Членистоногие
| Членистоногие Временной диапазон: Самый ранний кембрий ( фортуниан ) – недавний. | |
|---|---|
 | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Subkingdom: | Эуметазоа |
| Клэйд : | ПараХоксозоа |
| Клэйд : | билатерия |
| Клэйд : | Нефрозоа |
| (без рейтинга): | протостомия |
| Супертип: | Экдисозоа |
| (без рейтинга): | панартроподы |
| (без рейтинга): | Тактопода |
| Тип: | Членистоногие Грейвенхорст , 1843 год. [1] [2] |
| Подтипы, неразмещенные роды и классы | |
| Разнообразие | |
| около 1 170 000 видов . | |
| Синонимы | |
Кондилипода Латрейль, 1802 г. | |
Членистоногие ( / ˈ ɑːr θ r ə p ɒ d / ARTH -rə-pod ) [20] беспозвоночные из типа Членистоногие . Они обладают экзоскелетом с кутикулой из хитина , часто минерализованной карбонатом кальция , телом с дифференцированными ( метамерными ) сегментами и парными членистыми придатками . Чтобы продолжать расти, они должны пройти этапы линьки — процесса, в ходе которого они сбрасывают свой экзоскелет , чтобы обнажить новый. Это чрезвычайно разнообразная группа, насчитывающая до 10 миллионов видов.
Гемолимфа является аналогом крови для большинства членистоногих. Членистоногие имеют открытую систему кровообращения с полостью тела, называемой гемоцелем, через которую гемолимфа циркулирует во внутренние органы . Как и их внешний вид, внутренние органы членистоногих обычно состоят из повторяющихся сегментов. Их нервная система имеет «лестничную форму»: парные вентральные нервные шнуры проходят через все сегменты и образуют парные ганглии в каждом сегменте . Их головы образуются в результате слияния различного числа сегментов, а мозг формируется в результате слияния ганглиев этих сегментов и окружает пищевод . Дыхательная подтипа и выделительная системы членистоногих различаются в зависимости как от окружающей среды, так и от , к которому они принадлежат.
используют комбинацию сложных глаз и с пигментными ямками глазков Членистоногие для зрения . У большинства видов глазки могут определять только направление, откуда падает свет, а сложные глаза являются основным источником информации, но у пауков основными глазами являются глазки, которые могут формировать изображения и, в некоторых случаях, поворачиваться в сторону. отслеживать добычу. Членистоногие также обладают широким спектром химических и механических сенсоров, в основном основанных на модификациях множества щетинок, известных как щетинки , которые выступают через их кутикулы. Точно так же разнообразны их размножение и развитие; все наземные виды используют внутреннее оплодотворение , но иногда это происходит путем непрямой передачи спермы через придаток или землю, а не путем прямой инъекции. Водные виды используют либо внутреннее, либо внешнее оплодотворение . Почти все членистоногие откладывают яйца, причем многие виды рожают живых детенышей после того, как яйца вылупились внутри матери; но некоторые из них действительно живородящие , например тля . Детеныши членистоногих варьируются от миниатюрных взрослых особей до личинок и гусеницы , у которых отсутствуют суставчатые конечности и в конечном итоге претерпевают полную метаморфозу , образуя взрослую форму. Уровень материнской заботы о вылупившихся птенцах варьируется от полного отсутствия до продолжительной заботы, оказываемой социальными насекомыми .
Эволюционное происхождение членистоногих восходит к кембрийскому периоду. Группа обычно считается монофилетической , и многие анализы подтверждают размещение членистоногих с циклонейралиями (или их составляющими кладами) в супертипе Ecdysozoa . Однако в целом базальные взаимоотношения животных еще не вполне решены. Точно так же до сих пор активно обсуждаются взаимоотношения между различными группами членистоногих. Сегодня членистоногие вносят вклад в обеспечение человека продовольствием как непосредственно в качестве пищи, так и, что более важно, косвенно в качестве опылителей сельскохозяйственных культур. Известно, что некоторые виды передают тяжелые заболевания людям, домашнему скоту и сельскохозяйственным культурам .
Этимология [ править ]
Слово членистоногие происходит от греческого ἄρθρον arthron « сустав » и πούς pous ( род. ποδός podos ) « ступня » или « нога » , которые вместе означают «суставная нога», [21] со словом «членистоногие», первоначально использованным в анатомических описаниях Бартелеми Шарля Жозефа Дюмортье, опубликованных в 1832 году. [1] Название «Членистоногие», по-видимому, впервые было использовано в 1843 году немецким зоологом Иоганном Людвигом Кристианом Гравенхорстом (1777–1857). [22] [1] Происхождение имени было предметом серьезной путаницы: вместо этого часто ошибочно отдавали должное Пьеру Андре Латрею или Карлу Теодору Эрнсту фон Зибольду , а также многим другим. [1]
Наземных членистоногих часто называют клопами. [Примечание 1] Этот термин также иногда распространяется на разговорные названия пресноводных или морских ракообразных (например, клоп Балмейн , клоп Мортон-Бей , грязевой клоп ) и используется врачами и бактериологами для обозначения болезнетворных микробов (например, супербактерий ), [25] но энтомологи сохраняют этот термин для узкой категории « настоящих клопов », насекомых отряда Hemiptera . [25]
Описание [ править ]
Членистоногие — беспозвоночные с сегментированным телом и членистыми конечностями. [26] Экзоскелет ацетилглюкозамина или кутикула состоит из хитина , полимера N- . [27] Кутикула многих ракообразных, жуков-клещей , клад Penetini и Archaeoglenini внутри подсемейства жуков Phrenapatinae , [28] и многоножек (кроме щетинистых многоножек ) также биоминерализован карбонатом кальция . Кальцификация эндостернита, внутренней структуры, используемой для прикрепления мышц, также встречается у некоторых опилионов . [29] а кутикула куколки мухи Bactrocera dorsalis содержит фосфат кальция. [30]
Разнообразие [ править ]

Членистоногие являются крупнейшим типом животных , число видов членистоногих варьируется от 1 170 000 до 5–10 миллионов и составляет более 80 процентов всех известных ныне видов животных. [31] [32] Одна подгруппа членистоногих , насекомые , включает больше описанных видов , чем любой другой таксономический класс . [33] Общее количество видов определить пока сложно. Это связано с допущениями моделирования переписи, проецируемыми на другие регионы с целью масштабирования подсчетов в конкретных местах применительно ко всему миру. насчитывается 500 000 видов животных и растений Исследование, проведенное в 1992 году, показало, что только в Коста-Рике , из которых 365 000 являются членистоногими. [33]
Они являются важными членами морских, пресноводных, наземных и воздушных экосистем и одной из двух основных групп животных, приспособившихся к жизни в засушливой среде; другой — амниоты , живыми членами которых являются рептилии, птицы и млекопитающие. [34] И самые маленькие, и самые крупные членистоногие относятся к ракообразным . Самые маленькие принадлежат к классу Tantulocarida , некоторые из них имеют длину менее 100 микрометров (0,0039 дюйма). [35] Самыми крупными являются виды класса Malacostraca , длина ног японского краба-паука может достигать 4 метров (13 футов). [36] и американский омар , достигающий веса более 20 кг (44 фунта).
Сегментация [ править ]

Эмбрионы всех членистоногих сегментированы, построены из ряда повторяющихся модулей. Последний общий предок современных членистоногих, вероятно, состоял из ряда недифференцированных сегментов, каждый из которых имел пару придатков, выполнявших функцию конечностей. Однако все известные живые и ископаемые членистоногие сгруппировали сегменты в тагматы , в которых сегменты и их конечности различным образом специализированы. [34]
трехчастный вид тел многих насекомых и двухчастный облик пауков . Результатом этой группировки является [38] нет Внешних признаков сегментации у клещей . [34] У членистоногих также есть два элемента тела, которые не являются частью этого последовательно повторяющегося набора сегментов: глазной сомит спереди, где возникли рот и глаза, [34] [39] и тельсон сзади, за анусом .
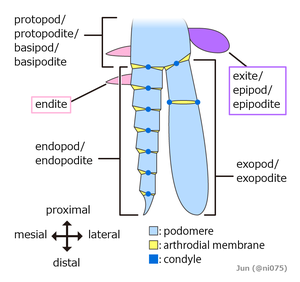
Первоначально казалось, что каждый сегмент, несущий придатки, имел две отдельные пары придатков: верхний несегментированный выход и нижний сегментированный эндопод. Позже они слились в одну пару двуветвистых придатков, объединенных базальным сегментом (протоподом или базиподом), при этом верхняя ветвь действовала как жабры , а нижняя ветвь использовалась для передвижения. [40] [41] [37] Придатки большинства ракообразных и некоторых вымерших таксонов, таких как трилобиты, имеют еще одну сегментированную ветвь, известную как экзоподы , но вопрос о том, имеют ли эти структуры единое происхождение, остается спорным. [42] [43] [37] У некоторых сегментов всех известных членистоногих придатки видоизменились, например, образовав жабры, ротовой аппарат, усики для сбора информации, [38] или когти для хватания; [44] Членистоногие «подобны швейцарским армейским ножам , каждый из которых оснащен уникальным набором специализированных инструментов». [34] У многих членистоногих придатки в некоторых участках тела исчезли; особенно часто брюшные придатки исчезают или сильно видоизменяются. [34]

Наиболее заметна специализация сегментов в голове. Четыре основные группы членистоногих — Chelicerata ( морские пауки , мечехвосты и паукообразные ), Myriapoda ( симфиланы , павроподы , многоножки и многоножки ), Pancrustacea ( олигостраканы , веслоногие раки , малакостраканы , жабероподы , шестиногие и т. д.) и вымершие трилобиты — имеют головы, состоящие из различных комбинаций сегментов, с придатками, которые отсутствуют или специализируются по-разному. [34] Несмотря на то, что многоножки и шестиногие имеют схожие комбинации голов, гексаподы глубоко гнездятся внутри ракообразных, а многоножки - нет, поэтому считается, что эти черты развивались отдельно. Кроме того, некоторые вымершие членистоногие, такие как Маррелла , не принадлежат ни к одной из этих групп, поскольку их головы образованы собственными особыми комбинациями сегментов и специализированных придатков. [46]
Выяснить эволюционные этапы, на которых могли возникнуть все эти различные комбинации, настолько сложно, что это уже давно известно как « проблема головы членистоногих ». [47] В 1960 году Р.Э. Снодграсс даже надеялся, что проблема не будет решена, поскольку находить решения было для него забавным занятием. [Примечание 2]
Экзоскелет [ править ]
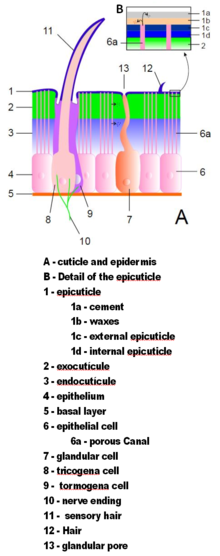
Экзоскелеты членистоногих состоят из кутикулы — неклеточного материала, выделяемого эпидермисом . [34] Их кутикулы различаются по деталям своей структуры, но обычно состоят из трех основных слоев: эпикутикулы , тонкого внешнего воскового слоя, который защищает от влаги другие слои и дает им некоторую защиту; экзокутикула и химически , состоящая из хитина затвердевших белков ; и эндокутикула , состоящая из хитина и незатвердевших белков. Экзокутикула и эндокутикула вместе известны как прокутикула . [49] Каждый сегмент тела и часть конечностей покрыты затвердевшей кутикулой. Соединения между сегментами тела и между отделами конечностей покрыты гибкой кутикулой. [34]
Экзоскелеты большинства водных ракообразных биоминерализованы , карбонатом кальция извлеченным из воды. Некоторые наземные ракообразные разработали способы хранения минерала, поскольку на суше они не могут рассчитывать на постоянный запас растворенного карбоната кальция. [50] Биоминерализация обычно затрагивает экзокутикулу и внешнюю часть эндокутикулы. [49] Две недавние гипотезы об эволюции биоминерализации у членистоногих и других групп животных предполагают, что она обеспечивает более прочную защитную броню. [51] и что это позволяет животным становиться больше и сильнее за счет более жесткого скелета; [52] и в любом случае экзоскелет из минерально-органического композита дешевле построить, чем полностью органический экзоскелет сопоставимой прочности. [52] [53]
Кутикула может иметь щетинки (щетинки), растущие из особых клеток эпидермиса. Щетинки столь же разнообразны по форме и функциям, как и придатки. Например, их часто используют в качестве датчиков для обнаружения потоков воздуха или воды или контакта с объектами; водные членистоногие используют перьевые щетинки для увеличения площади поверхности плавательных придатков и фильтрации частиц пищи из воды; водные насекомые, дышащие воздухом, используют толстые войлочные покровы щетинок для улавливания воздуха, продлевая время, которое они могут провести под водой; тяжелые, жесткие щетинки служат защитными шипами. [34]
Хотя все членистоногие используют мышцы, прикрепленные к внутренней части экзоскелета, для сгибания конечностей, некоторые все еще используют гидравлическое давление для их разгибания - система, унаследованная от их предшественников-членистоногих; [54] например, все пауки вытягивают ноги гидравлически и могут создавать давление, в восемь раз превышающее уровень покоя. [55]
Линька [ править ]

Экзоскелет не может растягиваться и, таким образом, ограничивает рост. Таким образом, членистоногие заменяют свои экзоскелеты, подвергаясь шелушению (линьке) или сбрасывая старый экзоскелет, экзувии , после выращивания нового, еще не затвердевшего. Циклы линьки происходят почти непрерывно, пока членистоногое не достигнет полного размера. Стадии развития между каждой линькой (шелушением) до достижения половой зрелости называются возрастом . Различия между возрастами часто можно увидеть в изменении пропорций тела, цвета, рисунка, изменении количества сегментов тела или ширины головы. После линьки, то есть потери экзоскелета, молодые членистоногие продолжают свой жизненный цикл до тех пор, пока они либо не окукливаются, либо не линяют снова. [56]
В начальной фазе линьки животное перестает питаться, и его эпидермис выделяет линяющую жидкость — смесь ферментов , которая переваривает эндокутикулу и таким образом отделяет старую кутикулу. Эта фаза начинается, когда эпидермис выделяет новую эпикутикулу для защиты от ферментов, и эпидермис секретирует новую экзокутикулу, в то время как старая кутикула отслаивается. Когда эта стадия завершена, животное заставляет свое тело разбухать, поглощая большое количество воды или воздуха, в результате чего старая кутикула расщепляется по заранее определенным слабым местам, где старая экзокутикула была самой тонкой. Обычно животному требуется несколько минут, чтобы выбраться из старой кутикулы. В этот момент новая эндокутикула морщинистая и настолько мягкая, что животное не может поддерживать себя и ей очень трудно двигаться, а новая эндокутикула еще не сформировалась. Животное продолжает накачиваться, чтобы максимально растянуть новую кутикулу, затем затвердевает новая экзокутикула и удаляет лишний воздух или воду. К концу этой фазы формируется новая эндокутикула. Многие членистоногие затем поедают выброшенную кутикулу, чтобы восстановить ее материалы. [56]
Поскольку членистоногие не защищены и почти обездвижены до тех пор, пока новая кутикула не затвердеет, они подвергаются опасности как оказаться в ловушке старой кутикулы, так и подвергнуться нападению хищников . Линька может быть причиной от 80 до 90% всех смертей членистоногих. [56]
Внутренние органы [ править ]

Тела членистоногих также сегментированы внутри, а нервная, мышечная, кровеносная и выделительная системы имеют повторяющиеся компоненты. [34] Членистоногие происходят от животных, у которых есть целом — выстланная мембраной полость между кишечником и стенкой тела, в которой расположены внутренние органы. Сильные сегментированные конечности членистоногих устраняют необходимость в одной из основных наследственных функций целома, а именно в гидростатическом скелете , который мышцы сжимают, чтобы изменить форму животного и, таким образом, дать ему возможность двигаться. Следовательно, целом членистоногих сокращается до небольших участков вокруг репродуктивной и выделительной систем. Его место в значительной степени занимает гемоцель — полость, которая проходит по большей части тела и через которую течет кровь . [57]
Дыхание и кровообращение [ править ]

Членистоногие имеют открытую систему кровообращения . Большинство из них имеют несколько коротких открытых артерий . У хелицератов и ракообразных кровь переносит кислород к тканям, а у шестиногих используется отдельная система трахей . Многие ракообразные, а также некоторые хелицераты и трахеаты используют дыхательные пигменты для облегчения транспорта кислорода. Наиболее распространенным дыхательным пигментом у членистоногих является меди на основе гемоцианин ; это используется многими ракообразными и некоторыми многоножками . Некоторые ракообразные и насекомые используют гемоглобин на основе железа — дыхательный пигмент, используемый позвоночными . Как и у других беспозвоночных, дыхательные пигменты тех членистоногих, у которых они есть, обычно растворены в крови и редко заключены в тельца , как у позвоночных. [57]
Сердце представляет собой мышечную трубку, которая проходит под спиной и занимает большую часть длины гемоцеля. Он сжимается волнами, идущими сзади вперед, выталкивая кровь вперед. Участки, не сдавливаемые сердечной мышцей, расширяются либо эластичными связками , либо мелкими мышцами , в том и другом случае соединяющими сердце со стенкой тела. Вдоль сердца проходит ряд парных отверстий, обратных клапанов, которые позволяют крови проникать в сердце, но не позволяют ей покинуть ее, прежде чем она достигнет передней части сердца. [57]
Членистоногие имеют большое разнообразие дыхательных систем. У мелких видов их часто нет, поскольку их высокое соотношение площади поверхности к объему обеспечивает простую диффузию через поверхность тела для снабжения достаточным количеством кислорода. У ракообразных обычно есть жабры, представляющие собой видоизмененные придатки. У многих паукообразных имеются книжные легкие . [58] Трахеи, системы ветвящихся туннелей, идущих от отверстий в стенках тела, доставляют кислород непосредственно к отдельным клеткам у многих насекомых, многоножек и паукообразных . [59]
Нервная система [ править ]

У современных членистоногих есть парные основные нервные тяжи, проходящие вдоль их тела ниже кишечника, и в каждом сегменте тяжи образуют пару ганглиев , от которых сенсорные и двигательные нервы идут к другим частям сегмента. Хотя пары ганглиев в каждом сегменте часто кажутся физически слитыми, они соединены комиссурами (относительно большими пучками нервов), которые придают нервной системе членистоногих характерный «лестничный» вид. Мозг находится в голове, окружая и преимущественно над пищеводом. Он состоит из сросшихся ганглиев акрона и одного или двух передних сегментов, образующих голову - всего три пары ганглиев у большинства членистоногих и только две у хелицератов, не имеющих усиков или соединенного с ними ганглия. . Ганглии других сегментов головы часто располагаются близко к мозгу и функционируют как его части. У насекомых эти другие головные ганглии объединяются в пару подпищеводных ганглиев , расположенных под и позади пищевода. Пауки идут еще дальше в этом процессе, поскольку все сегментарные ганглии включены в подпищеводные ганглии, занимающие большую часть пространства головогруди (передний «сверхсегмент»). [60]
Выделительная система [ править ]
Существует два разных типа выделительных систем членистоногих. У водных членистоногих конечным продуктом биохимических реакций метаболизма азота является аммиак , который настолько токсичен, что его необходимо максимально разбавлять водой. Затем аммиак удаляется через любую проницаемую мембрану, главным образом через жабры. [58] Все ракообразные используют эту систему, и высокое потребление воды может быть причиной относительной неудачи ракообразных в качестве наземных животных. [61] Различные группы наземных членистоногих независимо друг от друга выработали разную систему: конечным продуктом азотистого обмена является мочевая кислота , которая может выделяться в виде сухого вещества; Система мальпигиевых канальцев фильтрует мочевую кислоту и другие азотистые отходы из крови в гемоцеле и сбрасывает эти материалы в заднюю кишку, откуда они выводятся в виде фекалий . [61] У большинства водных членистоногих и некоторых наземных также есть органы, называемые нефридиями («маленькие почки »), которые извлекают другие отходы для выведения их в виде мочи . [61]
Чувства [ править ]

Жесткая кутикула членистоногих блокировала бы информацию о внешнем мире, за исключением того, что через них пронизано множество датчиков или соединений датчиков с нервной системой. Фактически, членистоногие превратили свои кутикулы в сложные наборы сенсоров. Различные сенсоры прикосновения, в основном щетинки , реагируют на разные уровни силы: от сильного прикосновения до очень слабых потоков воздуха. Химические сенсоры обеспечивают эквиваленты вкуса и запаха , часто с помощью щетинок. Датчики давления часто имеют форму мембран, которые функционируют как барабанные перепонки , но связаны непосредственно с нервами, а не со слуховыми косточками . Усики влажность большинства гексапод включают в себя пакеты датчиков, которые контролируют , влажность и температуру. [62]
Большинству членистоногих не хватает датчиков равновесия и ускорения , и они полагаются на свои глаза, чтобы определить, в какую сторону идти вверх. начинают самовосстанавливаться Тараканы , когда датчики давления на нижней стороне лап сообщают об отсутствии давления. Однако у многих ракообразных -малакостраканов есть статоцисты , которые предоставляют ту же информацию, что и датчики баланса и движения внутреннего уха позвоночных . [62]
Проприорецепторы членистоногих — сенсоры , которые сообщают о силе, оказываемой мышцами, а также степени сгибания тела и суставов — хорошо изучены. Однако мало что известно о том, какие еще внутренние сенсоры могут быть у членистоногих. [62]
Оптический [ править ]


Большинство членистоногих обладают сложной зрительной системой, которая включает в себя один или несколько сложных глаз с пигментными чашечками и глазков («маленькие глазки»). В большинстве случаев глазки способны определять направление света только по тени, отбрасываемой стенками чашки. Однако основные глаза пауков представляют собой пигментные чашечки глазков, способные образовывать изображения. [62] а пауки -прыгуны могут вращаться, чтобы выслеживать добычу. [63]
Сложные глаза состоят из пятнадцати — нескольких тысяч независимых омматидий — столбиков, обычно шестиугольных в сечении . Каждый омматидий представляет собой самостоятельный сенсор со своими светочувствительными клетками и часто со своей хрусталиком и роговицей . [62] Сложные глаза имеют широкое поле зрения и могут обнаруживать быстрое движение и, в некоторых случаях, поляризацию света . [64] С другой стороны, относительно большой размер омматидий делает изображения довольно грубыми, а фасеточные глаза близоруки, чем у птиц и млекопитающих, хотя это не является серьезным недостатком, поскольку объекты и события в пределах 20 см (8 дюймов) являются наиболее важными для большинства членистоногих. [62] Некоторые членистоногие обладают цветовым зрением, а у некоторых насекомых оно детально изучено; например, омматидии пчел содержат рецепторы как зеленого, так и ультрафиолетового света . [62]
Обоняние [ править ]
и развитие Размножение


Некоторые членистоногие, например ракообразные , гермафродиты , то есть каждый может иметь органы обоих полов . Однако особи большинства видов всю жизнь остаются представителями одного пола. [65] Некоторые виды насекомых и ракообразных могут размножаться партеногенезом , особенно если условия благоприятствуют «демографическому взрыву». Однако большинство членистоногих полагаются на половое размножение , а партеногенетические виды часто возвращаются к половому размножению, когда условия становятся менее благоприятными. [66] Способность к мейозу широко распространена среди членистоногих, включая как тех, которые размножаются половым путем, так и тех, которые размножаются партеногенетически . [67] Хотя мейоз является основной характеристикой членистоногих, понимание его фундаментального адаптивного преимущества долгое время считалось нерешенной проблемой. [68] похоже, это осталось нерешенным.
Водные членистоногие могут размножаться путем внешнего оплодотворения, как, например, мечехвосты . [69] или путем внутреннего оплодотворения , при котором яйцеклетки остаются в организме самки, а сперму необходимо каким-то образом внедрить. Все известные наземные членистоногие используют внутреннее оплодотворение. Опилионы (сборщики урожая), многоножки и некоторые ракообразные используют модифицированные придатки, такие как гоноподы или пенисы, для передачи спермы непосредственно самке. Однако большинство самцов наземных членистоногих производят сперматофоры — водонепроницаемые пакеты спермы , которые самки принимают в свои тела. Некоторые из таких видов полагаются на самок, чтобы найти сперматофоры, которые уже были отложены на земле, но в большинстве случаев самцы откладывают сперматофоры только тогда, когда сложные ритуалы ухаживания кажутся успешными. [65]

Большинство членистоногих откладывают яйца. [65] но скорпионы яйцеживородящие : они производят живых детенышей после того, как яйца вылупились внутри матери, и отличаются длительной материнской заботой. [70] Новорожденные членистоногие имеют разнообразные формы, и только насекомые охватывают диапазон крайностей. Некоторые вылупляются как миниатюрные взрослые особи (прямое развитие), а в некоторых случаях, например, у чешуйницы , птенцы не питаются и могут быть беспомощными до первой линьки. Многие насекомые вылупляются в виде личинок или гусениц , у которых нет сегментированных конечностей или затвердевшей кутикулы, и метаморфизируются во взрослые формы, вступая в неактивную фазу, в которой ткани личинки разрушаются и повторно используются для построения взрослого тела. [71] Личинки стрекоз имеют типичную кутикулу и членистые конечности членистоногих, но являются нелетающими вододышащими с выдвижными челюстями. [72] Ракообразные обычно вылупляются в виде крошечных личинок науплиусов , имеющих всего три сегмента и пару придатков. [65]
Эволюционная история [ править ]
Последний общий предок [ править ]
На основании распределения общих плезиоморфных особенностей среди современных и ископаемых таксонов можно сделать вывод, что последний общий предок всех членистоногих представлял собой модульный организм, каждый модуль которого был покрыт собственным склеритом (броневой пластиной) и имел пару двуветвистых конечностей . [73] Однако вопрос о том, была ли предковая конечность одноветвистой или двуветвистой, еще далеко не решен.У этого ур-членистоногого был брюшной рот, предротовые усики и спинные глаза в передней части тела. Предполагалось, что он был недискриминационным питателем отложений , перерабатывающим любые осадки, попадавшие на его пути в пищу. [73] но находки окаменелостей намекают на то, что последний общий предок членистоногих и приапулид имел один и тот же специализированный ротовой аппарат; круглый рот с кольцами зубов, используемый для захвата добычи животных. [74]
Ископаемая запись [ править ]

Было высказано предположение, что эдиакарские животные Парванкорина и Сприггина , жившие около 555 миллионов лет назад , были членистоногими. [75] [76] [77] но более поздние исследования показывают, что их сходство с происхождением членистоногих недостоверно. [78] Маленькие членистоногие с двустворчатыми раковинами были обнаружены в ископаемых пластах раннего кембрия, датируемых 541–539 миллионами лет назад в Китае и Австралии. [79] [80] [81] [82] Самым ранним кембрийским окаменелостям трилобитов около 520 миллионов лет, но этот класс уже был довольно разнообразным и распространился по всему миру, что позволяет предположить, что они существовали уже довольно давно. [83] В сланцах Маотяньшань , возраст которых составляет 518 миллионов лет назад, членистоногие, такие как Kylinxia и Erratus, были обнаружены которые, по-видимому, представляют собой переходные окаменелости между стволовыми (например, Radiodonta, такими как Anomalocaris ) и настоящими членистоногими. [84] [6] [41] Повторное исследование в 1970-х годах окаменелостей сланцев Берджесс возрастом около 505 миллионов лет назад выявило множество членистоногих, некоторых из которых нельзя было отнести ни к одной из известных групп, и тем самым усилило споры о кембрийском взрыве . [85] [86] [87] Окаменелость Марреллы из сланцев Бёрджесс предоставила самое раннее явное свидетельство линьки . [88]


Самая ранняя окаменелость личинок панкрустообразных датируется примерно 514 миллионами лет назад в кембрийском периоде , за ней следуют уникальные таксоны, такие как Yicaris и Wujicaris . [89] Предполагаемое с панкрустообразными и ракообразными родство некоторых кембрийских членистоногих Phosphatocopina , Bradoriida и Hymenocarine , такие как ваптииды) (например, таксоны [90] [91] [92] были оспорены последующими исследованиями, поскольку они могли разветвляться раньше коронковой группы нижней челюсти . [89] В кроновой группе панракообразных только Malacostraca , Branchiopoda и Pentastomida имеют кембрийские окаменелости. [89] Окаменелости ракообразных распространены начиная с ордовикского периода. [93] Они остались почти полностью водными, возможно, потому, что у них так и не развилась выделительная система , сохраняющая воду. [61]
Членистоногие представляют собой самые ранние идентифицируемые окаменелости наземных животных, датированные примерно 419 миллионами лет назад в позднем силуре . [58] а земные следы, датированные примерно 450 миллионами лет назад, по-видимому, были оставлены членистоногими. [94] Членистоногие обладали качествами, которые легко использовать для жизни на суше; их существующие сочлененные экзоскелеты обеспечивали защиту от высыхания, поддержку от гравитации и средство передвижения, не зависящее от воды. [95] Примерно в то же время водные скорпионоподобные эвриптериды стали самыми крупными когда-либо членистоногими, длина некоторых из них достигала 2,5 м (8 футов 2 дюйма). [96]
Самый старый известный паукообразный — тригонотарбид Palaeotarbus jerami , живший примерно 420 миллионов лет назад в силурийский период. [97] [Примечание 3] Attercopus fimbriunguis , появившийся 386 миллионов лет назад в девонский период, несет на себе самые ранние известные патрубки для производства шелка, но отсутствие у него фильер означает, что он не был одним из настоящих пауков . [99] которые впервые появились в позднем карбоне более 299 миллионов лет назад . [100] Юрский меловой и периоды дают большое количество ископаемых пауков, в том числе представителей многих современных семейств. [101] Самый старый известный скорпион – Dolichophonus , датируемый 436 миллионами лет назад . [102] Раньше считалось, что многие силурийские и девонские скорпионы дышат жабрами , отсюда и возникла идея, что скорпионы были примитивно водными и развили легкие-книжки, дышащие воздухом. позже [103] Однако последующие исследования показали, что большинству из них не хватает надежных доказательств водного образа жизни. [104] в то время как исключительные водные таксоны (например, Waeringoscorpio ), скорее всего, произошли от предков наземных скорпионов. [105]
Самая старая запись окаменелостей гексапод неясна, поскольку большинство кандидатов плохо сохранились, а их родство с гексаподами оспаривалось. Ярким примером является девонский Rhyniognatha hirsti , датируемый 396–407 миллионами лет назад . Считается, что его челюсти относятся к типу, встречающемуся только у крылатых насекомых , что позволяет предположить, что самые ранние насекомые появились в силурийский период. [106] Однако более позднее исследование показало, что Rhyniognatha, скорее всего, представляет собой многоножку, а даже не шестиножку. [107] Однозначно старейшими известными гексаподами и насекомыми являются коллембола Rhyniella , появившаяся примерно 410 миллионов лет назад в девонском периоде, и палеодиктиоптеран Delitzschala bitterfeldensis , появившаяся примерно 325 миллионов лет назад в каменноугольном периоде соответственно. [107] Lagerstätten Мазон -Крик из позднего карбона, около 300 миллионов лет назад , включает около 200 видов, некоторые из которых гигантские по современным меркам, и указывает на то, что насекомые заняли свои основные современные экологические ниши в качестве травоядных , детритофагов и насекомоядных . Социальные термиты и муравьи впервые появляются в раннем мелу , а развитые социальные пчелы были обнаружены в позднемеловых породах, но их численность не становилась массовой до среднего кайнозоя . [108]
отношения с другими Эволюционные типами животных

С 1952 по 1977 год зоолог Сидни Мэнтон и другие утверждали, что членистоногие полифилетичны , другими словами, что у них нет общего предка, который сам был членистоногим. Вместо этого они предположили, что три отдельные группы «членистоногих» развились отдельно от общих червеобразных предков: хелицераты , включая пауков и скорпионов ; ракообразные; и унирамия , состоящая из онихофоров , многоножек и шестиногих . Эти аргументы обычно обходили трилобитов стороной , так как эволюционные связи этого класса были неясны. Сторонники полифилии утверждали следующее: сходство между этими группами является результатом конвергентной эволюции , как естественное следствие наличия жестких, сегментированных экзоскелетов ; что эти три группы используют разные химические средства отверждения кутикулы; что существуют значительные различия в строении их сложных глаз; что трудно понять, как такие разные конфигурации сегментов и придатков головы могли произойти от одного и того же предка; и что ракообразные имеют двуветвистые конечности с отдельными жаберными и ножными ветвями, тогда как у двух других групп конечности одноветвистые , у которых единственная ветвь служит ногой. [110]
| ||||||||||||||||||||||||||||
Дальнейший анализ и открытия 1990-х годов изменили эту точку зрения и привели к признанию того, что членистоногие монофилетичны , иными словами, предполагается, что они имеют общего предка, который сам был членистоногим. [111] [112] Например, Грэмом Баддом анализ Kerygmachela в 1993 году и Opabinia в 1996 году убедил его, что эти животные похожи на онихофоров и различных раннекембрийских « лобоподов », и он представил «эволюционное генеалогическое древо», которое показало их как «тетушек». " и "двоюродные братья" всех членистоногих. [109] [113] Эти изменения сделали неясным содержание термина «членистоногие», и Клаус Нильсен предложил называть более широкую группу « Panarthropoda » («все членистоногие»), а животных с суставчатыми конечностями и затвердевшей кутикулой следует называть «Euarthropoda» (Euarthropoda). «настоящие членистоногие»). [114]
Противоположная точка зрения была высказана в 2003 году, когда Ян Бергстрем и Хоу Сянь-гуан утверждали, что, если членистоногие были «сестринской группой» любого из аномалокарид, они, должно быть, утратили, а затем вновь развили черты, которые были хорошо развиты у аномалокарид. аномалокариды. Самые ранние известные членистоногие питались грязью, чтобы извлечь из нее частицы пищи, и обладали разным количеством сегментов с неспециализированными придатками, которые выполняли функции как жабр, так и ног. По стандартам того времени аномалокариды были огромными и сложными хищниками со специализированными ртами и хватательными придатками, фиксированным количеством сегментов, некоторые из которых были специализированными, хвостовыми плавниками и жабрами, сильно отличавшимися от таковых у членистоногих. В 2006 году они предположили, что членистоногие более тесно связаны с лобоподами и тихоходками, чем с аномалокаридами. [115] В 2014 году было обнаружено, что тихоходки более тесно связаны с членистоногими, чем с бархатными червями. [116]
| Протостомы |
| |||||||||||||||||||||
Отношения Ecdysozoa друг к другу, к кольчатым червям и т . д ., [117] [ не удалось пройти проверку ] в том числе эутикарциноиды [118] | ||||||||||||||||||||||




Выше «генеалогического древа» Annelida традиционно считались ближайшими родственниками Panarthropoda, поскольку обе группы имели сегментированные тела, а комбинация этих групп получила название Articulata . Высказывались конкурирующие предположения о том, что членистоногие тесно связаны с другими группами, такими как нематоды , приапулиды и тихоходки , но эти мнения оставались меньшинством, поскольку было трудно подробно определить отношения между этими группами.
В 1990-х годах молекулярно-филогенетический анализ последовательностей ДНК позволил создать последовательную схему, показывающую членистоногих как членов супертипа, названного Ecdysozoa («животные, которые линяют»), который содержал нематод, приапулид и тихоходок, но исключал кольчатых червей. Это было подтверждено исследованиями анатомии и развития этих животных, которые показали, что многие из особенностей, подтверждающих гипотезу Articulata, демонстрируют существенные различия между кольчатыми червями и самыми ранними панартроподами в своих деталях, а некоторые вообще почти не присутствовали у членистоногих. Эта гипотеза объединяет кольчатых червей с моллюсками и брахиоподами в другой супертип, Lophotrochozoa .
Если гипотеза Ecdysozoa верна, то сегментация членистоногих и кольчатых червей либо развилась конвергентно , либо была унаследована от гораздо более древнего предка и впоследствии потеряна в нескольких других линиях, таких как представители Ecdysozoa, не относящиеся к членистоногим. [119] [117]
Эволюция ископаемых членистоногих [ править ]
| Ископаемая филогения членистоногих [120] | |||
| |||
| Сводная кладограмма взаимоотношений вымерших групп членистоногих. Подробнее см. Deuteropoda . |
Помимо четырех основных современных групп ( ракообразные , хелицераты , многоножки и шестиногие ), ряд ископаемых форм, в основном из раннего кембрийского периода, трудно определить таксономически либо из-за отсутствия очевидного родства с какой-либо из основных групп, либо из-за отсутствия очевидного родства с какой-либо из основных групп. явная близость к некоторым из них. Маррелла была первой, которую признали существенно отличающейся от известных групп. [46]
Современные интерпретации базальной вымершей стеблевой группы членистоногих выделяют следующие группы, от самых базальных до самых кроновых: [121] [120]
- « Гигантские » или «сибириидные лобоподии» , такие как Jianshanopodia , Siberion и Megadictyon , являются самой базовой ступенью в общей группе Arthropoda.
- « Жаберные лобоподии» , такие как Kerygmachela , Pambdelurion и Opabinia , являются вторым наиболее базальным сортом.
- Radiodonta , традиционно известные как аномалокаридиды , занимают третье место и считаются монофилетическими .
- Возможный комплекс «верхней стволовой группы» более неопределенного положения. [120] но содержится внутри Deuteropoda : [121] Fuxianhuiida включая , Megacheira и многочисленные «двустворчатые формы», Isoxyida и Hymenocarina .
Deuteropoda - это недавно созданная клада , объединяющая членистоногих кроновой группы (живых) с этими возможными ископаемыми таксонами «верхней стеблевой группы». [121] Клада определяется важными изменениями в структуре области головы, такими как появление дифференцированной пары дейтоцеребральных придатков, которая исключает больше базальных таксонов, таких как радиодонты и «жаберные лобоподии». [121]
До сих пор остаются споры о положении различных вымерших групп членистоногих. Некоторые исследования обнаруживают, что Megacheira тесно связаны с хелицератами, в то время как другие обнаруживают, что они находятся за пределами группы, содержащей Chelicerate и Mandibulata, как эуартроподы стеблевой группы. [122] Размещение парнокопытных ( содержащих вымерших трилобитов и подобные формы) также является частым предметом споров. [123] Основные гипотезы относят их к кладе Arachnomorpha с хелицератами. Однако одна из новейших гипотез заключается в том, что хелицеры произошли от той же пары придатков, которые превратились в усики у предков Mandibulata , что поместило бы трилобитов, имевших усики, ближе к Mandibulata, чем к Chelicerata, в кладу Antennulata . [122] [124] Фуксианхуиды , которых обычно относят к членистоногим стеблевой группы, в некоторых недавних исследованиях были предложены как мандибулаты. [122] Hymenocarina , группа двустворчатых членистоногих, которые ранее считались членами стеблевой группы этой группы, как было продемонстрировано, являются нижнечелюстными на основании наличия нижних челюстей. [120]
- Радиодонты, опабинииды, жаберные лобоподии и более традиционные лобоподии являются примерами линий членистоногих базальной стеблевой группы из кембрия.







- Маррелломорфы, мегахерии, функсианхуиды и фосфатокопины - некоторые примеры кембрийских членистоногих, классификация которых остается сложной.








- Другие примеры ныне вымерших групп членистоногих включают:








современных классификация членистоногих Эволюция и
Тип Arthropoda обычно подразделяется на четыре подтипа , один из которых вымер : [125]
- Парнокопытные — это вымершая группа ранее многочисленных морских животных , исчезнувших во время пермско-триасового вымирания сократились до одного порядка , хотя до этого смертельного удара они находились в упадке, а в позднедевонском вымирании . Они содержат такие группы, как трилобиты .
- Хелицеры включают морских морских пауков и мечехвостов , а также наземных паукообразных, таких как клещи , сенокосцы , пауки , скорпионы и родственные организмы, характеризующиеся наличием хелицер , придатков чуть выше или перед ротовым аппаратом . Хелицеры у скорпионов и мечехвостов появляются в виде крошечных клешней , которые они используют для питания, но у пауков они развились в виде клыков , впрыскивающих яд .
- К многоножкам относятся многоножки , многоножки , пауроподы и симфиланы , характеризующиеся наличием множества сегментов тела , каждый из которых несет одну или две пары ног (или в некоторых случаях вообще безногих). Все члены исключительно наземные.
- К панкрустообразным относятся остракоды , усоногие раки , веслоногие раки , малакостраканы , головоногие раки , жаберные моллюски , ремипеды и шестиногие моллюски . Большинство групп в основном ведут водный образ жизни (два заметных исключения — мокрицы и шестиногие, которые являются чисто наземными ) и характеризуются наличием двуветвистых придатков. Наиболее многочисленной группой панракообразных являются наземные шестиногие, в состав которых входят насекомые , двуногие , коллемболы и протураны с шестью грудными ногами.
Филогения . основных современных групп членистоногих была областью значительного интереса и споров [126] Недавние исследования убедительно свидетельствуют о том, что ракообразные в традиционном понимании являются парафилетическими , а Hexapoda развились изнутри них. [127] [128] так что Crustacea и Hexapoda образуют кладу Pancrustacea . Положение Myriapoda , Chelicerata и Pancrustacea по состоянию на апрель 2012 г. остается неясным. [update]. В некоторых исследованиях Myriapoda группируют с Chelicerata (образуя Myriochelata ); [129] [130] в других исследованиях Myriapoda группируется с Pancrustacea (образуя Mandibulata ), [127] или Myriapoda может быть сестрой Chelicerata плюс Pancrustacea. [128]
Следующая кладограмма показывает внутренние взаимоотношения между всеми современными классами членистоногих по состоянию на конец 2010-х годов. [131] [132] а также предполагаемые сроки для некоторых клад: [133]
| Членистоногие |
| |||||||||||||||||||||||||||||||||||||||||||||||||




Взаимодействие с людьми [ править ]

Ракообразные, такие как крабы , омары , раки , креветки и креветки , издавна были частью человеческой кухни, и теперь их выращивают в коммерческих целях. [134] Насекомые и их личинки, по крайней мере, так же питательны, как мясо, и во многих культурах их едят как сырыми, так и приготовленными, но не в большинстве европейских, индуистских и исламских культур. [135] [136] Вареные тарантулы считаются в Камбодже деликатесом . [137] [138] [139] и индейцами Пиароа на юге Венесуэлы после удаления сильно раздражающих волосков – основной защитной системы паука. [140] Люди также непреднамеренно едят членистоногих с другими продуктами питания. [141] а правила безопасности пищевых продуктов устанавливают приемлемые уровни загрязнения для различных видов пищевых материалов. [Примечание 4] [Примечание 5] Целенаправленное выращивание членистоногих и других мелких животных в пищу человеку, называемое мини-животноводством , в настоящее время становится в животноводстве . экологически обоснованной концепцией [145] Коммерческое разведение бабочек обеспечивает поголовье чешуекрылых для зимних садов бабочек , образовательных выставок, школ, исследовательских центров и культурных мероприятий.
Однако наибольший вклад членистоногих в снабжение человека продовольствием вносит опыление : исследование 2008 года изучило 100 культур, которые ФАО перечисляет как выращиваемые для производства продуктов питания, и оценило экономическую ценность опыления в 153 миллиарда евро, или 9,5 процента стоимости мирового сельскохозяйственного производства. продукции, используемой в пищу человеку в 2005 году. [146] Помимо опыления, пчелы производят мед , который является основой быстрорастущей промышленности и международной торговли. [147]
Красный краситель кошениль , производимый из насекомых Центральной Америки, имел экономическое значение для ацтеков и майя . [148] Хотя этот регион находился под контролем Испании , он стал . вторым по прибыльности экспортным товаром Мексики [149] и теперь восстанавливает часть позиций, которые он потерял у синтетических конкурентов. [150] Шеллак , смола, выделяемая видом насекомых, обитающих в Южной Азии, исторически использовалась в больших количествах для многих применений, в которых ее в основном заменяли синтетические смолы, но она все еще используется в деревообработке и в качестве пищевой добавки . Кровь мечехвостов содержит свертывающий агент Limulus Amebocyte Lysate , который сейчас используется для проверки в антибиотиках отсутствия опасных бактерий и почечных аппаратах , а также для выявления спинального менингита и некоторых видов рака . [151] Судебная энтомология использует доказательства членистоногих для установления времени, а иногда и места смерти человека, а в некоторых случаях и причины. [152] В последнее время насекомые также привлекли внимание как потенциальные источники лекарств и других лекарственных веществ. [153]
Относительная простота строения тела членистоногих, позволяющая им передвигаться по самым разным поверхностям как на суше, так и в воде, сделала их полезными в качестве моделей для робототехники . Избыточность, обеспечиваемая сегментами, позволяет членистоногим и биомиметическим роботам нормально передвигаться даже с поврежденными или утраченными придатками. [154] [155]
| Болезнь [156] | Насекомое | Случаев в год | Смертей в год |
|---|---|---|---|
| Малярия | анофелес Комар | 267 М | от 1 до 2 М |
| Лихорадка Денге | Aedes Комар | ? | ? |
| Желтая лихорадка | Aedes Комар | 4,432 | 1,177 |
| филяриоз | Кулекс комар | 250 М | неизвестный |
Хотя членистоногие являются самым многочисленным типом на Земле, а тысячи видов членистоногих ядовиты, они наносят человеку относительно мало серьезных укусов и ужалений. Гораздо более серьезными являются последствия для людей таких заболеваний, как малярия, переносимых кровососущими насекомыми. Другие кровососущие насекомые заражают домашний скот болезнями, которые убивают многих животных и значительно снижают полезность других. [156] Клещи могут вызывать клещевой паралич и некоторые паразитарные заболевания у людей. [157] Некоторые из близкородственных клещей также заражают людей, вызывая сильный зуд, [158] и другие вызывают аллергические заболевания, включая сенную лихорадку , астму и экзему . [159]
Многие виды членистоногих, главным образом насекомые, а также клещи, являются вредителями сельского хозяйства и леса. [160] [161] Клещ- деструктор Варроа стал крупнейшей проблемой, с которой сталкиваются пчеловоды во всем мире. [162] Усилия по борьбе с членистоногими вредителями путем широкомасштабного использования пестицидов привели к долгосрочным последствиям для здоровья человека и биоразнообразия . [163] Повышение устойчивости членистоногих к пестицидам привело к развитию комплексной борьбы с вредителями с использованием широкого спектра мер, включая биологический контроль . [160] Хищные клещи могут быть полезны для борьбы с некоторыми вредителями. [164] [165]
См. также [ править ]
Примечания [ править ]
- ↑ Музей Новой Зеландии отмечает, что «в повседневном разговоре» клоп «относится к наземным членистоногим, имеющим как минимум шесть ног, таким как насекомые, пауки и многоножки». [23] В главе «Насекомые, не являющиеся насекомыми» энтомолог Гилберт Вальбауэр описывает многоножек, многоножек, паукообразных (пауков, длинноногих папочек , скорпионов, клещей , чиггеров и клещей), а также немногочисленных наземных ракообразных ( насекомых и клопов ). [24] но утверждает, что «включение в число жуков безногих существ, таких как черви, слизни и улитки, слишком преувеличивает». [25]
- ^ «Было бы очень плохо, если бы вопрос о сегментации головы когда-либо был окончательно решен; он так долго был настолько благодатной почвой для теоретизирования, что членистоногие упустили бы его как поле для умственных упражнений». [48]
- ^ Первоначально ископаемое называлось Эотарбус, но было переименовано, когда выяснилось, что паукообразное каменноугольного периода уже носило имя Эотарбус . [98]
- ^ Упоминание о загрязнении насекомыми в международном стандарте качества пищевых продуктов см. в разделах 3.1.2 и 3.1.3 Кодекса 152 1985 г. Кодекса Алиментариус. [142]
- ^ Примеры количественного допустимого загрязнения пищевых продуктов насекомыми см. в последней записи («Пшеничная мука») и определении «Посторонние материалы» в Кодексе Алиментариус . [143] и стандарты, опубликованные FDA. [144]
Ссылки [ править ]
- ↑ Перейти обратно: Перейти обратно: а б с д Мартинес-Муньос, Карлос А. (4 мая 2023 г.). «Правильное авторство Arthropoda — переоценка» . Интегративная систематика . 6 (1): 1–8. дои : 10.18476/2023.472723 . ISSN 2628-2380 . S2CID 258497632 .
- ^ Грейвенхорст, JLC (1843 г.). Сравнительная зоология . Бреслау: Печать и издательство Graß, Barth and Comp.
- ^ Мойсюк Дж., Кэрон Дж.Б. (январь 2019 г.). «Окаменелости сланцев Бёрджесс проливают свет на проблему агностидов» . Слушания. Биологические науки . 286 (1894): 20182314. doi : 10.1098/rspb.2018.2314 . ПМК 6367181 . ПМИД 30963877 .
- ^ Фу, Д.; Легг, Д.А.; Дейли, AC; Бадд, GE; Ву, Ю.; Чжан, X. (2022). «Эволюция двуветвистых придатков, выявленная кембрийским членистоногим с панцирем» . Философские труды Королевского общества B: Биологические науки . 377 (1847): Идентификатор статьи 20210034. doi : 10.1098/rstb.2021.0034 . ПМЦ 8819368 . ПМИД 35125000 .
- ^ О'Флинн, Роберт Дж.; Уильямс, Марк; Ю, Мэнсяо; Харви, Томас; Лю, Ю (2022). «Новый эуартропод с крупными лобными придатками из биоты Чэнцзян раннего кембрия» . Электронная палеонтология . 25 (1): 1–21. дои : 10.26879/1167 . S2CID 246779634 .
- ↑ Перейти обратно: Перейти обратно: а б с Цзэн, Хан; Чжао, Фанчэн; Ню, Кеченг; Чжу, Маоянь; Хуан, Диин (декабрь 2020 г.). «Раннекембрийский эуартропод с хищными придатками, похожими на радиодонтов» . Природа . 588 (7836): 101–105. Бибкод : 2020Natur.588..101Z . дои : 10.1038/s41586-020-2883-7 . ISSN 1476-4687 . ПМИД 33149303 . S2CID 226248177 . Проверено 8 декабря 2020 г.
- ^ Легг, Дэвид А.; Саттон, Марк Д.; Эджкомб, Грегори Д. (30 сентября 2013 г.). «Данные об окаменелостях членистоногих повышают соответствие морфологической и молекулярной филогении» . Природные коммуникации . 4 (1): 2485. Бибкод : 2013NatCo...4.2485L . дои : 10.1038/ncomms3485 . ISSN 2041-1723 . ПМИД 24077329 .
- ^ Пульсифер, Массачусетс; Андерсон, EP; Райт, Л.С.; Клюссендорф, Дж.; Микулич, Д.Г.; Шиффбауэр, доктор юридических наук (2022). «Описание Acheronauta gen. nov., возможного мандибулата из силура Waukesha Lagerstätte, Висконсин, США». Журнал систематической палеонтологии . 20 (1). 2109216. дои : 10.1080/14772019.2022.2109216 . S2CID 252839113 .
- ^ Кларк, Нил Д.Л.; Фельдманн, Родни М; Шрам, Фредерик Р.; Швейцер, Кэрри Э (2020). «Переописание Americlus Rankini (Woodward, 1868) (Pancrustacea: Cyclida: Americlidae) и интерпретация его систематического размещения, морфологии и палеоэкологии» (PDF) . Журнал биологии ракообразных . 40 (2): 181–193. doi : 10.1093/jcbiol/ruaa001 .
- ^ Пил, Дж. С.; Штейн, М. «Новое членистоногое из нижнего кембрия Sirius Passet Fossil-Lagerstätten из Северной Гренландии» (PDF) . Бюллетень геонаук . 84 (4): 1158.
- ^ Эджкомб, Грегори Д. (1 сентября 2017 г.). «Выводы о филогении членистоногих: окаменелости и их взаимодействие с другими источниками данных» . Интегративная и сравнительная биология . 57 (3): 467–476. дои : 10.1093/icb/icx061 . ISSN 1540-7063 . ПМИД 28957518 .
- ^ Гарвуд, Р.; Саттон, М. (18 февраля 2012 г.), «Загадочное членистоногое Camptophyllia» , Palaeontologia Electronica , 15 (2): 12, doi : 10.1111/1475-4983.00174 , заархивировано (PDF) из оригинала 2 декабря 2013 г. , получено 11 июня. 2012 год
- ^ Чжай, Даю; Уильямс, Марк; Сиветер, Дэвид Дж.; Сиветер, Дерек Дж.; Харви, Томас HP; Сансом, Роберт С.; Май, Хуэйцзюань; Чжоу, Жуньцин; Хоу, Сянгуан (22 февраля 2022 г.). «Chuandianella ovata: раннекембрийское стеблевое моллюск с перистыми придатками» . Электронная палеонтология . 25 (1): 1–22. дои : 10.26879/1172 . ISSN 1094-8074 . S2CID 247123967 .
- ^ Валошек, Дитер; Мюллер, Клаус (1 октября 1990 г.). «Раки стволового происхождения верхнего кембрия и их влияние на монофилию ракообразных и положение агностуса» . Летайя . 23 : 409–427. дои : 10.1111/j.1502-3931.1990.tb01373.x .
- ^ Ван Рой, Питер; Рак, Штепан; Будил, Петр; Фатка, Олдржих (13 июня 2022 г.). «Переописание хелониеллидного эуартропода Triopus draboviensis из верхнего ордовика Богемии с комментариями о сходстве Parioscorpio venator ». Геологический журнал . 159 (9): 1471–1489. Бибкод : 2022ГеоМ..159.1471В . дои : 10.1017/s0016756822000292 . hdl : 1854/LU-8756253 . ISSN 0016-7568 . S2CID 249652930 .
- ^ Хауг, Дж. Т.; Маас, А.; Хауг, К.; Валошек, Д. (1 ноября 2011 г.). «Sarotrocercus oblitus — маленькое членистоногое, оказавшее большое влияние на понимание эволюции членистоногих?» . Бюллетень геонаук : 725–736. дои : 10.3140/bull.geosci.1283 . ISSN 1802-8225 .
- ^ Ортега-Эрнандес, Хавьер; Легг, Дэвид А.; Брэдди, Саймон Дж. (2013). «Филогения агласпидовых членистоногих и внутренние взаимоотношения внутри Artiopoda» . Кладистика . 29 (1): 15–45. дои : 10.1111/j.1096-0031.2012.00413.x . ПМИД 34814371 . S2CID 85744103 .
- ^ Кюль, Габриель; Раст, Джес (2009). « Devonoгексаподус bocksbergensis является синонимом Wingertshellicus backesi (Euarthropoda) – нет свидетельств существования морских шестиногих моллюсков, обитающих в девонском море Хунсрюк» . Разнообразие и эволюция организмов . 9 (3): 215–231. Бибкод : 2009ODivE...9..215K . дои : 10.1016/j.ode.2009.03.002 .
- ^ Пейтс, Стивен; Лерози-Обрил, Руди; Дейли, Эллисон С.; Кир, Карло; Бонино, Энрико; Ортега-Эрнандес, Хавьер (19 января 2021 г.). «Разнообразная фауна радиодонтов из формации Марджум в штате Юта, США (кембрий: драмиан)» . Палеонтология и эволюционная наука . 9 : е10509. дои : 10.7717/peerj.10509 . ПМЦ 7821760 . ПМИД 33552709 .
- ^ «членистоногие» . Словарь Merriam-Webster.com . Проверено 12 мая 2024 г.
- ^ «Членистоногие» . Интернет-словарь этимологии . Архивировано из оригинала 7 марта 2013 года . Проверено 23 мая 2013 г.
- ^ Грейвенхорст, JLC (1843 г.). Сравнительная зоология [ Сравнительная зоология ] (на немецком языке). Бреслау, (Пруссия): Graß, Barth & Comp. п. раскладной.
«С шарнирными органами движения»
- ^ «Что такое ошибка? Насекомые, паукообразные и многоножки» на сайте Музея Новой Зеландии Те Папа Тонгарева. По состоянию на 10 марта 2022 г.
- ^ Гилберт Вальдбауэр. Удобная книга ответов на ошибки. Видимые чернила, 1998. стр. 5–26. ISBN 978-1-57859-049-0
- ↑ Перейти обратно: Перейти обратно: а б с Гилберт Вальдбауэр. Удобная книга ответов на ошибки. Видимые чернила, 1998. с. 1. ISBN 978-1-57859-049-0
- ^ Валентайн, JW (2004), О происхождении Филы , University of Chicago Press , стр. 33, ISBN 978-0-226-84548-7
- ^ Катлер, Б. (август 1980 г.), «Особенности кутикулы членистоногих и монофилия членистоногих», Cellular and Molecular Life Sciences , 36 (8): 953, doi : 10.1007/BF01953812 , S2CID 84995596
- ^ Австралийские жуки, том 2: Archostemata, Myxophaga, Adephaga, Polyphaga.
- ^ Ковур, Дж. (1978). «Естественная кальцификация просоматического эндостернита у Phalangiidae (Arachnida: Opiliones)». Исследование кальцифицированной ткани . 26 (3): 267–9. дои : 10.1007/BF02013269 . ПМИД 750069 . S2CID 23119386 .
- ^ Ронг, Цзинцзин; Лин, Юбо; Суй, Чжуосяо; Ван, Сидзя; Вэй, Сюньфань; Сяо, Цзиньхуа; Хуан, Давэй (ноябрь – декабрь 2019 г.). «Аморфный фосфат кальция в кутикуле куколки Bactrocera dorsalis Hendel (Diptera: Tephritidae): новое открытие, позволяющее пересмотреть минерализацию кутикулы насекомых» . Журнал физиологии насекомых . 119 : 103964. Бибкод : 2019JInsP.11903964R . дои : 10.1016/j.jinsphys.2019.103964 . ПМИД 31604063 .
- ^ Танукос, Анна, История членистоногих , Калифорнийский университет, Беркли , заархивировано из оригинала 16 июня 2008 г. , получено 29 сентября 2008 г.
- ^ Одегаард, Фроде (декабрь 2000 г.), «Сколько видов членистоногих? Пересмотренная оценка Эрвина» (PDF) , Биологический журнал Линнеевского общества , 71 (4): 583–597, Бибкод : 2000BJLS...71..583O , doi : 10.1006/bijl.2000.0468 , заархивировано (PDF) из оригинала 26 декабря 2010 г. , получено 6 мая 2010 г.
- ↑ Перейти обратно: Перейти обратно: а б Томпсон, Дж. Н. (1994), Коэволюционный процесс , University of Chicago Press , стр. 9, ISBN 978-0-226-79760-1
- ↑ Перейти обратно: Перейти обратно: а б с д и ж г час я дж к л Руперт, Фокс и Барнс (2004) , стр. 518–522.
- ^ Инга Морбек; Педро Мартинес Арбису; Томас Глатцель (октябрь 2010 г.). «Tantulocarida (Crustacea) глубоководных районов Южного океана и описание трех новых видов Tantulacus Huys, Andersen & Kristensen, 1992». Систематическая паразитология . 77 (2): 131–151. дои : 10.1007/s11230-010-9260-0 . ПМИД 20852984 . S2CID 7325858 .
- ^ Шмидт-Нильсен, Кнут (1984), «Прочность костей и скелетов» , Масштабирование: почему размер животного так важен? , Издательство Кембриджского университета , стр. 42–55 , ISBN. 978-0-521-31987-4
- ↑ Перейти обратно: Перейти обратно: а б с Лю, Ю; Эджкомб, Грегори Д.; Шмидт, Мишель; Бонд, Эндрю Д.; Мельцер, Роланд Р.; Чжай, Даю; Май, Хуэйцзюань; Чжан, Маоинь; Хоу, Сянгуан (30 июля 2021 г.). «Экситы у кембрийских членистоногих и гомология ветвей конечностей членистоногих» . Природные коммуникации . 12 (1): 4619. Бибкод : 2021NatCo..12.4619L . дои : 10.1038/s41467-021-24918-8 . ISSN 2041-1723 . ПМЦ 8324779 . ПМИД 34330912 .
- ↑ Перейти обратно: Перейти обратно: а б Гулд (1990) , стр. 102–106.
- ↑ Перейти обратно: Перейти обратно: а б Ортега-Эрнандес, Хавьер; Янссен, Ральф; Бадд, Грэм Э. (2017). «Происхождение и эволюция головы панартропод - палеобиологическая перспектива и перспектива развития» . Строение и развитие членистоногих . 46 (3): 354–379. Бибкод : 2017ArtSD..46..354O . дои : 10.1016/j.asd.2016.10.011 . ПМИД 27989966 .
- ^ «Гигантское морское существо намекает на раннюю эволюцию членистоногих» . 11 марта 2015 г. Архивировано из оригинала 2 февраля 2017 г. Проверено 22 января 2017 г.
- ↑ Перейти обратно: Перейти обратно: а б Фу, Д.; Легг, Д.А.; Дейли, AC; Бадд, GE; Ву, Ю.; Чжан, X. (2022). «Эволюция двуветвистых придатков, выявленная кембрийским членистоногим с панцирем» . Философские труды Королевского общества B: Биологические науки . 377 (1847): Идентификатор статьи 20210034. doi : 10.1098/rstb.2021.0034 . ПМЦ 8819368 . ПМИД 35125000 . S2CID 246608509 .
- ^ Хейнол, Андреас; Шольц, Герхард (1 октября 2004 г.). «Клональный анализ бесдистальных и закреплённых паттернов экспрессии во время раннего морфогенеза одноветвистых и двуветвистых конечностей ракообразных». Гены развития и эволюция . 214 (10): 473–485. дои : 10.1007/s00427-004-0424-2 . ISSN 1432-041X . ПМИД 15300435 . S2CID 22426697 .
- ^ Вольф, Карстен; Шольц, Герхард (7 мая 2008 г.). «Клональный состав двуветвистых и одноветвистых конечностей членистоногих» . Труды Королевского общества B: Биологические науки . 275 (1638): 1023–1028. дои : 10.1098/rspb.2007.1327 . ПМК 2600901 . ПМИД 18252674 .
- ^ Шубин, Нил ; Табин, К.; Кэрролл, Шон (2000), «Ископаемые, гены и эволюция конечностей животных» , в книге Джи, Х. (редактор), « Встряхивание дерева: чтения природы в истории жизни» , University of Chicago Press , стр. 110, ISBN 978-0-226-28497-2
- ^ Данлоп, Джейсон А.; Ламсделл, Джеймс К. (2017). «Сегментация и тагмос у Chelicerata» . Строение и развитие членистоногих . 46 (3): 395–418. Бибкод : 2017ArtSD..46..395D . дои : 10.1016/j.asd.2016.05.002 . ПМИД 27240897 .
- ↑ Перейти обратно: Перейти обратно: а б Уиттингтон, HB (1971), «Переописание Marrella splendens (Trilobitoidea) из сланцев Бёрджесс, средний кембрий, Британская Колумбия», Бюллетень геологической службы Канады , 209 : 1–24 , кратко изложенное в Gould (1990) , стр. 107–121. .
- ^ Бадд, GE (16 мая 2002 г.). «Палеонтологическое решение проблемы головы членистоногих». Природа . 417 (6886): 271–275. Бибкод : 2002Natur.417..271B . дои : 10.1038/417271a . ПМИД 12015599 . S2CID 4310080 .
- ^ Снодграсс, Р.Э. (1960), «Факты и теории, касающиеся головы насекомых», Разные коллекции Смитсоновского института , 142 : 1–61.
- ↑ Перейти обратно: Перейти обратно: а б Уэйнрайт, ЮАР; Биггс, В.Д. и Гослайн, Дж.М. (1982). Механическое проектирование в организмах . Издательство Принстонского университета . стр. 162–163 . ISBN 978-0-691-08308-7 .
- ^ Лоуэнстам, штат Ха; Вайнер С. (1989), О биоминерализации , Oxford University Press, стр. 111, ISBN 978-0-19-504977-0
- ^ Дзик, Дж (2007), «Синдром Вердена: одновременное возникновение защитных панцирей и инфаунистических укрытий в период докембрия-кембрия» (PDF) , в Викерс-Рич, Патрисия; Комарауэр, Патрисия (ред.), Взлет и падение эдиакарской биоты , Специальные публикации, том. 286, Лондон: Геологическое общество, стр. 405–414, doi : 10.1144/SP286.30 , ISBN. 978-1-86239-233-5 , OCLC 156823511
- ↑ Перейти обратно: Перейти обратно: а б Коэн, Б.Л. (2005), «Не броня, а биомеханика, экологические возможности и повышенная плодовитость как ключи к происхождению и расширению минерализованной донной многоклеточной фауны» (PDF) , Биологический журнал Линнеевского общества , 85 (4): 483 –490, doi : 10.1111/j.1095-8312.2005.00507.x , заархивировано (PDF) из оригинала 3 октября 2008 г. , получено 25 сентября 2008 г.
- ^ Бенгтсон, С. (2004). «Ранние скелетные окаменелости». Ин Липпс, Дж. Х.; Ваггонер, Б.М. (ред.). Неопротерозойско-кембрийские биологические революции (PDF) . Документы Палеонтологического общества. Том. 10. С. 67–78. дои : 10.1017/S1089332600002345 . Архивировано из оригинала (PDF) 3 октября 2008 года.
- ^ Барнс, РСК; Калоу, П.; Олив, П.; Голдинг Д. и Спайсер Дж. (2001), «Беспозвоночные с ногами: членистоногие и подобные группы» , «Беспозвоночные: синтез» , Blackwell Publishing , стр. 168, ISBN 978-0-632-04761-1
- ^ Парри, Д.А. и Браун, Р.Х.Дж. (1959), «Гидравлический механизм ноги паука» (PDF) , Журнал экспериментальной биологии , 36 (2): 423–433, номер документа : 10.1242/jeb.36.2.423 , в архиве ( PDF) из оригинала от 3 октября 2008 г. , получено 25 сентября 2008 г.
- ↑ Перейти обратно: Перейти обратно: а б с Руперт, Фокс и Барнс (2004) , стр. 523–524.
- ↑ Перейти обратно: Перейти обратно: а б с Руперт, Фокс и Барнс (2004) , стр. 527–528.
- ↑ Перейти обратно: Перейти обратно: а б с Гарвуд, Рассел Дж.; Эджкомб, Грег (2011). «Ранние наземные животные, эволюция и неопределенность» . Эволюция: образование и информационно-пропагандистская деятельность . 4 (3): 489–501. дои : 10.1007/s12052-011-0357-y .
- ^ Рупперт, Fox & Barnes (2004) , стр. 530, 733.
- ^ Рупперт, Fox & Barnes (2004) , стр. 531–532.
- ↑ Перейти обратно: Перейти обратно: а б с д Руперт, Фокс и Барнс (2004) , стр. 529–530.
- ↑ Перейти обратно: Перейти обратно: а б с д и ж г Руперт, Фокс и Барнс (2004) , стр. 532–537.
- ^ Рупперт, Fox & Barnes (2004) , стр. 578–580.
- ^ Фёлькель, Р.; Эйснер, М.; Вейбл, К.Дж. (июнь 2003 г.). «Миниатюрные системы визуализации» (PDF) . Микроэлектронная инженерия . 67–68: 461–472. дои : 10.1016/S0167-9317(03)00102-3 . Архивировано из оригинала (PDF) 1 октября 2008 года.
- ↑ Перейти обратно: Перейти обратно: а б с д Руперт, Фокс и Барнс (2004) , стр. 537–539.
- ^ Олив, PJW (2001). «Размножение и жизненные циклы беспозвоночных». Энциклопедия наук о жизни . Джон Уайли и сыновья. дои : 10.1038/npg.els.0003649 . ISBN 978-0-470-01617-6 .
- ^ Щурко А.М.; Мазур, диджей; Логсдон, Дж. М. (февраль 2010 г.). «Инвентаризация и филогеномное распределение мейотических генов у Nasonia vitripennis и среди различных членистоногих». Молекулярная биология насекомых . 19 (Приложение 1): 165–180. дои : 10.1111/j.1365-2583.2009.00948.x . ПМИД 20167026 . S2CID 11617147 .
- ^ Бернштейн, Х.; Хопф, ФА; Мишод, RE (1987). «Молекулярная основа эволюции пола». Молекулярная генетика развития . Достижения генетики. Том. 24. С. 323–370. дои : 10.1016/s0065-2660(08)60012-7 . ISBN 978-0-12-017624-3 . ПМИД 3324702 .
- ^ «Факты о мечехвостах и часто задаваемые вопросы» . Проверено 19 января 2020 г.
- ^ Лоренсо, Уилсон Р. (2002), «Размножение скорпионов с особым упором на партеногенез», в Тофт, С.; Шарфф, Н. (ред.), Европейская арахнология 2000 (PDF) , Издательство Орхусского университета , стр. 71–85, ISBN 978-87-7934-001-5 , заархивировано (PDF) из оригинала 3 октября 2008 г. , получено 28 сентября 2008 г.
- ^ Трумэн, Дж.В.; Риддифорд, LM (сентябрь 1999 г.). «Происхождение метаморфоза насекомых» (PDF) . Природа . 401 (6752): 447–452. Бибкод : 1999Natur.401..447T . дои : 10.1038/46737 . ПМИД 10519548 . S2CID 4327078 . Архивировано (PDF) из оригинала 3 октября 2008 г. Проверено 28 сентября 2008 г.
- ^ Смит, Г., Разнообразие и адаптация водных насекомых (PDF) , Новый колледж Флориды , заархивировано из оригинала (PDF) 3 октября 2008 г. , получено 28 сентября 2008 г.
- ↑ Перейти обратно: Перейти обратно: а б Бергстрем, Ян; Хоу, Сянь-Гуан (2005), «Раннепалеозойские непластинчатые членистоногие», у Стефана Кенеманна; Рональд А. Дженнер (ред.), Взаимоотношения ракообразных и членистоногих , Проблемы ракообразных, том. 16, Бока-Ратон: Тейлор и Фрэнсис , стр. 73–93, doi : 10.1201/9781420037548.ch4 , ISBN 978-0-8493-3498-6
- ^ Маккивер, Конор (30 сентября 2016 г.). «Предок членистоногих имел пасть полового червя» . Музей естественной истории . Архивировано из оригинала 2 февраля 2017 года.
- ^ Глесснер, МФ (1958). «Новые окаменелости из основания кембрия в Южной Австралии» (PDF) . Труды Королевского общества Южной Австралии . 81 : 185–188. Архивировано из оригинала (PDF) 16 декабря 2008 года.
- ^ Лин, JP; Гон, С.М.; Гелинг, Дж.Г.; Бэбкок, Луизиана; Чжао, ЮЛ; Чжан, XL; Ху, SX; Юань, JL; Ю, МОЙ; Пэн, Дж. (2006). « Парванкориноподобное членистоногое из кембрия Южного Китая». Историческая биология . 18 (1): 33–45. Бибкод : 2006HBio...18...33L . дои : 10.1080/08912960500508689 . S2CID 85821717 .
- ^ МакМенамин, MAS (2003), « Сприггина - трилобитоид экдисозой» (аннотация) , Abstracts with Programs , 35 (6): 105, заархивировано из оригинала 30 августа 2008 г. , получено 21 октября 2008 г.
- ^ Дейли, Эллисон С.; Антклифф, Джонатан Б.; Дрейдж, Харриет Б.; Пейтс, Стивен (22 мая 2018 г.). «Ранние окаменелости Euarthropoda и кембрийский взрыв» . Труды Национальной академии наук Соединенных Штатов Америки . 115 (21): 5323–5331. Бибкод : 2018PNAS..115.5323D . дои : 10.1073/pnas.1719962115 . ПМК 6003487 . ПМИД 29784780 .
- ^ Браун, А.; Чен, Дж.; Валошек, Д.; Маас, А. (2007). «Первые раннекембрийские радиолярии» (PDF) . Специальные публикации . 286 (1): 143–149. Бибкод : 2007GSLSP.286..143B . дои : 10.1144/SP286.10 . S2CID 129651908 . Архивировано из оригинала (PDF) 18 июля 2011 года.
- ^ Юань, X.; Сяо, С.; Петрушка, РЛ; Чжоу, К.; Чен, З.; Ху, Дж. (апрель 2002 г.). «Высокие губки в раннекембрийском Lagerstätte: Несоответствие между недвусторонними и двусторонними эпифаунистическими ярусами на переходном этапе неопротерозой-кембрий» . Геология . 30 (4): 363–366. Бибкод : 2002Geo....30..363Y . doi : 10.1130/0091-7613(2002)030<0363:TSIAEC>2.0.CO;2 .
- ^ Сковстед, Кристиан; Брок, Гленн; Патерсон, Джон (2006), «Двустворчатые членистоногие из формации Мернмерна нижнего кембрия в Южной Австралии и их значение для идентификации кембрийских« мелких ракушковых окаменелостей » » , Мемуары Ассоциации австралийских палеонтологов , 32 : 7–41, ISSN 0810-8889
- ^ Беттс, Марисса; Топпер, Тимоти; Валентин, Джеймс; Сковстед, Кристиан; Патерсон, Джон; Брок, Гленн (январь 2014 г.), «Новый раннекембрийский комплекс брадориид (Arthropoda) из северных хребтов Флиндерс, Южная Австралия» , Gondwana Research , 25 (1): 420–437, Бибкод : 2014GondR..25..420B , дои : 10.1016/j.gr.2013.05.007
- ^ Либерман, Б.С. (1 марта 1999 г.), «Проверка дарвиновского наследия кембрийского излучения с использованием филогении и биогеографии трилобитов» , Journal of Paleontology , 73 (2): 176, Бибкод : 1999JPal...73..176L , doi : 10.1017 /S0022336000027700 , S2CID 88588171 , заархивировано из оригинала 19 октября 2008 г. , получено 21 октября 2008 г.
- ↑ Перейти обратно: Перейти обратно: а б «Пятиглазая окаменелость возрастом 520 миллионов лет свидетельствует о происхождении членистоногого» . физ.орг . Проверено 8 декабря 2020 г.
- ^ Уиттингтон, HB (1979). Ранние членистоногие, их придатки и взаимоотношения. В MR House (ред.), «Происхождение основных групп беспозвоночных» (стр. 253–268). Специальный том Ассоциации систематики, 12. Лондон: Academic Press.
- ^ Уиттингтон, HB ; Геологическая служба Канады (1985), Сланец Берджесс , издательство Йельского университета, ISBN 978-0-660-11901-4 , OCLC 15630217
- ^ Гулд (1990) , с. [ нужна страница ] .
- ^ Гарсиа-Беллидо, округ Колумбия; Коллинз, Д.Х. (май 2004 г.). «Линьющее членистоногое поймано с поличным» . Природа . 429 (6987): 40. Бибкод : 2004Natur.429...40G . дои : 10.1038/429040a . ПМИД 15129272 . S2CID 40015864 .
- ↑ Перейти обратно: Перейти обратно: а б с Хегна, Томас А.; Луке, Хавьер; Вулф, Джоанна М. (10 сентября 2020 г.). «Ископаемая летопись панкрустацеи» . Эволюция и биогеография . Издательство Оксфордского университета: 21–52. дои : 10.1093/oso/9780190637842.003.0002 . ISBN 978-0-19-063784-2 . Проверено 5 января 2024 г.
- ^ Хоу, Сянь-Гуан; Сиветер, Дерек Дж.; Олдридж, Ричард Дж.; Сиветер, Дэвид Дж. (10 октября 2008 г.). «Коллективное поведение раннекембрийских членистоногих» . Наука . 322 (5899): 224. Бибкод : 2008Sci...322..224H . дои : 10.1126/science.1162794 . ISSN 0036-8075 . ПМИД 18845748 .
- ^ Бадд, GE; Баттерфилд, Нью-Джерси; Дженсен, С. (декабрь 2001 г.), «Рачки и «кембрийский взрыв» », Science , 294 (5549): 2047, doi : 10.1126/science.294.5549.2047a , PMID 11739918
- ^ Сиань-Гуан, Хоу; Сиветер, Дерек Дж.; Олдридж, Ричард Дж.; Сиветер, Дэвид Дж. (2009). «Новое членистоногое в цепочечных ассоциациях из Cnengjiang Lagerstätte (нижний кембрий), Юньнань, Китай» . Палеонтология . 52 (4): 951–961. Бибкод : 2009Palgy..52..951X . дои : 10.1111/j.1475-4983.2009.00889.x . ISSN 0031-0239 .
- ^ Чжан, X.-G.; Сивер, диджей; Валошек, Д.; Маас, А. (октябрь 2007 г.). «Рачок кроновой группы, несущий эпиподит, из нижнего кембрия». Природа . 449 (7162): 595–598. Бибкод : 2007Natur.449..595Z . дои : 10.1038/nature06138 . ПМИД 17914395 . S2CID 4329196 .
- ^ Пизани, Д.; Полинг, LL; Лайонс-Вейлер М.; Хеджес, С.Б. (2004). «Колонизация суши животными: молекулярная филогения и время дивергенции среди членистоногих» . БМК Биология . 2 :1. дои : 10.1186/1741-7007-2-1 . ПМЦ 333434 . ПМИД 14731304 .
- ^ Коуэн, Р. (2000). История жизни (3-е изд.). Блэквелл Наука. п. 126. ИСБН 978-0-632-04444-3 .
- ^ Брэдди, С.Дж.; Маркус Пошманн, М. и Тетли, OE (2008). «Гигантский коготь обнаружил самое большое членистоногое в истории» . Письма по биологии . 4 (1): 106–109. дои : 10.1098/rsbl.2007.0491 . ПМЦ 2412931 . ПМИД 18029297 .
- ^ Данлоп, JA (сентябрь 1996 г.). «Тригонотарбидный паукообразный из верхнего силура Шропшира» (PDF) . Палеонтология . 39 (3): 605–614. Архивировано из оригинала (PDF) 16 декабря 2008 года.
- ^ Данлоп, Дж.А. (1999). «Заменительное название тригонотарбидного паукообразного Эотарбуса Данлопа» . Палеонтология . 42 (1): 191. Бибкод : 1999Palgy..42..191D . дои : 10.1111/1475-4983.00068 . S2CID 83825904 .
- ^ Селден, Пенсильвания; Шир, Вашингтон (декабрь 2008 г.). «Ископаемые свидетельства происхождения пауков-прядильщиков» . ПНАС . 105 (52): 20781–5. Бибкод : 2008PNAS..10520781S . дои : 10.1073/pnas.0809174106 . ПМЦ 2634869 . ПМИД 19104044 .
- ^ Селден, Пенсильвания (февраль 1996 г.). «Ископаемые мезотелиальные пауки». Природа . 379 (6565): 498–499. Бибкод : 1996Natur.379..498S . дои : 10.1038/379498b0 . S2CID 26323977 .
- ^ Воллрат, Ф. и Селден, Пенсильвания (декабрь 2007 г.). «Роль поведения в эволюции пауков, шелка и паутины» (PDF) . Ежегодный обзор экологии, эволюции и систематики . 38 : 819–846. doi : 10.1146/annurev.ecolsys.37.091305.110221 . Архивировано из оригинала (PDF) 9 декабря 2008 года.
- ^ Андерсон, Эван П.; Шиффбауэр, Джеймс Д.; Жаке, Сара М.; Ламсделл, Джеймс К.; Клюссендорф, Джоан; Микулич, Дональд Г. (2021). Чжан, Си-Гуан (ред.). «Незнакомец, чем скорпион: переоценка Parioscorpio venator, проблемного членистоногого из лландоверского Waukesha Lagerstätte». Палеонтология . 64 (3): 429–474. Бибкод : 2021Palgy..64..429A . дои : 10.1111/пала.12534 . ISSN 0031-0239 .
- ^ Джерам, Эй-Джей (январь 1990 г.). «Книжные легкие скорпиона нижнего карбона». Природа . 343 (6256): 360–361. Бибкод : 1990Natur.343..360J . дои : 10.1038/343360a0 . S2CID 4327169 .
- ^ Ховард, Ричард Дж.; Эджкомб, Грегори Д.; Легг, Дэвид А.; Пизани, Давиде; Лосано-Фернандес, Хесус (1 марта 2019 г.). «Изучение эволюции и наземного развития скорпионов (Arachnida: Scorpiones) с помощью камней и часов» . Разнообразие и эволюция организмов . 19 (1): 71–86. дои : 10.1007/s13127-019-00390-7 . hdl : 1983/9ab6548b-b4de-47b5-b1d0-8008d225c375 . ISSN 1618-1077 .
- ^ Пошманн, Маркус; Данлоп, Джейсон А.; Каменц, Карстен; Шольц, Герхард (декабрь 2008 г.). «Нижнедевонский скорпион Waeringoscorpio и дыхательная природа его нитевидных структур с описанием нового вида из района Вестервальда, Германия». Палеонтологический журнал . 82 (4): 418–436. Бибкод : 2008PalZ...82..418P . дои : 10.1007/BF03184431 . ISSN 0031-0220 .
- ^ Энгель, М.С. ; Гримальди, Д.А. (февраль 2004 г.). «Новый свет, пролитый на старейшее насекомое». Природа . 427 (6975): 627–630. Бибкод : 2004Natur.427..627E . дои : 10.1038/nature02291 . ПМИД 14961119 . S2CID 4431205 .
- ↑ Перейти обратно: Перейти обратно: а б Хауг, Кэролин; Хауг, Иоахим Т. (30 мая 2017 г.). «Предположительно старейшее летающее насекомое: скорее многоножка?» . ПерДж . 5 : е3402. дои : 10.7717/peerj.3402 . ПМЦ 5452959 . ПМИД 28584727 .
- ^ Лабандейра, К.; Эбл, Дж.Дж. (2000). «Ископаемая летопись разнообразия и неравенства насекомых». В Андерсоне, Дж.; Теккерей, Ф.; ван Вик, Б.; де Вит, М. (ред.). Гондвана жива: биоразнообразие и развивающаяся биосфера (PDF) . Издательство Университета Витватерсранда . Архивировано из оригинала (PDF) 11 сентября 2008 года . Проверено 21 октября 2008 г.
- ↑ Перейти обратно: Перейти обратно: а б с Бадд, GE (1996). «Морфология Opabinia regalis и реконструкция стеблевой группы членистоногих». Летайя . 29 (1): 1–14. Бибкод : 1996Лета..29....1Б . дои : 10.1111/j.1502-3931.1996.tb01831.x .
- ^ Гиллотт, К. (1995). Энтомология . Спрингер. стр. 17–19. ISBN 978-0-306-44967-3 .
- ^ Адрен, Дж. (15 марта 1999 г.). « Окаменелости и филогения членистоногих » под редакцией Грегори Д. Эджкомба . Рецензия на книгу. Электронная палеонтология . Архивировано из оригинала 8 сентября 2008 года . Проверено 28 сентября 2008 г.
- Книга
- ^ Чен, Ж.-Ю.; Эджкомб, Джорджия; Рамшельд, Л.; Чжоу, Г.-К. (2 июня 1995 г.). «Сегментация головы у раннего кембрия Fuxianhuia : значение для эволюции членистоногих». Наука . 268 (5215): 1339–1343. Бибкод : 1995Sci...268.1339C . дои : 10.1126/science.268.5215.1339 . ПМИД 17778981 . S2CID 32142337 .
- ^ Бадд, GE (1993). «Кембрийский жаберный лобопод из Гренландии». Природа . 364 (6439): 709–711. Бибкод : 1993Natur.364..709B . дои : 10.1038/364709a0 . S2CID 4341971 .
- ^ Нильсен, К. (2001). Эволюция животных: взаимоотношения живых типов (2-е изд.). Издательство Оксфордского университета . стр. 194–196. ISBN 978-0-19-850681-2 .
- ^ Хоу, С.-Г. ; Бергстрем, Дж.; Цзе, Ю. (2006). «Отличие аномалокаридид от членистоногих и приапулид». Геологический журнал . 41 (3–4): 259–269. Бибкод : 2006GeolJ..41..259X . дои : 10.1002/gj.1050 . S2CID 83582128 .
- ^ «Неправильно понятое червеобразное ископаемое находит свое место на Древе Жизни» (пресс-релиз). Кембриджский университет . 17 августа 2014 года. Архивировано из оригинала 7 января 2017 года . Проверено 24 января 2017 г.
- ↑ Перейти обратно: Перейти обратно: а б Телфорд, MJ; Бурла, SJ; Эконому, А.; Папийон, Д.; Рота-Стабелли, О. (январь 2008 г.). «Эволюция Ecdysozoa» . Философские труды Королевского общества B: Биологические науки . 363 (1496): 1529–1537. дои : 10.1098/rstb.2007.2243 . ПМК 2614232 . ПМИД 18192181 .
- ^ Ваккари, штат Невада; Эджкомб, Джорджия; Эскудеро, К. (29 июля 2004 г.). «Кембрийское происхождение и сходство загадочной ископаемой группы членистоногих». Природа . 430 (6999): 554–557. Бибкод : 2004Natur.430..554V . дои : 10.1038/nature02705 . ПМИД 15282604 . S2CID 4419235 .
- ^ Шмидт-Раэса, А.; Бартоломеус, Т.; Лембург, К.; Элерс, У.; Гэри, младший (январь 1999 г.). «Положение членистоногих в филогенетической системе». Журнал морфологии . 238 (3): 263–285. doi : 10.1002/(SICI)1097-4687(199812)238:3<263::AID-JMOR1>3.0.CO;2-L . ПМИД 29852696 . S2CID 46920478 .
- ↑ Перейти обратно: Перейти обратно: а б с д Грегори Д. Эджкомб (2020). «Происхождение членистоногих: объединение палеонтологических и молекулярных данных». Анну. Преподобный Экол. Эвол. Сист . 51 : 1–25. doi : 10.1146/annurev-ecolsys-011720-124437 . S2CID 225478171 .
- ↑ Перейти обратно: Перейти обратно: а б с д Ортега-Эрнандес, Хавьер (2016). «Осмысление «нижней» и «верхней» стеблевой группы Euarthropoda с комментариями о строгом использовании названия Arthropoda von Siebold, 1848». Биологические обзоры . 91 (1): 255–273. дои : 10.1111/brv.12168 . ПМИД 25528950 . S2CID 7751936 .
- ↑ Перейти обратно: Перейти обратно: а б с Ария, Седрик (26 апреля 2022 г.). «Происхождение и ранняя эволюция членистоногих» . Биологические обзоры . 97 (5): 1786–1809. дои : 10.1111/brv.12864 . ISSN 1464-7931 . ПМИД 35475316 . S2CID 243269510 .
- ^ Дженнер, РА (апрель 2006 г.). «Сложные знания: некоторый вклад новой микроскопии в новую филогению животных» . Интегративная и сравнительная биология . 46 (2): 93–103. дои : 10.1093/icb/icj014 . ПМИД 21672726 .
- ^ Данлоп, Джейсон А. (31 января 2011 г.). «Ископаемый фокус: Хелицераты» . Палеонтология онлайн . стр. 1–8. Архивировано из оригинала 12 сентября 2017 года . Проверено 15 марта 2018 г.
- ^ «Членистоногие» . Интегрированная таксономическая информационная система . Проверено 15 августа 2006 г.
- ^ Карапелли, Антонио; Лио, Пьетро; Нарди, Франческо; ван дер Ват, Элизабет; Фрати, Франческо (16 августа 2007 г.). «Филогенетический анализ генов, кодирующих митохондриальные белки, подтверждает взаимную парафилию Hexapoda и Crustacea» . Эволюционная биология BMC . 7 (Дополнение 2): S8. Бибкод : 2007BMCEE...7S...8C . дои : 10.1186/1471-2148-7-S2-S8 . ЧВК 1963475 . ПМИД 17767736 .
- ↑ Перейти обратно: Перейти обратно: а б Режье, Джером К.; Шульц, Дж.В.; Цвик, А.; Хасси, А.; Болл, Б.; Ветцер, Р.; Мартин, JW; Каннингем, CW; и др. (2010). «Отношения членистоногих, выявленные с помощью филогеномного анализа последовательностей, кодирующих ядерные белки». Природа . 463 (7284): 1079–1084. Бибкод : 2010Natur.463.1079R . дои : 10.1038/nature08742 . ПМИД 20147900 . S2CID 4427443 .
- ↑ Перейти обратно: Перейти обратно: а б фон Роймонт, Бьерн М.; Дженнер, Рональд А.; Уиллс, Мэтью А.; Делл'Ампио, Эмилиано; Пасс, Гюнтер; Эберсбергер, Инго; Мейер, Бенджамин; Кенеманн, Стефан; Илифф, Томас М.; Стаматакис, Александрос; Нихейс, Оливер; Мейсеманн, Карен; Мисоф, Бернхард (2011). «Филогения панкрустообразных в свете новых филогеномических данных: поддержка Remipedia как возможной сестринской группы Hexapoda» . Молекулярная биология и эволюция . 29 (3): 1031–45. дои : 10.1093/molbev/msr270 . ПМИД 22049065 .
- ^ Хасанин, Александр (2006). «Филогения членистоногих, выведенная на основе митохондриальных последовательностей: стратегии ограничения вводящих в заблуждение эффектов множественных изменений в структуре и скорости замен» (PDF) . Молекулярная филогенетика и эволюция . 38 (1): 100–116. Бибкод : 2006MolPE..38..100H . дои : 10.1016/j.ympev.2005.09.012 . ПМИД 16290034 . Архивировано (PDF) из оригинала 10 января 2011 года . Проверено 16 апреля 2010 г.
- ^ Гирибет, Г.; Рихтер, С.; Эджкомб, Джорджия; Уиллер, WC (2005). Положение ракообразных внутри членистоногих - данные девяти молекулярных локусов и морфологии (PDF) . Проблемы ракообразных. Том. 16. С. 307–352. дои : 10.1201/9781420037548.ch13 . ISBN 978-0-8493-3498-6 . Архивировано (PDF) из оригинала 16 сентября 2006 г. Проверено 23 августа 2006 г.
- ^ Лосано-Фернандес, Хесус; Джакомелли, Маттиа; Флеминг, Джеймс Ф.; Чен, Альберт; Винтер, Якоб; Томсен, Филип Фрэнсис; Гленнер, Хенрик; Палеро, Ферран; Легг, Дэвид А.; Илифф, Томас М.; Пизани, Давиде; Олесен, Йорген (2019). «Эволюция панкрустообразных, освещенная богатыми таксонами наборами данных геномного масштаба с расширенной выборкой реминожек» . Геном Биол. Эвол . 11 (8): 2055–2070. дои : 10.1093/gbe/evz097 . ПМК 6684935 . ПМИД 31270537 .
- ^ Гирибет, Гонсало; Эджкомб, Грегори (июнь 2019 г.). «Филогения и эволюционная история членистоногих» . Современная биология . 29 (12): Р592–Р602. Бибкод : 2019CBio...29.R592G . дои : 10.1016/j.cub.2019.04.057 . ПМИД 31211983 . S2CID 189926344 .
- ^ Мисоф, Бернхард; и др. (2014). «Филогеномика определяет время и характер эволюции насекомых». Наука . 346 (6210): 763–767. Бибкод : 2014Sci...346..763M . дои : 10.1126/science.1257570 . ПМИД 25378627 . S2CID 36008925 .
- ^ Викинс, Дж. Ф. и Ли, Д. О'К. (2002). Разведение ракообразных: разведение и культура (2-е изд.). Блэквелл. ISBN 978-0-632-05464-0 . Архивировано из оригинала 6 декабря 2008 года . Проверено 3 октября 2008 г.
- ^ Бэйли С., Bugfood II: Насекомые как еда!?! , Факультет энтомологии Университета Кентукки, заархивировано из оригинала 16 декабря 2008 г. , получено 3 октября 2008 г.
- ^ Унгер, Л., Bugfood III: Закуски из насекомых со всего мира , факультет энтомологии Университета Кентукки, заархивировано из оригинала 10 октября 2008 г. , получено 3 октября 2008 г.
- ^ Ригби, Р. (21 сентября 2002 г.), «Tuck in a Tarantula» , Sunday Telegraph , заархивировано из оригинала 18 июля 2009 г. , получено 24 августа 2009 г.
- ^ «Женщины-пауки подают жуткую икру Камбоджи» , ABC News Online , 2 сентября 2002 г., заархивировано из оригинала 3 июня 2008 г. , получено 24 августа 2009 г.
- ^ Рэй, Н. (2002). Одинокая планета Камбоджа . Публикации Lonely Planet. п. 308. ИСБН 978-1-74059-111-9 .
- ^ Вейл, К. (2006), Fierce Food , Plume, ISBN 978-0-452-28700-6 , архивировано из оригинала 11 мая 2011 года , получено 3 октября 2008 года.
- ^ Тейлор, Р.Л. (1975), Бабочки в моем желудке (или: Насекомые в питании человека) , Издательство Woodbridge Press Publishing Company, Санта-Барбара, Калифорния
- ^ Комиссия Кодекса по гигиене пищевых продуктов (1985 г.), «Стандарт Кодекса 152 от 1985 г. (на «пшеничную муку»)» (PDF) , Codex Alimentarius , Продовольственная и сельскохозяйственная организация , заархивировано (PDF) из оригинала 31 декабря 2010 г. , получено 8 мая 2010 .
- ^ «Полный список официальных стандартов» , Codex Alimentarius , Продовольственная и сельскохозяйственная организация, заархивировано из оригинала 31 января 2010 г. , получено 8 мая 2010 г.
- ^ Уровни действий по устранению дефектов пищевых продуктов , Управление по санитарному надзору за качеством пищевых продуктов и медикаментов США , заархивировано из оригинала 18 декабря 2006 г. , получено 16 декабря 2006 г.
- ^ Паолетти, М.Г. (2005), Экологические последствия мелкого домашнего скота: потенциал насекомых, грызунов, лягушек и улиток , Science Publishers , с. 648, ISBN 978-1-57808-339-8
- ^ Галлай, Н.; Саллес, Ж.-М.; Сеттеле, Дж.; Васьер, Бельгия (август 2008 г.). «Экономическая оценка уязвимости мирового сельского хозяйства перед лицом сокращения количества опылителей» (PDF) . Экологическая экономика . 68 (3): 810–821. дои : 10.1016/j.ecolecon.2008.06.014 . S2CID 54818498 . Архивировано (PDF) из оригинала 27 апреля 2019 г. Проверено 24 ноября 2018 г. Бесплатное резюме на Галлай, Н.; Саллес, Дж.; Сеттеле, Дж.; Васьер, Б. (2009). «Экономическая ценность опыления насекомыми во всем мире оценивается в 153 миллиарда евро» . Экологическая экономика . 68 (3): 810–821. дои : 10.1016/j.ecolecon.2008.06.014 . S2CID 54818498 . Архивировано из оригинала 3 декабря 2008 года . Проверено 3 октября 2008 г.
- ^ Apiservices — Международный рынок меда — Мировое производство, импорт и экспорт меда , архивировано из оригинала 6 декабря 2008 г. , получено 3 октября 2008 г.
- ^ Временная шкала тканей , Threads In Tyme, LTD, заархивировано из оригинала 28 октября 2005 г. , получено 14 июля 2005 г.
- ^ Джефф Бехан, Ошибка, изменившая историю , заархивировано из оригинала 21 июня 2006 г. , получено 26 июня 2006 г.
- ^ Домашняя страница производителей кошенили на Канарских островах , заархивировано из оригинала 24 июня 2005 г. , получено 14 июля 2005 г.
- ^ Херд, В., Кост (PDF) , Университет Южной Флориды , заархивировано из оригинала (PDF) 19 февраля 2017 г. , получено 25 августа 2008 г.
- ^ Холл, РД; Кастнер, Дж. Л. (2000), «Введение» , в Берде, Дж. Х.; Кастнер, Дж.Л. (ред.), Судебная энтомология: полезность членистоногих в юридических расследованиях , CRC Press , стр. 3–4 , ISBN 978-0-8493-8120-1
- ^ Досси, Аарон (декабрь 2010 г.). «Насекомые и их химическое оружие: новый потенциал для открытия лекарств». Отчеты о натуральных продуктах . 27 (12): 1737–1757. дои : 10.1039/C005319H . ПМИД 20957283 .
- ^ Спанья, JC; Гольдман Д.И.; Лин П.-К.; Кодичек Д.Э.; Р.Дж. Фулл (март 2007 г.). «Распределенная механическая обратная связь у членистоногих и роботов упрощает управление быстрым бегом по сложной местности» (PDF) . Биоинспирация и биомиметика . 2 (1): 9–18. Бибкод : 2007БиБи....2....9S . дои : 10.1088/1748-3182/2/1/002 . ПМИД 17671322 . S2CID 21564918 . Архивировано из оригинала (PDF) 10 марта 2012 года.
- ^ Кадзуо Цучия; Шинья Аой и Кацуёси Цудзита (2006), «Стратегия поворота многоногого передвижного робота», Адаптивное движение животных и машин , стр. 227–236, CiteSeerX 10.1.1.573.1846 , doi : 10.1007/4-431- 31381-8_20 , ISBN 978-4-431-24164-5
- ↑ Перейти обратно: Перейти обратно: а б Хилл, Д. (1997), Экономическое значение насекомых , Springer, стр. 77–92, ISBN. 978-0-412-49800-8
- ^ Гудман, Джесси Л.; Деннис, Дэвид Тэппен; Соненшайн, Дэниел Э. (2005), Клещевые болезни человека , ASM Press , стр. 114, ISBN 978-1-55581-238-6 , получено 29 марта 2010 г.
- ^ Поттер, М.Ф., Паразитические клещи человека , Сельскохозяйственный колледж Университета Кентукки, заархивировано из оригинала 8 января 2009 г. , получено 25 октября 2008 г.
- ^ Кленерман, Пол; Липуорт, Брайан, Аллергия на клещей домашней пыли , NetDoctor, архивировано из оригинала 11 февраля 2008 г. , получено 20 февраля 2008 г.
- ↑ Перейти обратно: Перейти обратно: а б Коган, М.; Крофт, бакалавр; Сазерст, РФ (1999), «Применение экологии для комплексной борьбы с вредителями» , в Хаффейкере, Карле Б.; Гутьеррес, AP (ред.), Экологическая энтомология , John Wiley & Sons, стр. 681–736, ISBN 978-0-471-24483-7
- ^ Горхэм, Дж. Ричард (1991), «Насекомые и клещи-вредители в продуктах питания: иллюстрированный ключ» (PDF) , Справочник по сельскому хозяйству, номер 655 , Министерство сельского хозяйства США , стр. 1–767, заархивировано из оригинала (PDF) на 25 октября 2007 г. , дата обращения 6 мая 2010 г.
- ^ Чон, Д.Д.; Морс, Р.А. и Эйкворт, GC (январь 1982 г.). «Клещи-вредители медоносных пчел». Ежегодный обзор энтомологии . 27 : 229–252. дои : 10.1146/annurev.en.27.010182.001305 .
- ^ Меткалф, Роберт Ли; Лакманн, Уильям Генри (1994), Введение в борьбу с насекомыми-вредителями , Wiley-IEEE, стр. 4, ISBN 978-0-471-58957-0
- ^ Шульц, JW (2001), «Chelicerata (паукообразные, включая пауков, клещей и скорпионов)», Энциклопедия наук о жизни , John Wiley & Sons, Ltd., doi : 10.1038/npg.els.0001605 , ISBN 978-0-470-01617-6
- ^ Осакабе, М. (март 2002 г.), «Какой хищный клещ может контролировать как доминирующего клеща-вредителя Tetranychus urticae , так и латентного клеща-вредителя Eotetranychus asiaticus на клубнике?», Experimental and Applied Acarology , 26 (3–4): 219–230, doi : 10.1023/A:1021116121604 , PMID 12542009 , S2CID 10823576
Библиография [ править ]
- Гулд, С.Дж. (1990). Прекрасная жизнь: сланцы Бёрджесс и природа истории . Хатчинсон Радиус. Бибкод : 1989wlbs.book.....G . ISBN 978-0-09-174271-3 .
- Руперт, Э.Э.; Р. С. Фокс; Р. Д. Барнс (2004). Зоология беспозвоночных (7-е изд.). Брукс/Коул . ISBN 978-0-03-025982-1 .
Внешние ссылки [ править ]
- «Членистоногие» в Энциклопедии жизни
- по ядовитым членистоногим Отделение в Агентстве по охране окружающей среды США и Университете Флориды / Институте пищевых и сельскохозяйственных наук, Национальное учебное пособие для специалистов по применению пестицидов в области общественного здравоохранения.
- Членистоногие – членистоногие формы жизни насекомых