. The incisors developed into tusks of different shapes and sizes.[29] Several species of proboscideans became isolated on islands and experienced insular dwarfism,[30] some dramatically reducing in body size, such as the 1 m (3 ft 3 in) tall dwarf elephant species Palaeoloxodon falconeri.[31]
Living species
| Name | Size | Appearance | Distribution | Image |
|---|---|---|---|---|
| African bush elephant (Loxodonta africana) | Male: 304–336 cm (10 ft 0 in – 11 ft 0 in) (shoulder height), 5.2–6.9 t (5.7–7.6 short tons) (weight); Female: 247–273 cm (8 ft 1 in – 8 ft 11 in) (shoulder height), 2.6–3.5 t (2.9–3.9 short tons) (weight).[32] | Relatively large and triangular ears, concave back, diamond shaped molar ridges, wrinkled skin, sloping abdomen, and two finger-like extensions at the tip of the trunk.[33] | Sub-Saharan Africa; forests, savannahs, deserts, wetlands, and near lakes[34] |  |
| African forest elephant (Loxodonta cyclotis) | 209–231 cm (6 ft 10 in – 7 ft 7 in) (shoulder height), 1.7–2.3 t (1.9–2.5 short tons) (weight).[32] | Similar to the bush species, but with smaller and more rounded ears and thinner and straighter tusks.[33][34] | West and Central Africa; equatorial forests, but occasionally gallery forests and forest/grassland ecotones.[34] |  |
| Asian elephant (Elephas maximus) | Male: 261–289 cm (8 ft 7 in – 9 ft 6 in) (shoulder height), 3.5–4.6 t (3.9–5.1 short tons) (weight); Female: 228–252 cm (7 ft 6 in – 8 ft 3 in) (shoulder height), 2.3–3.1 t (2.5–3.4 short tons) (weight).[32] | Relatively small ears, convex or level back, dish-shaped forehead with two large bumps, narrow molar ridges, smooth skin with some blotches of depigmentation, a straightened or saggy abdomen, and one extension at the tip of the trunk.[33] | South and Southeast Asia; habitats with a mix of grasses, low woody plants, and trees, including dry thorn-scrub forests in southern India and Sri Lanka and evergreen forests in Malaya.[35] |  |
Anatomy

Elephants are the largest living terrestrial animals.[36] The skeleton is made up of 326–351 bones.[37] The vertebrae are connected by tight joints, which limit the backbone's flexibility. African elephants have 21 pairs of ribs, while Asian elephants have 19 or 20 pairs.[38] The skull contains air cavities (sinuses) that reduce the weight of the skull while maintaining overall strength. These cavities give the inside of the skull a honeycomb-like appearance. By contrast, the lower jaw is dense. The cranium is particularly large and provides enough room for the attachment of muscles to support the entire head.[37] The skull is built to withstand great stress, particularly when fighting or using the tusks. The brain is surrounded by arches in the skull, which serve as protection.[39] Because of the size of the head, the neck is relatively short to provide better support.[29]Elephants are homeotherms and maintain their average body temperature at ~ 36 °C (97 °F), with a minimum of 35.2 °C (95.4 °F) during the cool season, and a maximum of 38.0 °C (100.4 °F) during the hot dry season.[40]
Ears and eyes

Elephant ear flaps, or pinnae, are 1–2 mm (0.039–0.079 in) thick in the middle with a thinner tip and supported by a thicker base. They contain numerous blood vessels called capillaries. Warm blood flows into the capillaries, releasing excess heat into the environment. This effect is increased by flapping the ears back and forth. Larger ear surfaces contain more capillaries, and more heat can be released. Of all the elephants, African bush elephants live in the hottest climates and have the largest ear flaps.[37][41] The ossicles are adapted for hearing low frequencies, being most sensitive at 1 kHz.[42]
Lacking a lacrimal apparatus (tear duct), the eye relies on the harderian gland in the orbit to keep it moist. A durable nictitating membrane shields the globe. The animal's field of vision is compromised by the location and limited mobility of the eyes.[43] Elephants are dichromats[44] and they can see well in dim light but not in bright light.[45]
Trunk

The elongated and prehensile trunk, or proboscis, consists of both the nose and upper lip, which fuse in early fetal development.[29] This versatile appendage contains up to 150,000 separate muscle fascicles, with no bone and little fat. These paired muscles consist of two major types: superficial (surface) and internal. The former are divided into dorsal, ventral, and lateral muscles, while the latter are divided into transverse and radiating muscles. The muscles of the trunk connect to a bony opening in the skull. The nasal septum consists of small elastic muscles between the nostrils, which are divided by cartilage at the base.[46] A unique proboscis nerve – a combination of the maxillary and facial nerves – lines each side of the appendage.[47]
As a muscular hydrostat, the trunk moves through finely controlled muscle contractions, working both with and against each other.[47] Using three basic movements: bending, twisting, and longitudinal stretching or retracting, the trunk has near unlimited flexibility. Objects grasped by the end of the trunk can be moved to the mouth by curving the appendage inward. The trunk can also bend at different points by creating stiffened "pseudo-joints". The tip can be moved in a way similar to the human hand.[48] The skin is more elastic on the dorsal side of the elephant trunk than underneath; allowing the animal to stretch and coil while maintaining a strong grasp.[49] The African elephants have two finger-like extensions at the tip of the trunk that allow them to pluck small food. The Asian elephant has only one and relies more on wrapping around a food item.[33] Asian elephant trunks have better motor coordination.[46]

The trunk's extreme flexibility allows it to forage and wrestle other elephants with it. It is powerful enough to lift up to 350 kg (770 lb), but it also has the precision to crack a peanut shell without breaking the seed. With its trunk, an elephant can reach items up to 7 m (23 ft) high and dig for water in the mud or sand below. It also uses it to clean itself.[50] Individuals may show lateral preference when grasping with their trunks: some prefer to twist them to the left, others to the right.[47] Elephant trunks are capable of powerful siphoning. They can expand their nostrils by 30%, leading to a 64% greater nasal volume, and can breathe in almost 30 times faster than a human sneeze, at over 150 m/s (490 ft/s).[51] They suck up water, which is squirted into the mouth or over the body.[29][51] The trunk of an adult Asian elephant is capable of retaining 8.5 L (2.2 US gal) of water.[46] They will also sprinkle dust or grass on themselves.[29] When underwater, the elephant uses its trunk as a snorkel.[52]
The trunk also acts as a sense organ. Its sense of smell may be four times greater than a bloodhound's nose.[53] The infraorbital nerve, which makes the trunk sensitive to touch, is thicker than both the optic and auditory nerves. Whiskers grow all along the trunk, and are particularly packed at the tip, where they contribute to its tactile sensitivity. Unlike those of many mammals, such as cats and rats, elephant whiskers do not move independently ("whisk") to sense the environment; the trunk itself must move to bring the whiskers into contact with nearby objects. Whiskers grow in rows along each side on the ventral surface of the trunk, which is thought to be essential in helping elephants balance objects there, whereas they are more evenly arranged on the dorsal surface. The number and patterns of whiskers are distinctly different between species.[54]
Damaging the trunk would be detrimental to an elephant's survival,[29] although in rare cases, individuals have survived with shortened ones. One trunkless elephant has been observed to graze using its lips with its hind legs in the air and balancing on its front knees.[46] Floppy trunk syndrome is a condition of trunk paralysis recorded in African bush elephants and involves the degeneration of the peripheral nerves and muscles. The disorder has been linked to lead poisoning.[55]
Teeth


Elephants usually have 26 teeth: the incisors, known as the tusks; 12 deciduous premolars; and 12 molars. Unlike most mammals, teeth are not replaced by new ones emerging from the jaws vertically. Instead, new teeth start at the back of the mouth and push out the old ones. The first chewing tooth on each side of the jaw falls out when the elephant is two to three years old. This is followed by four more tooth replacements at the ages of four to six, 9–15, 18–28, and finally in their early 40s. The final (usually sixth) set must last the elephant the rest of its life. Elephant teeth have loop-shaped dental ridges, which are more diamond-shaped in African elephants.[56]
Tusks
The tusks of an elephant are modified second incisors in the upper jaw. They replace deciduous milk teeth at 6–12 months of age and keep growing at about 17 cm (7 in) a year. As the tusk develops, it is topped with smooth, cone-shaped enamel that eventually wanes. The dentine is known as ivory and has a cross-section of intersecting lines, known as "engine turning", which create diamond-shaped patterns. Being living tissue, tusks are fairly soft and about as dense as the mineral calcite. The tusk protrudes from a socket in the skull, and most of it is external. At least one-third of the tusk contains the pulp, and some have nerves that stretch even further. Thus, it would be difficult to remove it without harming the animal. When removed, ivory will dry up and crack if not kept cool and wet. Tusks function in digging, debarking, marking, moving objects, and fighting.[57]
Elephants are usually right- or left-tusked, similar to humans, who are typically right- or left-handed. The dominant, or "master" tusk, is typically more worn down, as it is shorter and blunter. For African elephants, tusks are present in both males and females and are around the same length in both sexes, reaching up to 300 cm (9 ft 10 in),[57] but those of males tend to be more massive.[58] In the Asian species, only the males have large tusks. Female Asians have very small tusks, or none at all.[57] Tuskless males exist and are particularly common among Sri Lankan elephants.[59] Asian males can have tusks as long as Africans', but they are usually slimmer and lighter; the largest recorded was 302 cm (9 ft 11 in) long and weighed 39 kg (86 lb). Hunting for elephant ivory in Africa[60] and Asia[61] has led to natural selection for shorter tusks[62][63] and tusklessness.[64][65]
Skin

An elephant's skin is generally very tough, at 2.5 cm (1 in) thick on the back and parts of the head. The skin around the mouth, anus, and inside of the ear is considerably thinner. Elephants are typically grey, but African elephants look brown or reddish after rolling in coloured mud. Asian elephants have some patches of depigmentation, particularly on the head. Calves have brownish or reddish hair, with the head and back being particularly hairy. As elephants mature, their hair darkens and becomes sparser, but dense concentrations of hair and bristles remain on the tip of the tail and parts of the head and genitals. Normally, the skin of an Asian elephant is covered with more hair than its African counterpart.[66] Their hair is thought to help them lose heat in their hot environments.[67]
Although tough, an elephant's skin is very sensitive and requires mud baths to maintain moisture and protection from burning and insect bites. After bathing, the elephant will usually use its trunk to blow dust onto its body, which dries into a protective crust. Elephants have difficulty releasing heat through the skin because of their low surface-area-to-volume ratio, which is many times smaller than that of a human. They have even been observed lifting up their legs to expose their soles to the air.[66] Elephants only have sweat glands between the toes,[68] but the skin allows water to disperse and evaporate, cooling the animal.[69][70] In addition, cracks in the skin may reduce dehydration and allow for increased thermal regulation in the long term.[71]
Legs, locomotion, and posture


To support the animal's weight, an elephant's limbs are positioned more vertically under the body than in most other mammals. The long bones of the limbs have cancellous bones in place of medullary cavities. This strengthens the bones while still allowing haematopoiesis (blood cell creation).[72] Both the front and hind limbs can support an elephant's weight, although 60% is borne by the front.[73] The position of the limbs and leg bones allows an elephant to stand still for extended periods of time without tiring. Elephants are incapable of turning their manus as the ulna and radius of the front legs are secured in pronation.[72] Elephants may also lack the pronator quadratus and pronator teres muscles or have very small ones.[74] The circular feet of an elephant have soft tissues, or "cushion pads" beneath the manus or pes, which allow them to bear the animal's great mass.[73] They appear to have a sesamoid, an extra "toe" similar in placement to a giant panda's extra "thumb", that also helps in weight distribution.[75] As many as five toenails can be found on both the front and hind feet.[33]
Elephants can move both forward and backward, but are incapable of trotting, jumping, or galloping. They can move on land only by walking or ambling: a faster gait similar to running.[72][76] In walking, the legs act as pendulums, with the hips and shoulders moving up and down while the foot is planted on the ground. The fast gait does not meet all the criteria of running, since there is no point where all the feet are off the ground, although the elephant uses its legs much like other running animals, and can move faster by quickening its stride. Fast-moving elephants appear to 'run' with their front legs, but 'walk' with their hind legs and can reach a top speed of 25 km/h (16 mph). At this speed, most other quadrupeds are well into a gallop, even accounting for leg length. Spring-like kinetics could explain the difference between the motion of elephants and other animals.[76][77] The cushion pads expand and contract, and reduce both the pain and noise that would come from a very heavy animal moving.[73] Elephants are capable swimmers: they can swim for up to six hours while completely waterborne, moving at 2.1 km/h (1 mph) and traversing up to 48 km (30 mi) continuously.[78]
Internal systems
The brain of an elephant weighs 4.5–5.5 kg (10–12 lb) compared to 1.6 kg (4 lb) for a human brain.[79] It is the largest of all terrestrial mammals.[80] While the elephant brain is larger overall, it is proportionally smaller than the human brain. At birth, an elephant's brain already weighs 30–40% of its adult weight. The cerebrum and cerebellum are well developed, and the temporal lobes are so large that they bulge out laterally.[79] Their temporal lobes are proportionally larger than those of other animals, including humans.[80] The throat of an elephant appears to contain a pouch where it can store water for later use.[29] The larynx of the elephant is the largest known among mammals. The vocal folds are anchored close to the epiglottis base. When comparing an elephant's vocal folds to those of a human, an elephant's are proportionally longer, thicker, with a greater cross-sectional area. In addition, they are located further up the vocal tract with an acute slope.[81]

The heart of an elephant weighs 12–21 kg (26–46 lb). Its apex has two pointed ends, an unusual trait among mammals.[79] In addition, the ventricles of the heart split towards the top, a trait also found in sirenians.[82] When upright, the elephant's heart beats around 28 beats per minute and actually speeds up to 35 beats when it lies down.[79] The blood vessels are thick and wide and can hold up under high blood pressure.[82] The lungs are attached to the diaphragm, and breathing relies less on the expanding of the ribcage.[79] Connective tissue exists in place of the pleural cavity. This may allow the animal to deal with the pressure differences when its body is underwater and its trunk is breaking the surface for air.[52] Elephants breathe mostly with the trunk but also with the mouth. They have a hindgut fermentation system, and their large and small intestines together reach 35 m (115 ft) in length. Less than half of an elephant's food intake gets digested, despite the process lasting a day.[79] An elephant's kidneys can produce more than 50 litres of urine per day.[83]
Sex characteristics
A male elephant's testes, like other Afrotheria,[84] are internally located near the kidneys.[85] The penis can be as long as 100 cm (39 in) with a 16 cm (6 in) wide base. It curves to an 'S' when fully erect and has an orifice shaped like a Y. The female's clitoris may be 40 cm (16 in). The vulva is found lower than in other herbivores, between the hind legs instead of under the tail. Determining pregnancy status can be difficult due to the animal's large belly. The female's mammary glands occupy the space between the front legs, which puts the suckling calf within reach of the female's trunk.[79] Elephants have a unique organ, the temporal gland, located on both sides of the head. This organ is associated with sexual behaviour, and males secrete a fluid from it when in musth.[86] Females have also been observed with these secretions.[53]
Behaviour and ecology

Elephants are herbivorous and will eat leaves, twigs, fruit, bark, grass, and roots. African elephants mostly browse, while Asian elephants mainly graze.[34] They can eat as much as 300 kg (660 lb) of food and drink 40 L (11 US gal) of water in a day. Elephants tend to stay near water sources.[34][87] They have morning, afternoon, and nighttime feeding sessions. At midday, elephants rest under trees and may doze off while standing. Sleeping occurs at night while the animal is lying down.[87] Elephants average 3–4 hours of sleep per day.[88] Both males and family groups typically move no more than 20 km (12 mi) a day, but distances as far as 180 km (112 mi) have been recorded in the Etosha region of Namibia.[89] Elephants go on seasonal migrations in response to changes in environmental conditions.[90] In northern Botswana, they travel 325 km (202 mi) to the Chobe River after the local waterholes dry up in late August.[91]
Because of their large size, elephants have a huge impact on their environments and are considered keystone species. Their habit of uprooting trees and undergrowth can transform savannah into grasslands;[92] smaller herbivores can access trees mowed down by elephants.[87] When they dig for water during droughts, they create waterholes that can be used by other animals. When they use waterholes, they end up making them bigger.[92] At Mount Elgon, elephants dig through caves and pave the way for ungulates, hyraxes, bats, birds, and insects.[92] Elephants are important seed dispersers; African forest elephants consume and deposit many seeds over great distances, with either no effect or a positive effect on germination.[93] In Asian forests, large seeds require giant herbivores like elephants and rhinoceros for transport and dispersal. This ecological niche cannot be filled by the smaller Malayan tapir.[94] Because most of the food elephants eat goes undigested, their dung can provide food for other animals, such as dung beetles and monkeys.[92] Elephants can have a negative impact on ecosystems. At Murchison Falls National Park in Uganda, elephant numbers have threatened several species of small birds that depend on woodlands. Their weight causes the soil to compress, leading to runoff and erosion.[87]
Elephants typically coexist peacefully with other herbivores, which will usually stay out of their way. Some aggressive interactions between elephants and rhinoceros have been recorded.[87] The size of adult elephants makes them nearly invulnerable to predators.[35] Calves may be preyed on by lions, spotted hyenas, and wild dogs in Africa[95] and tigers in Asia.[35] The lions of Savuti, Botswana, have adapted to hunting elephants, targeting calves, juveniles or even sub-adults.[96][97] There are rare reports of adult Asian elephants falling prey to tigers.[98] Elephants tend to have high numbers of parasites, particularly nematodes, compared to many other mammals. This may be due to elephants being less vulnerable to predation; in other mammal species, individuals weakened by significant parasite loads are easily killed off by predators, removing them from the population.[99]
Social organisation

Elephants are generally gregarious animals. African bush elephants in particular have a complex, stratified social structure.[100] Female elephants spend their entire lives in tight-knit matrilineal family groups.[101] They are led by the matriarch, who is often the eldest female.[102] She remains leader of the group until death[95] or if she no longer has the energy for the role;[103] a study on zoo elephants found that the death of the matriarch led to greater stress in the surviving elephants.[104] When her tenure is over, the matriarch's eldest daughter takes her place instead of her sister (if present).[95] One study found that younger matriarchs take potential threats less seriously.[105] Large family groups may split if they cannot be supported by local resources.[106]
At Amboseli National Park, Kenya, female groups may consist of around ten members, including four adults and their dependent offspring. Here, a cow's life involves interaction with those outside her group. Two separate families may associate and bond with each other, forming what are known as bond groups. During the dry season, elephant families may aggregate into clans. These may number around nine groups, in which clans do not form strong bonds but defend their dry-season ranges against other clans. The Amboseli elephant population is further divided into the "central" and "peripheral" subpopulations.[101]
Female Asian elephants tend to have more fluid social associations.[100] In Sri Lanka, there appear to be stable family units or "herds" and larger, looser "groups". They have been observed to have "nursing units" and "juvenile-care units". In southern India, elephant populations may contain family groups, bond groups, and possibly clans. Family groups tend to be small, with only one or two adult females and their offspring. A group containing more than two cows and their offspring is known as a "joint family". Malay elephant populations have even smaller family units and do not reach levels above a bond group. Groups of African forest elephants typically consist of one cow with one to three offspring. These groups appear to interact with each other, especially at forest clearings.[101]

Adult males live separate lives. As he matures, a bull associates more with outside males or even other families. At Amboseli, young males may be away from their families 80% of the time by 14–15 years of age. When males permanently leave, they either live alone or with other males. The former is typical of bulls in dense forests. A dominance hierarchy exists among males, whether they are social or solitary. Dominance depends on age, size, and sexual condition.[107] Male elephants can be quite sociable when not competing for mates and form vast and fluid social networks.[108][109] Older bulls act as the leaders of these groups.[110] The presence of older males appears to subdue the aggression and "deviant" behaviour of younger ones.[111] The largest all-male groups can reach close to 150 individuals. Adult males and females come together to breed. Bulls will accompany family groups if a cow is in oestrous.[107]
Sexual behaviour
Musth

Adult males enter a state of increased testosterone known as musth. In a population in southern India, males first enter musth at 15 years old, but it is not very intense until they are older than 25. At Amboseli, no bulls under 24 were found to be in musth, while half of those aged 25–35 and all those over 35 were. In some areas, there may be seasonal influences on the timing of musths. The main characteristic of a bull's musth is a fluid discharged from the temporal gland that runs down the side of his face. Behaviours associated with musth include walking with a high and swinging head, nonsynchronous ear flapping, picking at the ground with the tusks, marking, rumbling, and urinating in the sheath. The length of this varies between males of different ages and conditions, lasting from days to months.[112]
Males become extremely aggressive during musth. Size is the determining factor in agonistic encounters when the individuals have the same condition. In contests between musth and non-musth individuals, musth bulls win the majority of the time, even when the non-musth bull is larger. A male may stop showing signs of musth when he encounters a musth male of higher rank. Those of equal rank tend to avoid each other. Agonistic encounters typically consist of threat displays, chases, and minor sparring. Rarely do they full-on fight.[112]
Mating

Elephants are polygynous breeders,[113] and most copulations occur during rainfall.[114] An oestrous cow uses pheromones in her urine and vaginal secretions to signal her readiness to mate. A bull will follow a potential mate and assess her condition with the flehmen response, which requires him to collect a chemical sample with his trunk and taste it with the vomeronasal organ at the roof of the mouth.[115] The oestrous cycle of a cow lasts 14–16 weeks, with the follicular phase lasting 4–6 weeks and the luteal phase lasting 8–10 weeks. While most mammals have one surge of luteinizing hormone during the follicular phase, elephants have two. The first (or anovulatory) surge, appears to change the female's scent, signaling to males that she is in heat, but ovulation does not occur until the second (or ovulatory) surge.[116] Cows over 45–50 years of age are less fertile.[103]
Bulls engage in a behaviour known as mate-guarding, where they follow oestrous females and defend them from other males.[117] Most mate-guarding is done by musth males, and females seek them out, particularly older ones.[118] Musth appears to signal to females the condition of the male, as weak or injured males do not have normal musths.[119] For young females, the approach of an older bull can be intimidating, so her relatives stay nearby for comfort.[120] During copulation, the male rests his trunk on the female.[121] The penis is mobile enough to move without the pelvis.[83] Before mounting, it curves forward and upward. Copulation lasts about 45 seconds and does not involve pelvic thrusting or an ejaculatory pause.[122]
Homosexual behaviour is frequent in both sexes. As in heterosexual interactions, this involves mounting. Male elephants sometimes stimulate each other by playfighting, and "championships" may form between old bulls and younger males. Female same-sex behaviours have been documented only in captivity, where they engage in mutual masturbation with their trunks.[123]
Birth and development


Gestation in elephants typically lasts between one and a half and two years and the female will not give birth again for at least four years.[124] The relatively long pregnancy is supported by several corpus luteums and gives the foetus more time to develop, particularly the brain and trunk.[125] Births tend to take place during the wet season.[114] Typically, only a single young is born, but twins sometimes occur.[125] Calves are born roughly 85 cm (33 in) tall and with a weight of around 120 kg (260 lb).[120] They are precocial and quickly stand and walk to follow their mother and family herd.[126] A newborn calf will attract the attention of all the herd members. Adults and most of the other young will gather around the newborn, touching and caressing it with their trunks. For the first few days, the mother limits access to her young. Alloparenting – where a calf is cared for by someone other than its mother – takes place in some family groups. Allomothers are typically aged two to twelve years.[120]
For the first few days, the newborn is unsteady on its feet and needs its mother's help. It relies on touch, smell, and hearing, as its eyesight is less developed. With little coordination in its trunk, it can only flop it around which may cause it to trip. When it reaches its second week, the calf can walk with more balance and has more control over its trunk. After its first month, the trunk can grab and hold objects but still lacks sucking abilities, and the calf must bend down to drink. It continues to stay near its mother as it is still reliant on her. For its first three months, a calf relies entirely on its mother's milk, after which it begins to forage for vegetation and can use its trunk to collect water. At the same time, there is progress in lip and leg movements. By nine months, mouth, trunk, and foot coordination are mastered. Suckling bouts tend to last 2–4 min/hr for a calf younger than a year. After a year, a calf is fully capable of grooming, drinking, and feeding itself. It still needs its mother's milk and protection until it is at least two years old. Suckling after two years may improve growth, health, and fertility.[126]
Play behaviour in calves differs between the sexes; females run or chase each other while males play-fight. The former are sexually mature by the age of nine years[120] while the latter become mature around 14–15 years.[107] Adulthood starts at about 18 years of age in both sexes.[127][128] Elephants have long lifespans, reaching 60–70 years of age.[56] Lin Wang, a captive male Asian elephant, lived for 86 years.[129]
Communication

Elephants communicate in various ways. Individuals greet one another by touching each other on the mouth, temporal glands, and genitals. This allows them to pick up chemical cues. Older elephants use trunk-slaps, kicks, and shoves to control younger ones. Touching is especially important for mother–calf communication. When moving, elephant mothers will touch their calves with their trunks or feet when side-by-side or with their tails if the calf is behind them. A calf will press against its mother's front legs to signal it wants to rest and will touch her breast or leg when it wants to suckle.[130]
Visual displays mostly occur in agonistic situations. Elephants will try to appear more threatening by raising their heads and spreading their ears. They may add to the display by shaking their heads and snapping their ears, as well as tossing around dust and vegetation. They are usually bluffing when performing these actions. Excited elephants also raise their heads and spread their ears but additionally may raise their trunks. Submissive elephants will lower their heads and trunks, as well as flatten their ears against their necks, while those that are ready to fight will bend their ears in a V shape.[131]
Elephants produce several vocalisations—some of which pass though the trunk[132]—for both short and long range communication. This includes trumpeting, bellowing, roaring, growling, barking, snorting, and rumbling.[132][133] Elephants can produce infrasonic rumbles.[134] For Asian elephants, these calls have a frequency of 14–24 Hz, with sound pressure levels of 85–90 dB and last 10–15 seconds.[135] For African elephants, calls range from 15 to 35 Hz with sound pressure levels as high as 117 dB, allowing communication for many kilometres, possibly over 10 km (6 mi).[136] Elephants are known to communicate with seismics, vibrations produced by impacts on the earth's surface or acoustical waves that travel through it. An individual foot stomping or mock charging can create seismic signals that can be heard at travel distances of up to 32 km (20 mi). Seismic waveforms produced by rumbles travel 16 km (10 mi).[137][138]
Intelligence and cognition
Elephants are among the most intelligent animals. They exhibit mirror self-recognition, an indication of self-awareness and cognition that has also been demonstrated in some apes and dolphins.[139] One study of a captive female Asian elephant suggested the animal was capable of learning and distinguishing between several visual and some acoustic discrimination pairs. This individual was even able to score a high accuracy rating when re-tested with the same visual pairs a year later.[140] Elephants are among the species known to use tools. An Asian elephant has been observed fine-tuning branches for use as flyswatters.[141] Tool modification by these animals is not as advanced as that of chimpanzees. Elephants are popularly thought of as having an excellent memory. This could have a factual basis; they possibly have cognitive maps which give them long lasting memories of their environment on a wide scale. Individuals may be able to remember where their family members are located.[45]
Scientists debate the extent to which elephants feel emotion. They are attracted to the bones of their own kind, regardless of whether they are related.[142] As with chimpanzees and dolphins, a dying or dead elephant may elicit attention and aid from others, including those from other groups. This has been interpreted as expressing "concern";[143] however, the Oxford Companion to Animal Behaviour (1987) said that "one is well advised to study the behaviour rather than attempting to get at any underlying emotion".[144]
Conservation
Status

African bush elephants were listed as Endangered by the International Union for Conservation of Nature (IUCN) in 2021,[145] and African forest elephants were listed as Critically Endangered in the same year.[146] In 1979, Africa had an estimated population of at least 1.3 million elephants, possibly as high as 3.0 million. A decade later, the population was estimated to be 609,000; with 277,000 in Central Africa, 110,000 in Eastern Africa, 204,000 in Southern Africa, and 19,000 in Western Africa. The population of rainforest elephants was lower than anticipated, at around 214,000 individuals. Between 1977 and 1989, elephant populations declined by 74% in East Africa. After 1987, losses in elephant numbers hastened, and savannah populations from Cameroon to Somalia experienced a decline of 80%. African forest elephants had a total loss of 43%. Population trends in southern Africa were various, with unconfirmed losses in Zambia, Mozambique and Angola while populations grew in Botswana and Zimbabwe and were stable in South Africa.[147] The IUCN estimated that total population in Africa is estimated at to 415,000 individuals for both species combined as of 2016.[148]
Африканские слоны получают хоть какую-то правовую защиту в каждой стране, где они обитают. Успешные усилия по сохранению в определенных районах привели к высокой плотности населения, в то время как неудачи привели к снижению на 70% и более в течение десяти лет. По состоянию на 2008 год местная численность контролировалась с помощью контрацепции или транслокации . Масштабные выбраковки прекратились в конце 1980-х – начале 1990-х годов. В 1989 году африканский слон был внесен в Приложение I Конвенции о международной торговле видами дикой фауны и флоры, находящимися под угрозой исчезновения (СИТЕС), что сделало торговлю незаконной. Статус Приложения II (который разрешает ограниченную торговлю) был присвоен слонам в Ботсване, Намибии и Зимбабве в 1997 году и в Южной Африке в 2000 году. В некоторых странах спортивная охота на животных разрешена; Ботсвана, Камерун, Габон, Мозамбик, Намибия, Южная Африка, Танзания, Замбия и Зимбабве имеют экспортные квоты СИТЕС на трофеи слонов. [145]
В 2020 году МСОП включил азиатского слона в список находящихся под угрозой исчезновения из -за того, что его популяция сократилась вдвое за «последние три поколения». [149] Азиатские слоны когда-то обитали от Западной до Восточной Азии и на юг до Суматры . [150] и Ява. В настоящее время в этих районах он вымер. [149] а нынешний ареал азиатских слонов сильно фрагментирован. [150] Общая популяция азиатских слонов оценивается примерно в 40 000–50 000 особей, хотя это может быть приблизительная оценка. Около 60% населения проживает в Индии. Хотя численность азиатских слонов в целом сокращается, особенно в Юго-Восточной Азии, популяция в Западных Гатах, возможно, стабилизировалась. [149]
Угрозы

Браконьерство слонов ради их слоновой кости, мяса и шкур было одной из главных угроз их существованию. [149] Исторически сложилось так, что многие культуры изготавливали украшения и другие произведения искусства из слоновой кости, и ее использование было сопоставимо с использованием золота. [151] Торговля слоновой костью способствовала сокращению популяции африканских слонов в конце 20 века. [145] Это вызвало международные запреты на импорт слоновой кости, начиная с США в июне 1989 года, а затем последовали запреты в других странах Северной Америки, странах Западной Европы и Японии. [151] Примерно в то же время Кения уничтожила все свои запасы слоновой кости. [152] Слоновая кость была запрещена на международном уровне СИТЕС в 1990 году. После запретов выросла безработица в Индии и Китае, где промышленность слоновой кости имела важное экономическое значение. Напротив, Япония и Гонконг, которые также были частью отрасли, смогли адаптироваться и не пострадали так сильно. [151] Зимбабве, Ботсвана, Намибия, Замбия и Малави хотели продолжить торговлю слоновой костью, и им было разрешено это делать, поскольку их местное население было здоровым, но только в том случае, если их поставки поступали от отбракованных особей или тех, кто умер естественной смертью. [152]
Запрет позволил слону выздороветь в некоторых частях Африки. [151] В феврале 2012 года 650 слонов в национальном парке Буба-Нджида в Камеруне были убиты чадскими рейдерами. [153] Это событие назвали «одним из худших концентрированных убийств» со времен запрета на слоновую кость. [152] Азиатские слоны потенциально менее уязвимы для торговли слоновой костью, поскольку у самок обычно нет бивней. представителей этого вида убивали ради слоновой кости . Тем не менее, в некоторых районах, например, в национальном парке Перияр в Индии, [149] Китай был крупнейшим рынком сбыта браконьерской слоновой кости, но в мае 2015 года они объявили о поэтапном отказе от легального внутреннего производства и продажи изделий из слоновой кости, а в сентябре 2015 года Китай и США заявили, что «они введут почти полный запрет на импорт и продажу изделий из слоновой кости». экспорт слоновой кости» из-за причин исчезновения. [154]
Другие угрозы для слонов включают разрушение и фрагментацию среды обитания . Азиатский слон обитает в районах с самой высокой численностью населения и может быть ограничен небольшими лесными островками среди ландшафтов, где доминирует человек. Слоны обычно вытаптывают и поедают урожай, что способствует конфликтам с людьми, в результате чего и слоны, и люди гибнут сотнями. Смягчение этих конфликтов важно для сохранения природы. Одним из предлагаемых решений является защита коридоров дикой природы , которые дают населению большую взаимосвязанность и пространство. [149] Было обнаружено, что продукты из перца чили, а также охрана с помощью защитных инструментов эффективны в предотвращении набегов слонов на урожай. Менее эффективная тактика включает в себя улей и электрические заборы . [155]
Человеческие отношения
Рабочее животное

Слоны были рабочими животными, по крайней мере, со времен цивилизации долины Инда, более 4000 лет назад. [156] и продолжают использоваться в настоящее время. В 2000 году в Азии работало 13 000–16 500 рабочих слонов. Этих животных обычно отлавливают в дикой природе в возрасте 10–20 лет, когда они лучше поддаются дрессировке и могут работать больше лет. [157] Традиционно их ловили ловушками и арканами , но с 1950 года транквилизаторы . стали использовать [158] Особей азиатского вида часто дрессировали как рабочих животных. Азиатские слоны используются для переноски и вытаскивания предметов и людей в места и из них, а также для руководства людьми на религиозных праздниках. Их ценят перед механизированными инструментами, поскольку они могут выполнять те же задачи, но на более сложной местности, обладая силой, памятью и деликатностью. Слоны могут выучить более 30 команд. [157] С быками муста трудно и опасно работать, поэтому их приковывают цепью до тех пор, пока их состояние не пройдет. [159]
В Индии многие рабочие слоны, как утверждается, подвергались жестокому обращению. Таким образом, они и другие слоны в неволе находятся под защитой Закона о предотвращении жестокого обращения с животными 1960 года . [160] И в Мьянме, и в Таиланде вырубка лесов и другие экономические факторы привели к появлению значительных популяций безработных слонов, что привело к проблемам со здоровьем самих слонов, а также экономическим проблемам и проблемам безопасности для людей, среди которых они живут. [161] [162]
Практика использования слонов была предпринята и в Африке. Приручение африканских слонов в Бельгийском Конго началось по указу Леопольда II Бельгийского в 19 веке и продолжается по настоящее время в Центре приручения слонов Апи . [163]
Война

Исторически слоны считались грозным орудием войны. Они были описаны в санскритских текстах еще в 1500 году до нашей эры. Из Южной Азии использование слонов в войне распространилось на запад, в Персию. [164] и на восток в Юго-Восточную Азию. [165] Персы использовали их во времена империи Ахеменидов (между VI и IV веками до нашей эры). [164] в то время как государства Юго-Восточной Азии впервые использовали боевых слонов, возможно, еще в 5 веке до нашей эры и продолжали использовать их до 20 века. [165] Боевые слоны также использовались в Средиземноморье и Северной Африке на протяжении всего классического периода, начиная с правления Птолемея II в Египте. Карфагенский как полководец Ганнибал, известно, переправил африканских слонов через Альпы во время войны с римлянами и достиг долины реки По в 218 г. до н. э., все они были живы, но год спустя умер от болезней и боевых действий. [164]
Голова и бока слона были снабжены доспехами, к хоботу мог быть привязан меч, а бивни иногда покрывались заточенным железом или латунью. Дрессированные слоны нападали своими бивнями как на людей, так и на лошадей. Они могли схватить вражеского солдата хоботом и бросить его в свой погонщик , или прижать солдата к земле и пронзить его копьем. Некоторые недостатки боевых слонов включали их большую видимость, благодаря которой их было легко нацелить, и ограниченную маневренность по сравнению с лошадьми. Александр Великий добился победы над армиями с помощью боевых слонов, заставив своих солдат повредить хоботы и ноги животных, что заставило их запаниковать и стать неуправляемыми. [164]
Зоопарки и цирки

Слоны традиционно составляли значительную часть зоопарков и цирков по всему миру. В цирках их обучают выполнять трюки. Самым известным цирковым слоном, вероятно, был Джамбо (1861 – 15 сентября 1885), который был главной достопримечательностью цирка Барнума и Бейли . [166] [167] Эти животные плохо размножаются в неволе из-за сложности обращения с быками и ограниченного понимания эстральных циклов самок. Азиатские слоны всегда были более распространены в современных зоопарках и цирках, чем их африканские собратья. После того, как СИТЕС включил азиатского слона в Приложение I в 1975 году, к концу 1980-х годов импорт этого вида практически прекратился. Впоследствии США получили много содержащихся в неволе африканских слонов из Зимбабве, где животных было переизбыток. [167]
Содержание слонов в зоопарках вызвало некоторые споры. Сторонники зоопарков утверждают, что они обеспечивают легкий доступ к животным и предоставляют средства и знания для сохранения их естественной среды обитания, а также обеспечения безопасности видов. Оппоненты утверждают, что животные в зоопарках испытывают физический и психический стресс. [168] Было зарегистрировано, что слоны демонстрируют стереотипное поведение в виде покачивания телом или головой и прохождения одного и того же маршрута вперед и назад. Это наблюдалось у 54% особей в зоопарках Великобритании. [169] Слоны в европейских зоопарках, по-видимому, имеют более короткую продолжительность жизни, чем их дикие собратья, всего 17 лет, хотя другие исследования показывают, что слоны в зоопарках живут так же долго. [170]
Использование слонов в цирках также вызывает споры; Общество защиты животных США обвинило цирки в жестоком обращении и причинении страданий животным. [171] В своих показаниях перед федеральным судом США в 2009 году генеральный директор Barnum & Bailey Circus Кеннет Фелд признал, что цирковых слонов бьют за уши, под подбородок и по ногам штырями с металлическими наконечниками, называемыми бычьими крюками или анкусами. Фельд заявил, что эти методы необходимы для защиты работников цирка, и признал, что дрессировщика слонов упрекали за использование электрошокера на слоне. Несмотря на это, он отрицал, что какие-либо из этих действий причинили вред животным. [172] Некоторые дрессировщики пытались дрессировать слонов без применения физических наказаний. Известно, что Ральф Хелфер полагался на положительное подкрепление при дрессировке своих животных. [173] Цирк Барнума и Бейли отказался от гастрольных слонов в мае 2016 года. [174]
Атаки
Слоны могут проявлять приступы агрессивного поведения и совершать разрушительные действия против людей. [175] В Африке группы слонов-подростков повредили дома в деревнях после выбраковки в 1970-х и 1980-х годах. Из-за своевременности эти нападения были истолкованы как месть. [176] [177] В некоторых частях Индии слоны-самцы по ночам проникали в деревни, разрушая дома и убивая людей. С 2000 по 2004 год в Джаркханде погибло 300 человек , а в Ассаме , как сообщается, в период с 2001 по 2006 год было убито 239 человек. [175] В период с 2019 по 2022 год по всей стране слоны убили 1500 человек, в результате чего 300 слонов были убиты в натуральной форме. [178] Местные жители сообщили, что некоторые слоны во время нападений были пьяны, хотя официальные лица это оспаривают. [179] [180] Предположительно пьяные слоны напали на индийскую деревню в декабре 2002 года, убив шесть человек, что привело к ответной резне местных жителей около 200 слонов. [181]
Культурное значение
Слоны повсеместно присутствуют в мировой культуре. Они были представлены в искусстве со времен палеолита . Африка, в частности, содержит множество примеров наскального искусства со слонами , особенно в Сахаре и на юге Африки. [182] В Азии животные изображаются в качестве мотивов в индуистских и буддийских святынях и храмах. [183] Слонов часто было трудно изобразить людям, не имеющим непосредственного опыта их общения. [184] Древние римляне , которые держали животных в неволе, изображали слонов более точно, чем средневековые европейцы, которые изображали их скорее как фантастических существ, с чертами лошади, быка и кабана, а также хоботами, похожими на трубы. Поскольку европейцы получили больше доступа к плененным слонам в 15 веке, их изображения стали более точными, в том числе изображение, сделанное Леонардо да Винчи . [185]

Слоны были предметом религиозных верований. Народ мбути в Центральной Африке верит, что души их умерших предков обитали в слонах. [183] Подобные идеи существовали и среди других африканских обществ, которые верили, что их вожди перевоплотятся в слонов. В 10 веке нашей эры жители Игбо-Укву , на территории современной Нигерии, положили слоновьи бивни под ноги своего умершего вождя в могиле. [186] Значение животных тотемическое только в Африке, но гораздо более значимое в Азии. [187] На Суматре слоны ассоциировались с молниями. Точно так же в индуизме они связаны с грозами, поскольку Айравата , отец всех слонов, олицетворяет и молнию, и радугу. [183] с головой слона Одно из самых важных индуистских божеств, Ганеша приравнивается к верховным богам Шиве , Вишну и Брахме . , в некоторых традициях [188] Ганеша ассоциируется с писателями и торговцами, и считается, что он может дать людям успех и исполнить их желания, но также может и отнять их. [183] В буддизме говорится, что Будда был белым слоном, перевоплотившимся в человека. [189]
В западной массовой культуре слоны символизируют экзотику, тем более, что, как и в случае с жирафом , бегемотом и носорогом , подобных животных, знакомых западной аудитории, не существует. Как персонажи слоны чаще всего встречаются в детских сказках , где они изображены положительно. Обычно они являются суррогатами людей с идеальными человеческими ценностями. Многие истории рассказывают об изолированных молодых слонах, возвращающихся в семью или находящих ее, например, «Слоновий ребенок» из « Редьярда Киплинга » Такие истории , и » Диснея «Дамбо « Кэтрин и Байрона Джексонов Обвисший мешковатый слон» . Другие герои-слоны, человеческими качествами, Жана Брунхоффа де включают Бабара , Дэвида Макки Элмера наделенные и доктора Сьюза Хортона . [190]
Некоторые культурные отсылки подчеркивают размер и странность слона. Например, « белый слон » — это синоним чего-то странного, нежелательного и не имеющего никакой ценности. [190] Выражение « слон в комнате » относится к чему-то, что игнорируется, но в конечном итоге должно быть решено. [191] В истории о слепых и слоне слепые трогают разные части слона и пытаются понять, что это такое. [192]
См. также
- След животного
- Пустынный слон
- Слоновье кладбище
- Список отдельных слонов
- Мотти , пленный гибрид азиатского и африканского слона.
- Национальный день слонов (Таиланд)
- Всемирный день слонов
Ссылки
- ^ Перейти обратно: а б ἐλέφας . Лидделл, Генри Джордж ; Скотт, Роберт ; Греко-английский лексикон в проекте «Персей»
- ^ Перейти обратно: а б Харпер, Д. «Слон» . Интернет-словарь этимологии . Архивировано из оригинала 24 декабря 2013 года . Проверено 25 октября 2012 г.
- ^ Лухан, скорая помощь; Бернабе, А. «Изготовление слоновой кости и рогов в микенских текстах» . Космос. Ювелирные изделия, украшения и текстиль в Эгейском бронзовом веке . Архивировано из оригинала 20 октября 2021 года . Проверено 22 января 2013 г.
- ^ «слон» . Палеолексикон, инструмент для изучения слов древних языков. Архивировано из оригинала 4 декабря 2012 года . Проверено 19 января 2013 г.
- ^ Табусе, Р.; Ашер, Р.Дж.; Леманн, Т. (2008). «Афротерийские млекопитающие: обзор современных данных» (PDF) . Млекопитающие . 72 : 2–14. дои : 10.1515/МАММ.2008.004 . ISSN 0025-1461 . S2CID 46133294 . Архивировано из оригинала (PDF) 24 февраля 2021 года . Проверено 19 июня 2017 г.
- ^ Келлог, М.; Беркетт, С.; Деннис, ТР; Стоун, Г.; Грей, бакалавр; Макгуайр, премьер-министр; Зори, RT; Станьон, Р. (2007). «Хромосомная окраска ламантина поддерживает Afrotheria и Paenungulata» . Эволюционная биология . 7 (1): 6. Бибкод : 2007BMCEE...7....6K . дои : 10.1186/1471-2148-7-6 . ПМК 1784077 . ПМИД 17244368 .
- ^ Одзава, Т.; Хаяши, С.; Михельсон, В.М. (1997). «Филогенетическое положение мамонта и стеллеровой морской коровы внутри тетитерии, продемонстрированное последовательностями митохондриальной ДНК». Журнал молекулярной эволюции . 44 (4): 406–13. Бибкод : 1997JMolE..44..406O . дои : 10.1007/PL00006160 . ПМИД 9089080 . S2CID 417046 .
- ^ Шошани, Дж. (2005). «Отряд Хоботки». В Уилсоне, Делавэр; Ридер, Д.М. (ред.). Виды млекопитающих мира: таксономический и географический справочник . Том. 1 (3-е изд.). Издательство Университета Джонса Хопкинса. стр. 90–91. ISBN 978-0-8018-8221-0 . OCLC 62265494 . Архивировано из оригинала 1 февраля 2015 года . Проверено 11 ноября 2016 г.
- ^ Роланд, Н.; Райх, Д.; Маллик, С.; Мейер, М.; Зеленый, RE; Георгиадис, Нью-Джерси; Рока, Алабама; Хофрейтер, М. (2010). Пенни, Дэвид (ред.). «Последовательности геномной ДНК мастодонта и шерстистого мамонта раскрывают глубокое видообразование лесных слонов и слонов саванны» . ПЛОС Биология . 8 (12): e1000564. дои : 10.1371/journal.pbio.1000564 . ПМК 3006346 . ПМИД 21203580 .
- ^ Исида, Ю.; Олексик, ТК; Георгиадис, Нью-Джерси; Дэвид, Вирджиния; Чжао, К.; Стивенс, РМ; Колокотронис, С.-О.; Рока, Алабама (2011). Мерфи, Уильям Дж (ред.). «Урегулирование очевидных конфликтов между митохондриальной и ядерной филогенией у африканских слонов» . ПЛОС ОДИН . 6 (6): e20642. Бибкод : 2011PLoSO...620642I . дои : 10.1371/journal.pone.0020642 . ПМК 3110795 . ПМИД 21701575 .
- ^ Рока, Альфред Л.; Исида, Ясуко; Брандт, Адам Л.; Бенджамин, Нил Р.; Чжао, Кай; Георгиадис, Николас Дж. (2015). «Естественная история слонов: геномная перспектива». Ежегодный обзор биологических наук о животных . 3 (1): 139–167. doi : 10.1146/annurev-animal-022114-110838 . ПМИД 25493538 .
- ^ Рока, Альфред Л.; Исида, Ясуко; Брандт, Адам Л.; Бенджамин, Нил Р.; Чжао, Кай; Георгиадис, Николас Дж. (2015). «Естественная история слонов: геномная перспектива». Ежегодный обзор биологических наук о животных . 3 (1): 139–167. doi : 10.1146/annurev-animal-022114-110838 . ПМИД 25493538 .
- ^ Перейти обратно: а б Палкопулу, Элефтерия; Липсон, Марк; Маллик, Свапан; Нильсен, Свенд; Роланд, Надин; Балека, Сина; Карпински, Эмиль; Иванцевич, Атма М.; То, Ту-Хиен; Корчак, Р. Дэниел; Рэйсон, Джой М. (13 марта 2018 г.). «Всеобъемлющая геномная история вымерших и ныне живущих слонов» . Труды Национальной академии наук . 115 (11): E2566–E2574. Бибкод : 2018PNAS..115E2566P . дои : 10.1073/pnas.1720554115 . ISSN 0027-8424 . ПМЦ 5856550 . ПМИД 29483247 .
- ^ Кингдон, Дж. (2013). Млекопитающие Африки . Блумсбери. п. 173. ИСБН 9781408189962 . Архивировано из оригинала 21 марта 2023 года . Проверено 6 июня 2020 г.
- ^ Гербрант, Э. (2009). «Палеоценовое появление родственников слонов и быстрая радиация африканских копытных» . Труды Национальной академии наук Соединенных Штатов Америки . 106 (26): 10717–10721. Бибкод : 2009PNAS..10610717G . дои : 10.1073/pnas.0900251106 . ПМК 2705600 . ПМИД 19549873 .
- ^ Перейти обратно: а б Сукумар, стр. 13–16.
- ^ Лю, AGSC; Зейферт, ER; Саймонс, Э.Л. (2008). «Стабильные изотопы свидетельствуют о земноводной фазе ранней эволюции хоботных» . Труды Национальной академии наук . 105 (15): 5786–5791. Бибкод : 2008PNAS..105.5786L . дои : 10.1073/pnas.0800884105 . ПМК 2311368 . ПМИД 18413605 .
- ^ Шошани, Дж.; Уолтер, Р.К.; Абраха, М.; Берхе, С.; Тасси, П.; Сандерс, WJ; Маршан, GH; Либсекаль, Ю.; Гирмаи, Т.; Зиннер, Д. (2006). «Хобоцид из позднего олигоцена Эритреи, «недостающее звено» между ранними Elephantiformes и Elephantimorpha, а также биогеографические последствия» . Труды Национальной академии наук . 103 (46): 17296–17301. Бибкод : 2006PNAS..10317296S . дои : 10.1073/pnas.0603689103 . ПМК 1859925 . ПМИД 17085582 .
- ^ Балека, С.; Варела, Л.; Тамбуссо, PS; Пайманс, JLA; Моте, Д.; Стаффорд-младший, ТВ; Фаринья, РА; Хофрейтер, М. (2022). «Возврат к филогении и эволюции хоботных на основе полных доказательств и палеогенетического анализа, включая нотиомастодонов древнюю ДНК » . iScience . 25 (1): 103559. Бибкод : 2022iSci...25j3559B . дои : 10.1016/j.isci.2021.103559 . ПМЦ 8693454 . ПМИД 34988402 .
- ^ Бенуа, Дж.; Лирас, Джорджия; Шмитт, А.; Нксумало, М.; Табусе, Р.; Обада, Т.; Марарсекул, В.; Мангер, П. (2023). «Палеоневрология хоботков (млекопитающих, афротерий): данные из эндокаста их мозга и лабиринта». В Дозо, Монтана; Паулина-Карабахал, А.; Макрини, Т.Э.; Уолш, С. (ред.). Палеоневрология амниотов . Чам: Международное издательство Springer. стр. 579–644. дои : 10.1007/978-3-031-13983-3_15 . ISBN 978-3-031-13982-6 .
- ^ Перейти обратно: а б с д Канталапьедра, JL; Санисидро, О.; Чжан, Х.; Альберди, Монтана; Прадо, Дж.Л.; Бланко, Ф.; Сааринен, Дж. (2021). «Взлет и падение экологического разнообразия хоботных» . Экология и эволюция природы . 5 (9): 1266–1272. Бибкод : 2021NatEE...5.1266C . дои : 10.1038/s41559-021-01498-w . ПМИД 34211141 . S2CID 235712060 .
- ^ Саэгуса, Х.; Накая, Х.; Кунимацу, Ю.; Накацукаса, М.; Цудзикава, Х.; Савада, Ю.; Санеёси М. и Сакаи Т. (2014). «Останки самых ранних слонов из местонахождения позднего миоцена, Накали, Кения» (PDF) . В Костопулосе, Д.С.; Влахос Э. и Цукала Э. (ред.). VI Международная конференция по мамонтам и их родственникам . Том. 102. Салоники: Школа геологии, Университет Аристотеля в Салониках. п. 175. ИСБН 978-960-9502-14-6 .
- ^ Листер, AM (2013). «Роль поведения в адаптивной морфологической эволюции африканских хоботных». Природа . 500 (7462): 331–334. Бибкод : 2013Natur.500..331L . дои : 10.1038/nature12275 . ПМИД 23803767 . S2CID 883007 .
- ^ Яннуччи, Алессио; Сарделла, Рафаэле (28 февраля 2023 г.). «Что означает событие «Слон- Эквус » сегодня? Размышления о событиях расселения млекопитающих вокруг границы плиоцена и плейстоцена и гибкой двусмысленности биохронологии» . Четвертичный период . 6 (1): 16. дои : 10,3390/quat6010016 . hdl : 11573/1680082 .
- ^ Моте, Димила; дос Сантос Авилья, Леонардо; Асеведо, Лидиана; Борхес-Сильва, Леон; Росас, Мариан; Лабарка-Энсина, Рафаэль; Суберлих, Рикардо; Сойбельзон, Эстебан; Роман-Каррион, Хосе Луис; Риос, Серхио Д.; Ринкон, Асканио Д.; Кардосо де Оливейра, Джина; Перейра Лопес, Ренато (30 сентября 2016 г.). «Шестьдесят лет после« Мастодонтов Бразилии »: современное состояние южноамериканских хоботных (Proboscidea, Gomphotheriidae)» (PDF) . Четвертичный интернационал . 443 :52–64. Бибкод : 2017QuInt.443...52M . дои : 10.1016/j.quaint.2016.08.028 .
- ^ Листер, AM; Шер, А.В. (2015). «Эволюция и распространение мамонтов по Северному полушарию». Наука . 350 (6262): 805–809. Бибкод : 2015Sci...350..805L . дои : 10.1126/science.aac5660 . ПМИД 26564853 . S2CID 206639522 .
- ^ Листер, AM (2004). «Экологические взаимодействия слонов в плейстоценовой Евразии» . Палеоэкология человека в Левантийском коридоре . Книги Оксбоу. стр. 53–60. ISBN 978-1-78570-965-4 .
- ^ Роджерс, РЛ; Слаткин, М. (2017). «Избыток геномных дефектов у шерстистого мамонта на острове Врангеля» . ПЛОС Генетика . 13 (3): e1006601. дои : 10.1371/journal.pgen.1006601 . ISSN 1553-7404 . ПМЦ 5333797 . ПМИД 28253255 .
- ^ Перейти обратно: а б с д и ж г Шошани, Дж. (1998). «Понимание эволюции хоботных: непростая задача». Тенденции экологии и эволюции . 13 (12): 480–487. дои : 10.1016/S0169-5347(98)01491-8 . ПМИД 21238404 .
- ^ Сукумар, стр. 31–33.
- ^ Романо, М.; Мануччи, Ф.; Паломбо, MR (2021). «Самый маленький из крупнейших: новая оценка объемной массы тела и восстановление in vivo карликового слона Palaeoloxodon ex gr. P. falconeri из пещеры Спинагалло (Сицилия)». Историческая биология . 33 (3): 340–353. Бибкод : 2021HBio...33..340R . дои : 10.1080/08912963.2019.1617289 . S2CID 181855906 .
- ^ Перейти обратно: а б с Ларраменди, А. (2015). «Высота плеч, масса тела и форма хоботков» . Acta Palaeontologica Polonica . дои : 10.4202/app.00136.2014 .
- ^ Перейти обратно: а б с д и Шошани, с. 38–41.
- ^ Перейти обратно: а б с д и Шошани, с. 42–51.
- ^ Перейти обратно: а б с Шошани, Дж.; Айзенберг, Дж. Ф. (1982). « Большой слон » (PDF) . Виды млекопитающих (182): 1–8. дои : 10.2307/3504045 . JSTOR 3504045 . Архивировано из оригинала (PDF) 24 сентября 2015 года . Проверено 27 октября 2012 г.
- ^ «Самые крупные наземные животные в мире» . Факт Животное . Проверено 29 сентября 2023 г.
- ^ Перейти обратно: а б с Шошани, с. 68–70.
- ^ Сомгрид, К. «Анатомия и биология слона: скелетная система» . Центр исследований и образования слонов, отделение клиники для домашних животных и дикой природы, факультет ветеринарной медицины, Университет Чиангмая. Архивировано из оригинала 13 июня 2012 года . Проверено 21 сентября 2012 г.
- ^ Кингдон, с. 11.
- ^ Моул, Майкл А.; Родригес Дараужо, Шон; Ван Аарде, Руди Дж.; Митчелл, Дункан; Фуллер, Андреа (2018). «Слоны саванны сохраняют гомеотермию в условиях африканской жары» . Журнал сравнительной физиологии Б. 188 (5): 889–897. дои : 10.1007/s00360-018-1170-5 . ПМИД 30008137 . S2CID 51626564 . Архивировано из оригинала 15 мая 2021 года . Проверено 14 мая 2021 г.
- ^ Нарасимхан, А. (2008). «Почему у слонов большие уши?». Резонанс . 13 (7): 638–647. дои : 10.1007/s12045-008-0070-5 . S2CID 121443269 .
- ^ Рейтер, Т.; Нуммела, С.; Хемила, С. (1998). «Слоновий слух» (PDF) . Журнал Акустического общества Америки . 104 (2): 1122–1123. Бибкод : 1998ASAJ..104.1122R . дои : 10.1121/1.423341 . ПМИД 9714930 . Архивировано из оригинала (PDF) 7 декабря 2012 года.
- ^ Сомгрид, К. «Анатомия и биология слона: особые органы чувств» . Центр исследований и образования слонов, отделение клиники для домашних животных и дикой природы, факультет ветеринарной медицины, Университет Чиангмая. Архивировано из оригинала 29 июля 2013 года . Проверено 21 сентября 2012 г.
- ^ Ёкояма, С.; Такенака, Н.; Агнью, Д.В.; Шошани, Дж. (2005). «Слоны и люди-дейтеранопы, страдающие дальтонизмом, имеют идентичный набор зрительных пигментов» . Генетика . 170 (1): 335–344. дои : 10.1534/genetics.104.039511 . ПМЦ 1449733 . ПМИД 15781694 .
- ^ Перейти обратно: а б Бирн, RW; Бейтс, Л.; Мосс CJ (2009). «Познание слона с точки зрения приматов» . Сравнительные обзоры познания и поведения . 4 : 65–79. дои : 10.3819/ccbr.2009.40009 . hdl : 10023/1612 .
- ^ Перейти обратно: а б с д Шошани, с. 74–77.
- ^ Перейти обратно: а б с Мартин, Ф.; Нимиц К. (2003). « «Правые хоботы» и «левые хоботы»: боковые предпочтения движений хобота у диких азиатских слонов ( Elephas maximus )». Журнал сравнительной психологии . 117 (4): 371–379. дои : 10.1037/0735-7036.117.4.371 . ПМИД 14717638 .
- ^ Дагенайс, П; Хенсман, С; Хэхлер, В; Милинкович, MC (2021). «Слоны развили стратегии, уменьшающие биомеханическую сложность их хобота» . Современная биология . 31 (21): 4727–4737. Бибкод : 2021CBio...31E4727D . дои : 10.1016/j.cub.2021.08.029 . ПМИД 34428468 . S2CID 237273086 .
- ^ Шульц, АК; Бойл, М; Бойл, К; Сордилья, С; Ринкон, К; Хупер, К; Обюшон, К; Рейденберг, Дж. С.; Хиггинс, К; Ху, Д.Л. (2022). «Морщины и складки на коже способствуют асимметричному растяжению хобота слона» . Труды Национальной академии наук . 119 (31): e2122563119. Бибкод : 2022PNAS..11922563S . дои : 10.1073/pnas.2122563119 . ПМЦ 9351381 . PMID 35858384 .
- ^ Кингдон, с. 9.
- ^ Перейти обратно: а б Шульц, АК; Нин Ву, Цзя; Сара Ха, Ю.С.; Ким, Г. (2021). «Всасывающее кормление слонами» . Журнал интерфейса Королевского общества . 18 (179). дои : 10.1098/rsif.2021.0215 . ПМК 8169210 . ПМИД 34062103 .
- ^ Перейти обратно: а б Уэст, Дж. Б. (2002). «Почему у слона нет плевральной полости?» . Физиология . 17 (2): 47–50. дои : 10.1152/nips.01374.2001 . ПМИД 11909991 . S2CID 27321751 .
- ^ Перейти обратно: а б Сукумар, с. 149.
- ^ Дейрингер, Нора; Белоснежка, Ундина; Кауфманн, Лена В.; Эйген, Леннарт; Шпайсеггер, Селина; Герхардт, Бен; Хольце, Сюзанна; Фрич, Гвидо; Гёриц, Франк; Беккер, Рольф; Окс, Андреас; Хильдебрандт, Томас; Брехт, Майкл (8 июня 2023 г.). «Функциональная анатомия усов хобота слона» . Коммуникационная биология . 6 (1): 591. doi : 10.1038/s42003-023-04945-5 . ISSN 2399-3642 . ПМЦ 10250425 . ПМИД 37291455 .
- ^ Коул, М. (14 ноября 1992 г.). "Свинца в озере обвиняют в гибких стволах" . Новый учёный . Архивировано из оригинала 17 мая 2008 года . Проверено 25 июня 2009 г.
- ^ Перейти обратно: а б Шошани, с. 70–71.
- ^ Перейти обратно: а б с Шошани, с. 71–74.
- ^ Сукумар, с. 120
- ^ Клаттон-Брок, Дж. (1986). Естественная история одомашненных млекопитающих . Британский музей (естественная история). п. 208. ИСБН 978-0-521-34697-9 .
- ^ «Из-за браконьерства у слонов появились бивни меньшего размера» . Сеть экологических новостей. 20 января 2008 г. Архивировано из оригинала 21 ноября 2015 г. . Проверено 25 сентября 2012 г.
- ^ «Под давлением браконьерства слоны теряют свои бивни» . Нэшнл Географик . 9 ноября 2018 года. Архивировано из оригинала 3 марта 2021 года . Проверено 28 октября 2021 г.
- ^ Грей, Р. (20 января 2008 г.). «Почему у слонов не такой длинный бивень» . «Дейли телеграф» . Архивировано из оригинала 18 октября 2009 года . Проверено 27 января 2013 г.
- ^ Чиё, ИП; Обанда, В.; Корир, ДК (2015). «Незаконный сбор бивней и уменьшение размера бивней африканского слона» . Экология и эволюция . 5 (22): 5216–5229. Бибкод : 2015EcoEv...5.5216C . дои : 10.1002/ece3.1769 . ПМК 6102531 . ПМИД 30151125 .
- ^ Яхманн, Х.; Берри, ПСМ; Имаэ, Х. (1995). «Без клыков у африканских слонов: тенденция будущего». Африканский журнал экологии . 33 (3): 230–235. Бибкод : 1995AfJEc..33..230J . дои : 10.1111/j.1365-2028.1995.tb00800.x .
- ^ Курт, Ф.; Хартл, Г.; Тидеманн, Р. (1995). «Бесклыковые быки у азиатского слона Elephas maximus . История и популяционная генетика рукотворного феномена» . Акта Териол . 40 : 125–144. дои : 10.4098/at.arch.95-51 .
- ^ Перейти обратно: а б Шошани, с. 66–67.
- ^ Мирволд, CL; Стоун, штат Ха; Бу-Зейд, Э. (10 октября 2012 г.). «Для чего нужна слоновья шерсть?» . ПЛОС ОДИН . 7 (10): е47018. Бибкод : 2012PLoSO...747018M . дои : 10.1371/journal.pone.0047018 . ПМЦ 3468452 . ПМИД 23071700 .
- ^ Лампы ДВ; Смоллер, БР; Расмуссен, ЛЕЛ; Слэйд, Британская Колумбия; Фрич, Г; Годвин, TE (2001). «Характеристика межпальцевых желез азиатского слона (Elephas maximus)». Исследования в области ветеринарии . 71 (3): 197–200. дои : 10.1053/rvsc.2001.0508 . ПМИД 11798294 .
- ^ Райт, П.Г.; Удача, КП (1984). «Нужно ли слонам потеть?» . Журнал зоологии . 19 (4): 270–274. дои : 10.1080/02541858.1984.11447892 .
- ^ Спирмен, РИК (1970). «Эпидермис и его ороговение у африканского слона (Loxodonta Africana)» . Зоология Африканская . 5 (2): 327–338. дои : 10.1080/00445096.1970.11447400 .
- ^ Мартинс, Антониу Ф.; Беннетт, Найджел С.; Клавель, Сильви; Гроеневальд, Герман; Хенсман, Шон; Хоби, Стефан; Жорис, Антуан; Мангер, Пол Р.; Милинкович, Мишель К. (2018). «Локально-изогнутая геометрия приводит к образованию трещин на коже африканского слона» . Природные коммуникации . 9 (1): 3865. Бибкод : 2018NatCo...9.3865M . дои : 10.1038/s41467-018-06257-3 . ПМК 6168576 . ПМИД 30279508 .
- ^ Перейти обратно: а б с Шошани, с. 69–70.
- ^ Перейти обратно: а б с Вайсенгрубер, GE; Эггер, Г.Ф.; Хатчинсон-младший; Грёневальд, HB; Эльсэссер, Л.; Фамини, Д.; Форстенпойнтнер, Г. (2006). «Строение подушечек стоп африканских слонов ( Loxodonta africana )» . Журнал анатомии . 209 (6): 781–792. дои : 10.1111/j.1469-7580.2006.00648.x . ПМК 2048995 . ПМИД 17118065 .
- ^ Шошани, с. 74.
- ^ Хатчинсон-младший; Делмер, К; Миллер, CE; Хильдебрандт, Т; Пициллидес, А.А.; Бойд, А. (2011). «От плоскостопия к толстой стопе: строение, онтогенез, функции и эволюция «шестых пальцев» слона » (PDF) . Наука . 334 (6063): 1699–1703. Бибкод : 2011Sci...334R1699H . дои : 10.1126/science.1211437 . ПМИД 22194576 . S2CID 206536505 .
- ^ Перейти обратно: а б Хатчинсон-младший; Шверда, Д.; Фамини, диджей; Дейл, Р.Х.; Фишер, М.С.; Крам, Р. (2006). «Локомоторная кинематика азиатских и африканских слонов: меняется в зависимости от скорости и размера» . Журнал экспериментальной биологии . 209 (19): 3812–3827. дои : 10.1242/jeb.02443 . ПМИД 16985198 .
- ^ Хатчинсон-младший; Фамини, Д.; Лэр, Р.; Крам, Р. (2003). «Биомеханика: действительно ли быстро движущиеся слоны бегут?» . Природа . 422 (6931): 493–494. Бибкод : 2003Natur.422..493H . дои : 10.1038/422493а . ПМИД 12673241 . S2CID 4403723 . Архивировано из оригинала 9 августа 2022 года . Проверено 3 января 2023 г.
- ^ Шошани, с. 60.
- ^ Перейти обратно: а б с д и ж г Шошани, с. 78–79.
- ^ Перейти обратно: а б О'Коннелл, Кейтлин (20 июля 2016 г.). «Слон Дон: Политика отряда толстокожих» . Издательство Чикагского университета. ISBN 9780226380056 . Архивировано из оригинала 21 марта 2023 года . Проверено 16 сентября 2022 г.
- ^ Хербест, Коннектикут; Швец, Ю.Г.; Лошеллер, Дж.; Фрей, Р.; Гумпенбергер, М.; Штегер, А.; Фитч, WT (2013). «Сложные образцы вибрации в гортани слона» . Журнал экспериментальной биологии . 216 (21): 4054–4064. дои : 10.1242/jeb.091009 . ПМИД 24133151 .
- ^ Перейти обратно: а б Анон (2010). Анатомия млекопитающих: Иллюстрированное руководство . Маршалл Кавендиш. п. 59. ИСБН 978-0-7614-7882-9 .
- ^ Перейти обратно: а б Мюррей Э. Фаулер; Сьюзан К. Микота (2006). Биология, медицина и хирургия слонов . Джон Уайли и сыновья. п. 353. ИСБН 978-0-8138-0676-1 . Архивировано из оригинала 21 марта 2023 года . Проверено 17 октября 2020 г.
- ^ Шарма, Вираг; Леманн, Томас; Стукас, Хейко; Функе, Лиана; Хиллер, Майкл (2018). «Потеря генов RXFP2 и INSL3 у Afrotheria показывает, что происхождение семенников является наследственным состоянием плацентарных млекопитающих» . ПЛОС Биология . 16 (6): e2005293. doi : 10.1371/journal.pbio.2005293 . ISSN 1545-7885 . ПМК 6023123 . ПМИД 29953435 .
- ^ Короткий, Р.В.; Манн, Т.; Хэй, Мэри Ф. (1967). «Мужские репродуктивные органы африканского слона Loxodonta africana» (PDF) . Журнал репродукции и фертильности . 13 (3): 517–536. дои : 10.1530/jrf.0.0130517 . ПМИД 6029179 . Архивировано (PDF) из оригинала 20 июля 2018 года . Проверено 10 июля 2017 г.
- ^ Шошани, с. 80.
- ^ Перейти обратно: а б с д и Элтрингем, стр. 124–27.
- ^ Сигел, Дж. М. (2005). «Подсказки к функциям сна млекопитающих» . Природа . 437 (7063): 1264–1271. Бибкод : 2005Natur.437.1264S . дои : 10.1038/nature04285 . ПМЦ 8760626 . ПМИД 16251951 . S2CID 234089 .
- ^ Сукумар, с. 159.
- ^ Сукумар, с. 174.
- ^ Хоар, Б. (2009). Миграция животных: замечательные путешествия в дикой природе . Издательство Калифорнийского университета. стр. 58–59. ISBN 978-0-520-25823-5 .
- ^ Перейти обратно: а б с д Шошани, с. 226–29.
- ^ Кампос-Арсеис, А.; Блейк, С. (2011). «Мега-садовники леса – роль слонов в распространении семян» (PDF) . Акта Экологика . 37 (6): 542–553. Бибкод : 2011AcO....37..542C . дои : 10.1016/j.actao.2011.01.014 . Архивировано (PDF) из оригинала 18 июня 2013 года . Проверено 22 ноября 2012 г.
- ^ Кампос-Арсеис, А.; Трэхольт, К.; Джаффар, Р.; Сантамария, Л.; Корлетт, RT (2012). «Азиатские тапиры — не слоны, когда дело доходит до распространения семян». Биотропика . 44 (2): 220–227. Бибкод : 2012Биотр..44..220С . дои : 10.1111/j.1744-7429.2011.00784.x . hdl : 10261/56573 . S2CID 83604698 .
- ^ Перейти обратно: а б с Лаурсен, Л.; Бекофф, М. (1978). « Локсодонта африканская » (PDF) . Виды млекопитающих (92): 1–8. дои : 10.2307/3503889 . JSTOR 3503889 . S2CID 253949585 . Архивировано из оригинала (PDF) 30 апреля 2013 года.
- ^ Пауэр, Р.Дж.; Шем Компион, RX (2009). «Похищение львами слонов в Савути, национальный парк Чобе, Ботсвана». Африканская зоология . 44 (1): 36–44. дои : 10.3377/004.044.0104 . S2CID 86371484 .
- ^ Жубер, Д. (2006). «Охотничье поведение львов ( Panthera leo ) на слонах ( Loxodonta africana ) в Национальном парке Чобе, Ботсвана». Африканский журнал экологии . 44 (2): 279–281. Бибкод : 2006AfJEc..44..279J . дои : 10.1111/j.1365-2028.2006.00626.x .
- ^ «Тигр убивает слона в парке Эравикулам» . Новый Индийский экспресс . 2009. Архивировано из оригинала 11 мая 2016 года . Проверено 21 июня 2014 г.
- ^ Сукумар, с. 121.
- ^ Перейти обратно: а б де Сильва, С.; Виттемайер, Г. (2012). «Сравнение социальной организации азиатских слонов и слонов африканской саванны». Международный журнал приматологии . 33 (5): 1125–1141. дои : 10.1007/s10764-011-9564-1 . S2CID 17209753 .
- ^ Перейти обратно: а б с Сукумар, стр. 172, 175–79.
- ^ Сукумар, с. 186.
- ^ Перейти обратно: а б Кингдон, с. 53.
- ^ Харрис, М.; Шервин, К.; Харрис, С. (2008). «Окончательный отчет Defra о благополучии слонов» (PDF) . Бристольский университет. Архивировано (PDF) из оригинала 24 ноября 2014 года . Проверено 30 октября 2012 г.
- ^ МакКомб, К.; Шеннон, Г.; Дюрант, С.М.; Сайялель, К.; Слотоу, Р.; Пул, Дж.; Мосс, К. (2011). «Лидерство слонов: адаптивное значение возраста» (PDF) . Труды Королевского общества B: Биологические науки . 278 (1722): 3270–3276. дои : 10.1098/rspb.2011.0168 . ПМК 3169024 . ПМИД 21411454 . Архивировано (PDF) из оригинала 29 апреля 2013 года . Проверено 26 декабря 2012 г.
- ^ Воган, Т.; Райан, Дж.; Чаплевский, Н. (2011). Маммология . Джонс и Бартлетт Обучение. п. 136. ИСБН 978-0763762995 .
- ^ Перейти обратно: а б с Сукумар, стр. 179–83.
- ^ Гольденберг, СЗ; де Сильва, С.; Расмуссен, Х.Б.; Дуглас-Гамильтон, И.; Виттемайер, Г. (2014). «Контроль поведенческого состояния выявляет социальную динамику среди самцов африканских слонов Loxodonta africana ». Поведение животных . 95 : 111–119. дои : 10.1016/j.anbehav.2014.07.002 . S2CID 53152412 .
- ^ Ладью, Калифорния; Вандерконе, РПГ; Кисо, Западная Келли; Фриман, EW (2022). «Социальное поведение и формирование групп самцов азиатских слонов (Elephas maximus): влияние возраста и потребности у диких животных и животных, содержащихся в зоопарках» . Животные . 12 (9): 1215. дои : 10.3390/ani12091215 . ПМК 9100748 . ПМИД 35565641 .
- ^ Аллен, CRB; Брент, LJN; Моцентва, Т; Вайс, Миннесота; Крофт, ДП (2020). «Значение старых быков: лидеры и последователи в коллективных движениях мужских групп слонов африканской саванны ( Loxodonta africana )» . Научные отчеты . 10 (1): 13996. Бибкод : 2020NatSR..1013996A . дои : 10.1038/s41598-020-70682-y . ПМЦ 7471917 . ПМИД 32883968 .
- ^ Слотоу, Р.; ван Дайк, Г.; Пул, Дж.; Пейдж, Б.; Клок, А. (2000). «Старые слоны-быки контролируют молодых самцов». Природа . 408 (6811): 425–426. Бибкод : 2000Natur.408..425S . дои : 10.1038/35044191 . ПМИД 11100713 . S2CID 136330 .
- ^ Перейти обратно: а б Сукумар, стр. 100–08.
- ^ Сукумар, с. 89.
- ^ Перейти обратно: а б Сукумар, с. 262.
- ^ Сукумар, стр. 98–99.
- ^ «Проект размножения слонов: эстральный цикл слонов» . Смитсоновский национальный зоопарк. Архивировано из оригинала 6 июня 2012 года . Проверено 8 октября 2012 года .
- ^ Пул Джойс Х (1989). «Охрана партнера, репродуктивный успех и выбор самок африканских слонов» (PDF) . Поведение животных . 37 : 842–849. дои : 10.1016/0003-3472(89)90068-7 . S2CID 53150105 . Архивировано из оригинала 24 октября 2018 года . Проверено 24 октября 2018 г.
- ^ Сукумар, с. 113.
- ^ Сукумар, с. 117.
- ^ Перейти обратно: а б с д Мосс, стр. 106–13.
- ^ Кингдон, с. 69.
- ^ Эстес, Р. (1991). Руководство по поведению млекопитающих Африки: в том числе копытных, хищников, приматов . Издательство Калифорнийского университета. стр. 263–266 . ISBN 978-0-520-08085-0 .
- ^ Багемил, Б. (1999). Биологическое изобилие: гомосексуализм животных и естественное разнообразие . Пресса Святого Мартина. стр. 427–430 . ISBN 978-1-4668-0927-7 .
- ^ Сукумар, стр. 259–60.
- ^ Перейти обратно: а б Людерс, И.; Нимюллер, К.; Рич, П.; Грей, К.; Гермес, Р.; Гериц, Ф.; Хильдебрандт, ТБ (2012). «Гестация 22 месяца: развитие лютеина и поддержание беременности у слонов» . Труды Королевского общества B: Биологические науки . 279 (1743): 3687–3696. дои : 10.1098/rspb.2012.1038 . ПМЦ 3415912 . ПМИД 22719030 .
- ^ Перейти обратно: а б Сукумар, стр. 126–29.
- ^ «Жизненный цикл слона» . ElephantsForever.co.za. Архивировано из оригинала 29 сентября 2014 года . Проверено 21 июня 2014 г.
- ^ «Жизненный цикл слона – подростковый возраст» . ElephantsForever.co.za. Архивировано из оригинала 30 мая 2014 года . Проверено 21 июня 2014 г.
- ^ «Умер слон-ветеран войны» . Новости Би-би-си . 26 февраля 2003 г. Архивировано из оригинала 12 ноября 2012 г. Проверено 8 января 2013 г.
- ^ Пейн и Лангбауэр, с. 116.
- ^ Пейн и Лангбауэр, стр. 119–20.
- ^ Перейти обратно: а б Пейн и Лангбауэр, стр. 120–21.
- ^ Сукумар, с. 141.
- ^ Хербест, Коннектикут; Штегер, А.; Фрей, Р.; Лошеллер, Дж.; Титце, ИК; Гумпенбергер, М.; Фитч, WT (2012). «Насколько низко вы можете опуститься? Физический механизм производства инфразвуковых вокализаций слонов». Наука . 337 (6094): 595–599. Бибкод : 2012Sci...337..595H . дои : 10.1126/science.1219712 . ПМИД 22859490 . S2CID 32792564 .
- ^ Пейн, КБ; Лангбауэр, WR; Томас, Э.М. (1986). «Инфразвуковые крики азиатского слона ( Elephas maximus )». Поведенческая экология и социобиология . 18 (4): 297–301. дои : 10.1007/BF00300007 . S2CID 1480496 .
- ^ Ларом, Д.; Гарстанг, М.; Пейн, К.; Распет, Р.; Линдек, М. (1997). «Влияние приземных атмосферных условий на дальность и территорию, достигаемую вокализацией животных» (PDF) . Журнал экспериментальной биологии . 200 (Часть 3): 421–431. дои : 10.1242/jeb.200.3.421 . ПМИД 9057305 .
- ^ О'Коннелл-Родвелл, CE; Вуд, Джей Ди; Родвелл, Техас; Пурия, С.; Партан, СР; Киф, Р.; Шрайвер, Д.; Арнасон, Британская Колумбия; Харт, Луизиана (2006). «Стада диких слонов ( Loxodonta africana ) реагируют на искусственно передаваемые сейсмические стимулы» (PDF) . Поведенческая и экологическая социобиология . 59 (6): 842–850. дои : 10.1007/s00265-005-0136-2 . S2CID 33221888 . Архивировано из оригинала (PDF) 3 декабря 2013 года.
- ^ О'Коннелл-Родвелл CE; Арнасон, Б.; Харт, Луизиана (2000). «Сейсмические свойства вокализации и передвижения азиатского слона ( Elephas maximus )». Журнал Акустического общества Америки . 108 (6): 3066–3072. Бибкод : 2000ASAJ..108.3066O . дои : 10.1121/1.1323460 . ПМИД 11144599 .
- ^ Плотник, Ю.М.; де Ваал, ФБМ и Рейсс, Д. (2006). «Самоузнавание в азиатском слоне» . Труды Национальной академии наук . 103 (45): 17053–17057. Бибкод : 2006PNAS..10317053P . дои : 10.1073/pnas.0608062103 . ПМЦ 1636577 . ПМИД 17075063 .
- ^ Ренш, Б. (1957). «Интеллект слонов». Научный американец . 196 (2): 44–49. Бибкод : 1957SciAm.196b..44R . doi : 10.1038/scientificamerican0257-44 .
- ^ Харт, Би Джей; Харт, Луизиана; МакКори, М.; Сарат, ЧР (2001). «Когнитивное поведение азиатских слонов: использование и модификация ветвей для переключения мух». Поведение животных . 62 (5): 839–847. дои : 10.1006/anbe.2001.1815 . S2CID 53184282 .
- ^ МакКомб, К.; Бейкер, Л.; Мосс, К. (2006). «Африканские слоны проявляют большой интерес к черепам и слоновой кости представителей своего вида» . Письма по биологии . 2 (1): 26–28. дои : 10.1098/rsbl.2005.0400 . ПМК 1617198 . ПМИД 17148317 .
- ^ Дуглас-Гамильтон, И.; Бхаллаа, С.; Виттемьера, Г.; Воллрата, Ф. (2006). «Поведенческие реакции слонов на умирающего и умершего матриарха» (PDF) . Прикладная наука о поведении животных . 100 (1): 87–102. дои : 10.1016/j.applanim.2006.04.014 . Архивировано из оригинала (PDF) 11 мая 2012 года.
- ^ Массон, Джеффри Мусаев ; Сьюзан Маккарти (1996). Когда слоны плачут: эмоциональная жизнь животных . Винтаж. п. 272. ИСБН 978-0-09-947891-1 .
- ^ Перейти обратно: а б с Гобуш, К.С.; Эдвардс, СТТ; Мейзелс, Ф.; Виттемайер, Г.; Бальфур, Д.; Тейлор, РД (2021). « Локсодонта африканская » . Красный список исчезающих видов МСОП . 2021 : e.T181008073A181022663 . Проверено 25 марта 2021 г.
- ^ Гобуш, К.С.; Эдвардс, СТТ; Мейзелс, Ф.; Виттемайер, Г.; Бальфур, Д. и Тейлор, Р.Д. (2021). « Локсодонта циклотис » . Красный список исчезающих видов МСОП . 2021 : e.T181007989A181019888 . Проверено 25 марта 2021 г.
- ^ Дуглас-Гамильтон, стр. 178–82.
- ^ Таулесс, ЧР; Дублин, штат ХТ; Блан, Джей-Джей; Скиннер, ДП; Дэниел, TE; Тейлор, доктор медицинских наук; Мейзелс, Ф.; Фредерик, HL; Буше, П. (2016). «Отчет о состоянии африканских слонов за 2016 год» (PDF) . База данных МСОП по африканским слонам. Архивировано (PDF) из оригинала 27 июня 2021 года . Проверено 5 июня 2021 г.
- ^ Перейти обратно: а б с д и ж Уильямс, К.; и др. (2020). « Большой слон » . Красный список исчезающих видов МСОП . 2020 . Проверено 3 июня 2020 г.
- ^ Перейти обратно: а б Дэниел, с. 174.
- ^ Перейти обратно: а б с д Мартин, стр. 202–07.
- ^ Перейти обратно: а б с Кристи, Б. (октябрь 2012 г.). «Поклонение слоновой кости» . Нэшнл Географик . Архивировано из оригинала 26 сентября 2015 года . Проверено 17 октября 2012 г.
- ^ Хикс, Селеста (19 марта 2013 г.). «86 слонов убиты в результате браконьерской резни в Чаде» . Хранитель . Архивировано из оригинала 25 сентября 2015 года . Проверено 23 сентября 2015 г.
- ^ Райан, Ф. (26 сентября 2015 г.). «Китай и США договорились о запрете на слоновую кость, стремясь положить конец незаконной торговле во всем мире» . Хранитель . Архивировано из оригинала 21 декабря 2019 года . Проверено 12 октября 2015 г.
- ^ Монтгомери, РА; Раупп, Дж; Мухвана, М; Гринлиф, А; Мудумба, Т; Мурути, П. (2022). «Эффективность мер по защите посевов от набегов слонов» . Амбио . 51 (3): 716–727. Бибкод : 2022Амбио..51..716М . дои : 10.1007/s13280-021-01587-x . ПМЦ 8800974 . ПМИД 34173175 .
- ^ Сукумар, с. 57.
- ^ Перейти обратно: а б Макнили, стр. 149–50.
- ^ Уайли, стр. 120–23.
- ^ Иса, с. 86.
- ^ Бист, СС; Чиран, СП; Чоудри, С.; Баруа, П.; Мисра, М.К. «Одомашненный азиатский слон в Индии» . Региональное отделение для Азии и Тихого океана. Архивировано из оригинала 1 июня 2015 года . Проверено 25 декабря 2012 г.
- ^ Фуллер, Томас (30 января 2016 г.). «Безработные слоны Мьянмы становятся нервными и тяжелее» . Нью-Йорк Таймс . Архивировано из оригинала 1 января 2022 года . Проверено 31 января 2016 г.
- ^ Лоханан, Роджер (февраль 2001 г.). «Ситуация со слонами в Таиланде и призыв к сотрудничеству» . ФАО . Архивировано из оригинала 4 апреля 2016 года . Проверено 31 января 2016 г.
- ^ Смит, стр. 152–54.
- ^ Перейти обратно: а б с д Уайли (2000), стр. 146–48.
- ^ Перейти обратно: а б Гриффин, Б. (2004). «Слоны: от священного к обыденному». В Джин Оой, К. (ред.). Юго-Восточная Азия: Историческая энциклопедия, от Ангкор-Вата до Восточного Тимора . Том. 1. Академик Блумсбери. стр. 487–489. ISBN 978-1-57607-770-2 .
- ^ Шошани, с. 168–69.
- ^ Перейти обратно: а б Таттл, стр. 184–88.
- ^ Стем, А. (28 февраля 2005 г.). «Смерть слонов в зоопарках вновь разжигает дебаты о животных: сторонники зоопарков ссылаются на сохранение, активисты ссылаются на замкнутые пространства» . MSNBC/Рейтер. Архивировано из оригинала 23 сентября 2015 года . Проверено 24 октября 2012 г.
- ^ Харрис, М.; Шервин, К.; Харрис, С. (10 ноября 2008 г.). «Заключительный отчет Defra о благополучии слонов» (PDF) . Бристольский университет. Архивировано (PDF) из оригинала 24 ноября 2014 года . Проверено 16 ноября 2011 г.
- ^ Мотт, М. (11 декабря 2008 г.). «Дикие слоны живут дольше, чем их собратья из зоопарка» . Национальные географические новости. Архивировано из оригинала 4 мая 2015 года . Проверено 24 октября 2012 г.
- ^ «Цирковые мифы: Настоящая жестокость под шапкой» . Общество защиты животных США. 25 сентября 2009 г. Архивировано из оригинала 5 сентября 2015 г. Проверено 24 октября 2012 г.
- ^ Пиклер, Н. (4 марта 2009 г.). «Генеральный директор цирка говорит, что слонов бьют, но не причиняют вреда» . Ассошиэйтед Пресс. Архивировано из оригинала 20 января 2013 года . Проверено 25 октября 2012 г.
- ^ Уайли, с. 142.
- ^ Карими, Фейт (2 мая 2016 г.). «Слоны братьев Ринглинг дают последнее представление» . Санкт-Петербург, Флорида: CNN. Архивировано из оригинала 22 сентября 2017 года . Проверено 21 сентября 2017 г.
- ^ Перейти обратно: а б Хагглер, Дж. (12 октября 2006 г.). «Поведение животных: Слоны-бродяги» . Независимый . Лондон. Архивировано из оригинала 14 мая 2008 года . Проверено 16 июня 2007 г.
- ^ Хайфилд, Р. (17 февраля 2006 г.). «Слоновья ярость: они тоже никогда не прощают» . Сидней Морнинг Геральд . Архивировано из оригинала 20 сентября 2013 года . Проверено 16 июня 2007 г.
- ^ Зиберт, К. (8 октября 2006 г.). "Слоновья авария?" . Нью-Йорк Таймс . Архивировано из оригинала 14 ноября 2012 года . Проверено 25 октября 2012 г.
- ^ Кришнан, Мурали (19 августа 2022 г.). «В Индии наблюдается всплеск смертоносных нападений слонов» . Немецкая волна . Проверено 11 июня 2023 г.
- ^ «Буйство индийского слона» . Новости Би-би-си . 24 декабря 1998 года. Архивировано из оригинала 26 мая 2007 года . Проверено 16 июня 2007 г.
- ^ «Пьяные слоны топчут деревню» . Новости Би-би-си . 21 октября 1999 года. Архивировано из оригинала 26 декабря 2007 года . Проверено 16 июня 2007 г.
- ^ «Пьяные слоны убили шесть человек» . Новости Би-би-си . 17 декабря 2002 года. Архивировано из оригинала 8 апреля 2013 года . Проверено 16 июня 2007 г.
- ^ Уайли, стр. 62–65.
- ^ Перейти обратно: а б с д Макнили, стр. 158–65.
- ^ Кингдон, с. 31.
- ^ Уайли, стр. 83–84.
- ^ Уайли, с. 79.
- ^ Сукумар, с. 87.
- ^ Сукумар, с. 64.
- ^ Сукумар, с. 62.
- ^ Перейти обратно: а б Ван Рипер; АБ (2002). Наука в массовой культуре: Справочное руководство . Гринвуд Пресс. стр. 73–75. ISBN 978-0-313-31822-1 .
- ^ Уайли, с. 90.
- ^ Уайли, стр. 27–28.
Библиография
- Кингдон, Дж. (29 декабря 1988 г.). Млекопитающие Восточной Африки: Атлас эволюции Африки, Том 3, Часть B: Крупные млекопитающие . Издательство Чикагского университета. ISBN 978-0-226-43722-4 . OCLC 468569394 . Архивировано из оригинала 21 марта 2023 года . Проверено 10 марта 2017 г.
- Шошани, Дж., изд. (2000). Слоны: величественные создания дикой природы . Книги с галочками. ISBN 978-0-87596-143-9 . OCLC 475147472 .
- Шошани, Дж.; Шошани, С.Л. «Что такое слон?». Слоны: величественные создания дикой природы . стр. 14–15.
- Шошани, Дж. «Сравнение живых слонов». Слоны: величественные создания дикой природы . стр. 36–51.
- Шошани, Дж. «Анатомия и физиология». Слоны: величественные создания дикой природы . стр. 66–80.
- Иса, П.С. «Сусло у азиатских слонов». Слоны: величественные создания дикой природы . стр. 85–86.
- Мосс, К. «Слонята: история двух полов». Слоны: величественные создания дикой природы . стр. 106–113.
- Пейн, КБ; Лангауэр, Всемирный банк «Слоновья коммуникация». Слоны: величественные создания дикой природы . стр. 116–123.
- Элтрингем, С.К. «Экология и поведение». Слоны: величественные создания дикой природы . стр. 124–127.
- Уайли, К.К. «Слоны как военные машины». Слоны: величественные создания дикой природы . стр. 146–148.
- Макнили, Дж. А. «Слоны как вьючные животные». Слоны: величественные создания дикой природы . стр. 149–150.
- Смит, К.Х. «Африканский центр приручения слонов». Слоны: величественные создания дикой природы . стр. 152–154.
- Макнили, Дж. А. «Слоны в фольклоре, религии и искусстве». Слоны: величественные создания дикой природы . стр. 158–165.
- Шошани, С.Л. «Знаменитые слоны». Слоны: величественные создания дикой природы . стр. 168–171.
- Дэниел, Дж. К. «Популяция азиатских слонов сегодня». Слоны: величественные создания дикой природы . стр. 174–177.
- Дуглас-Гамильтон, И. «Популяция африканских слонов сегодня». Слоны: величественные создания дикой природы . стр. 178–183.
- Таттл, компакт-диск «Слоны в неволе». Слоны: величественные создания дикой природы . стр. 184–193.
- Мартин, EB «Взлет и падение рынка слоновой кости». Слоны: величественные создания дикой природы . стр. 202–207.
- Шошани, Дж. «Зачем спасать слонов?». Слоны: величественные создания дикой природы . стр. 226–229.
- Сукумар, Р. (11 сентября 2003 г.). Живые слоны: эволюционная экология, поведение и охрана . Издательство Оксфордского университета, США. ISBN 978-0-19-510778-4 . OCLC 935260783 .
- Уайли, Д. (15 января 2009 г.). Слон . Книги реакции. ISBN 978-1-86189-615-5 . OCLC 740873839 . Архивировано из оригинала 21 марта 2023 года . Проверено 10 марта 2017 г.
Дальнейшее чтение
- Кэррингтон, Ричард (1958). Слоны: краткий отчет об их естественной истории, эволюции и влиянии на человечество . Чатто и Виндус. OCLC 911782153 .
- Нэнси, Сьюзен (2013). Развлекательные слоны: Агентство животных и бизнес американского цирка . Балтимор, Мэриленд: Издательство Университета Джонса Хопкинса.
- Сакс, Джон Годфри (1872). «Слепцы и слон» в Wikisource . Стихи Джона Годфри Сакса .
- Уильямс, Хиткот (1989). Священный слон . Нью-Йорк: Книги Гармонии. ISBN 978-0-517-57320-4 .
Внешние ссылки
 Определения из Викисловаря
Определения из Викисловаря  СМИ из Commons
СМИ из Commons  Новости из Викиновостей
Новости из Викиновостей  Цитаты из Wikiquote
Цитаты из Wikiquote  Тексты из Wikisource
Тексты из Wikisource  Учебники из Wikibooks
Учебники из Wikibooks  Ресурсы из Викиверситета
Ресурсы из Викиверситета  Туристическая информация от Wikivoyage
Туристическая информация от Wikivoyage
Роды отряда Хоботковые. |
|---|


