Нейроанатомия
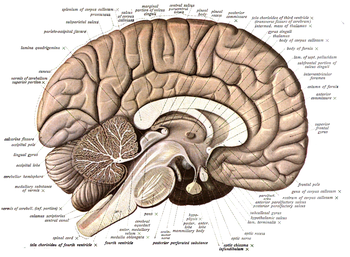
Нейроанатомия — наука о строении и организации нервной системы . В отличие от животных с радиальной симметрией , чья нервная система состоит из распределенной сети клеток, животные с двусторонней симметрией имеют отдельные, определенные нервные системы. Поэтому их нейроанатомия лучше изучена. У позвоночных нервная система разделена на внутреннюю структуру головного и спинного мозга (вместе называемые центральной нервной системой или ЦНС) и ряд нервов, которые соединяют ЦНС с остальной частью тела (известный как периферическая нервная система) . система или ПНС). Разрушение и идентификация конкретных частей нервной системы имеет решающее значение для выяснения того, как она работает. Например, большая часть того, что узнали нейробиологи, получена в результате наблюдения за тем, как повреждения или «поражения» определенных областей мозга влияют на поведение или другие нервные функции.
Информацию о составе нервной системы животных, отличных от человека, см. в разделе Нервная система . Информацию о типичном строении нервной системы Homo sapiens см. в разделе « Мозг человека или периферическая нервная система» . В этой статье обсуждается информация, имеющая отношение к изучению нейроанатомии.
История [ править ]

Первым известным письменным свидетельством изучения анатомии человеческого мозга является древнеегипетский документ, Папирус Эдвина Смита . [1] В Древней Греции интерес к мозгу начался с работы Алкмеона , который, по-видимому, препарировал глаз и связал мозг со зрением. Он также предположил, что мозг, а не сердце, является органом, управляющим телом (то, что стоики назвали гегемониконом ) и что чувства зависят от мозга. [2]
Споры о гегемониконе продолжались среди древнегреческих философов и врачей очень долгое время. [3] Те, кто выступал за мозг, также часто вносили свой вклад в понимание нейроанатомии. Герофил и Эрасистрат Александрийские, пожалуй, оказали наибольшее влияние своими исследованиями, включавшими в себя препарирование человеческого мозга, подтверждение различия между головным мозгом и мозжечком , а также идентификацию желудочков и твердой мозговой оболочки . [4] [5] Греческий врач и философ Гален также решительно выступал за мозг как орган, ответственный за ощущения и произвольные движения , о чем свидетельствуют его исследования нейроанатомии быков , берберийских обезьян и других животных. [3] [6]
Культурное табу на вскрытие человека продолжалось еще несколько сотен лет после этого, что не принесло существенного прогресса в понимании анатомии мозга или нервной системы. Однако Папа Сикст IV эффективно возобновил изучение нейроанатомии, изменив папскую политику и разрешив вскрытие человека. Это привело к всплеску новой деятельности художников и учёных эпохи Возрождения. [7] такие как Мондино де Луцци , Беренгарио да Карпи и Жак Дюбуа , а кульминацией стали работы Андреаса Везалия . [8] [9]
В 1664 году Томас Уиллис , врач и профессор Оксфордского университета, ввёл термин «неврология», когда опубликовал свой текст « Анатомия головного мозга» , который считается основой современной нейроанатомии. [10] Последующие триста пятьдесят лет привели к появлению большого количества документации и исследований нервной системы.
Состав [ править ]
На тканевом уровне нервная система состоит из нейронов , глиальных клеток и внеклеточного матрикса . И нейроны, и глиальные клетки бывают разных типов (см., например, раздел «Нервная система» в списке различных типов клеток в организме взрослого человека ). Нейроны — это клетки нервной системы, обрабатывающие информацию: они чувствуют окружающую среду, общаются друг с другом посредством электрических сигналов и химических веществ, называемых нейротрансмиттерами, которые обычно действуют через синапсы (тесные контакты между двумя нейронами или между нейроном и мышечной клеткой; обратите внимание). возможны также экстрасинаптические эффекты, а также выброс нейромедиаторов в нервное внеклеточное пространство), и производят наши воспоминания, мысли и движения. Глиальные клетки поддерживают гомеостаз, производят миелин (олигодендроциты, шванновские клетки) и обеспечивают поддержку и защиту нейронов головного мозга. Некоторые глиальные клетки ( астроциты ) в ответ на стимуляцию могут даже распространять межклеточные волны кальция на большие расстояния и выделять глиотрансмиттеров в ответ на изменение концентрации кальция. Рубцы от ран головного мозга в основном содержат астроциты. Внеклеточный матрикс также обеспечивает поддержку на молекулярном уровне клеткам мозга, транспортируя вещества в кровеносные сосуды и обратно.
На уровне органов нервная система состоит из областей мозга, таких как гиппокамп у млекопитающих или грибовидные тела плодовой мухи . [11] Эти области часто имеют модульную структуру и играют особую роль в общих системных путях нервной системы. Например, гиппокамп имеет решающее значение для формирования воспоминаний, как и многие другие области мозга. Периферическая нервная система также содержит афферентные или эфферентные нервы , которые представляют собой пучки волокон, исходящих из головного и спинного мозга или от сенсорных или моторных периферических ганглиев и неоднократно разветвляющихся, иннервирующих каждую часть тела. Нервы состоят в основном из аксонов или дендритов нейронов (аксонов в случае эфферентных двигательных волокон и дендритов в случае афферентных сенсорных волокон нервов), а также множества мембран, которые окружают их и разделяют на нервные пучки .
Нервная система позвоночных делится на центральную и периферическую нервную систему. Центральная нервная система (ЦНС) состоит из головного мозга , сетчатки и спинного мозга , тогда как периферическая нервная система (ПНС) состоит из всех нервов и ганглиев (пакетов периферических нейронов) за пределами ЦНС, которые соединяют ее с остальная часть тела. ПНС подразделяется на соматическую и вегетативную нервные системы. Соматическая нервная система состоит из «афферентных» нейронов, которые передают сенсорную информацию от соматических органов чувств (тела) в ЦНС, и «эфферентных» нейронов, которые передают двигательные инструкции произвольным мышцам тела. Автономная нервная система может работать под контролем ЦНС или без нее (поэтому ее называют «автономной»), а также имеет два подразделения, называемые симпатическим и парасимпатическим , которые важны для передачи двигательных команд основным внутренним органам организма. таким образом контролируя такие функции, как сердцебиение, дыхание, пищеварение и слюноотделение. Вегетативные нервы, в отличие от соматических, содержат только эфферентные волокна. Сенсорные сигналы, поступающие из внутренних органов в ЦНС через соматические сенсорные нервы (например, висцеральная боль) или через некоторые определенные черепные нервы (например, химиочувствительные или механические сигналы).
Ориентация по нейроанатомии [ править ]

В анатомии в целом и нейроанатомии в частности для обозначения ориентации и местоположения используется несколько наборов топографических терминов, которые обычно относятся к оси тела или мозга (см. Анатомические термины местоположения ). Ось ЦНС часто ошибочно считают более или менее прямой, но на самом деле она всегда имеет два вентральных изгиба (шейный и головной изгиб) и дорсальный изгиб (мостовой изгиб), все из-за дифференцированного роста во время эмбриогенеза. В нейроанатомии чаще всего используются следующие пары терминов:
- Дорсальный и вентральный: Дорсальный относится более или менее к верхней или верхней стороне мозга, которая символизируется пластиной дна, а вентральный — к нижней или нижней стороне. Эти дескрипторы первоначально использовались для спинной и вентральной части тела, спины и живота; брюхо большинства животных обращено к земле; в прямоугольной позе человека брюшная часть тела оказывается впереди, а спинная часть становится задней. Случай с головой и мозгом своеобразен, поскольку брюхо не переходит в голову должным образом, если только мы не предположим, что рот представляет собой расширенный элемент брюшка. Поэтому в обиходе те части мозга, которые лежат близко к основанию черепа и через него к ротовой полости, называются вентральными, т. е. на его нижней или нижней стороне, как определено выше, тогда как дорсальные части располагаются ближе к охватывающий свод черепа. Ссылка на верхнюю и нижнюю пластинки мозга менее подвержена путанице, а также позволяет нам следить за упомянутыми выше осевыми изгибами. Таким образом, дорсальный и вентральный — относительные термины в мозге, точное значение которых зависит от конкретного местоположения.
- Ростральный и каудальный: ростральный в общей анатомии относится к передней части тела (к носу или роструму на латыни), а каудальный относится к хвостовому концу тела (к хвосту; cauda на латыни). Ростро-каудальный размер головного мозга соответствует его оси длины, которая проходит через указанные изгибы от каудального конца спинного мозга к ростральному концу примерно на перекресте зрительных нервов. У прямоходящего человека термины «высший» и «низший», по существу, относятся к этому рострокаудальному измерению, поскольку в вертикальном положении оси нашего тела и мозга примерно ориентированы вертикально. Однако у всех позвоночных развивается очень выраженный вентральный излом нервной трубки, который все еще обнаруживается во взрослой центральной нервной системе, известный как цефалический изгиб . Последняя изгибает ростральную часть ЦНС под углом 180 градусов относительно каудальной части, в месте перехода между передним мозгом (ось, заканчивающаяся рострально на перекресте зрительных нервов) и стволом мозга и спинным мозгом (ось примерно вертикальная, но включая дополнительные незначительные изломы моста и шейных изгибов) Эти изгибные изменения осевого размера проблематичны при попытке описать относительное положение и плоскости срезов головного мозга. Существует множество литературы, в которой ошибочно игнорируются осевые изгибы и предполагается относительно прямая ось мозга.
- Медиальный и латеральный: медиальный относится к расположению близко или относительно ближе к средней линии (дескриптор « медиана» означает положение точно на средней линии). Латеральная – противоположная (позиция более или менее отделена от средней линии).
Обратите внимание, что такие дескрипторы (дорсальный/вентральный, ростральный/каудальный; медиальный/латеральный) являются скорее относительными, чем абсолютными (например, можно сказать, что латеральная структура лежит медиально по отношению к чему-то другому, что лежит еще более латерально).
Обычно используемые термины для плоскостей ориентации или плоскостей сечения в нейроанатомии: «сагиттальная», «поперечная» или «корональная», «аксиальная» или «горизонтальная». Опять же в этом случае ситуация у плавающих, ползающих или четвероногих (лежачих) животных иная, чем у Человека или других прямоходящих видов, из-за измененного положения оси. Из-за осевых изгибов мозга ни одна плоскость сечения никогда не достигает полной серии сечений в выбранной плоскости, поскольку некоторые сечения неизбежно получаются косыми или даже перпендикулярными к ней, когда они проходят через изгибы. Опыт позволяет разглядеть порции, которые в результате нарезаются по желанию.
- Срединно-сагиттальная плоскость делит тело и мозг на левую и правую половины; сагиттальные сечения, как правило, параллельны этой срединной плоскости, перемещаясь по медиально-латеральному измерению (см. изображение выше). Термин «сагиттальный» этимологически относится к срединному шву между правой и левой теменными костями черепа, известному классически как стреловидный шов, поскольку он выглядит примерно как стрела из-за слияния с другими швами ( sagitta ; стрелка на латыни).
- Плоскость сечения, ортогональная оси любой удлиненной формы, в принципе считается поперечной (например, поперечный разрез пальца или позвоночного столба); если оси длины нет, нет возможности определить такие сечения или существуют бесконечные возможности. Поэтому поперечные участки тела у позвоночных параллельны ребрам, ортогональным позвоночному столбу, представляющему ось тела как у животных, так и у человека. Мозг также имеет внутреннюю продольную ось – ось первичной удлиненной нервной трубки – которая становится в значительной степени вертикальной при прямохождении человека, подобно оси тела, за исключением ее рострального конца, как отмечалось выше. Это объясняет, что поперечные участки спинного мозга примерно параллельны нашим ребрам или земле. Однако это верно только для спинного мозга и ствола головного мозга, поскольку переднемозговой конец нервной оси изгибается во время раннего морфогенеза в хиазматический гипоталамус, где и заканчивается; соответственно изменяется ориентация истинных поперечных сечений, которые уже не параллельны ребрам и земле, а перпендикулярны им; Неосведомленность об этой морфологической особенности мозга (присутствующей у всех без исключения позвоночных) вызвала и вызывает множество ошибочных представлений о передних отделах мозга. Признавая своеобразие ростральных поперечных срезов, традиция ввела для них другое обозначение, а именно корональные срезы. Корональные отделы делят передний мозг от рострального (спереди) до каудального (заднего), образуя ряд, ортогональный (поперечный) к локальной изогнутой оси. Эту концепцию нельзя осмысленно применить к стволу головного мозга и спинному мозгу, поскольку там корональные срезы становятся горизонтальными по отношению к осевому измерению и параллельны оси. В любом случае концепция «корональных» срезов менее точна, чем концепция «поперечных», поскольку часто используются плоскости корональных срезов, которые не совсем ортогональны ростральному концу оси мозга. Этот термин этимологически связан с венечным швом кранума, а значит, и с местом ношения коронок (лат. corona означает корона). Неясно, какой тип короны имелся в виду первоначально (возможно, просто диадема), и это, к сожалению, приводит к неоднозначности в плоскости сечения, определяемой просто как корональная.
- В настоящее время считается, что корональная плоскость, проходящая через человеческую голову и мозг, параллельна лицу (плоскость, в которой королевская корона сидит на его голове, не совсем параллельна лицу, и эта концепция экспортируется на животных, менее наделенных фронтальной частью, чем мы). очевидно, еще более противоречив, но здесь имеется неявная ссылка на венечный шов черепа, который образуется между лобными и височными/теменными костями, создавая своего рода конфигурацию диадемы, которая примерно параллельна лицу). Таким образом, плоскости коронального сечения по существу относятся только к голове и мозгу, где диадема имеет смысл, а не к шее и телу ниже.
- Горизонтальные участки по определению выровнены (параллельны) горизонту. У плавающих, ползающих и четвероногих животных сама ось тела горизонтальна, и, таким образом, по длине спинного мозга проходят горизонтальные участки, отделяющие вентральную часть от спинной. Горизонтальные сечения ортогональны как поперечному, так и сагиттальному сечениям и теоретически параллельны оси длины. Из-за осевого изгиба головного мозга (переднего мозга) истинные горизонтальные сечения в этой области ортогональны корональным (поперечным) сечениям (как и горизонт относительно лица).
Согласно этим соображениям, три направления пространства представлены именно сагиттальной, поперечной и горизонтальной плоскостями, тогда как корональные срезы могут быть поперечными, косыми или горизонтальными, в зависимости от того, как они относятся к оси мозга и ее извилинам.
Инструменты [ править ]
Современные разработки в области нейроанатомии напрямую связаны с технологиями, используемыми для проведения исследований . Поэтому необходимо обсудить различные доступные инструменты. Многие гистологические методы, используемые для изучения других тканей, могут быть применены и к нервной системе. Однако есть некоторые методы, разработанные специально для изучения нейроанатомии.
Окрашивание клеток [ править ]
В биологических системах окрашивание — это метод, используемый для усиления контраста определенных особенностей микроскопических изображений.
При окраске по Нисслю используются основные анилиновые красители для интенсивного окрашивания кислых полирибосом в шероховатой эндоплазматической сети , которой много в нейронах. Это позволяет исследователям различать различные типы клеток (например, нейроны и глия ), а также формы и размеры нейронов в различных областях цитоархитектуры нервной системы .
Классическое окрашивание по Гольджи использует дихромат калия и нитрат серебра для избирательного заполнения осадком хромата серебра нескольких нервных клеток (нейронов или глии, но в принципе любые клетки могут реагировать аналогичным образом). Эта так называемая процедура импрегнации хроматом серебра окрашивает полностью или частично тела клеток и нейриты некоторых нейронов — дендриты , аксоны — в коричневый и черный цвет, что позволяет исследователям проследить их пути вплоть до их тончайших концевых ветвей в срезе нервной ткани, благодаря прозрачности, обусловленной отсутствием окрашивания большинства окружающих клеток. В настоящее время материал, пропитанный аппаратом Гольджи, адаптирован для электронно-микроскопической визуализации неокрашенных элементов, окружающих окрашенные отростки и тела клеток, что добавляет дополнительную разрешающую способность.
Гистохимия [ править ]
Гистохимия использует знания о свойствах биохимических реакций химических компонентов мозга (включая, в частности, ферменты), чтобы применять избирательные методы реакций для визуализации того, где они происходят в мозге, а также любых функциональных или патологических изменений. Это в первую очередь применимо к молекулам, связанным с производством и метаболизмом нейротрансмиттеров, но также применимо и ко многим другим направлениям химиоархитектуры или химической нейроанатомии.
Иммуноцитохимия - это особый случай гистохимии, в которой используются селективные антитела против различных химических эпитопов нервной системы для избирательного окрашивания определенных типов клеток, аксональных пучков, нейропилей, глиальных отростков или кровеносных сосудов или специфических внутрицитоплазматических или внутриядерных белков и других иммуногенетических молекул. например, нейромедиаторы. Иммунореагировавшие белки транскрипционных факторов демонстрируют геномное считывание с точки зрения транслируемого белка. Это значительно увеличивает возможности исследователей различать разные типы клеток (например, нейроны и глия ) в различных областях нервной системы.
В гибридизации in situ используются синтетические зонды РНК, которые избирательно прикрепляются (гибридизируются) к комплементарным транскриптам мРНК экзонов ДНК в цитоплазме, чтобы визуализировать считывание генома, то есть различать активную экспрессию генов с точки зрения мРНК, а не белка. Это позволяет идентифицировать гистологически (in situ) клетки, участвующие в производстве генетически закодированных молекул, которые часто представляют собой дифференцированные или функциональные признаки, а также молекулярные границы, разделяющие отдельные домены мозга или популяции клеток.
маркеры Генетически закодированные
Экспрессируя различное количество красных, зеленых и синих флуоресцентных белков в мозге, так называемая мутантная мышь с « мозговой дугой » позволяет комбинаторно визуализировать множество различных цветов в нейронах. Это помечает нейроны настолько уникальными цветами, что их часто можно отличить от своих соседей с помощью флуоресцентной микроскопии , что позволяет исследователям составить карту локальных связей или взаимного расположения (мозаики) между нейронами.
Оптогенетика использует трансгенную конститутивную и сайт-специфическую экспрессию (обычно у мышей) заблокированных маркеров, которые можно избирательно активировать путем освещения световым лучом. Это позволяет исследователям очень избирательно изучать связь аксонов в нервной системе.
Неинвазивная визуализация мозга
Магнитно-резонансная томография широко используется для исследования структуры и функционирования неинвазивного мозга у здоровых людей. Важным примером является диффузионно-тензорная визуализация , которая основана на ограниченной диффузии воды в тканях для создания изображений аксонов. В частности, вода движется быстрее в направлении аксонов, что позволяет сделать вывод об их структуре.
Вирусные методы [ править ]
Некоторые вирусы могут реплицироваться в клетках головного мозга и пересекать синапсы. Таким образом, вирусы, модифицированные для экспрессии маркеров (таких как флуоресцентные белки), можно использовать для отслеживания связей между областями мозга через многочисленные синапсы. [12] Два вируса-индикатора, которые реплицируются и распространяются транснейронально/транссинаптически, представляют собой вирус простого герпеса типа 1 (ВПГ). [13] и рабдовирусы . [14] Вирус простого герпеса использовался для отслеживания связей между мозгом и желудком, чтобы исследовать области мозга, участвующие в висцеро-сенсорной обработке. [15] В другом исследовании вирус простого герпеса вводился в глаз, что позволяло визуализировать оптический путь от сетчатки к зрительной системе . [16] Примером вируса-индикатора, который реплицируется из синапса в сому, является вирус псевдобешенства . [17] Используя вирусы псевдобешенства с различными флуоресцентными репортерами, модели двойной инфекции могут анализировать сложную синаптическую архитектуру. [18]
Методы на основе красителей [ править ]
В методах аксонального транспорта используются разнообразные красители (варианты пероксидазы хрена, флуоресцентные или радиоактивные маркеры, лектины, декстраны), которые более или менее жадно поглощаются нейронами или их отростками. Эти молекулы избирательно транспортируются антероградно (от сомы к окончаниям аксонов) или ретроградно (от окончаний аксонов к соме), что свидетельствует о наличии первичных и побочных связей в головном мозге. Эти «физиологические» методы (поскольку используются свойства живых неповрежденных клеток) можно комбинировать с другими процедурами и по существу заменили более ранние процедуры, изучающие дегенерацию поврежденных нейронов или аксонов. Детальные синаптические связи можно определить с помощью корреляционной электронной микроскопии.
Коннектомика [ править ]
Серийная секционная электронная микроскопия получила широкое развитие для использования при изучении нервной системы. Например, первое применение серийной блочной сканирующей электронной микроскопии было на кортикальной ткани грызунов. [19] Реконструкция схемы на основе данных, полученных с помощью этого высокопроизводительного метода, является сложной задачей, и научная игра Citizen EyeWire для помощи исследованиям в этой области была разработана .
нейроанатомия Вычислительная
Это область, в которой используются различные методы визуализации и вычислительные методы для моделирования и количественной оценки пространственно-временной динамики нейроанатомических структур как в нормальных, так и в клинических популяциях.
Модельные системы [ править ]
Помимо человеческого мозга , существует множество других животных, чей мозг и нервная система широко изучались в качестве модельных систем , в том числе мыши, рыбки данио , [20] плодовая мушка , [21] и разновидность круглых червей под названием C. elegans . Каждая из них имеет свои преимущества и недостатки как модельная система. Например, нервная система C. elegans чрезвычайно стереотипна от одного червя к другому. Это позволило исследователям с помощью электронной микроскопии составить карту путей и связей всех 302 нейронов этого вида. Плодовая мушка широко изучается отчасти потому, что ее генетика очень хорошо изучена и ею легко манипулировать. Мышь используется потому, что ее мозг, как млекопитающего, по структуре больше похож на наш (например, у нее шестислойная кора , однако ее гены легко модифицируются, а ее репродуктивный цикл относительно быстр).
Caenorhabditis elegans [ править ]

Мозг у некоторых видов небольшой и простой, например у червя- нематоды , у которого строение тела довольно простое: трубка с полой кишечной полостью, идущая от рта к заднему проходу, и нервный тяж с расширением ( ганглием ). для каждого сегмента тела, с особенно большим ганглием спереди, называемым мозгом. Нематода Caenorhabditis elegans изучалась из-за ее важности в генетике. [22] В начале 1970-х годов Сидней Бреннер выбрал ее в качестве модельной системы для изучения того, как гены контролируют развитие, включая развитие нейронов. Одним из преимуществ работы с этим червем является то, что нервная система гермафродита содержит ровно 302 нейрона, всегда находящихся в одних и тех же местах, образующих одинаковые синаптические связи у каждого червя. [23] Команда Бреннера разрезала червей на тысячи ультратонких срезов и сфотографировала каждый срез под электронным микроскопом, а затем визуально сопоставила волокна от среза к срезу, чтобы составить карту каждого нейрона и синапса во всем теле и получить полный коннектом нематоды. [24] Ни для одного другого организма не существует ничего, что могло бы приблизиться к такому уровню детализации, и эта информация была использована для проведения множества исследований, которые без нее были бы невозможны. [25]
Drosophila melanogaster [ править ]
Drosophila melanogaster — популярное экспериментальное животное, поскольку его легко массово культивировать в дикой природе, у него короткий период генерации и легко доступны мутантные животные.
У членистоногих есть центральный мозг с тремя отделами и большими оптическими долями позади каждого глаза для обработки визуальной информации. Мозг плодовой мухи содержит несколько миллионов синапсов по сравнению с по меньшей мере 100 миллиардами в человеческом мозге. Примерно две трети мозга дрозофилы отвечает за обработку зрительной информации .
Томас Хант Морган начал работать с дрозофилой в 1906 году, и эта работа принесла ему Нобелевскую премию по медицине 1933 года за определение хромосом как вектора наследования генов. Из-за большого количества инструментов, доступных для изучения генетики дрозофил, они стали естественным объектом для изучения роли генов в нервной системе. [26] Геном был секвенирован и опубликован в 2000 году. Около 75% известных генов болезней человека имеют узнаваемое совпадение с геномом плодовых мух. Дрозофила используется в качестве генетической модели нескольких неврологических заболеваний человека, включая нейродегенеративные расстройства Паркинсона, Хантингтона, спиноцеребеллярную атаксию и болезнь Альцгеймера. Несмотря на большую эволюционную дистанцию между насекомыми и млекопитающими, многие основные аспекты нейрогенетики дрозофилы оказались актуальными и для человека. Например, первые гены биологических часов были идентифицированы путем изучения мутантов дрозофилы , у которых были нарушены ежедневные циклы активности. [27]
См. также [ править ]
Цитаты [ править ]
- ^ Атта, HM (1999). «Хирургический папирус Эдвина Смита: старейший известный хирургический трактат». Американский хирург . 65 (12): 1190–1192. дои : 10.1177/000313489906501222 . ПМИД 10597074 . S2CID 30179363 .
- ^ Роуз, Ф (2009). «Церебральная локализация в древности». Журнал истории нейронаук . 18 (3): 239–247. дои : 10.1080/09647040802025052 . ПМИД 20183203 . S2CID 5195450 .
- ^ Jump up to: Перейти обратно: а б Рокка, Дж. (2003). Гален о мозге: анатомические знания и физиологические предположения во втором веке нашей эры . Том. 26. Брилл. стр. 1–313. ISBN 978-90-474-0143-8 . ПМИД 12848196 .
{{cite book}}:|journal=игнорируется ( помогите ) - ^ Поттер, П. (1976). «Герофил Халкидонский: Оценка его места в истории анатомии» . Бюллетень истории медицины . 50 (1): 45–60. ISSN 0007-5140 . JSTOR 44450313 . ПМИД 769875 .
- ^ Реверон, Р.Р. (2014). «Герофил и Эрасистрат, пионеры анатомического препарирования человека» . Везалий: Международный журнал истории медицины . 20 (1): 55–58. ПМИД 25181783 .
- ^ Аджита, Р. (2015). «Гален и его вклад в анатомию: обзор» . Журнал эволюции медицинских и стоматологических наук . 4 (26): 4509–4517. дои : 10.14260/jemds/2015/651 .
- ^ Джинн, СР; Лоруссо, Л. (2008). «Мозг, разум и тело: взаимодействие с искусством в Италии эпохи Возрождения». Журнал истории нейронаук . 17 (3): 295–313. дои : 10.1080/09647040701575900 . ПМИД 18629698 . S2CID 35600367 .
- ^ Маркатос, К.; Читас, Д.; Цакотос, Г.; Караману, М.; Пьягку, М.; Мазаракис, А.; Джонсон, Э. (2020). «Андреас Везалий Брюссельский (1514–1564): его вклад в область функциональной нейроанатомии и критика своих предшественников» . Акта Хирургика Бельгика . 120 (6): 437–441. дои : 10.1080/00015458.2020.1759887 . ПМИД 32345153 . S2CID 216647830 .
- ^ Сплавски, Б. (2019). «Андреас Везалий, предшественник нейрохирургии: как его прогрессивные научные достижения повлияли на его профессиональную жизнь и судьбу» . Мировая нейрохирургия . 129 : 202–209. дои : 10.1016/j.wneu.2019.06.008 . ПМИД 31201946 . S2CID 189897890 .
- ^ Неер, А (2009). «Кристофер Рен, Томас Уиллис и изображение мозга и нервов». Журнал медицинских гуманитарных наук . 30 (3): 191–200. дои : 10.1007/s10912-009-9085-5 . ПМИД 19633935 . S2CID 11121186 .
- ^ Грибные тела плодовой мухи. Архивировано 16 июля 2012 г., archive.today .
- ^ Джинджер, М.; Хаберл, М.; Конзельманн, К.-К.; Шварц, М.; Фрик, А. (2013). «Раскрытие секретов нейронных цепей с помощью технологии рекомбинантного вируса бешенства» . Передний. Нейронные цепи . 7 :2. дои : 10.3389/fncir.2013.00002 . ПМЦ 3553424 . ПМИД 23355811 .
- ^ Макговерн, А.Е.; Дэвис-Пойнтер, Н.; Ракоци, Дж; Фиппс, С; Симмонс, Д.Г.; Маццоне, С.Б. (2012). «Отслеживание антероградных нейрональных цепей с использованием генетически модифицированного вируса простого герпеса, экспрессирующего EGFP». J Неврологические методы . 209 (1): 158–67. doi : 10.1016/j.jneumeth.2012.05.035 . ПМИД 22687938 . S2CID 20370171 .
- ^ Кайперс Х.Г., Уголини Дж. (февраль 1990 г.). «Вирусы как транснейрональные индикаторы». Тенденции в нейронауках . 13 (2): 71–5. дои : 10.1016/0166-2236(90)90071-H . ПМИД 1690933 . S2CID 27938628 .
- ^ Ринаман Л., Шварц Г. (март 2004 г.). «Антероградное транснейрональное вирусное отслеживание центральных висцеросенсорных путей у крыс» . Журнал неврологии . 24 (11): 2782–6. doi : 10.1523/JNEUROSCI.5329-03.2004 . ПМК 6729508 . ПМИД 15028771 .
- ^ Норгрен Р.Б., Маклин Дж.Х., Бубель Х.К., Вандер А., Бернштейн Д.И., Леман М.Н. (март 1992 г.). «Антероградный транспорт ВПГ-1 и ВПГ-2 в зрительной системе». Бюллетень исследований мозга . 28 (3): 393–9. дои : 10.1016/0361-9230(92)90038-Y . ПМИД 1317240 . S2CID 4701001 .
- ^ Кард, JP (2001). «Нейроинвазивность вируса псевдобешенства: окно в функциональную организацию мозга». Достижения в области исследования вирусов . 56 : 39–71. дои : 10.1016/S0065-3527(01)56004-2 . ISBN 9780120398560 . ПМИД 11450308 .
- ^ Кард, JP (2011). «Подход к условному репортеру вируса псевдобешенства с двойной инфекцией для идентификации проекций на коллатерализованные нейроны в сложных нейронных цепях» . ПЛОС ОДИН . 6 (6): e21141. Бибкод : 2011PLoSO...621141C . дои : 10.1371/journal.pone.0021141 . ПМК 3116869 . ПМИД 21698154 .
- ^ Денк, В; Хорстманн, Х (2004). «Серийная блочная сканирующая электронная микроскопия для реконструкции трехмерной наноструктуры ткани» . ПЛОС Биология . 2 (11): е329. дои : 10.1371/journal.pbio.0020329 . ПМК 524270 . ПМИД 15514700 .
- ^ Вуллиманн, Марио Ф.; Рупп, Барбар; Райхерт, Генрих (1996). Нейроанатомия мозга рыбок данио: топологический атлас . Бирк[Ux9451]пользователь Верлаг. ISBN 3-7643-5120-9 . Архивировано из оригинала 15 июня 2013 г. Проверено 16 октября 2016 г.
- ^ «Атлас мозга дрозофилы» . Архивировано из оригинала 16 июля 2011 г. Проверено 24 марта 2011 г.
- ^ «WormBook: Интернет-обзор биологии C. elegans » . Архивировано из оригинала 11 октября 2011 г. Проверено 14 октября 2011 г.
- ^ Хоберт, Оливер (2005). Исследовательское сообщество C. elegans (ред.). «Спецификация нервной системы» . Червячная книга : 1–19. дои : 10.1895/wormbook.1.12.1 . ПМЦ 4781215 . ПМИД 18050401 . Архивировано из оригинала 17 июля 2011 г. Проверено 5 ноября 2011 г.
- ^ Уайт, Дж. Г.; Саутгейт, Восточная Европа ; Томсон, Дж. Н.; Бреннер, С. (1986). «Строение нервной системы нематоды Caenorhabditis elegans». Философские труды Королевского общества Б. 314 (1165): 1–340. Бибкод : 1986RSPTB.314....1W . дои : 10.1098/rstb.1986.0056 . ПМИД 22462104 .
- ^ Ходжкин Дж (2001). « Каенорхабдитис элегантный ». В Бреннер С., Миллер Дж. Х. (ред.). Энциклопедия генетики . Эльзевир. стр. 251–256. ISBN 978-0-12-227080-2 .
- ^ «Flybrain: онлайн-атлас и база данных нервной системы дрозофилы » . Архивировано из оригинала 16 мая 2016 г. Проверено 14 октября 2011 г.
- ^ Конопка, Р.Дж.; Бензер, С. (1971). «Часовые мутанты Drosophila melanogaster» . Учеб. Натл. акад. наук. США . 68 (9): 2112–6. Бибкод : 1971ПНАС...68.2112К . дои : 10.1073/pnas.68.9.2112 . ПМЦ 389363 . ПМИД 5002428 .
Источники [ править ]
- Родитель, Андре ; Карпентер, Малкольм Б. (1996). Нейроанатомия человека Карпентера (9-е изд.). Уильямс и Уилкинс. ISBN 978-0683067521 .
- Патестас, Мария А.; Гартнер, Лесли П. (2016). Учебник нейроанатомии (2-е изд.). Уайли Блэквелл. ISBN 978-1-118-67746-9 .
- Сплиттгербер, Райан (2019). Клиническая нейроанатомия Снелла (8-е изд.). Уолтерс Клювер. ISBN 978-1496346759 .
- Ваксман, Стивен (2020). Клиническая нейроанатомия (29-е изд.). Макгроу-Хилл Образование. ISBN 978-1260452358 .
Внешние ссылки [ править ]
- Нейроанатомия , ежегодный журнал клинической нейроанатомии.
- Атласы мозга мыши, крысы, примата и человека (Центр вычислительной биологии Калифорнийского университета в Лос-Анджелесе)
- Brainmaps.org: Нейроанатомически аннотированные атласы головного мозга высокого разрешения
- BrainInfo для нейроанатомии
- Система управления архитектурой мозга , несколько атласов анатомии мозга
- Атлас белого вещества , Атлас диффузионно-тензорной визуализации участков белого вещества головного мозга


| Базы данных органов управления : Национальные |
|---|