Азиатский слон
| Азиатский слон Временной диапазон: | |
|---|---|
 | |
| Клыкастый бык Национальный парк Бандипур , Карнатака , Индия | |
 | |
| Корова и подростки Национальный парк Яла , Шри-Ланка | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Сорт: | Млекопитающие |
| Заказ: | Хоботки |
| Семья: | Слоновиды |
| Род: | слон |
| Разновидность: | Э. Максимус [1] |
| Биномиальное имя | |
| Самый большой слон [1] | |
| Подвиды [1] | |
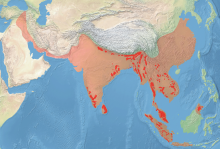 | |
исторический диапазон диапазон в начале 21 века | |

Азиатский слон ( Elephas maximus ), также известный как азиатский слон , — вид слонов , распространенный на Индийском субконтиненте и в Юго-Восточной Азии , от Индии на западе до Борнео на востоке и от Непала на севере до Суматры на юге. . три подвида Выделены — E. m. Максимус , Э.М. indicus и Э. м. суматранус . Азиатский слон отличается длинным хоботом с единственной пальцевой отделкой; большие бивни у самцов; сложенные по бокам большие уши, но меньшего размера в отличие от африканских слонов; и морщинистая серая кожа. Кожа более гладкая, чем у африканских слонов, и может быть депигментирована на туловище, ушах или шее. Взрослые самцы весят в среднем 4 т (3,9 длинных тонны; 4,4 коротких тонны), а самки - 2,7 т (2,7 длинных тонны; 3,0 коротких тонны).
Это один из трех ныне живущих видов слонов или слонов в мире, остальные — африканский кустарниковый слон и африканский лесной слон . Далее, азиатский слон — единственный ныне живущий вид рода Elephas . Это второй по величине вид слонов после африканского кустарникового слона. Он часто населяет луга , тропические вечнозеленые леса , полувечнозеленые леса, влажные лиственные леса , сухие лиственные леса и сухие терновые леса. Они травоядны и съедают около 150 кг (330 фунтов) растительности в день. Коровы и телята образуют группы, а самцы остаются поодиночке или образуют «холостяцкие группы» с другими самцами. В период размножения самцы временно присоединяются к женским группам для спаривания. Азиатские слоны имеют большой и хорошо развитый неокортекс головного мозга, они очень умны и самосознательны, способны демонстрировать поведение, связанное с горем, обучением, приветствием и т. д.
Азиатский слон — самое крупное живое наземное животное в Азии . С 1986 года азиатский слон занесен как находящийся под угрозой исчезновения в Красный список МСОП , поскольку за последние три поколения слонов, что составляет примерно 60–75 лет, популяция сократилась как минимум на 50 процентов. В первую очередь ему угрожают потеря среды обитания , деградация среды обитания , фрагментация и браконьерство . Дикие азиатские слоны живут около 60 лет. В то время как зарегистрировано, что самки слонов в неволе живут более 60 лет при содержании в полуестественной среде, азиатские слоны умирают в неволе в гораздо более молодом возрасте; Численность пленников сокращается из-за низкой рождаемости и высокого уровня смертности. Самыми ранними свидетельствами использования азиатских слонов в неволе являются гравюры на печатях цивилизации долины Инда, датированные III тысячелетием до нашей эры.
Таксономия
Карл Линней предложил Научное название Elephas maximus в 1758 году для слона с Цейлона . [4] Elephas indicus был предложен Жоржем Кювье в 1798 году, который описал слона из Индии. [5] Коенраад Джейкоб Темминк назвал слона с Суматры Elephas sumatranus в 1847 году. [6] В 1940 году Фредерик Наттер Чейзен классифицировал всех троих как подвиды азиатского слона. [7] Эти три подвида в настоящее время признаны действительными таксонами . [8] [9] Результаты филогеографического и морфологического анализа показывают, что шриланкийские и индийские слоны недостаточно различны, чтобы их можно было классифицировать как отдельные подвиды. [10]
- Шри-ланкийский слон ( E. maximus maximus Linnaeus, 1758 г. )

- Индийский слон ( E. maximus indicus Cuvier, 1798 )

- Суматранский слон ( E. maximus sumatranus Temminck, 1847 )

Шри-ланкийские слоны — самый крупный подвид. Цвет кожи у них темнее, чем у E. m. indicus и E. m. sumatranus с более крупными и отчетливыми пятнами депигментации на ушах, морде, туловище и животе. [11] Цвет кожи индийского слона обычно серый и светлее, чем у E. m. maximus , но темнее, чем у E. m. суматранус . [9]
Потенциальный четвертый подвид, слон Борнео ( Elephas maximus Borneensis ), встречается в Сабахе северо-восточных частях Борнео, в первую очередь в ( Малайзия ) , а иногда и на Калимантане ( Индонезия ). [12] Его предложил Паулес Дераниягала в 1950 году, который описал слона на иллюстрации, опубликованной в журнале National Geographic , но не как живого слона в соответствии с правилами Международного кодекса зоологической номенклатуры . [13] [14] Эти слоны, живущие на севере Борнео, меньше всех остальных подвидов, но у них были более крупные уши, более длинный хвост и прямые бивни. [15] Результаты генетического анализа показывают, что их предки отделились от материкового населения около 300 000 лет назад. [16] Исследование 2003 года с использованием анализа митохондриальной ДНК и микроспутниковых данных показало, что популяция слонов Борнео произошла от поголовья, происходящего из района Зондских островов , и предполагает, что популяция Борнео была отделена от других популяций слонов Юго-Восточной Азии. начиная с плейстоцена . [17]
Следующие азиатские слоны были предложены как вымершие подвиды, но теперь считаются синонимами индийского слона: [9]
- Сирийский слон ( E.m.asurus ), предложенный Дераниягалой на основе ископаемых останков и иллюстраций бронзового века . [18] [19]
- Китайский слон ( E.m.rubridens ), также предложенный Дераниягалой, на основе китайской бронзовой статуэтки. [20]
- Яванский слон ( E.m. sondaicus ), также предложенный Дераниягалой на основе иллюстрации резьбы на буддийском памятнике Боробудур . [21] [14]
Эволюция
Род Elephas , единственным живым представителем которого является азиатский слон, является ближайшим родственником вымерших мамонтов . По оценкам, эти две группы отделились друг от друга около 7 миллионов лет назад. [22] Слоны возникли в Африке к югу от Сахары в плиоцене и распространились по всей Африке, а затем распространились на южную половину Азии. [23] Самый ранний вид Elephas , Elephas ekorensis , известен из раннего плиоцена Восточной Африки, около 5-4,2 миллиона лет назад. [24] Самые старые остатки этого рода в Азии известны с холмов Сивалик на Индийском субконтиненте, датируются поздним плиоценом, около 3,6-3,2 миллиона лет назад, и отнесены к виду Elephas planifrons . [25] Предполагается, что современный азиатский слон произошел от вида Elephas hysudricus , который впервые появился в начале раннего плейстоцена около 2,6 миллиона лет назад и известен в первую очередь по останкам ранне- среднеплейстоценового возраста, найденным на Индийском субконтиненте. [26] Скелетные останки E. m. asurus были зарегистрированы на Ближнем Востоке : в Иране , Ираке , Сирии и Турции в периоды, датируемые как минимум 1800 г. до н.э. и, вероятно, 700 г. до н.э. [27]
Описание

В целом азиатский слон меньше африканского кустарникового слона и имеет самую высокую точку тела на голове. Спина выпуклая или ровная. Уши . маленькие, со спинными краями, загнутыми по бокам Имеет до 20 пар ребер и 34 хвостовых позвонка . Ступни имеют по пять гвоздевидных образований на каждой передней части стопы и по четыре на каждой задней ноге. [9] Лоб имеет две полусферические выпуклости, в отличие от плоской передней части африканских слонов. [28] Его длинный хобот , или хобот, имеет только один палец, в отличие от африканских слонов, у которых их два. [9] Следовательно, азиатские виды больше полагаются на то, что оборачивают пищу и сжимают ее в рот, а не хватают кончиком. Азиатские слоны обладают лучшей мышечной координацией и могут выполнять более сложные задачи. [29]

У коров обычно нет клыков ; если бивни - в данном случае называемые «кистями» - присутствуют, они едва заметны и видны только тогда, когда рот открыт. [30] Эмалевых пластинок у коренных зубов азиатских слонов больше и они расположены ближе друг к другу. [31] У некоторых быков также могут отсутствовать бивни; этих особей называют «махнами», и они особенно распространены среди популяции слонов Шри-Ланки. [32] Бивень слона высотой 11 футов (3,4 м), убитого сэром Виктором Бруком, имел длину 8 футов (2,4 м), окружность почти 17 дюймов (43 см) и весил 90 фунтов (41 кг). Однако вес этого бивня превышает вес более короткого бивня длиной около 6 футов (1,8 м), который весил 100 фунтов (45 кг), и, как сообщается, были бивни весом более 150 фунтов (68 кг). [33]

Цвет кожи обычно серый и может быть замаскирован почвой из-за пыли и валяния . Их морщинистая кожа подвижна и содержит множество нервных центров. Он более гладкий, чем у африканских слонов, и может быть лишен пигментации на туловище, ушах или шее. Толщина эпидермиса ; и дермы тела составляет в среднем 18 мм (0,71 дюйма) Толщина кожи на спине составляет 30 мм (1,2 дюйма), что обеспечивает защиту от укусов, ударов и неблагоприятных погодных условий. Его складки увеличивают площадь поверхности для рассеивания тепла. Они лучше переносят холод, чем чрезмерную жару. Температура кожи варьируется от 24 до 32,9 °C (от 75,2 до 91,2 °F). Температура тела в среднем составляет 35,9 ° C (96,6 ° F). [9]
Размер
В среднем, взрослые быки имеют высоту около 2,75 м (9,0 футов) в холке и вес 4,0 т (4,4 коротких тонны), тогда как коровы меньше — около 2,40 м (7,9 футов) в холке и 2,7 т. (3,0 коротких тонны) по весу. [34] [35] [36] Половой диморфизм размеров тела у азиатских слонов относительно менее выражен, чем у африканских кустарниковых слонов; при этом быки в среднем на 15% и 23% выше в первом и втором случаях соответственно. [34] Длина тела и головы, включая туловище, составляет 5,5–6,5 м (18–21 фут), длина хвоста — 1,2–1,5 м (3,9–4,9 фута). [9] Самый крупный из когда-либо зарегистрированных слонов-быков был застрелен махараджей Сусанга на холмах Гаро в Ассаме, Индия , в 1924 году. Он весил примерно 7 т (7,7 коротких тонн), имел высоту в холке 3,43 м (11,3 фута) и был Длина от головы до хвоста 8,06 м (26,4 фута). [34] [37] [38] Есть сообщения о более крупных особях ростом до 3,7 м (12 футов). [33]
Распространение и среда обитания

Азиатские слоны распространены по всему Индийскому субконтиненту и Юго-Восточной Азии , от Индии на западе до Борнео на востоке и от Непала на севере до Суматры на юге. [2] Они населяют луга, тропические вечнозеленые леса , полувечнозеленые леса, влажные лиственные леса , сухие лиственные леса и сухие терновые леса, а также культивируемые и вторичные леса и кустарники. В этом диапазоне типов среды обитания слоны встречаются от уровня моря до высоты более 3000 м (9800 футов). В восточных Гималаях на северо-востоке Индии они летом в нескольких местах регулярно поднимаются на высоту более 3000 м (9800 футов). [39]
В Бангладеш некоторые изолированные популяции сохранились на юго-востоке холмов Читтагонг в начале 1990-х годов. [40] В северном малайзийском национальном парке Джохор и Теренггану два азиатских слона, выслеженные с помощью технологии спутникового слежения, проводили большую часть своего времени во второстепенных или «вырубленных лесах»; они путешествовали 75% своего времени на расстоянии менее 1,5 км (0,93 мили) от источника воды. [41] В Китае азиатский слон сохранился только в префектурах Сишуанбаньна , Симао и Линьцан на юге Юньнани . По состоянию на 2020 год [update]Предполагаемая популяция составляла около 300 особей. [42]
По оценкам, по состоянию на 2017 год дикая популяция в Индии составляет почти три четверти нынешней популяции - 27 312 особей. [43] В 2019 году популяция азиатских слонов в Индии увеличилась примерно до 27 000–29 000 особей. [44] [45] По состоянию на 2019 год [update]Общая численность дикой популяции оценивалась в 48 323–51 680 особей. [46]
Экология и поведение

Азиатские слоны сумеречные . [9] Они классифицируются как мегатравоядные и потребляют до 150 кг (330 фунтов) растительных веществ в день. [47] Приблизительно от 50 до 75% дня посвящено еде. [48] Они универсальные кормушки, одновременно и травоядные , и браузерные . Известно, что они питаются как минимум 112 различными видами растений, чаще всего из отряда Malvales , а также семействами бобовых , пальмовых , осоки и настоящих трав . [49] Они больше пасутся в засушливый сезон, а кора составляет основную часть их рациона в прохладную часть этого сезона. [50] Они пьют не реже одного раза в день и никогда не отходят далеко от постоянного источника пресной воды. [9] Им требуется 80–200 литров воды в день, а для купания они используют еще больше. Иногда они очищают почву от глины или минералов. [51] [52]

Коровы и телята передвигаются вместе, группами, а быки, достигнув подросткового возраста, расходятся от своих матерей. Быки ведут одиночный образ жизни или образуют временные «холостяцкие группы». [53] Отряды «корова-телята», как правило, небольшие и обычно состоят из трех взрослых особей (скорее всего, родственных самок) и их потомства. [54] Также были зарегистрированы более крупные группы, насчитывающие до 15 взрослых самок. [55] Сезонные скопления 17 особей, включая телят и молодых особей, наблюдались в национальном парке Уда-Валаве на Шри-Ланке . До недавнего времени считалось, что азиатские слоны, как и африканские слоны, находятся под руководством пожилых самок или матриархов . Сейчас признано, что коровы образуют обширные и очень подвижные социальные сети с разной степенью связей между особями. [56] Социальные связи обычно слабее, чем у африканских кустарниковых слонов. [55] В отличие от африканских слонов, которые редко используют передние лапы для чего-либо, кроме рытья или соскабливания почвы, азиатские слоны более ловко используют ноги вместе с хоботом для манипулирования объектами. Иногда они могут быть известны своим агрессивным поведением. [28]

Записано, что азиатские слоны издают три основных звука: рычание, писк и фырканье. Рычание в своей основной форме используется для общения на близких расстояниях. При легком возбуждении рычание резонирует в туловище и переходит в грохот, а при общении на дальние расстояния перерастает в рев. Низкочастотное рычание является инфразвуковым и производится во многих контекстах. Писки бывают двух видов: щебетание и трубы. Щебетание состоит из нескольких коротких писков и сигнализирует о конфликте и нервозности. Трубы представляют собой удлиненные звуки повышенной громкости, издаваемые при сильном возбуждении. Фырканье сигнализирует об изменении активности и повышении громкости во время легкого или сильного возбуждения. В последнем случае, когда слон подпрыгивает кончиком хобота, он создает удары, которые служат индикатором угрозы. [57] Слоны способны различать звуки низкой амплитуды. [58]
Редко , что тигры регистрируется нападали и убивали телят, особенно если телята были разлучены со своими матерями, остались в стаде или остались сиротами. Взрослые особи в значительной степени неуязвимы для естественных хищников. Известен уникальный случай, когда азиатскую слониху-мать предположительно убили рядом со своим детенышем; однако эта версия является спорной. [59] [60] В 2011 и 2014 годах было зарегистрировано два случая, когда тигры успешно убили взрослых слонов; один - от одиночного тигра в национальном парке Джима Корбетта на 20-летней молодой взрослой слоновьей корове, а другой - на 28-летнем больном взрослом быке в национальном парке Казиранга дальше на восток, который был сбит и съеден несколькими тиграми. охота в сотрудничестве. [61] [62] Слоны, кажется, различают рычание более крупных хищников, таких как тигры, и более мелких хищников, таких как леопарды ; на леопардов они реагируют менее боязливо и более агрессивно. [63]
Воспроизведение

Размножение азиатских слонов можно объяснить выработкой и восприятием сигнальных соединений, называемых феромонами . Эти сигналы передаются через различные жидкости организма. Они обычно выделяются с мочой, но у мужчин они также обнаруживаются в секрете височных желез. [64] После интеграции и восприятия эти сигналы предоставляют получателю информацию о репродуктивном статусе отправителя. Если обе стороны готовы к размножению, происходит репродуктивное ритуальное поведение и продолжается процесс полового размножения. [65]

Быки будут сражаться друг с другом, чтобы получить доступ к течным коровам. Сильные драки за доступ к самкам случаются крайне редко. Быки достигают половой зрелости примерно в 12–15 лет. В возрасте от 10 до 20 лет быки ежегодно подвергаются явлению, известному как « муст ». Это период, когда уровень тестостерона в 100 раз выше, чем в периоды отсутствия мускуса, и они становятся агрессивными. Выделения, содержащие феромоны, происходят в этот период из парных височных желез, расположенных на голове между латеральным краем глаза и основанием уха. [66] Агрессивное поведение, наблюдаемое во время сусла, можно объяснить различным количеством фронталина (1,5-диметил-6,8-диоксабицикло[3.2.1]октана) на протяжении всего процесса созревания быков. Фронталин — это феромон , который впервые был выделен у короедов, но также может вырабатываться у быков как азиатских, так и африканских слонов. Соединение может выводиться через мочу, а также через височные железы быка, обеспечивая передачу сигналов. Во время сусла повышенная концентрация фронталина в моче быка сообщает слонам-самкам о репродуктивном статусе быка. [67]
Как и у других млекопитающих, секреция гормонов у слонов-самок регулируется эстральным циклом . Этот цикл регулируется скачками лютеинизирующего гормона , которые наблюдаются с интервалом в три недели. Этот тип эстрального цикла также наблюдался у африканских слонов, но не известно, что он влияет на других млекопитающих. Первый всплеск лютеинизирующего гормона не сопровождается выходом яйцеклетки из яичников. [68] Тем не менее, некоторые самки слонов во время этого всплеска все еще демонстрируют ожидаемый протокол спаривания. Самки слонов подают сигналы овуляции, используя половые феромоны . Было также обнаружено, что его основной компонент, (Z)-7-додецен-1-илацетат, является половым феромоном у многих видов насекомых. [69] [70] И у насекомых, и у слонов это химическое соединение используется в качестве аттрактанта, способствующего процессу спаривания. У слонов это химическое вещество выделяется при мочеиспускании, что способствует привлечению быков к спариванию. После обнаружения химическое вещество стимулирует сошниково-носовой орган быка, предоставляя тем самым информацию о зрелости самки. [71]
Репродуктивный обмен сигналами между слонами-самцами и самками передается через обонятельные сигналы в жидкостях организма. [65] У самцов повышение уровня фронталина во время сусла повышает их чувствительность к (Z)-7-додецен-1-илацетату, вырабатываемому самками слонов. [67] После восприятия рецепторами туловища следует последовательность ритуальных действий. Реакция самцов варьируется в зависимости от стадии развития и темперамента слона. [65] Этот процесс приема и обработки сигналов через хобот называется флеменом. Разница в движениях тела дает понять, заинтересован ли самец в спаривании с самкой, производящей секрет. [72] Бык, готовый к размножению, приближается к моче, и в некоторых случаях у него возникает эрекция. Бык, который не готов к размножению, будет робок и попытается дистанцироваться от сигнала. [65] Помимо репродуктивной коммуникации, хемосенсорная передача сигналов используется для облегчения однополых взаимодействий. Когда менее развитые самцы обнаруживают феромоны самца в сусле, они часто отступают, чтобы избежать контакта с агрессивным поведением. Также было замечено, что самки слонов общаются друг с другом посредством феромонов в моче. [65] Цель этого типа интерсексуального общения все еще исследуется. Однако существуют явные различия в силе сигнала и реакции приемника на разных стадиях эстрального цикла. [72]

Период беременности составляет 18–22 месяца, корова рожает одного теленка , лишь изредка двойню . Теленок полностью развивается к 19-му месяцу, но остается в утробе матери , чтобы вырасти и добраться до матери для кормления. При рождении теленок весит около 100 кг (220 фунтов) и находится на вскармливании до трех лет. После того, как самка рожает, она обычно не размножается снова, пока первый теленок не будет отнят от груди, в результате чего интервал между родами составляет четыре-пять лет. [73] [74] В этот период общение матери с теленком происходит преимущественно временными средствами. Однако известно, что у телят-самцов в молодом возрасте развиваются органы, вырабатывающие половые феромоны. Ранняя зрелость сошниково-носового органа позволяет неполовозрелым слонам производить и получать феромоны . [75] Маловероятно, что интеграция этих феромонов приведет к реакции Флемена у теленка. [72] Самки остаются в стаде, а половозрелых самцов прогоняют. [76]
Самки азиатских слонов достигают половой зрелости примерно в возрасте 10–15 лет и продолжают расти до 30 лет, тогда как самцы полностью созревают в возрасте старше 25 лет и постоянно растут на протяжении всей своей жизни. [77] [78] Средняя продолжительность жизни слонов составляет 60 лет в дикой природе и 80 лет в неволе, хотя в прошлом эта цифра преувеличивалась. [9] Продолжительность поколения азиатского слона составляет 22 года. [79]
Интеллект
У азиатских слонов очень большой и высокоразвитый неокортекс — черта, присущая людям , обезьянам и некоторым видам дельфинов . У них больший объем коры головного мозга, доступный для когнитивной обработки, чем у всех других существующих наземных животных. Результаты исследований показывают, что азиатские слоны обладают когнитивными способностями к использованию и изготовлению орудий, сходными с человекообразными обезьянами . [80] Они демонстрируют широкое разнообразие поведения, в том числе связанное с горем , обучением, алломотерингом , мимикрией , игрой, альтруизмом , использованием инструментов , состраданием , сотрудничеством , самосознанием , памятью и языком . [81] Сообщается, что слоны направляются в более безопасное место во время стихийных бедствий, таких как цунами и землетрясения, но данные двух шри-ланкийских слонов с ошейниками, установленными спутниками, показывают, что это может быть неправдой. [82] Несколько исследователей, изучающих когнитивные способности слонов и нейроанатомию, убеждены, что азиатские слоны очень умны и обладают самосознанием. [83] [84] [85] Другие оспаривают эту точку зрения. [86] [87]
Угрозы
Главной угрозой для азиатского слона сегодня являются потеря, деградация и фрагментация среды его обитания, что приводит к увеличению конфликтов между людьми и слонами. Азиатских слонов охотятся ради слоновой кости и множества других продуктов, включая мясо и кожу . [8] Спрос на кожу слона возрос в связи с тем, что она становится все более распространенным ингредиентом в традиционной китайской медицине. [88] [89]
Конфликт человека и слона


В некоторых частях Азии люди и слоны сосуществовали на протяжении тысячелетий. [90] В других районах люди и слоны вступают в конфликт, что приводит к насилию и, в конечном итоге, к перемещению слонов. [91] Основными причинами конфликта между людьми и слонами являются растущая численность населения, крупномасштабные проекты развития и плохое управление сверху вниз . Непосредственные причины включают утрату среды обитания из-за вырубки лесов , нарушение миграционных маршрутов слонов, расширение сельского хозяйства и незаконное вторжение на охраняемые территории. [92]
Уничтожение лесов в результате вырубки , вторжения, подсечек и поджогов , сменного земледелия и монокультурных плантаций деревьев представляют собой серьезную угрозу выживанию слонов. Конфликты между человеком и слонами происходят, когда слоны совершают набеги на посевы вахтовых земледельцев на полях, которые разбросаны по большой территории, перемежающейся лесами. Разграбление населенных пунктов — еще одна серьезная область конфликтов между людьми и слонами, происходящая в небольших лесных массивах, вторжения в среду обитания слонов и на пути миграции слонов. [93] Однако исследования, проведенные в Шри-Ланке, показывают, что традиционное подсечно-огневое земледелие может создать оптимальную среду обитания для слонов, создавая мозаику сукцессионной растительности. Популяции, населяющие небольшие фрагменты среды обитания, гораздо более склонны к конфликту с человеком. [94]
Такие разработки, как ограждение границы вдоль границы Индии и Бангладеш, стали серьезным препятствием для свободного передвижения слонов. [95] В Ассаме более 1150 человек и 370 слонов погибли в результате конфликта между людьми и слонами в период с 1980 по 2003 год. [93] По оценкам исследования 2010 года, только в Индии слоны ежегодно убивают более 400 человек и повреждают от 0,8 до 1 миллиона гектаров, что затрагивает не менее 500 000 семей по всей стране. [96] [97] [98] Более того, известно, что слоны уничтожают посевы на сумму до 2–3 миллионов долларов США в год. [99] Это оказывает серьезное влияние на благосостояние и средства к существованию местных сообществ, а также на будущее сохранение этого вида. [92] В таких странах, как Бангладеш и Шри-Ланка , азиатский слон является одним из самых страшных диких животных, хотя он менее смертоносен, чем другие местные животные, такие как ядовитые змеи (которые, по оценкам, унесли в Шри-Ланке в 30 раз больше жизней, чем слоны). [100] [101]
В целом азиатские слоны демонстрируют очень сложное, а иногда и непредсказуемое поведение. Большинство диких слонов стараются избегать людей, но если они будут застигнуты врасплох какой-либо предполагаемой физической угрозой, включая людей, они, скорее всего, бросятся в атаку. Особенно это касается самцов в сусле и самок с молодняком. Стрельба и другие подобные методы отпугивания, которые, как известно, эффективны против многих видов диких животных, включая тигров, могут сработать, а могут и не сработать со слонами, и могут даже ухудшить ситуацию. Слоны, которые в прошлом подвергались насилию со стороны людей, часто становятся «слонами-изгоями», которые регулярно нападают на людей без каких-либо провокаций. [102] [103] [104]
Браконьерство
Для слоновой кости
Спрос на слоновую кость в 1970-е и 1980-е годы, особенно в Восточной Азии , привел к безудержному браконьерству и серьезному сокращению поголовья слонов как в Африке, так и в Азии. В Таиланде до сих пор процветает нелегальная торговля живыми слонами и слоновой костью. Хотя количество открыто продаваемой слоновой кости существенно сократилось с 2001 года, в Таиланде по-прежнему существует один из крупнейших и наиболее активных черных рынков слоновой кости в мире. На рынок также поступают бивни слонов, отловленных в тайском браконьерстве; в период с 1992 по 1997 год по меньшей мере 24 слона-самца были убиты из-за бивней. [105]
До начала 1990-х годов вьетнамские мастера по изготовлению изделий из слоновой кости использовали исключительно кость азиатского слона из Вьетнама и соседних Лаоса и Камбоджи. До 1990 года туристов было мало, а низкий спрос на обработанную слоновую кость мог удовлетворяться за счет домашних слонов. Экономическая либерализация и рост туризма повысили спрос как у местных жителей, так и у посетителей на обработанную слоновую кость, что привело к масштабному браконьерству. [106]
Для кожи
Кожа азиатского слона используется в качестве ингредиента в китайской медицине , а также при изготовлении декоративных бус. Этой практике способствовала Государственная администрация лесного хозяйства Китая (SFA), которая выдала лицензии на производство и продажу фармацевтических продуктов, содержащих кожу слона, тем самым сделав торговлю законной. В 2010 году в лесу Мьянмы были найдены четыре слона со шкурой; В 2013 году браконьеры убили 26 слонов, а в 2016 году — 61. По данным неправительственной организации Elephant Family , Мьянма является основным источником слоновьих шкур, где с 2010 года быстро развивается кризис браконьерства. [107]
Болезнь
( Эндотелиотропный герпесвирус слона EEHV) является представителем рода Proboscivirus , новой клады, наиболее близкой к бетагерпевирусам млекопитающих . По состоянию на 2011 год он стал причиной смерти 70 слонов в зоопарках и диких азиатских слонов во всем мире, особенно молодых телят. [108] [109] несколько случаев гибели телят от эндотелиотропного герпесвируса слонов . В частности, в Мьянме зафиксировано [110] Слоновья шистосома — паразитическая трематода , которая использует азиатского слона в качестве окончательного хозяина. Двумя другими хозяевами могут быть индийский слон и большой однорогий носорог. [111]
Сохранение


Азиатский слон внесен в Приложение I СИТЕС . [8] Это типичный флагманский вид , используемый для стимулирования ряда природоохранных целей, включая сохранение среды обитания в ландшафтных масштабах, повышение осведомленности общественности о вопросах сохранения и мобилизацию в качестве популярного культурного символа как в Индии, так и на Западе. [112] [113] [92] Ключевым аспектом сохранения является соединение предпочтительных маршрутов передвижения азиатских слонов через территории с высоким растительным покровом и низкой плотностью населения. [114]
Всемирный день слона отмечается ежегодно 12 августа с 2012 года. Мероприятия организуются с целью распространения информации и привлечения людей к проблемам, с которыми сталкивается азиатский слон. [115] Зоопарки и партнеры по охране природы в Соединенных Штатах объявили август Месяцем осведомленности об азиатских слонах. [116]
В штате Карнатака в Индии обитает больше всего азиатских слонов из всех известных территорий, и он составляет 20% от общей численности населения страны. Распространение слонов в штате, по одной из оценок, составляет примерно 38 310 км2. 2 (14 790 квадратных миль). [117] По данным исследования 2013 года, около 10 000 слонов населяли Западные Гаты , и им в первую очередь угрожало браконьерство и фрагментация среды обитания. В качестве вероятной проблемы также было названо увеличение числа конфликтов с людьми. Планы сохранения направлены на создание коридоров для дикой природы, прекращение браконьерства на быков, а также на защиту земельных территорий или управление ими. [118] Проект «Слон» был инициирован в 1992 году как программа централизованного спонсирования (CSS) Министерством окружающей среды, лесов и изменения климата правительства Индии . Проект был инициирован с целью защиты индийского слона и среды его обитания, а также создания специальных заповедников для поддержания популяции слонов. [119]
Распространение слонов на Шри-Ланке составляет лишь две пятых того, что было в конце 19 — начале 20 веков. Таким образом, взаимодействия с людьми происходят гораздо чаще. Во время опроса 2003 года местные жители выразили некоторую форму неодобрения по отношению к сохранению азиатских слонов, поскольку фермеры считали их вредителями, однако большинство участников поддержали эту идею. [120]
В Китае азиатские слоны находятся под охраной первого уровня. В провинции Юньнань имеется 11 национальных и региональных заповедников. В общей сложности крытая охраняемая территория в Китае составляет около 510 000 га (1 300 000 акров). В 2020 году популяция азиатских слонов в Юньнани оценивалась примерно в 300 особей. Поскольку в последние годы вокруг охраняемых территорий возникли конфликты между людьми и дикими слонами, префектура Сишуанбаньна построила продовольственные базы и посадила бананы и бамбук, чтобы создать лучшую среду обитания. [42]
В Таиланде заповедник дикой природы Салакпра и национальный парк Тхам Тан Лот являются охраняемыми территориями, на которых, по данным за 2013 год, обитает около 250–300 слонов. [update]. [121] В последние годы Национальный парк столкнулся с проблемами из-за посягательств и чрезмерной эксплуатации. [122] В Индии Национальный совет дикой природы рекомендовал разрешить добычу угля в национальном парке Дехинг Паткай в апреле 2020 года. Это решение вызвало обеспокоенность среди студентов и активистов-экологов, которые начали онлайн-кампанию с целью остановить проект. [123]
В плену

Около половины мировой популяции слонов в зоопарках содержится в европейских зоопарках, где их продолжительность жизни составляет менее половины (18,9 года) средней продолжительности жизни сородичей (41,6 года) в охраняемых популяциях в странах ареала. Это несоответствие наиболее очевидно у азиатских слонов: детская смертность более чем в два-три раза выше, чем в бирманских лесных лагерях, а выживаемость взрослых особей в зоопарках за последние годы существенно не улучшилась. Одним из факторов риска для азиатских слонов в зоопарках является их перемещение из одного учреждения в другое, при этом раннее отделение от матери обычно приводит к дополнительным неблагоприятным последствиям. Еще одним фактором риска является рождение в зоопарке, а не завоз из дикой природы, при этом низкая выживаемость взрослых азиатов, рожденных в зоопарке, очевидно, передается пренатально или в раннем детстве. Вероятными причинами снижения выживаемости являются стресс и/или ожирение . [124] Проблемы со стопами обычно наблюдаются у содержащихся в неволе слонов. Это связано с отсутствием физических упражнений, долгим стоянием на твердом грунте и заражением в результате стояния в навозе. Многие из этих проблем поддаются лечению. Однако неправильное обращение может привести к серьезной инвалидности или смерти. [125]
Демографический анализ содержащихся в неволе азиатских слонов в Северной Америке показывает, что популяция не является самодостаточной. Смертность в первый год жизни составляет почти 30 процентов, а плодовитость в первые репродуктивные годы крайне низка. [126] Данные региональных племенных книг Северной Америки и Европы за период с 1962 по 2006 год были проанализированы на наличие отклонений в соотношении полов при рождении и смертности молодняка. Из 349 рожденных в неволе телят 142 умерли преждевременно. Они умерли в течение одного месяца после рождения, основными причинами были мертворождение и детоубийство, совершенное матерью теленка или одним из товарищей по выставке. Установлено, что соотношение полов мертворожденных в Европе имеет тенденцию к избытку мужского пола. [127]
Методы обработки
Молодых слонов отлавливают в дикой природе и незаконно ввозят в Таиланд из Мьянмы для использования в туристической индустрии; телят используют в основном в парках развлечений и обучают выполнять различные трюки для туристов. [105] Телят часто подвергают «взлому», который может включать в себя связывание, запирание, морение голодом, избиение и пытки; в результате две трети могут погибнуть. [128] Дрессировщики используют технику, известную как тренировочное давление , при которой «хендлеры используют лишение сна, голод и жажду, чтобы «сломать» дух слонов и сделать их покорными своим владельцам»; кроме того, проводники забивают слонам гвозди в уши и ноги. [129]
В культуре

Азиатский слон – национальное животное Таиланда и Лаоса. [130] [131] Он также был объявлен животным национального наследия Индии. [132] Кости азиатских слонов, раскопанные в Мохенджо-Даро в долине Инда, указывают на то, что они были приручены в цивилизации долины Инда и использовались для работы. Украшенные слоны также изображались на печатях и вылеплялись из глины. [133] азиатский слон стал осадной машиной , скакуном на войне, символом статуса , вьючным животным и возвышенной площадкой для охоты В исторические времена в Южной Азии . [134]

Азиатских слонов поймали в дикой природе и приручили для использования людьми. Их способность работать под руководством делает их особенно полезными для переноски тяжелых предметов. Их использовали, в частности, для перевозки леса в джунглях. Помимо рабочего использования, их использовали на войне, на церемониях и для перевозки. [135] Сообщается, что Боевые слоны до сих пор используются Армией независимости Качина (KIA) в штате Качин на севере Мьянмы против вооруженных сил Мьянмы. Для перевозки грузов КИА использует около четырех десятков слонов. [136]
Азиатский слон играет важную роль в культуре субконтинента и за его пределами, он занимает видное место в баснях Панчатантры и буддийских сказках Джатака . Они играют важную роль в индуизме : голова бога Ганеши — голова слона, а «благословения» храмового слона высоко ценятся. Слонов часто используют в процессиях , где животных украшают праздничными нарядами. [137]
Азиатский слон изображен в нескольких индийских рукописях и трактатах, среди которых выделяется Матанга Лила (слоновий спорт) Нилаканты. [137] Рукопись Хастивидьярнава родом из Ассама на северо-востоке Индии. [138] В бирманском, тайском и сингальском животном и планетарном зодиаке азиатский слон, как с клыками, так и без клыков, является четвертым и пятым животным зодиаком бирманцев , четвертым животным зодиаком тайцев и вторым животным зодиаком сингальского народа . Шри-Ланка. [139] [140] Точно так же слон - двенадцатое животное зодиака в зодиаке животных Дай народа Дай на юге Китая. [141]
См. также
Ссылки
- ^ Jump up to: Перейти обратно: а б Шошани, Дж. (2005). «Вид Elephas maximus » . В Уилсоне, Делавэр ; Ридер, Д.М. (ред.). Виды млекопитающих мира: таксономический и географический справочник (3-е изд.). Издательство Университета Джонса Хопкинса. п. 90. ИСБН 978-0-8018-8221-0 . OCLC 62265494 .
- ^ Jump up to: Перейти обратно: а б Уильямс, К.; Тивари, СК; Госвами, В.Р.; де Сильва, С.; Кумар, А.; Баскаран, Н.; Йогананд К. и Менон В. (2020). « Большой слон » . Красный список исчезающих видов МСОП . 2020 : e.T7140A45818198. doi : 10.2305/IUCN.UK.2020-3.RLTS.T7140A45818198.en . Проверено 15 января 2022 г.
- ^ «Приложения» . Конвенция о международной торговле видами дикой фауны и флоры, находящимися под угрозой исчезновения ( СИТЕС ). Архивировано из оригинала 5 декабря 2017 года . Проверено 6 июня 2022 г.
- ^ Линней, К. (1758). « Брута » . Карл Линней Система природы через три царства природы по классам, отрядам, родам, видам, с признаками, различиями, синонимами, местами . Том. Томус И. Холмиае: Impensis Direct. Лоран Сальвиус п. 33.
- ^ Кювье, Г. (1798). « Элефас индийский » . Элементарная таблица естественной истории животных . Париж: Бодуэн. стр. 148–149.
- ^ Темминк, CJ (1847). « Суматранский слон » . Общий обзор голландских владений в архипелаге Индия . Полет. Второй том. Лейде: А. Арнц и Комп. п. 91.
- ^ Чейзен, ФН (1940). «Список млекопитающих Малайзии, систематический список млекопитающих Малайского полуострова, Суматры, Борнео и Явы, включая прилегающие небольшие острова» (PDF) . Бюллетень музея Раффлз . 15 : 1–209. Архивировано из оригинала (PDF) 12 февраля 2020 года.
- ^ Jump up to: Перейти обратно: а б с д Чоудри, А.; Лахири Чоудхури, Дания; Десаи, А.; Дакворт, JW; Иса, ПС; Джонсингх, AJT; Фернандо, П.; Хеджес, С.; Гунавардена, М.; Курт, Ф.; Карант, Ю; Листер, А.; Менон, В.; Риддл, Х.; Рюбель, А.; Викраманаяке, Э.; Группа специалистов по азиатским слонам МСОП SSC (2008 г.). « Большой слон » . Красный список исчезающих видов МСОП . 2008 : e.T7140A12828813.
- ^ Jump up to: Перейти обратно: а б с д и ж г час я дж к Шошани, Дж.; Айзенберг, Дж. Ф. (1982). « Большой слон » (PDF) . Виды млекопитающих (182): 1–8. дои : 10.2307/3504045 . JSTOR 3504045 . Архивировано из оригинала (PDF) 30 апреля 2013 года.
- ^ Флейшер, Р.К.; Перри, Э.А.; Муралидхаран, К.; Стивенс, Э.Э. и Веммер, К.М. (2001). «Филогеография азиатского слона ( Elephas maximus ) на основе митохондриальной ДНК» . Эволюция . 55 (9): 1882–1892. дои : 10.1111/j.0014-3820.2001.tb00837.x . JSTOR 2680432 . ПМИД 11681743 .
- ^ Шошани, Дж . (2006). «Таксономия, классификация и эволюция слонов» . В Фаулере, Мэн; Микота, СК (ред.). Биология, медицина и хирургия слонов . Уайли-Блэквелл. стр. 3–14. ISBN 0813806763 . Архивировано из оригинала 20 октября 2022 года . Проверено 4 июня 2020 г.
- ^ Фэрроуз. «Борнейский слон: виды в заповедниках Всемирного земельного фонда» . Всемирный земельный фонд . Архивировано из оригинала 14 мая 2021 года . Проверено 26 января 2021 г.
- ^ Дераниягала 1955 , с. 123-124.
- ^ Jump up to: Перейти обратно: а б Крэнбрук, Э.; Пейн, Дж. и Лех, CMU (2007). «Происхождение слонов Elephas maximus L. с Борнео» (PDF) . Журнал музея Саравака . 63 : 95–125. Архивировано (PDF) из оригинала 27 сентября 2012 года . Проверено 13 марта 2011 г.
- ^ Нуржафарина О.; Марьяти, М.; Ахмад, Ах; Натан, С.; Пирсон, ХТ; Гусенс, Б. (2008). «Предварительное исследование морфометрии борнейского слона» (PDF) . Журнал тропической биологии и охраны природы . 4 (1): 109–113. Архивировано из оригинала (PDF) 7 марта 2012 года.
- ^ Фернандо, П.; Видья, ТНК; Пейн, Дж.; Стьюве, М.; Дэвисон, Г.; Альфред, Р.Дж.; Андау, П.; Боси Э. и Килборн А. (2003). «Анализ ДНК показывает, что азиатские слоны являются коренными жителями Борнео и поэтому являются приоритетной задачей для сохранения» . ПЛОС Биология . 1 (1): e6. doi : 10.1371/journal.pbio.0000006 . ПМК 176546 . ПМИД 12929206 .
- ^ Фернандо, П.; Видья ТНК; Пейн Дж.; Стьюве М.; Дэвисон Г.; Альфред, Р.Дж.; Андау, П.; Боси, Э.; Килборн, А.; Мельник, диджей (2003). «Анализ ДНК показывает, что азиатские слоны являются родными для Борнео и поэтому их сохранение является приоритетной задачей» . ПЛОС Биология . 1 (1): e6. дои : 10.1371/journal.pbio.0000006 . ПМК 176546 . ПМИД 12929206 .
- ^ Дераниягала 1955 , с. 116.
- ^ Гирдланд Флинк, Эль; Албайрак Э. и Листер А. (2018). «Генетический взгляд на вымершую популяцию азиатских слонов ( Elephas maximus ) на Ближнем Востоке» . Открытый четвертичный период . 4 (1): 1–9. дои : 10.5334/ок.36 . hdl : 2164/13525 .
- ^ Дераниягала 1955 , с. 125.
- ^ Дераниягала 1955 , с. 124.
- ^ Роланд, Надин; Райх, Дэвид; Маллик, Свапан; Мейер, Матиас; Грин, Ричард Э.; Георгиадис, Николас Дж.; Рока, Альфред Л.; Хофрейтер, Михаэль (21 декабря 2010 г.). «Последовательности геномной ДНК мастодонта и шерстистого мамонта раскрывают глубокое видообразование лесных слонов и слонов саванны» . ПЛОС Биология . 8 (12): e1000564. дои : 10.1371/journal.pbio.1000564 . ISSN 1545-7885 . ПМК 3006346 . ПМИД 21203580 .
- ^ Хейнс, Г. (1993). Мамонты, мастодонты и слоны: биология, поведение и летопись окаменелостей . Издательство Кембриджского университета. п. 8. ISBN 978-0521456913 . Архивировано из оригинала 17 марта 2024 года . Проверено 19 марта 2018 г.
- ^ Сандерс, Уильям Дж.; Хайле-Селассие, Йоханнес (июнь 2012 г.). «Новое собрание хоботных среднего плиоцена из района Ворансо-Милле, регион Афар, Эфиопия: таксономические, эволюционные и палеоэкологические соображения» . Журнал эволюции млекопитающих . 19 (2): 105–128. дои : 10.1007/s10914-011-9181-y . ISSN 1064-7554 . S2CID 254703858 . Архивировано из оригинала 17 марта 2024 года . Проверено 12 августа 2023 г.
- ^ Яннуччи, Алессио; Сарделла, Рафаэле (28 февраля 2023 г.). «Что означает событие «Слон-Эквус» сегодня? Размышления о событиях расселения млекопитающих вокруг границы плиоцена и плейстоцена и гибкой двусмысленности биохронологии» . Четвертичный период . 6 (1): 16. дои : 10,3390/quat6010016 . hdl : 11573/1680082 . ISSN 2571-550X .
- ^ Листер, Адриан М.; Диркс, Венди; Ассаф, Амнон; Чазан, Майкл; Гольдберг, Пол; Аппльбаум, Яаков Х.; Гринбаум, Натали; Хорвиц, Лиора Кольска (сентябрь 2013 г.). «Новые ископаемые останки слонов из южного Леванта: значение для истории эволюции азиатского слона» . Палеогеография, Палеоклиматология, Палеоэкология . 386 : 119–130. Бибкод : 2013PPP...386..119L . дои : 10.1016/j.palaeo.2013.05.013 . Архивировано из оригинала 2 января 2023 года . Проверено 12 августа 2023 г.
- ^ Чакырлар, Джанан; Икрам, Салима (3 мая 2016 г.). « Когда слоны дерутся, страдает трава». Власть, слоновая кость и сирийский слон» . Левант . 48 (2): 167–183. дои : 10.1080/00758914.2016.1198068 . ISSN 0075-8914 .
- ^ Jump up to: Перейти обратно: а б Клаттон-Брок, Джульетта (1987). Естественная история одомашненных млекопитающих . Издательство Кембриджского университета. п. 141. ИСБН 978-0-292-71532-5 .
- ^ Шошани, Дж., изд. (2000). Слоны: величественные создания дикой природы . Книги с галочками. стр. 38–41, 74–77. ISBN 978-0-87596-143-9 . OCLC 475147472 .
- ^ Челия, Карпагам; Сукумар, Раман (1 декабря 2013 г.). «Роль бивней, муста и размера тела в конкуренции между самцами азиатских слонов Elephas maximus» . Поведение животных . 86 (6): 1207–1214. дои : 10.1016/j.anbehav.2013.09.022 . ISSN 0003-3472 . Архивировано из оригинала 2 декабря 2018 года . Проверено 17 марта 2024 г.
- ^ Нэнси Э. Тодд (январь 2010 г.). «Качественное сравнение кранио-дентальной остеологии современных слонов Elephas Maximus (азиатский слон) и Loxodonta africana (африканский слон)» . Анатомическая запись . 293 (1): 62–73. дои : 10.1002/ar.21011 . ПМИД 19937635 . Архивировано из оригинала 21 мая 2024 года . Проверено 21 мая 2024 г.
- ^ «Махна» . База данных слонов . Архивировано из оригинала 21 мая 2024 года . Проверено 1 декабря 2023 г.
- ^ Jump up to: Перейти обратно: а б Лидеккер, Р. (1894). Королевская естественная история . Том. 2. Лондон: Фредерик Уорн и компания, стр. 522–523.
- ^ Jump up to: Перейти обратно: а б с Ларраменди, А. (2016). «Высота плеч, масса тела и форма хоботков» . Acta Palaeontologica Polonica . 61 (3): 537–574. дои : 10.4202/app.00136.2014 .
- ^ Сукумар, Р.; Джоши, Невада; Кришнамурти, В. (1988). «Рост азиатского слона». Труды: Науки о животных . 97 (6): 561–571. дои : 10.1007/BF03179558 . S2CID 84871322 .
- ^ Курт, Ф.; Кумарасингхе, Дж. К. (1998). «Замечания о росте тела и фенотипах азиатского слона Elephas maximus » . Акта Териологика . 5 (Приложение): 135–153. дои : 10.4098/AT.arch.98-39 .
- ^ Пиллаи, Н.Г. (1941). «О росте и возрасте слона». Журнал Бомбейского общества естествознания . 42 : 927–928.
- ^ Вуд, Г. (1983). Книга рекордов Гиннеса по фактам и подвигам животных . Энфилд, Миддлсекс: превосходная степень Книги Гиннеса. п. 17. ISBN 978-0-85112-235-9 .
- ^ Чоудри, Австралия (1999). «Статус и охрана азиатского слона Elephas maximus на северо-востоке Индии». Обзор млекопитающих . 29 (3): 141–173. дои : 10.1046/j.1365-2907.1999.00045.x .
- ^ Sukumar, R. (1993). The Asian Elephant: Ecology and Management (Second ed.). Cambridge: Cambridge University Press. p. 18. ISBN 978-0-521-43758-5. Archived from the original on 17 March 2024. Retrieved 15 December 2015.
- ^ Aini, S.; Sood, A. M.; Saaban, S. "Analysing elephant habitat parameters using GIS, remote sensing and analytic hierarchy process in Peninsular Malaysia". Pertanika Journal of Science & Technology. 23 (1): 37–50. Archived from the original on 17 May 2023. Retrieved 17 May 2023.
- ^ Jump up to: Jump up to: a b Shuang, F. (2020). "Solving a dilemma caused by elephants". ECNS. Archived from the original on 13 January 2021. Retrieved 6 September 2020.
- ^ "India has 27,312 elephants, census shows". The Hindu. 16 August 2017. Archived from the original on 3 December 2023. Retrieved 1 December 2023.
- ^ Srinivasaiah, Nishant M.; Vaidyanathan, Srinivas; Sukumar, Raman; Sinha, Anindya (2019). "Elephants on the Move: Implications for Human–Elephant Interaction". India International Centre Quarterly. 46 (3/4): 100–113. ISSN 0376-9771. JSTOR 26946286. Archived from the original on 21 May 2024. Retrieved 21 May 2024.
- ^ Baskaran, N.; Varma, S.; Sar, C.K.; Sukumar, R. (2011). "Current status of elephants in India". Gajah. 35: 47–54. Archived from the original on 21 May 2024. Retrieved 29 March 2024.
- ^ Menon, V.; Tiwari, S. (2019). "Population status of Asian elephants Elephas maximus and key threats". International Zoo Yearbook. 53 (1): 17–30. doi:10.1111/izy.12247. S2CID 209571148.
- ^ Samansiri, K. A. P.; Weerakoon, D. K. (2007). "Feeding Behaviour of Asian Elephants in the Northwestern Region of Sri Lanka" (PDF). Gajah. 2: 27–34. Archived (PDF) from the original on 18 November 2016. Retrieved 18 October 2011.
- ^ Sukumar, R. (July 2006). "A brief review of the status, distribution and biology of wild Asian elephants Elephas maximus". International Zoo Yearbook. 40 (1): 1–8. doi:10.1111/j.1748-1090.2006.00001.x. ISSN 0074-9664. Archived from the original on 18 March 2024. Retrieved 18 March 2024.
- ^ Sukumar, R. (1990). "Ecology of the Asian Elephant in southern India. II. Feeding habits and crop raiding patterns" (PDF). Journal of Tropical Ecology. 6: 33–53. doi:10.1017/S0266467400004004. S2CID 85129439. Archived from the original (PDF) on 12 October 2007.
- ^ Pradhan, N. M. B.; Wegge, P.; Moe, S. R.; Shrestha, A. K. (2008). "Feeding ecology of two endangered sympatric megaherbivores: Asian elephant Elephas maximus and greater one-horned rhinoceros Rhinoceros unicornis in lowland Nepal" (PDF). Wildlife Biology. 14: 147–154. doi:10.2981/0909-6396(2008)14[147:FEOTES]2.0.CO;2. S2CID 85964847. Archived (PDF) from the original on 13 March 2023. Retrieved 3 January 2023.
- ^ Sach, Fiona; Dierenfeld, Ellen S.; Langley-Evans, Simon C.; Hamilton, Elliott; Murray Lark, R.; Yon, Lisa; Watts, Michael J. (15 May 2020). "Potential bio-indicators for assessment of mineral status in elephants". Scientific Reports. 10 (1): 8032. Bibcode:2020NatSR..10.8032S. doi:10.1038/s41598-020-64780-0. ISSN 2045-2322. PMC 7229182. PMID 32415129.
- ^ Singh, A. P.; Sharma, Ramesh C. (24 September 2001). "Conflicts between linear developments and Asian elephants in sub - Himalayan zone of Uttranchal". Road Ecology Center. Archived from the original on 18 March 2024. Retrieved 18 March 2024.
- ^ McKay, G. M. (1973). "Behavior and ecology of the Asiatic elephant in southeastern Ceylon". Smithsonian Contributions to Zoology. 125 (125): 1–113. doi:10.5479/si.00810282.125. S2CID 128585445.
- ^ Fernando, P.; Lande, R. (2000). "Molecular genetic and behavioral analysis of social organization in the Asian elephant (Elephas maximus)". Behav Ecol Sociobiol. 48 (1): 84–91. doi:10.1007/s002650000218. S2CID 33707844.
- ^ Jump up to: Jump up to: a b de Silva, S.; Wittemyer, G. (2012). "A Comparison of Social Organization in Asian Elephants and African Savannah Elephants". International Journal of Primatology. 33 (5): 1125–1141. doi:10.1007/s10764-011-9564-1. S2CID 17209753.
- ^ de Silva, S.; Ranjeewa, A. D. G.; Kryazhimskiy, S. (2011). "The dynamics of social networks among female Asian elephants". BMC Ecology. 11 (1): 17. Bibcode:2011BMCE...11...17D. doi:10.1186/1472-6785-11-17. PMC 3199741. PMID 21794147.
- ^ Sukumar, R. (2003). The Living Elephants: Evolutionary Ecology, Behavior, and Conservation. Oxford: Oxford University Press. p. 142. ISBN 978-0-19-510778-4.
- ^ Heffner, R.; Heffner, H. (1980). "Hearing in the elephant (Elephas maximus)". Science. 208 (4443): 518–520. Bibcode:1980Sci...208..518H. doi:10.1126/science.7367876. PMID 7367876. Archived from the original on 17 March 2024. Retrieved 27 September 2013.
- ^ Karanth, K. U. & Nichols, J. D. (1998). "Estimation of tiger densities in India using photographic captures and recaptures" (PDF). Ecology. 79 (8): 2852–2862. doi:10.1890/0012-9658(1998)079[2852:EOTDII]2.0.CO;2. JSTOR 176521. Archived from the original (PDF) on 9 August 2017. Retrieved 7 December 2012.
- ^ "Tiger kills mother, baby elephant". Elephant News. 2006. Archived from the original on 14 July 2014.
- ^ "Tiger kills elephant in Corbett Reserve". The Hindu. 29 January 2011. Archived from the original on 8 February 2024. Retrieved 8 February 2024.
- ^ "Kaziranga elephant killed in tiger attack". The Times of India. 22 October 2014. Archived from the original on 8 February 2024. Retrieved 8 February 2024.
- ^ Thuppil, V.; Coss, R. G. (2013). "Wild Asian elephants distinguish aggressive tiger and leopard growls according to perceived danger". Biology Letters. 9 (5): 20130518. doi:10.1098/rsbl.2013.0518. PMC 3971691. PMID 24026347. Archived from the original on 14 July 2014.
- ^ Rasmussen, L. E. L. (1 June 1999). "Evolution of chemical signals in the Asian elephant,Elephas maximus: behavioural and ecological influences". Journal of Biosciences. 24 (2): 241–251. doi:10.1007/BF02941206. ISSN 0973-7138. S2CID 37477741. Archived from the original on 17 March 2024. Retrieved 27 November 2022.
- ^ Jump up to: Jump up to: a b c d e Schulte, Bruce A.; Bagley, Kathryn; Correll, Maureen; Gray, Amy; Heineman, Sarah M.; Loizi, Helen; Malament, Michelle; Scott, Nancy L.; Slade, Barbara E.; Stanley, Lauren; Goodwin, Thomas E.; Rasmussen, L. E. L. (2005). Mason, Robert T.; LeMaster, Michael P.; Müller-Schwarze, Dietland (eds.). "Assessing chemical communication in elephants". Chemical Signals in Vertebrates 10. Boston, MA: Springer US: 140–151. doi:10.1007/0-387-25160-X_18. ISBN 978-0-387-25160-8. Archived from the original on 22 November 2022. Retrieved 27 November 2022.
- ^ Jainudeen, M. R.; McKay, G. M.; Eisenberg, J. F. (1972). "Observation on musth in the domesticated Asiatic elephant (Elephas maximus)". Mammalia. 36 (2): 247–261. doi:10.1515/mamm.1972.36.2.247. S2CID 84275661.
- ^ Jump up to: Jump up to: a b Rasmussen, L.E.L (1 June 2003). "Frontalin: a Chemical Message of Musth in Asian Elephants (elephas maximus)". Chemical Senses. 28 (5): 433–446. doi:10.1093/chemse/28.5.433. PMID 12826539. Archived from the original on 3 November 2022. Retrieved 27 November 2022.
- ^ Brown, J.L.; Schmitt, D.L.; Bellem, A.; Graham, L.H.; Lehnhardt, J. (November 1999). "Hormone Secretion in the Asian Elephant (Elephas maximus): Characterizationof Ovulatory and Anovulatory Luteinizing Hormone Surges". Biology of Reproduction. 61 (5): 1294–1299. doi:10.1095/biolreprod61.5.1294. ISSN 0006-3363. PMID 10529277.
- ^ Rasmussen, L. E. L.; Lee, T. D.; Zhang, A. J.; Roelofs, W. L.; Daves, G. D. (1997). "Purification, identification, concentration and bioactivity of (Z)-7-dodecen-1-yl acetate: sex pheromone of the female Asian elephant, Elephas maximus". Chemical Senses. 22 (4): 417–437. doi:10.1093/chemse/22.4.417. PMID 9279465.
- ^ Rasmussen, L. E. L.; Lee, T. D.; Roelofs, W. L.; Zhang, A. J.; Daves, G. D. (1996). "Insect pheromone in elephants". Nature. 379 (6567): 684. Bibcode:1996Natur.379..684R. doi:10.1038/379684a0. PMID 8602213. S2CID 4330432.
- ^ Rasmussen, L.E.L (22 February 1996). "Insect Pheromones in Elephants" (PDF). Nature. 379 (6567): 684. Bibcode:1996Natur.379..684R. doi:10.1038/379684a0. PMID 8602213. S2CID 4330432. Archived (PDF) from the original on 3 November 2022. Retrieved 27 November 2022.
- ^ Jump up to: Jump up to: a b c Schulte, Bruce A.; LaDue, Chase A. (30 September 2021). "The Chemical Ecology of Elephants: 21st Century Additions to Our Understanding and Future Outlooks". Animals. 11 (10): 2860. doi:10.3390/ani11102860. ISSN 2076-2615. PMC 8532676. PMID 34679881.
- ^ Crawley, J. A. H.; Mumby, H. S.; Chapman, S. N.; Lahdenperä, M.; Mar, K. U.; Htut, W.; Thura Soe, A.; Aung, H. H.; Lummaa, V. (5 August 2017). "Is bigger better? The relationship between size and reproduction in female Asian elephants". Journal of Evolutionary Biology. 30 (10): 1836–1845. doi:10.1111/jeb.13143. ISSN 1010-061X. PMID 28703384. Archived from the original on 21 May 2024. Retrieved 21 May 2024.
- ^ Оуэн-Смит, Р. Норман (1988). Мегатравоядные: влияние очень крупных размеров тела на экологию . Издательство Кембриджского университета. стр. 145–150. ISBN 978-0-521-42637-4 . Архивировано из оригинала 21 мая 2024 года . Проверено 21 мая 2024 г.
- ^ Джонсон, Эдвард В.; Расмуссен, Лель (1 июля 2002 г.). «Морфологическая характеристика сошниково-носового органа новорожденного азиатского слона (Elephas maximus)» . Анатомическая запись . 267 (3): 252–259. дои : 10.1002/ar.10112 . ISSN 0003-276X . ПМИД 12115276 . S2CID 30345793 .
- ^ «Слоновья общественная организация» . ccrsl . Архивировано из оригинала 4 марта 2016 года . Проверено 29 августа 2015 г.
- ^ Фернандо, П.; Виджаякришнан, С.; Ранджива, AD; Пасторини, Дж. (2022). «Шкала размерно-возрастных классов азиатских слонов». Гаджа . 55 :30-39.
- ^ Мамби, HS; Чепмен, С.Н.; Кроули, Дж.А.; Мар, КУ; Хтут, В.; Тура Со, А.; Луммаа, В. (2015). «Различие детерминантного и неопределенного роста у млекопитающих-долгожителей» . Эволюционная биология BMC . 15 (1): 1-9. Бибкод : 2015BMCEE..15..214M . дои : 10.1186/s12862-015-0487-x . ПМЦ 4604763 . ПМИД 26464339 .
- ^ Пасифичи, М.; Сантини, Л.; Ди Марко, М.; Байсеро, Д.; Франкуччи, Л.; Гроттоло Маразини, Г.; Висконти, П.; Рондинини, К. (2013). «Продолжительность поколений млекопитающих» . Охрана природы . 5 : 87–94. doi : 10.3897/natureconservation.5.5734 . Архивировано из оригинала 26 января 2021 года . Проверено 20 ноября 2020 г.
- ^ Харт, Бенджамин Л.; Харт, Линетт А.; Маккой, Майкл; Сарат, Чехия (ноябрь 2001 г.). «Когнитивное поведение азиатских слонов: использование и модификация ветвей для переключения мух» . Поведение животных . 62 (5): 839–847. дои : 10.1006/anbe.2001.1815 . ISSN 0003-3472 . Архивировано из оригинала 5 февраля 2017 года . Проверено 18 марта 2024 г.
- ^ Барретт, Лиза П.; Бенсон-Амрам, Сара (август 2021 г.). «Множественные оценки личности и способности решать проблемы у содержащихся в неволе азиатских слонов (Elephas maximus) и африканских слонов саванны (Loxodonta africana)» . Журнал сравнительной психологии . 135 (3): 406–419. дои : 10.1037/com0000281 . ISSN 1939-2087 . ПМИД 34166041 .
- ^ Викраманаяке, Э.; Фернандо, П.; Леймгрубер, П. (2008). «Поведенческая реакция слонов со спутниковыми ошейниками на цунами на юге Шри-Ланки». Биотропика . 38 (6): 775–777. дои : 10.1111/j.1744-7429.2006.00199.x . S2CID 85358582 .
- ^ С., Дж.К.; Уильямс, Дж. Х. (октябрь 1950 г.). «Слон Билл» . Географический журнал . 116 (4/6): 229. Бибкод : 1950GeogJ.116..229S . дои : 10.2307/1789395 . ISSN 0016-7398 . JSTOR 1789395 . Архивировано из оригинала 21 мая 2024 года . Проверено 15 мая 2024 г.
- ^ Олдос, П. (2006). «Слоны видят себя в зеркале» . Новый учёный. Архивировано из оригинала 11 мая 2015 года . Проверено 24 августа 2017 г.
- ^ Пул, Дж. (1997). Слоны . Библиотека мировой жизни. п. 61. ИСБН 978-0-896-58357-3 .
- ^ Сандерсон, врач общей практики (1879 г.). Тринадцать лет среди диких зверей Индии . Лондон: WH Allen and Co., с. 80.
- ^ Ниссани, М. (2006). «Применяют ли азиатские слоны причинно-следственные связи к задачам, связанным с использованием инструментов?». Журнал экспериментальной психологии: процессы поведения животных . 31 (1): 91–96. дои : 10.1037/0097-7403.32.1.91 . ПМИД 16435969 .
- ^ Ван, Дэвид QH; Кэри, Мартин С. (7 августа 2014 г.). «Терапевтическое использование желчи животных в традиционной китайской медицине: этнофармакологический, биофизический, химический и медицинский обзор» . Всемирный журнал гастроэнтерологии . 20 (29): 9952–9975. дои : 10.3748/wjg.v20.i29.9952 . ISSN 1007-9327 . ПМЦ 4123376 . ПМИД 25110425 .
- ^ Нейман, Винсент; Шепард, Крис Р. (12 июля 2017 г.). «Этнозоологическая оценка животных, используемых торговцами традиционной медицины Мон в Кьяиктио, Мьянма» . Журнал этнофармакологии . 206 : 101–106. дои : 10.1016/j.jep.2017.05.010 . ISSN 0378-8741 . ПМИД 28506903 . Архивировано из оригинала 21 мая 2024 года . Проверено 18 марта 2024 г.
- ^ Лим, Теквин; Кампос-Арсеис, Ахимса (2022 г.). «Обзор экологических отношений человека и слона на Малайском полуострове: адаптация к сосуществованию» . Разнообразие . 14 (1): 36. дои : 10.3390/d14010036 . ISSN 1424-2818 .
- ^ де ла Торре, JA; Вонг, EP; Лехнер, AM; Зулайха, Н.; Завави, А.; Абдул-Патах, П.; Саабан, С.; Гуссенс, Б.; Кампос-Арсеис, А. (октябрь 2021 г.). «Конфликт будет – сельскохозяйственные ландшафты являются основной, а не маргинальной средой обитания азиатских слонов» . Охрана животных . 24 (5): 720–732. Бибкод : 2021AnCon..24..720D . дои : 10.1111/acv.12668 . ISSN 1367-9430 . Архивировано из оригинала 18 марта 2024 года . Проверено 18 марта 2024 г.
- ^ Jump up to: Перейти обратно: а б с Баруа, М. (2010). «Чья проблема? Репрезентации конфликта между человеком и слоном в индийских и международных СМИ» . Научная коммуникация . 32 : 55–75. дои : 10.1177/1075547009353177 . S2CID 143335411 . Архивировано из оригинала 8 октября 2013 года . Проверено 29 января 2012 г.
- ^ Jump up to: Перейти обратно: а б Чоудри, Австралия (2004). «Конфликты между людьми и слонами в Северо-Восточной Индии» (PDF) . Человеческое измерение дикой природы . 9 (4): 261–270. Бибкод : 2004HDW.....9..261C . дои : 10.1080/10871200490505693 . S2CID 145776270 . Архивировано (PDF) из оригинала 14 января 2014 года . Проверено 27 сентября 2013 г.
- ^ Фернандо, П. (2000). «Слоны в Шри-Ланке: прошлое, настоящее и будущее» (PDF) . Лорис . 22 (2): 38–44. Архивировано (PDF) из оригинала 14 января 2014 года . Проверено 8 декабря 2012 г.
- ^ Чоудри, Австралия (2007). «Влияние пограничного забора вдоль границы между Индией и Бангладеш на передвижение слонов» (PDF) . Гаджа . 26 : 27–30. Архивировано (PDF) из оригинала 25 апреля 2012 года . Проверено 18 октября 2011 г.
- ^ Рангараджан, М., Десаи, А., Сукумар, Р., Иса, П.С., Менон, В., Винсент, С., Гангули, С., Талукдар, Б.К., Сингх, Б., Мудаппа, Д., Чоудхари , С., Прасад, АН (2010). Гаджа: Обеспечение будущего слонов в Индии. Архивировано 19 июня 2013 года в Wayback Machine . Отчет Оперативной группы по слонам. Министерство окружающей среды и лесов, Нью-Дели.
- ^ Чоудхури, А. (2004). Конфликты между человеком и слонами на северо-востоке Индии . Человеческие измерения дикой природы, 9 (4), 261–270. дои : 10.1080/10871200490505693
- ^ Вудрофф, Рози; Тергуд, Саймон; Рабиновиц, Алан (2005), Рабиновиц, Алан; Вудрофф, Рози; Тергуд, Саймон (ред.), «Влияние конфликта между человеком и дикой природой на природные системы» , Люди и дикая природа, конфликт или сосуществование? , Биология сохранения, Кембридж: Издательство Кембриджского университета, стр. 1–12, ISBN. 978-0-521-82505-4 , заархивировано из оригинала 18 ноября 2019 года , получено 19 мая 2024 года.
- ^ Бист, СС (2006). «Охрана слонов в Индии – обзор» (PDF) . Гаджа . 25 : 27–35. Архивировано (PDF) из оригинала 3 июня 2013 года . Проверено 30 января 2012 г.
- ^ Райхан Саркер, AHM, и Рёскафт, Э. (2010). Конфликты между человеком и дикой природой и варианты управления в Бангладеш, с особым упором на азиатских слонов (Elephas maximus) . Международный журнал науки о биоразнообразии, экосистемных услугах и управлении, 6 (3–4), 164–175. дои : 10.1080/21513732.2011.554867
- ^ Сантьяпиллай, Чарльз; Виджеймохан, С.; Бандара, Ганга; Атурупана, Рукмали; Диссанаяке, Навин; Прочтите, Брюс (12 октября 2010 г.). «Оценка конфликта человека и слона в Шри-Ланке» . Джей. Дж. Наук. (Био. Науки) . 39 (1): 21. doi : 10.4038/cjsbs.v39i1.2350 .
- ^ Хоар, RE (1999). Детерминанты конфликта между человеком и слонами в мозаике землепользования . Журнал прикладной экологии, 36 (5), 689–700. дои : 10.1046/j.1365-2664.1999.00437.x
- ^ Расмуссен, ЛЕЛ (1999). Эволюция химических сигналов у азиатского слона Elephas maximus: поведенческие и экологические влияния . Журнал биологических наук, 24 (2), 241–251. дои : 10.1007/BF02941206
- ^ Сукумар, Р. (1995). Слоны-налетчики и мошенники . Естественная история, 104 (7), 52–61.
- ^ Jump up to: Перейти обратно: а б Стайлз, Д. (2009). Торговля слонами и слоновой костью в Таиланде. Архивировано 4 марта 2012 года в Wayback Machine TRAFFIC Southeast Asia, Петалинг-Джая, Селангор, Малайзия.
- ^ Стайлз, Д. (2009). «Состояние торговли слоновой костью в Таиланде и Вьетнаме» (PDF) . Бюллетень ДОРОЖНОГО ДВИЖЕНИЯ . 22 (2): 83–91. Архивировано из оригинала 17 апреля 2019 года . Проверено 8 декабря 2012 г.
- ^ Флинн, В.; Стюарт-Кокс, Б. и Мелидонис, К. (2018). со шкурой; Растущий аппетит к азиатским слонам (PDF) (Отчет). Лондон: Семья слонов. Архивировано (PDF) из оригинала 26 апреля 2018 г. Проверено 26 апреля 2018 г.
- ^ Латимер Э; Цзун Дж.К.; Хигганс С.Ю.; Ричман Л.К.; Хейворд Г.С. (2011). «Обнаружение и оценка новых герпесвирусов в рутинных и патологических образцах азиатских и африканских слонов: идентификация двух новых пробоскивирусов (EEHV5 и EEHV6) и двух новых гаммагерпесвирусов (EGHV3B и EGHV5)» . Ветеринарная микробиология . 147 (1–2): 28–41. дои : 10.1016/j.vetmic.2010.05.042 . ПМЦ 2976818 . ПМИД 20579821 .
- ^ Рид С.Э., Хильдебрандт Т.Б., Маркс Н., Хант М., Тай Н., Рейнс Дж.М., Шафтенаар В., Фикель Дж. (2006). «Эндотелиотропный вирус герпеса слона (EEHV). Первый смертельный случай, подтвержденный ПЦР, в Азии». Ветеринарный ежеквартальный журнал . 28 (2): 61–64. дои : 10.1080/01652176.2006.9695209 . ПМИД 16841568 . S2CID 25537428 .
- ^ Oo, Zaw Min; Aung, Ye Htut; Aung, Tin Tun; San, Nyo; Tun, Zaw Min; Hayward, Gary S.; Zachariah, Arun (January 2020). "Elephant Endotheliotropic Herpesvirus Hemorrhagic Disease in Asian Elephant Calves in Logging Camps, Myanmar". Emerging Infectious Diseases. 26 (1): 63–69. doi:10.3201/eid2601.190159. ISSN 1080-6040. PMC 6924905. PMID 31855135.
- ^ Devkota, R.; Brant, S.V.; Thapa, A.; Loker, E.S. (2014) [Published online 2012]. "Sharing schistosomes: the elephant schistosome Bivitellobilharzia nairi also infects the greater one-horned rhinoceros (Rhinoceros unicornis) in Chitwan National Park, Nepal". Journal of Helminthology. 88 (1): 32–40. doi:10.1017/S0022149X12000697. PMID 23113960. S2CID 7039264. Archived from the original on 6 January 2018. Retrieved 18 March 2024.
- ^ Bowen-Jones, E. & Entwistle, A. (2002). "Identifying appropriate flagship species: The importance of culture and local contexts". Oryx. 36 (2): 189–195. doi:10.1017/S0030605302000261.
- ^ Barua, M.; Tamuly, J. & Ahmed, R.A. (2010). "Mutiny or Clear Sailing? Examining the Role of the Asian Elephant as a Flagship Species". Human Dimensions of Wildlife. 15 (2): 145–160. Bibcode:2010HDW....15..145B. doi:10.1080/10871200903536176. S2CID 143021377. Archived from the original on 8 October 2013. Retrieved 6 May 2012.
- ^ Vasudev, D.; Goswami, V.; Srinivas, N.; La Nam Syiem, B.; Sarma, A. (2021). "Identifying important connectivity areas for the wide-ranging Asian elephant across conservation landscapes of Northeast India". Diversity and Distributions. 27 (12): 2510–2523. Bibcode:2021DivDi..27.2510V. doi:10.1111/ddi.13419. JSTOR 48632844. S2CID 244245085.
- ^ Shelden, D. (2020). "World Elephant Day 2020". The Atlantic. Archived from the original on 7 September 2020. Retrieved 6 September 2020.
- ^ Shelden, D. (2020). "OKC Zoo to host 2nd annual Asian Elephant Awareness Month". The City Sentinel. Archived from the original on 26 September 2020. Retrieved 6 September 2020.
- ^ Madhusudan, M.D.; Sharma, Narayan; Raghunath, R.; Baskaran, N.; Bipin, C.M.; Gubbi, Sanjay; Johnsingh, A.J.T.; Kulkarni, Jayant; Kumara, Honnavalli N.; Mehta, Prachi; Pillay, Rajeev; Sukumar, Raman (July 2015). "Distribution, relative abundance, and conservation status of Asian elephants in Karnataka, southern India". Biological Conservation. 187: 34–40. Bibcode:2015BCons.187...34M. doi:10.1016/j.biocon.2015.04.003. ISSN 0006-3207. Archived from the original on 21 May 2024. Retrieved 20 May 2024.
- ^ Baskaran, Nagarajan (26 October 2013). "An overview of Asian Elephants in the Western Ghats, southern India: implications for the conservation of Western Ghats ecology". Journal of Threatened Taxa. 5 (14): 4854–4870. doi:10.11609/jott.o3634.4854-70. ISSN 0974-7893. Archived from the original on 21 May 2024. Retrieved 20 May 2024.
- ^ "Project Elephant". Government of India. Archived from the original on 21 May 2024. Retrieved 1 April 2024.
- ^ Bandara, Ranjith; Tisdell, Clem (1 April 2003). "Comparison of rural and urban attitudes to the conservation of Asian elephants in Sri Lanka: empirical evidence". Biological Conservation. 110 (3): 327–342. doi:10.1016/S0006-3207(02)00241-0. ISSN 0006-3207. Archived from the original on 21 May 2024. Retrieved 20 May 2024.
- ^ Jeerawat na Thalang (2020). "It's risky, but elephants are feeling frisky". Bangkok Post. Archived from the original on 2 October 2021. Retrieved 6 September 2020.
- ^ Elephant Conservation Network. Proposed Eastern Extension to Salakpra Wildlife Sanctuary (PDF). The Elephant Conservation Network (Report). Archived (PDF) from the original on 21 September 2020. Retrieved 6 September 2020.
- ^ Parashar, U. (2020). "Assam students, environment activists launch online campaign to stop coal mining in elephant reserve". Hindustan Times. Archived from the original on 29 June 2020. Retrieved 6 September 2020.
- ^ Clubb, R.; Rowcliffe, M.; Lee, P.; Mar, K. U.; Moss, C. & Mason, G. J. (2008). "Compromised Survivorship in Zoo Elephants" (PDF). Science. 322 (5908): 1649. Bibcode:2008Sci...322.1649C. doi:10.1126/science.1164298. hdl:1893/974. PMID 19074339. S2CID 30756352. Archived (PDF) from the original on 14 May 2014. Retrieved 9 December 2012.
- ^ Roocroft, Alan; Oosterhuis, James (December 2000). "Foot Care for Captive Elephants". The Elephant's Foot: 1-5. doi:10.1002/9780470292150.ch5. ISBN 978-0-8138-2820-6. Archived from the original on 21 May 2024. Retrieved 19 May 2024.
- ^ Wiese, R. J. (2000). "Asian elephants are not self-sustaining in North America". Zoo Biology. 19 (5): 299–309. doi:10.1002/1098-2361(2000)19:5<299::AID-ZOO2>3.0.CO;2-Z.
- ^ Saragusty, J.; Hermes, R.; Goritz, F.; Schmitt, D.L. & Hildebrandt, T. B. (2009). "Skewed birth sex ratio and premature mortality in elephants". Animal Reproduction Science. 115 (1–4): 247–254. doi:10.1016/j.anireprosci.2008.10.019. PMID 19058933. Archived from the original on 21 June 2013. Retrieved 9 December 2012.
- ^ "Tourism driving illegal elephant trade in Burma and Thailand – video". The Guardian. 24 July 2012. Archived from the original on 2 December 2016. Retrieved 11 December 2016.
- ^ Hile, J. (2002). "Activists Denounce Thailand's Elephant "Crushing" Ritual". National Geographic Society. Archived from the original on 18 October 2002. Retrieved 1 October 2014.
Just before dawn in the remote highlands of northern Thailand, west of the village Mae Jaem, a four-year-old elephant bellows as seven village men stab nails into her ears and feet. She is tied up and immobilized in a small, wooden cage. Her cries are the only sounds to interrupt the otherwise quiet countryside. The cage is called a "training crush." It's the centerpiece of a centuries-old ritual in northern Thailand designed to domesticate young elephants. In addition to beatings, handlers use sleep-deprivation, hunger, and thirst to "break" the elephants' spirit and make them submissive to their owners.
- ^ Han, Jianghua (3 July 2019). "The Study of Thai Elephant Culture Based on the "Elephant Metaphors" in Thai Idioms". Comparative Literature: East & West. 3 (2): 148–162. doi:10.1080/25723618.2019.1701306. ISSN 2572-3618.
- ^ "Thailand and Laos elephant". South East Asia Globe. 28 February 2022. Archived from the original on 1 October 2023. Retrieved 1 December 2023.
- ^ Sukumar, Raman (26 July 2016). "Iconic Fauna of Heritage Significance in India" (PDF). Indian Journal of History of Science. 51 (2.2). doi:10.16943/ijhs/2016/v51i2.2/48450. ISSN 0019-5235. Archived (PDF) from the original on 4 February 2023. Retrieved 21 May 2024.
- ^ Baker, Iljas; Kashio, Masakazu (2002). Giants on Our Hands: Proceedings of the International Workshop on the Domesticated Asian Elephant, Bangkok, Thailand, 5 to 10 February 2001. FAO Regional Office for Asia and the Pacific. ISBN 978-974-90757-1-5. Archived from the original on 21 May 2024. Retrieved 21 May 2024.
- ^ Rangarajan, M. (2001). "The Forest and the Field in Ancient India". India's Wildlife History. Delhi: Permanent Black. pp. 1–10. ISBN 978-8-178-24140-1. Archived from the original on 17 March 2024. Retrieved 19 March 2018.
- ^ Liehrmann, Océane; Crawley, Jennie A. H.; Seltmann, Martin W.; Feillet, Sherine; Nyein, U. Kyaw; Aung, Htoo Htoo; Htut, Win; Lahdenperä, Mirkka; Lansade, Léa; Lummaa, Virpi (29 July 2021). "Handler familiarity helps to improve working performance during novel situations in semi-captive Asian elephants". Scientific Reports. 11 (1): 15480. doi:10.1038/s41598-021-95048-w. ISSN 2045-2322. PMC 8322261. PMID 34326446.
- ^ Winn, P. (2017). "War elephants still exist. But only in one forbidding place". Public Radio International (PRI). Archived from the original on 1 March 2017. Retrieved 1 March 2017.
- ^ Jump up to: Jump up to: a b Rajamangalam), Nīlakaṇṭha (1985). The Elephant-lore of the Hindus: The Elephant-sport (Matanga-lila) of Nilakantha. Motilal Banarsidass Publications. pp. 1-10. ISBN 978-8-12080-005-2.
- ^ Sharma, Jayeeta (August 2011). Empire's Garden: Assam and the Making of India. Duke University Press. p. 25. ISBN 978-0-82235-049-1. Archived from the original on 24 May 2022. Retrieved 15 May 2024.
- ^ Upham, E. (1829). The History and Doctrine of Budhism: Popularly Illustrated: with Notices of the Kappooism, Or Demon Worship, and of the Bali, Or Planetary Incantations, of Ceylon. London: R. Ackermann.
- ^ Ling, Trevor (March 1971). "Sinhalese buddhism in regent anthropological writing: Some implications". Religion. 1 (1): 49–59. doi:10.1016/0048-721X(71)90007-8. ISSN 0048-721X. Archived from the original on 15 May 2024. Retrieved 15 May 2024.
- ^ "Chinese Zodiac". Warriortours. Archived from the original on 15 September 2018. Retrieved 12 June 2017.
Bibliography
- Deraniyagala, Paulus Edward Pieris (1955). Some Extinct Elephants, Their Relatives, and the Two Living Species. Department of National Museums, Ceylon.
Further reading
- Bandara, Ranjith & Clem Tisdell (2004). "The net benefit of saving the Asian elephant a policy and contingent valuation study" (PDF). Ecological Economics. 48 (1): 93–107. Bibcode:2004EcoEc..48...93B. doi:10.1016/j.ecolecon.2003.01.001.
- Miall, L. C.; Greenwood, F. (1878). Anatomy of the Indian Elephant. London: Macmillan and Co.
External links
- Save Elephant Foundation
- International Elephant Foundation
- ElefantAsia: Protecting the Asian elephant
- Asian Elephants at the Zoological Gardens of the World
- Elephant Information Repository Archived 11 July 2017 at the Wayback Machine
- WWF—Asian elephant species profile
- National Zoo Facts on Asian Elephant and a Webcam of the Asian Elephant exhibit
- Environmental Investigation Agency: Illegal Wildlife Trade : Elephants

- IUCN Red List endangered species
- Elephants
- EDGE species
- Endangered animals
- Endangered biota of Asia
- Extant Pliocene first appearances
- Fauna of South Asia
- Fauna of Southeast Asia
- Herbivorous mammals
- Mammals described in 1758
- Mammals of Asia
- National symbols of India
- National symbols of Laos
- National symbols of Thailand
- Pliocene mammals of Asia
- Quaternary animals of Asia
- Species endangered by habitat fragmentation
- Species endangered by habitat loss
- Taxa named by Carl Linnaeus