Обоняние насекомых
Обоняние насекомых относится к функции химических рецепторов , которые позволяют насекомым обнаруживать и идентифицировать летучие соединения для поиска пищи , предотвращения хищников, поиска для спаривания партнеров (посредством феромонов ) и определения местонахождения мест откладки яиц . [1] Таким образом, это самое важное ощущение для насекомых. [1] Наиболее важные действия насекомых должны быть точно рассчитаны по времени, что зависит от того, что они чувствуют и когда они это чувствуют. [2] Например, обоняние необходимо для обнаружения растений-хозяев и охоты на добычу у многих видов насекомых, таких как моль Deilephila elpenor и оса Polybia sericea соответственно.
Два органа, которые насекомые в основном используют для обнаружения запахов, — это усики и специальные части рта, называемые верхнечелюстными щупиками. [3] Однако недавнее исследование продемонстрировало обонятельную роль яйцеклада инжирных ос. [4] Внутри этих органов обоняния есть нейроны, называемые нейронами обонятельных рецепторов, которые, как следует из названия, содержат рецепторы для молекул запаха в своей клеточной мембране. Большинство нейронов обонятельных рецепторов обычно расположены в антенне . Этих нейронов может быть очень много; например, у мух -дрозофил 2600 обонятельных сенсорных нейронов. [3]
Насекомые способны и избирательно чувствовать запахи и различать тысячи летучих соединений чутко . [1] [5] Чувствительность – это то, насколько насекомое настроено на очень небольшое количество запаха или небольшие изменения концентрации запаха. Селективность означает способность насекомых отличать один запах от другого. Среди кровососущих членистоногих эти соединения обычно делятся на три класса: карбоновые кислоты с короткой цепью , альдегиды и низкомолекулярные азотистые соединения. [5]
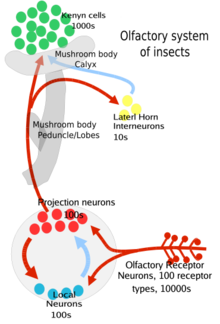
Насекомые использовались в качестве модельной системы млекопитающих и особенно человека для изучения обоняния . Тем не менее, в отличие от позвоночных, которые используют рецепторы, связанные с G-белком ( GPCR ), насекомые экспрессируют белки, включая OR ( обонятельные рецепторы ), GR (вкусовые рецепторы) и IR ( ионотропные рецепторы ), которые все являются гетеромерными лиганд-управляемыми ионными каналами . [3] Вид бабочки отряда чешуекрылых, известный как черная совка ( Agrotis ipsilon ), производит еще больше белков, включая OBP ( белки, связывающие запахи ), CSP (хемосенсорные связывающие белки) и SNMP (белки мембран сенсорных нейронов), которые помогают моли распознавать половые феромоны и пахучие вещества, например те, которые выделяются растениями-хозяевами. [6] Как и у позвоночных, аксоны сенсорных нейронов сходятся в клубочки, но различаются местом расположения клубочков. У млекопитающих они расположены в обонятельных луковицах , а у насекомых — в усиковой доле . [7]
Эволюционная история
[ редактировать ]Обоняние требует метаболических затрат. Эволюционные компромиссы требуют дальнейшего изучения, поскольку по состоянию на 2016 г. [update] большинство таких исследований проводилось в лабораторных условиях с использованием нереально надежных продуктов питания. [8]
Обонятельный каскад
[ редактировать ]Сенсорные нейроны антенны, верхнечелюстных щупиков и лабелл генерируют специфические для запаха электрические сигналы, называемые спайками ( потенциалами действия ), в ответ на связывание запахов с белками клеточной поверхности, такими как обонятельные рецепторы . Сенсорные нейроны антенны и верхней челюсти посылают эту информацию через свои аксоны в долю усика . [7] в то время как сенсорный нейрон лабеллы отправляет эту информацию через аксоны в подпищеводный ганглий . [9] Внутри антеннальной доли они образуют синапсы с другими нейронами в полуочерченных (с мембранными границами) структурах, называемых клубочками .
В частности, процесс выглядит следующим образом: сначала аромат доносится до антенны насекомого или верхнечелюстных щупиков , которые покрыты волосообразными выступами, называемыми сенсиллами . [5] Затем запах проникает через крошечные поры в экзоскелете (или кутикуле) этой сенсиллы и диффундирует в жидкость между клетками, называемую внеклеточной жидкостью . [1] Там молекула одоранта связывается с белком, связывающим одорант , который транспортирует его к рецептору. [1] и команда корецепторов (Orco) на поверхности нейрона обонятельного рецептора (ORN). [1] [3] Это приводит к тому, что нейрон запускает потенциал действия по аксону. [2] Этот сигнал посылается в антеннальную долю или подпищеводный ганглий мозга насекомых, где он затем может интегрировать информацию с другими сигналами от других сенсилл .
Эти ОРН являются биполярными: на одном конце находятся обонятельные дендриты с рецепторами запахов, а на другом конце — аксоны, которые передают потенциал действия в антеннальную долю мозга. [3] Усиковые доли имеют два типа нейронов: проекционные нейроны (в основном возбуждающие) и локальные нейроны (тормозные, с некоторыми возбуждающими). Проекционные нейроны посылают свои аксонные окончания в часть мозга насекомых, называемую грибовидными телами (важная для регуляции усвоенных реакций на запахи), и в другую часть мозга, называемую боковым рогом (важная для регуляции врожденных реакций на запахи). [3] ). Обе эти области являются частью протоцеребрума мозга насекомых.
Методы исследования
[ редактировать ]Запись потенциала действия проводится тремя различными способами: электроантенограммами, электропальпограммами и записью одиночных сенсилл (SSR). [5] На электроантенограммах (ЭАГ) и электропальпограммах (ЭПГ) регистрируются потенциалы действия всей антенны или верхнечелюстного щупика соответственно. EAG и EPG дают общее представление об обонянии соответствующего органа. [5] Во время SSR электрод вводится только в одну сенсиллу, и запись производится только из ORN, содержащихся внутри этой сенсиллы, предоставляя более подробную информацию. [5] .
Любой из этих методов можно комбинировать с газовой хроматографией высокого разрешения для выделения летучих соединений из важных животных или мест обитания. [5] Например, этот метод можно использовать для определения того, какое соединение из конкретного цветка является наиболее привлекательным для пчел. Записи проекционных нейронов показывают, что у некоторых насекомых существует сильная специализация и распознавание запахов, представляемых ОРН. Особенно это касается проекционных нейронов макрогломерул — специализированного комплекса клубочков, ответственного за обнаружение феромонов.
Репелленты и аттрактанты
[ редактировать ]Люди используют обонятельную систему насекомых для борьбы с сельскохозяйственными вредителями и переносчиками болезней. [3] Для некоторых сельскохозяйственных вредителей искусственные половые феромоны помещаются в ловушки для отлова взрослых особей до того, как они смогут отложить яйца (отложить яйца), что приводит к вылуплению их разрушительных личинок. [3] Хотя существуют тысячи химических веществ, которые насекомые могут обнаружить, существует ограниченный диапазон, который насекомые используют в качестве сигналов, чтобы двигаться к источнику запаха или от него. [5]
Искусство поиска аттрактанта или репеллента для конкретного интересующего насекомого — сложный и долгий и интенсивный процесс. Например, использование феромонов привлекает насекомых только на репродуктивной стадии, коротком периоде их жизни. [2] Хотя запахи еды могут быть привлекательными для голодных насекомых, они не будут эффективны на поле, полном урожая, который нравится этим насекомым. [2]
Ситуативно-зависимые аттрактанты/репелленты
[ редактировать ]Насекомые используют один и тот же сигнал для самых разных целей в зависимости от ситуации. Это называется химической экономией. [5] Ситуациями, которые могут изменить поведение насекомого в ответ на запах, являются такие вещи, как концентрация соединения, жизненный этап насекомого, его статус спаривания, другие обонятельные сигналы, состояние питания насекомых (голодное или сытое), время день или даже положение тела насекомых. [2] [3] [5] Например, дрозофилу очень привлекает яблочный уксус, но в очень высоких концентрациях активируется дополнительный обонятельный рецептор (имеющий низкое сродство к уксусу, Or85a), который меняет поведение мухи с влечения на отвращение. [3] Такое разное поведение по одному и тому же сигналу называется поведенческой пластичностью. [2]
Углекислый газ
[ редактировать ]Многие насекомые способны улавливать малейшие изменения концентрации CO 2 . [5] Хотя было обнаружено, что CO 2 является аттрактантом у каждого исследованного членистоногого. [5] и это очень важно для мониторинга и борьбы с комарами, даже эта стереотипная реакция может быть пластичной. Дрозофила избегает CO 2 при ходьбе, но движется к нему в полете. [3]
ДЭТА
[ редактировать ]Было показано, что многие насекомые (и другие членистоногие) избегают мест, содержащих N,N-диэтил-3-метилбензамид или ДЭТА . Они изначально избегают ДЭТА, вероятно, потому, что это «конфузант», который стимулирует вкусовые, ионотропные и обонятельные рецепторы и «искажает» взаимодействие других одорантов с этими рецепторами. [3]
См. также
[ редактировать ]- Электроантеннография , Электроольфактография
- Машинное обоняние , Электронный нос
- Билл С. Хэнсон
- Запись одиночной сенсиллы
Ссылки
[ редактировать ]- ^ Jump up to: а б с д и ж Каррахер, Колм; Далзил, Джули; Джордан, Мелисса Д.; Кристи, Дэвид Л.; Ньюкомб, Ричард Д.; Краличек, Эндрю В. (2015). «К пониманию структурной основы обоняния насекомых рецепторами запаха» . Биохимия насекомых и молекулярная биология . 66 : 31–41. Бибкод : 2015IBMB...66...31C . дои : 10.1016/j.ibmb.2015.09.010 . ПМИД 26416146 .
- ^ Jump up to: а б с д и ж Гаденн, Кристоф; Баррозо, Ромина Б.; Антон, Сильвия (2016). «Пластичность обоняния насекомых: пахнуть или не пахнуть?». Ежегодный обзор энтомологии . 61 : 317–333. doi : 10.1146/annurev-ento-010715-023523 . hdl : 11336/19586 . ПМИД 26982441 .
- ^ Jump up to: а б с д и ж г час я дж к л Ли, Цянь; Либерлес, Стивен Д. (2015). «Отвращение и влечение через обоняние» . Современная биология . 25 (3): Р120–Р1209. Бибкод : 2015CBio...25.R120L . дои : 10.1016/j.cub.2014.11.044 . ПМЦ 4317791 . ПМИД 25649823 .
- ^ Ядав, Пратибха; Борхес, Рене М. (2017). «Яйцеклад насекомого как изменчивый сенсор внутри замкнутого микромира» . Журнал экспериментальной биологии . 220 (9): 1554–1557. дои : 10.1242/jeb.152777 . ПМИД 28468812 .
- ^ Jump up to: а б с д и ж г час я дж к л Сайед, Зайнулабеддин (2015). «Химическая экология и обоняние членистоногих-переносчиков болезней». Современное мнение в области науки о насекомых . 10 : 83–89. Бибкод : 2015COIS...10...83S . дои : 10.1016/j.cois.2015.04.011 . ПМИД 29588018 .
- ^ Гу, Шао-Хуа; и др. (2014). «Молекулярная характеристика и дифференциальная экспрессия обонятельных генов в усиках черной совки Agrotis ipsilon » . ПЛОС ОДИН . 9 (8): е103420. Бибкод : 2014PLoSO...9j3420G . дои : 10.1371/journal.pone.0103420 . ПМЦ 4118888 . ПМИД 25083706 .
- ^ Jump up to: а б Уилсон, Рэйчел (2013). «Ранняя обонятельная обработка у дрозофилы: механизмы и принципы» . Ежегодный обзор неврологии . 36 : 217–241. doi : 10.1146/annurev-neuro-062111-150533 . ПМЦ 3933953 . ПМИД 23841839 .
- ^ Гаденн, Кристоф; Баррозо, Ромина Б.; Антон, Сильвия (11 марта 2016 г.). «Пластичность обоняния насекомых: пахнуть или не пахнуть?». Ежегодный обзор энтомологии . 61 (1). Годовые обзоры : 317–333. doi : 10.1146/annurev-ento-010715-023523 . hdl : 11336/19586 . ISSN 0066-4170 . ПМИД 26982441 . S2CID 207568844 .
- ^ Рябинина, Елена; Задача, Дарья; Марр, Элизабет; Лин, Чун-Чье; Алфорд, Роберт; О'Брочта, Дэвид А.; Поттер, Кристофер Дж. (3 октября 2016 г.). «Организация обонятельных центров у малярийного комара Anopheles gambiae» . Природные коммуникации . 7 : 13010. Бибкод : 2016NatCo...713010R . дои : 10.1038/ncomms13010 . ПМК 5063964 . ПМИД 27694947 .