Грибные тела

Грибовидные тела или corpora pedunculata — парные структуры головного мозга членистоногих числе , в том насекомых и ракообразных . [2] и некоторые кольчатые черви (особенно тряпичный червь Platynereis dumerilii ). [3] Известно, что они играют роль в обонятельном обучении и памяти . У большинства насекомых грибовидные тела и боковой рог являются двумя высшими областями мозга, которые получают обонятельную информацию от усиков через проекционные нейроны. [4] Впервые они были идентифицированы и описаны французским биологом Феликсом Дюжарденом в 1850 году. [5] [6]
Структура
[ редактировать ]Грибовидные тела обычно описываются как нейропили , т. е. как плотные сети нейрональных отростков ( дендриты и окончания аксонов ) и глии . Они получили свое название из-за своей примерно полусферической чашечки , выступа, который соединен с остальной частью мозга центральным нервным трактом или ножкой .
Большая часть наших текущих знаний о грибных телах получена в результате исследований нескольких видов насекомых, особенно таракана Periplaneta americana , медоносной пчелы Apis mellifera , [7] саранча и плодовая мушка Drosophila melanogaster . Исследования тел грибов плодовой мухи были особенно важны для понимания генетической основы функционирования грибных тел, поскольку их геном секвенирован и существует огромное количество инструментов для управления экспрессией их генов.
В мозгу насекомых ножки грибовидных тел проходят через средний мозг . Они в основном состоят из длинных, плотно упакованных нервных волокон клеток Кеньона , собственных нейронов грибовидных тел. Эти клетки были обнаружены в телах грибов всех исследованных видов, хотя их количество варьируется. Например, у плодовых мух их около 2500, а у тараканов — около 200 000.
Здесь можно увидеть вскрытие мозга саранчи с целью обнажения центрального мозга и проведения электрофизиологических записей. [8]
Эволюционная история
[ редактировать ]Исторически считалось, что грибовидные тела есть только у насекомых, поскольку у крабов и омаров их нет. Однако их открытие у креветок-богомолов в 2017 году привело к более позднему выводу. [2] что грибовидное тело является предковым состоянием всех членистоногих и что эта особенность позже была утеряна у крабов и омаров. [2]
Функция
[ редактировать ]Грибные тела наиболее известны своей ролью в обонятельном ассоциативном обучении. Эти обонятельные сигналы поступают от дофаминергических , октопаминэргических , холинергических , серотонинергических и ГАМКергических нейронов за пределами МБ. [9] Они самые крупные среди перепончатокрылых , которые, как известно, особенно тщательно контролируют обонятельное поведение. Однако, поскольку грибовидные тела обнаруживаются и у аносмичных примитивных насекомых, их роль, вероятно, выходит за рамки обонятельной обработки. Анатомические исследования предполагают определенную роль в обработке зрительной и механосенсорной информации у некоторых видов. [10] В частности, у перепончатокрылых субрегионы нейропиля грибовидного тела специализированы для получения обонятельной, зрительной или обоих типов сенсорной информации. [11] У перепончатокрылых обонятельный вход расположен в чашечке. У муравьев можно выделить несколько слоев, соответствующих разным скоплениям клубочков в долях усиков , возможно, для обработки разных классов запахов. [4] [12] Существуют две основные группы проекционных нейронов, разделяющих долю усика на две основные области: переднюю и заднюю. Группы проекционных нейронов разделены, иннервируют группы клубочков отдельно и посылают аксоны разными путями, либо через медиально-антенный протоцеребральный тракт (m-APT), либо через латеральный антенно протоцеребральный тракт (l-APT), и соединяясь с двумя слоями в чашечка грибовидных тел. В этих слоях топографически представлена организация двух эфферентных областей усика, что позволяет составить приблизительную одотопическую карту усика в области губ грибовидных тел. [4] [12]
Известно, что грибные тела участвуют в обучении и памяти , особенно в отношении обоняния , и поэтому являются предметом текущих интенсивных исследований. Исследования показывают, что у более крупных насекомых грибовидные тела обладают и другими функциями обучения и памяти, такими как ассоциативная память , сенсорная фильтрация, двигательный контроль и память места. Исследования показывают, что грибовидные тела обычно действуют как своего рода детектор совпадений , интегрируя мультимодальные входные данные. [4] и создают новые ассоциации, тем самым предполагая их роль в обучении и памяти. [13] Недавняя работа также демонстрирует доказательства участия грибовидного тела во врожденном обонятельном поведении посредством взаимодействия с боковым рогом. [14] [15] возможно, используя частично стереотипные сенсорные реакции выходных нейронов грибовидного тела (MBON) у разных людей. [16] Хотя связи между проекционными нейронами и клетками Кеньона случайны (т. е. не стереотипны для разных людей), [17] Стереотипность в ответах MBON стала возможной благодаря плотной конвергенции многих ячеек Кеньона на несколько MBON, а также других свойств сети. [16]
Информация о запахах может быть закодирована в теле гриба с помощью особенностей чувствительных нейронов, а также времени их импульсов. [18] Эксперименты на саранче показали, что клеток Кеньона активность синхронизируется с нейронными колебаниями частотой 20 Гц и особенно чувствительна к спайкам проекционных нейронов на определенных фазах колебательного цикла. [19]
Спать
[ редактировать ]Нейроны, которые получают сигналы от серотонинергических и ГАМКергических нейронов за пределами МБ, вызывают бодрствование, а экспериментальная стимуляция этих вышестоящих серотонинергических нейронов вызывает сон. Нейроны-мишени в MB ингибируются серотонином , ГАМК и их комбинацией. С другой стороны, октопамин , похоже, не влияет на функцию сна МБ. [9]
Дрозофила меланогастер
[ редактировать ]
Мы знаем, что грибовидные структуры тела важны для обонятельного обучения и памяти у дрозофилы, поскольку их абляция разрушает эту функцию. [21] Грибовидное тело также способно комбинировать информацию о внутреннем состоянии тела и обонятельные данные для определения врожденного поведения. [22] Точная роль конкретных нейронов, составляющих грибовидные тела, до сих пор неясна. Однако эти структуры широко изучаются, поскольку многое известно об их генетической структуре . Существует три конкретных класса нейронов, составляющих доли грибовидного тела: α/β, α’/β’ и γ-нейроны, все из которых имеют различную экспрессию генов. Темой текущих исследований является то, какие из этих подструктур грибовидного тела участвуют в каждой фазе и процессе обучения и памяти. [23] Грибные тела дрозофилы также часто используются для изучения обучения и памяти, и ими манипулируют из-за их относительно дискретной природы. Обычно анализы обонятельного обучения состоят из подвергания мух воздействию двух запахов по отдельности; один сопряжен с электрошоковыми импульсами ( условный раздражитель , или УС+), а второй нет ( безусловный раздражитель , или УЗ). После этого периода обучения мух помещают в Т-образный лабиринт , причем два запаха размещаются индивидуально на обоих концах горизонтальных Т-образных рычагов. Подсчитывается процент мух, избегающих CS+, при этом высокий уровень избегания считается свидетельством обучения и памяти. [24]
Следы клеточной памяти
[ редактировать ]Недавние исследования, сочетающие кондиционирование запаха и клеточную визуализацию, выявили шесть следов памяти, которые совпадают с молекулярными изменениями в дрозофилы обонятельной системе . Три из этих следов связаны с ранним формированием поведенческой памяти. Один из таких следов был визуализирован в антеннальной доле (AL) с помощью репортерных молекул синапто-флуорина . Сразу после кондиционирования дополнительный набор проекционных нейронов в наборе из восьми клубочков AL синаптически активируется условным запахом и длится всего 7 минут. [25] Второй след обнаруживается по экспрессии GCaMP и, следовательно, по увеличению Ca 2+ приток в α'/β'-аксоны нейронов грибовидного тела. [26] Это более продолжительный след, который сохраняется до одного часа после кондиционирования. Третий след памяти — снижение активности переднепарного латерального нейрона, который действует как супрессор формирования памяти через один из своих тормозных ГАМКергических рецепторов. Снижение кальциевого ответа нейронов APL и последующее снижение высвобождения ГАМК на грибовидные тела сохранялось до 5 минут после кондиционирования запаха. [27]
След промежуточной памяти зависит от экспрессии гена amn , расположенного в дорсальных парных медиальных нейронах. Увеличение притока кальция и синаптического высвобождения, иннервирующего грибовидные тела, становится заметным примерно через 30 минут после сочетания электрошока с запахом и сохраняется не менее часа. [28] Оба картированных следа долговременной памяти зависят от активности и синтеза белков CREB и CaMKII и существуют только после интервального кондиционирования. Первый след обнаруживается в α/β-нейронах между 9 и 24 часами после кондиционирования и характеризуется увеличением притока кальция в ответ на условный запах. [29] Второй след долговременной памяти формируется в телах γ-гриба и обнаруживается по увеличению притока кальция между 18 и 24 часами после кондиционирования. [30]
динамика цАМФ
[ редактировать ]Циклический аденозинмонофосфат (цАМФ или циклический АМФ) является вторичным мессенджером, который участвует в облегчении притока кальция в грибовидные тела в нейроны грибовидных тел Drosophila melanogaster . Повышение цАМФ индуцирует пресинаптическую пластичность у дрозофилы. На уровень цАМФ влияют как нейротрансмиттеры , такие как дофамин и октопамин , так и сами запахи. Дофамин и октопамин высвобождаются интернейронами грибовидных тел , а запахи напрямую активируют нейроны обонятельного пути, вызывая приток кальция через потенциалзависимые кальциевые каналы . [31]
В классической парадигме кондиционирования нейронов сочетание деполяризации (посредством применения ацетилхолина для обозначения запаха или CS ) с последующим применением дофамина (для обозначения шока или УЗИ ) приводит к синергетическому увеличению цАМФ в долях грибовидного тела. [31] Эти результаты позволяют предположить, что доли грибовидных тел являются критическим местом интеграции CS/US посредством действия цАМФ. Этот синергетический эффект первоначально наблюдался у Aplysia , где сочетание притока кальция с активацией G-белка передачи сигналов серотонином приводит к аналогичному синергическому увеличению цАМФ. [32]
Кроме того, это синергическое увеличение цАМФ опосредовано и зависит от аденилатциклазы брюквы (рут AC), которая чувствительна как к кальцию (что является результатом открытия потенциалзависимых кальциевых каналов запахами), так и к стимуляции G-белка (вызванной дофамином). [31] В то время как прямое сочетание деполяризации нейронов и дофамина (ацетилхолин с последующим дофамином) приводит к синергическому увеличению цАМФ, прямое сочетание деполяризации нейронов и октопамина оказывает субаддитивный эффект на цАМФ. [31] Более конкретно, это означает, что эта пара производит значительно меньше цАМФ, чем сумма каждого стимула по отдельности в долях. Следовательно, коловый AC в нейронах грибовидных тел работает как детектор совпадений , при этом дофамин и октопамин действуют двунаправленно, влияя на уровни цАМФ. [31]
Динамика ПКА
[ редактировать ]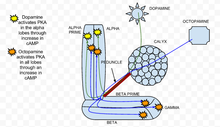
протеинкиназа А Было обнаружено, что (ПКА) играет важную роль в обучении и памяти у дрозофилы . [33] Когда кальций попадает в клетку и связывается с кальмодулином , он стимулирует аденилатциклазу (АС), которая кодируется геном брюквы ( рут ). [34] Эта активация AC увеличивает концентрацию цАМФ , который активирует PKA. [34] Когда применяется дофамин , аверсивный обонятельный стимулятор, он активирует ПКА конкретно в вертикальных долях грибовидного тела. [34] Эта пространственная специфичность регулируется dunce ( dnc ) PDE, цАМФ-специфической фосфодиэстеразой . Если ген dunce отменен, как это обнаружено у мутанта dnc , пространственная специфичность не сохраняется. Напротив, стимуляция аппетита, вызванная применением октопамина , увеличивает ПКА во всех долях. [34] У гонного мутанта, генотипа, в котором брюква отсутствует, реакции как на дофамин, так и на октопамин были значительно снижены и близки к экспериментальному шуму.
Ацетилхолин , который представляет собой условный раздражитель , приводит к сильному увеличению активации ПКА по сравнению со стимуляцией только дофамином или октопамином. [34] Эта реакция отменена у гонных мутантов, что демонстрирует, что PKA необходима для сенсорной интеграции. [34] Специфичность активации альфа-доли в присутствии дофамина сохраняется при сочетании дофамина с ацетилхолином. [34] По сути, во время парадигмы кондиционирования, когда условный стимул сочетается с безусловным стимулом, ПКА демонстрирует повышенную активацию. Это показывает, что PKA необходим для условного обучения у Drosophila melanogaster .
Апис меллифера
[ редактировать ]Стимул → выходные реакции являются продуктом пар возбуждения и торможения . Это та же модель организации, что и в мозге млекопитающих . Эти закономерности могут, как и у млекопитающих, предшествовать действию. По состоянию на 2021 год [update] эта область лишь недавно была освещена Звакой и др., 2018 г., Дуером и др., 2015 г. и Паффхаузеном и др., 2020 г. [7]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Дженетт А.; Шинделин Дж. Э.; Гейзенберг М. (2006). «Протокол виртуального мозга насекомых: создание и сравнение стандартизированной нейроанатомии» . БМК Биоинформатика . 7 : 544. дои : 10.1186/1471-2105-7-544 . ПМК 1769402 . ПМИД 17196102 .
- ^ Jump up to: а б с Штраусфельд, Николас Джеймс; Вольф, Габриэлла Ханна; Сэйр, Марсель Итан (3 марта 2020 г.). «Эволюция грибовидных тел демонстрирует гомологию и расхождение между Pancrustacea» . электронная жизнь . 9 . дои : 10.7554/eLife.52411 . ISSN 2050-084X . ПМК 7054004 . ПМИД 32124731 .
- ^ Томер, Р.; Денес, А.С.; Тессмар-Райбл, К.; Арендт, Д. (2010). «Профилирование путем регистрации изображений показывает общее происхождение грибовидных тел кольчатых червей и мантия позвоночных» . Клетка . 142 (5): 800–809. дои : 10.1016/j.cell.2010.07.043 . ПМИД 20813265 . S2CID 917306 .
- ^ Jump up to: а б с д Гроненберг, В.; Лопес-Рикельме, GO (февраль 2014 г.). «Мультисенсорная конвергенция в грибовидных телах муравьев и пчел». Акта Биологика Хунгарика . 55 (1–4): 31–37. дои : 10.1556/ABiol.55.2004.1-4.5 . ПМИД 15270216 .
- ^ Дюжарден, Ф. (1850). «Память о нервной системе насекомых». Энн. наук. Нат. Зоол . 14 : 195–206.
- ^ Штраусфельд, штат Нью-Джерси; Хансен Л; Ли Й; Гомес Р.С.; Ито К. (1998). «Эволюция, открытие и интерпретация грибовидных тел членистоногих» . Учиться. Память 5 (1–2): 11–37. дои : 10.1101/lm.5.1.11 . ПМК 311242 . ПМИД 10454370 .
- ^ Jump up to: а б Мензель, Рэндольф (13 августа 2020 г.). «Краткая история исследований интеллекта и мозга медоносных пчел» . Апидология . 52 (1). IRAE & DI ( Спрингер ): 23–34. дои : 10.1007/s13592-020-00794-x . ISSN 0044-8435 . S2CID 221111734 .
- ^ «Вскрытие мозга насекомых для электрофизиологии in vivo» . Ютуб .
- ^ Jump up to: а б Хелфрих-Фёрстер, Шарлотта (07 января 2018 г.). «Сон среди насекомых» . Ежегодный обзор энтомологии . 63 (1). Годовые обзоры : 69–86. doi : 10.1146/annurev-ento-020117-043201 . ISSN 0066-4170 . ПМИД 28938081 .
- ^ Зарс, Трой (декабрь 2000 г.). «Поведенческие функции тел насекомых-грибов». Курр Опин Нейробиол . 10 (6): 790–5. дои : 10.1016/S0959-4388(00)00147-1 . ПМИД 11240291 . S2CID 5946392 .
- ^ Моббс, PG (1982). «Мозг медоносной пчелы Apis Mellifera. I. Связи и пространственная организация тел грибов». Философские труды Лондонского королевского общества Б. 298 (1091): 309–354. Бибкод : 1982РСТБ.298..309М . дои : 10.1098/rstb.1982.0086 .
- ^ Jump up to: а б Лопес-Рикельме, GO (июнь 2014 г.). «Одотопическое афферентное представление клубочковой организации усиков в грибовидных телах муравьев (Hymenoptera: Formicidae): Сравнение двух видов» . TIP Revista Especializada en Ciencias Químico-Biológicas . 15 (1): 15–31. дои : 10.1016/S1405-888X(14)70317-1 .
- ^ Талли, Т; Куинн, WG (сентябрь 1985 г.). «Классическое кондиционирование и сохранение у нормальных и мутантных Drosophila melanogaster». J Comp Physiol A. 157 (2): 263–77. дои : 10.1007/bf01350033 . ПМИД 3939242 . S2CID 13552261 .
- ^ Долан, Майкл-Джон; Фрехтер, Шахар; Бейтс, Александр Шакил; Дэн, Чунтао; Хуовиала, Пааво; Робертс, СП Руайри; Шлегель, Филипп; Дхаван, Серен; Табано, Реми; Дионн, Хизер; Христофору, Кристина; Близко, Кари; Сатклифф, Бен; Джулиани, Бьянка; Ли, Фэн; Коста, Марта; Ильке, Гудрун; Мейснер, Джеффри Уилсон; Бок, Дэви Д; Асо, Ёсинори; Рубин, Джеральд М; Джефферис, Грегори SXE (21 мая 2019 г.). «Нейрогенетическое рассечение бокового рога дрозофилы выявляет основные результаты, разнообразные поведенческие функции и взаимодействие с грибовидным телом» . электронная жизнь . 8 : e43079. doi : 10.7554/eLife.43079 . ISSN 2050-084X . ПМК 6529221 . ПМИД 31112130 .
- ^ Льюис, LP; Сиджу, КП; Асо, Ю; Фридрих, АБ; Балтил, Эй Джей; Рубин, генеральный менеджер; Грюнвальд Кадов, IC (31 августа 2015 г.). «Высший мозговой контур для немедленной интеграции противоречивой сенсорной информации у дрозофилы» . Современная биология . 25 (17): 2203–14. дои : 10.1016/j.cub.2015.07.015 . ПМИД 26299514 . S2CID 16276500 .
- ^ Jump up to: а б Миттал, Ааруш Мохит; Гупта, Дикша; Сингх, Амрита; Лин, Эндрю С.; Гупта, Нитин (24 февраля 2020 г.). «Множественные сетевые свойства преодолевают случайные соединения, обеспечивая стереотипные сенсорные реакции» . Природные коммуникации . 11 (1): 1023. Бибкод : 2020NatCo..11.1023M . дои : 10.1038/s41467-020-14836-6 . ПМК 7039968 . ПМИД 32094345 .
- ^ Кэрон, С.Дж.; Рута, В; Эбботт, ЛФ; Аксель, Р. (2 мая 2013 г.). «Случайное сближение обонятельных сигналов в теле гриба дрозофилы» . Природа . 497 (7447): 113–7. Бибкод : 2013Natur.497..113C . дои : 10.1038/nature12063 . ПМК 4148081 . ПМИД 23615618 .
- ^ Гупта, Нитин; Стопфер, Марк (6 октября 2014 г.). «Временный канал для информации в разреженном сенсорном кодировании» . Современная биология . 24 (19): 2247–56. дои : 10.1016/j.cub.2014.08.021 . ПМК 4189991 . ПМИД 25264257 .
- ^ Гупта, Нитин; Сингх, Свикрити Саран; Стопфер, Марк (15 декабря 2016 г.). «Осцилляционные окна интеграции в нейронах» . Природные коммуникации . 7 : 13808. Бибкод : 2016NatCo...713808G . дои : 10.1038/ncomms13808 . ISSN 2041-1723 . ПМК 5171764 . ПМИД 27976720 .
- ^ Дэвис, Рональд (2011). «Следы памяти дрозофилы» . Нейрон . 70 (1): 8–19. дои : 10.1016/j.neuron.2011.03.012 . ПМЦ 3374581 . ПМИД 21482352 .
- ^ Макгуайр, Шон; Ле, Фуонг; Дэвис, Рональд (август 2001 г.). «Роль передачи сигналов тела гриба дрозофилы в обонятельной памяти» . Наука . 17 (293): 1330–33. Бибкод : 2001Sci...293.1330M . дои : 10.1126/science.1062622 . ПМИД 11397912 . S2CID 23489877 .
- ^ Брекер, Л.Б.; Сиджу, КП; Варела, Н.; Асо, Ю.; Чжан, М.; Хейн, И.; Кадоу, ICG (2013). «Важнейшая роль грибовидного тела в контекстно-зависимом избегании CO2 у дрозофилы» . Современная биология . 23 (13): 1228–1234. дои : 10.1016/j.cub.2013.05.029 . ПМИД 23770186 . S2CID 15112681 .
- ^ Йылдызоглу, Тугче; Вайслогель, Ян-Марек; Мохаммед, Фархан; Чан, Эдвин С.-Ю.; Ассам, Присели Н.; Кларидж-Чанг, Адам (08 декабря 2015 г.). «Оценка обработки информации в системе памяти: полезность метааналитических методов для генетики» . ПЛОС Генет . 11 (12): e1005718. дои : 10.1371/journal.pgen.1005718 . ISSN 1553-7404 . ПМЦ 4672901 . ПМИД 26647168 .
- ^ Акалал, Давид-Бенджамин; Уилсон, Кертис; Цзун, Лин; Танака, Нобуаки; Ито, Кей; Дэвис, Рональд (сентябрь 2006 г.). «Роль нейронов грибовидного тела дрозофилы в обонятельном обучении и памяти» . Обучение и память . 13 (1): 659–68. дои : 10.1101/lm.221206 . ПМЦ 1783621 . ПМИД 16980542 .
- ^ Ю, Динхуэй; Пономарев Артем; Дэвис, Рональд (май 2004 г.). «Измененное представление пространственного кода запахов после обонятельной классической обработки; формирование следов памяти путем синаптического рекрутирования» . Нейрон . 42 (3): 437–49. дои : 10.1016/S0896-6273(04)00217-X . ПМИД 15134640 . S2CID 5859632 .
- ^ Ван, Ялинь; Мамира, Акира; Чан, Анн-шин ; Чжун, Йи (апрель 2008 г.). «Изображение следов ранней памяти в теле гриба дрозофилы» . Журнал неврологии . 28 (17): 4368–76. doi : 10.1523/jneurosci.2958-07.2008 . ПМЦ 3413309 . ПМИД 18434515 .
- ^ Сюй, Лю; Дэвис, Рональд (январь 2009 г.). «ГАМКергический передний парный латеральный нейрон подавляет обонятельное обучение» . Природная неврология . 12 (1): 53–59. дои : 10.1038/nn.2235 . ПМК 2680707 . ПМИД 19043409 .
- ^ Ю, Динхуэй; Кин, Алекс (декабрь 2005 г.). «Нейроны DPM дрозофилы образуют отсроченный и отраслевой след памяти после классического обонятельного кондиционирования» . Клетка . 123 (5): 945–57. дои : 10.1016/j.cell.2005.09.037 . ПМИД 16325586 . S2CID 14152868 .
- ^ Ю, Динхуэй; Акалал, Бенджамин-Дэвид (декабрь 2006 г.). «Нейроны грибовидного тела дрозофилы a/b образуют отраслевой след долговременной клеточной памяти после разнесенного обонятельного кондиционирования» . Нейрон . 52 (1): 845–55. дои : 10.1016/j.neuron.2006.10.030 . ПМЦ 1779901 . ПМИД 17145505 .
- ^ Акалал, Бенджамин-Дэвид; Ю, Динхуэй (декабрь 2010 г.). «Поздняя фаза следа долговременной памяти в γ-нейронах грибных тел дрозофилы после классического обонятельного кондиционирования» . Журнал неврологии . 30 (49): 16699–16708. doi : 10.1523/jneurosci.1882-10.2010 . ПМЦ 3380342 . ПМИД 21148009 .
- ^ Jump up to: а б с д и Томчик, Сет; Дэвис, Рональд (ноябрь 2009 г.). «Динамика связанной с обучением передачи сигналов цАМФ и интеграции стимулов в обонятельный путь дрозофилы» . Нейрон . 64 (4): 510–21. дои : 10.1016/j.neuron.2009.09.029 . ПМК 4080329 . ПМИД 19945393 .
- ^ Абрамс, Томас; Карл, Кевин; Кандел, Эрик (сентябрь 1991 г.). «Биохимические исследования конвергенции стимулов во время классического кондиционирования у аплизии: двойная регуляция аденилатциклазы с помощью Ca2+/кальмодулина и медиатора» . Журнал неврологии . 11 (9): 2655–65. doi : 10.1523/JNEUROSCI.11-09-02655.1991 . ПМК 6575265 . ПМИД 1679120 . S2CID 16477962 .
- ^ Скулакис, Э.М.; Кальдерон, Д; Дэвис, Р.Л. (1993). «Преимущественная экспрессия в грибных тельцах каталитической субъединицы протеинкиназы А и ее роль в обучении и памяти». Нейрон . 11 (2): 197–201. дои : 10.1016/0896-6273(93)90178-т . ПМИД 8352940 . S2CID 23105390 .
- ^ Jump up to: а б с д и ж г Джерваси, Николас; Ченио, Пол; Преат, Томас (февраль 2010 г.). «Динамика PKA в учебном центре дрозофилы: обнаружение совпадений с помощью аденилатциклазы брюквы и пространственная регуляция с помощью фосфодиэстеразы Дунса» . Нейрон . 65 (4): 516–529. дои : 10.1016/j.neuron.2010.01.014 . ПМИД 20188656 . S2CID 14318460 .
Дальнейшее чтение
[ редактировать ]- Мартин Гейзенберг (1998). «Что грибные тела делают для мозга насекомых? Введение» . Учиться. Мем . 5 (1): 1–10. дои : 10.1101/lm.5.1.1 . ПМК 311238 . ПМИД 10454369 .
- С. Е. Фарбах (2006). «Строение грибовидных тел мозга насекомых». Анну. Преподобный Энтомол. 51 (3): 209–232. дои : 10.1006/jtbi.2001.2504 . ПМИД 12183125 .
- Лопес-Рикельме, Херман Октавио (2008). Муравьи как модельные системы сложного поведения. Нейробиологические основы химической связи и разделения труда у муравьев (к.б.н.). Национальный автономный университет Мексики. дои : 10.13140/RG.2.1.3145.1689 .