Четогната
| Стрелочные черви Временной диапазон: | |
|---|---|
 | |
| Chaetognatha и некоторые примеры их разнообразия. | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Subkingdom: | Эуметазоа |
| Клэйд : | ПараХоксозоа |
| Клэйд : | билатерия |
| Клэйд : | Нефрозоа |
| (без рейтинга): | протостомия |
| (без рейтинга): | Спиралия |
| Клэйд : | Гнатифера |
| Тип: | Четогната Лейкарт , 1854 г. |
| Сорт: | Стрелецоидея Клаус и Гроббен , 1905 год. [2] |
| Заказы | |
Chaetognatha t / k iː ˈ t ɒ ɡ n ə θ ə / или четогнаты / ˈ k iː ɒ ɡ n æ θ во всем s / (что означает щетинистые челюсти ) — тип хищных морских червей , которые являются основным компонентом планктона мире. Обычно известные как стрелы-черви , они в основном нектонны ; однако около 20% известных видов являются донными и могут прикрепляться к водорослям и камням . Они встречаются во всех морских водах: от поверхностных тропических вод и мелких приливных бассейнов до глубоководных и полярных регионов . Большинство щетогенных прозрачны и имеют торпедообразную форму, но некоторые глубоководные виды имеют оранжевый цвет . Их размер варьируется от 2 до 120 миллиметров (от 0,1 до 4,7 дюйма).
Четогнаты были впервые описаны голландским натуралистом Мартинусом Слаббером в 1775 году. [3] По состоянию на 2021 год биологи признают 133 современных вида, относящихся к более чем 26 родам и восьми семействам . [3] Несмотря на ограниченное видовое разнообразие, численность особей велика. [4]
Черви-стрелы тесно связаны и, возможно, принадлежат к Gnathifera , кладе протостом , которые не принадлежат ни к Ecdysozoa , ни к Lophotrochozoa .
Анатомия
[ редактировать ]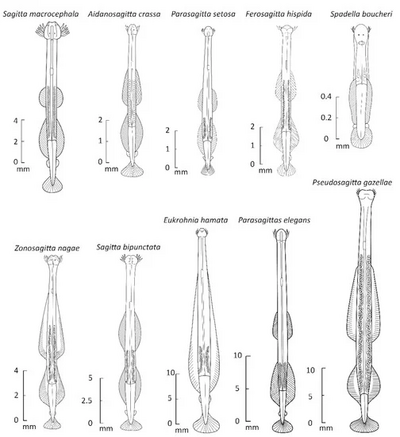
Четогнаты — прозрачные или полупрозрачные животные в форме дротика, покрытые кутикулой . Их длина варьируется от 1,5 до 105 мм у антарктических видов Pseudosagitta gasellae . [5] Размер тела как у особей одного вида, так и у разных видов, по-видимому, увеличивается с понижением температуры. [5] Тело разделено на голову, туловище и хвост. Около 80% тела занимают первичные продольные мышцы . [3]
Голова и пищеварительная система
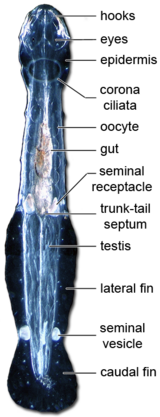
[ редактировать ]На каждой стороне головы имеется от четырех до четырнадцати крючковатых цепких шипов, расположенных по бокам полого преддверия, в котором находится рот. Шипы используются при охоте, а во время плавания животного прикрываются гибким капюшоном, возникающим из области шеи. Шипы и зубы состоят из α- хитина , а голова защищена хитиновым панцирем. [3]

Рот открывается в мускулистую глотку , содержащую железы, обеспечивающие смазку прохождения пищи. Отсюда прямая кишка проходит по всей длине туловища к анусу прямо перед хвостом. Кишечник является основным местом пищеварения и включает пару дивертикулов вблизи переднего конца. [6] Материалы перемещаются по полости тела с помощью ресничек . Отходы просто выводятся через кожу и задний проход. Виды Eucrohniid обладают масляной вакуолью, тесно связанной с кишечником. вне сезона производства Этот орган содержит эфиры воска, которые могут способствовать размножению и росту Eucrohnia hamata в арктических морях. [7] Благодаря расположению масляной вакуоли в центре тракта этот орган также может влиять на плавучесть, балансировку и передвижение. [8]
Обычно щетогенные не пигментированы, однако кишечник некоторых глубоководных видов содержит оранжево-красные каротиноидные пигменты. [3]
Нервная и сенсорная системы
[ редактировать ]Нервная система достаточно проста и имеет типичную анатомию протостома . [3] Состоит из ганглиозного нервного кольца, окружающего глотку. Мозг состоит из двух различных функциональных доменов: переднего домена нейропиля и заднего домена нейропиля. Первый, вероятно, управляет мышцами головы, приводящими в движение позвоночник и пищеварительную систему. Последний связан с глазами и ресничной короной. Предполагаемая сенсорная структура с неизвестной функцией, ретроцеребральный орган, также находится в заднем домене нейропиля. [3] Дорсальный ганглий — самый крупный, но от всех ганглиев по длине тела отходят нервы.
У Chaetognaths два сложных глаза, каждый из которых состоит из нескольких пигментных чашечковых глазков сросшихся между собой ; у некоторых глубоководных и троглобитовых видов глаза непигментированы или отсутствуют. [3] Кроме того, имеется ряд чувствующих щетинок, расположенных рядами по бокам тела, где они, вероятно, выполняют функцию, аналогичную функции боковой линии у рыб. Дополнительный изогнутый пучок сенсорных щетинок лежит над головой и шеей. [6] Почти все щетинкочелюстные имеют «непрямые» или «перевернутые» глаза в зависимости от ориентации фоторецепторных клеток; только некоторые виды Eukhroniidae имеют «прямые» или «вывернутые» глаза. [3] Уникальной особенностью глаза четогната является пластинчатая структура мембран фоторецепторов, содержащая сетку круглых пор шириной 35-55 нм. [3]
Важная механосенсорная система, состоящая из реснитчатых рецепторных органов, обнаруживает вибрации, позволяя четогнатам обнаруживать плавательные движения потенциальной добычи. Другой орган на дорсальной части шеи, ресничный венец, вероятно, участвует в хеморецепции. [3]
Внутренние органы
[ редактировать ]Полость тела выстлана брюшиной и поэтому представляет собой настоящий целом и разделена на по одному отсеку с каждой стороны туловища и дополнительные отсеки внутри головы и хвоста, полностью разделенные перегородками. Хотя у них есть рот с одним или двумя рядами крошечных зубов, сложные глаза и нервная система, у них нет выделительной и дыхательной систем. [9] [3] Хотя часто говорят, что у хетогнат отсутствует кровеносная система, у них есть рудиментарная гемальная система, напоминающая таковую у кольчатых червей . [3]
стрелы Рабдомеры происходят из микротрубочек длиной 20 нм и шириной 50 нм, которые, в свою очередь, образуют конические тельца, содержащие гранулы и нитевидные структуры. Тело колбочки происходит от реснички . [10]
Передвижение
[ редактировать ]Туловище несет одну или две пары боковых плавников со структурами, внешне похожими на плавниковые лучи рыб, с которыми они не гомологичны . В отличие от плавников позвоночных, эти боковые плавники состоят из утолщенной базальной мембраны, отходящей от эпидермиса . Дополнительный хвостовой плавник прикрывает постанальный хвост. [6] Два вида хетогнатов, Caecosagitta macrocephala и Eukrohnia fowleri , имеют биолюминесцентные органы на плавниках. [11] [12]
Четогнаты плавают короткими рывками, используя дорзо-вентральные волнообразные движения тела, при этом их хвостовой плавник способствует движению, а плавники тела - стабилизации и рулевому управлению. [13] Мышечные движения были описаны как одни из самых быстрых у многоклеточных животных . [3] Мышцы непосредственно возбудимы электрическим током или сильными К+ растворами ; основным нервно-мышечным медиатором является ацетилхолин. [3]
Размножение и жизненный цикл
[ редактировать ]Все виды являются гермафродитами и несут как яйцеклетки , так и сперматозоиды . [4] У каждого животного есть пара семенников в хвосте и пара яичников в задней части основной полости тела. Незрелые сперматозоиды выходят из семенников, созревают внутри полости хвоста, а затем плывут по короткому протоку к семенному пузырьку , где упаковываются в сперматофор . [6]
Во время спаривания каждая особь помещает сперматофор на шею своего партнера после разрыва семенного пузырька. Сперматозоиды быстро выходят из сперматофора и плывут вдоль средней линии животного, пока не достигнут пары маленьких пор прямо перед хвостом. Эти поры соединяются с яйцеводами , в которые из яичников уже перешли развитые яйцеклетки, и именно здесь происходит оплодотворение. [6] Семяприемники и яйцеводы накапливают и хранят сперматозоиды, необходимые для выполнения нескольких циклов оплодотворения. [3] некоторые донные представители Spadellidae Известно, что перед совокуплением проводят сложные ритуалы ухаживания . [3] например Параспаделла готовая . [14]
Яйца в основном планктонные, за исключением некоторых видов, таких как Ferosagitta hispida , которые прикрепляют яйца к субстрату. [3] У эукронии яйца развиваются в сумчатых мешках или прикрепляются к водорослям . [15] Яйца обычно вылупляются через 1-3 дня. Четогнаты не подвергаются метаморфозу и не имеют четко выраженной личиночной стадии. [6] [3] необычная черта среди морских беспозвоночных; [14] однако существуют значительные морфологические различия между новорожденными и взрослыми особями в отношении пропорций, хитиновых структур и развития плавников. [3] [16]
Продолжительность жизни щетогнатов варьируется, но коротка; самый длинный зарегистрированный период составил 15 месяцев в Sagitta friderici . [16]
Поведение
[ редактировать ]Мало что известно о поведении и физиологии стреловидных червей из-за сложности их культивирования и реконструкции их естественной среды обитания. [3] Известно, что при более высоких температурах они питаются чаще. Планктонные щетки часто должны плавать непрерывно, «подпрыгивая и тонув», чтобы оставаться в нужном месте в слое воды, и активно плавать, чтобы поймать добычу. Все они склонны держать тело слегка наклоненным, а голову — вниз. [3] Они часто демонстрируют «скользящее» поведение, на некоторое время медленно погружаясь, а затем быстро догоняя плавниками. [14] Бентические виды обычно остаются прикрепленными к субстратам, таким как камни, водоросли или морские травы, реже на вершине или между песчинками, и действуют более строго как хищники, устраивающие засады, оставаясь на месте, пока мимо не пройдет добыча. [3] Добычу обнаруживают благодаря ресничному забору и пучковым органам, улавливающим вибрации. [3] - Например, особи Spadella cephaloptera атакуют стеклянный или металлический зонд, вибрирующий с достаточной частотой. [14] Чтобы поймать добычу, черви-стрелы выпрыгивают вперед сильным взмахом хвостового плавника. [3] При контакте с добычей они надевают капюшон на хватательные шипы, так что он образует клетку вокруг добычи и приводит ее в контакт со ртом. Они заглатывают свою добычу целиком. [14]
Экология
[ редактировать ]Четогнаты встречаются во всех океанах мира, от полюсов до тропиков, а также в солоноватых и устьевых водах. Они обитают в самых разных средах: от гидротермальных источников до глубокого морского дна океана, зарослей морских водорослей и морских пещер. [3] Большинство из них являются планктонными, и они часто являются вторым по распространенности компонентом зоопланктона с биомассой от 10 до 30% от биомассы веслоногих ракообразных . [3] В Канадском бассейне одни только щетинкочелюстные составляют около 13% биомассы зоопланктона. [17] По существу, они экологически значимы и являются ключевым источником пищи для рыб и других хищников, включая коммерчески значимые рыбы, такие как скумбрия или сардины . [18] 58% известных видов являются пелагическими, [5] при этом около трети видов являются эпибентосными или мейобентосными или обитают в непосредственной близости от субстрата. [3] Четогнаты зарегистрированы на глубине до 5000, а возможно, и 6000 метров. [5]
Наибольшая плотность щетиконосцев наблюдается в фотической зоне мелководий. [3] Более крупные виды щеток, как правило, живут глубже в воде, но молодь проводят выше в толще воды. [14] Однако черви-стрелы ежедневно совершают вертикальную миграцию , проводя день на более низких глубинах, чтобы избежать хищников, а ночью приближаются к поверхности. Их положение в толще воды может зависеть от освещенности, температуры, солености, возраста и обеспеченности пищей. Они не могут плавать против океанических течений и используются как гидрологический индикатор течений и водных масс. [3]
Все четвероногие — хищники , устраивающие засаду и охотящиеся на других планктонных животных, в основном веслоногих и ветвистоусых ракообразных. [6] [3] но также амфиподы, криль и личинки рыб. [18] Взрослые особи могут питаться более молодыми особями того же вида. [19] Сообщается также, что некоторые виды всеядны и питаются водорослями и детритом. [20] Известно, что хетогнаты используют нейротоксин тетродотоксин , чтобы подчинить добычу. [21] возможно, синтезируется видами бактерий Vibrio . [3]
Генетика
[ редактировать ]Митохондриальный геном
[ редактировать ]МтДНК . стрелы-червя Spadella cephaloptera была секвенирована в 2004 году, и на тот момент это был самый маленький известный митохондриальный геном многоклеточных животных, его длина составляла 11 905 пар оснований [22] (теперь его превзошел михондриальный геном гребневика Mnemiopsis leidyi , длина которого составляет 10 326 пар оснований). [23] Все гены митохондриальной тРНК отсутствуют. Гены MT -ATP8 и MT-ATP6 также отсутствуют. [22] МтДНК . Paraspadella gotoi , также секвенированная в 2004 году, еще меньше (11 403 п.н.) и демонстрирует аналогичную структуру: в ней отсутствует 21 из 22 обычно присутствующих генов тРНК и имеется только 14 из 37 нормально присутствующих генов [24]
Четогнаты демонстрируют уникальное митохондриальное геномное разнообразие у особей одного и того же вида. [25]
Филогения
[ редактировать ]
Внешний
[ редактировать ]Эволюционные взаимоотношения щетиконосцев долгое время оставались загадочными. Чарльз Дарвин заметил, что черви-стрелы «удивительны своей неясностью своего родства». [14] В прошлом хетогнаты традиционно, но ошибочно, эмбриологами классифицировались как из вторичноротые . -за особенностей эмбриона, напоминающих вторичноротые Линн Маргулис и К.В. Шварц поместили щетогенных к вторичноротым в своей классификации Пяти царств . [27] Однако некоторые особенности развития расходятся с вторичноротыми и либо сходны со Spiralia , либо уникальны для Chaetognatha. [3]
| Краткое изложение взаимоотношений гнатифер в недавних исследованиях, включая Chaetognatha внутри клады, при этом спорные отношения представлены в виде политомии. [28] [29] [30] [31] [32] |
| |||
| Четогнаты на многоклеточном древе жизни считаются сестринской группой Gnathifera. [3] |
Молекулярная филогения показывает, что Chaetognatha на самом деле являются протостомами . Томас Кавальер-Смит относит их к протостомам в своей классификации Шести Королевств . [33] Сходство между щетогенными и нематодами, упомянутое выше, может поддержать тезис о протостомах - фактически, щетогенных иногда рассматривают как базальных экдизозойных или лофотрохозойных животных . [34] В большинстве исследований их молекулярной филогении Chaetognatha появляется близко к основанию протостомного дерева. [35] Это может объяснить их эмбриональные особенности вторичноротых. Если бы щетогенные ответвились от протостом до того, как у них развились характерные эмбриональные признаки протостомов, они могли бы сохранить признаки вторичноротых, унаследованные от ранних двулатеральных предков. Таким образом, щетогнаты могут быть полезной моделью предков билатерий. [36] Исследования нервной системы стреловидных червей показывают, что их следует размещать внутри протостомов. [37] [38] Согласно работам 2017 и 2019 годов, щетинкочелюстные относятся либо к [39] [40] или являются сестринской группой Gnathifera . [3]
Внутренний
[ редактировать ]Ниже представлено согласованное эволюционное древо Chaetognatha, основанное как на морфологических, так и на молекулярных данных по состоянию на 2021 год. [3]
Ископаемая запись
[ редактировать ]Из-за мягкого тела щетогенные плохо окаменевают. Несмотря на это, было описано несколько ископаемых видов щетинистых. [1] Четогнаты впервые появились в кембрийский период. Окаменелости полных тел были официально описаны из нижнего кембрия сланцев Маотяньшань в Юньнани, Китай ( Eognathacantha ercainella Chen & Huang [41] и Protosagitta spinosa Hu [42] ) и среднекембрийские сланцы Бёрджесс Британской Колумбии (( Oesia disjuncta Walcott [43] ), (точка зрения, которую оспаривают Конвей Моррис (2009) и Capinatator praetermissus .) Кембрийский щетогенный стволовой группы , Timorebestia , впервые описанный в 2024 году, был намного крупнее современных видов, что показывает, что щетогнаты играли разные роли в морских экосистемах по сравнению с сегодня. [44] Более поздний хетогнат, Paucijaculum samamithion Schram, был описан из биоты Мейзон-Крик из пенсильванского горизонта штата Иллинойс.
Считалось, что хетогнаты, возможно, связаны с некоторыми животными, сгруппированными с конодонтами . Однако было показано, что сами конодонты являются зубными элементами позвоночных . В настоящее время считается, что протоконодонтов элементы (например, Protohertzina anabarica Missarzhevsky, 1973), вероятно, захватывают не зубы конодонтов, а шипы щеток. Раньше подозревали, что в раннем кембрии существовали щетогенные особи только по этим элементам протоконодонтов, но более поздние открытия окаменелостей тела подтвердили их присутствие тогда. [45] Есть свидетельства того, что щетки были важными компонентами океанической пищевой сети уже в раннем кембрии. [46]
История
[ редактировать ]Первое известное описание хетогната было опубликовано голландским натуралистом Мартинусом Слаббером в 1770-х годах; он также придумал название «червь-стрела». [47] [5] Зоолог Анри Мари Дюкроте де Бленвиль также вкратце упомянул о вероятных щетогнатах, но понимал их как пелагических моллюсков. Первое описание принятого в настоящее время вида щетогеновых Sagitta bipunctata относится к 1827 году. [48] [5] Среди первых зоологов, описывающих стрельчатых червей, есть Чарльз Дарвин , который делал о них заметки во время путешествия « Бигля» и в 1844 году посвятил им статью. [49] В следующем году Август Дэвид Крон опубликовал раннее анатомическое описание Sagitta bipunctata . [50] [18]
Термин «хетогнат» был придуман в 1856 году Рудольфом Лейкартом . Он же был первым, кто предположил принадлежность рода Sagitta к отдельной группе: «В настоящий момент представляется наиболее естественным рассматривать Sagittas как представителей небольшой собственной группы, совершающей переход от настоящих кольчатых червей (вначале все любрицины) к нематодам, и их вполне можно назвать Chaetognathi». [51] [5]
Современная систематика Chaetognatha начинается в 1911 году Риттером-Захони. [52] [18] и позже объединен Такаси Токиока в 1965 году. [53] [5] [18] и Роберт Бьери в 1991 году. [54] Токиока ввел отряды Phragmophora и Aphragmophora и классифицировал четыре семейства, шесть родов, в общей сложности 58 видов — плюс вымершую Amiskwia , классифицированную как настоящие примитивные хетогнаты, в отдельный класс Archisagittoidea. [18]
Некоторое время Chaetognaths считались принадлежащими или родственными вторичноротым , но подозрения в их родстве со Spiralia или другими протостомами присутствовали уже в 1986 году. [22] Их сходство с протостомами было выяснено в 2004 году путем секвенирования и анализа мтДНК . [22]
Заражение гигантскими вирусами
[ редактировать ]
В 2018 году повторный анализ фотографий электронной микроскопии 1980-х годов позволил ученым идентифицировать гигантский вирус ( Meelsvirus ), поражающий Adhesisagitta hispida ; его место размножения является ядерным, а вирионы (длина: 1,25 мкм) покрыты оболочкой. [55] В 2019 году повторный анализ других предыдущих исследований показал, что структуры, которые были приняты в 1967 году за щетинки, присутствуют на поверхности вида Spadella cephaloptera . [56] и в 2003 году для бактерий, заражающих Paraspadella gotoi , [57] на самом деле представляли собой гигантские вирусы веретенообразной формы с оболочкой и цитоплазматическим местом размножения. [58] Вирусный вид, поражающий P. gotoi , максимальная длина которого составляет 3,1 мкм, был назван Klothovirus casanovai ( Klotho — греческое название одной из трех Судьб, атрибутом которых было веретено, и casanovai , в честь Пр. Ж.-П. Казанова, посвятивший большую часть своей научной жизни изучению щетогенных). Другой вид был назван Megaklothovirus horridgei (в честь Адриана Хорриджа , первого автора статьи 1967 года). На фотографии один из вирусов M. horridgei , хотя и усеченный, имеет длину 3,9 мкм, что примерно вдвое превышает длину бактерии Escherichia coli . В вирионах присутствует множество рибосом , но их происхождение остается неизвестным (клеточное, вирусное или частично вирусное). На сегодняшний день известно, что гигантские вирусы, поражающие многоклеточных животных, встречаются исключительно редко.
Ссылки
[ редактировать ]- ^ Перейти обратно: а б Ваннье Дж., Штайнер М., Ренвуаз Э., Ху С.С., Казанова Дж.П. (март 2007 г.). «Раннекембрийское происхождение современных пищевых сетей: свидетельства хищных червей-стрелок» . Труды Королевского общества B: Биологические науки . 274 (1610): 627–33. дои : 10.1098/rspb.2006.3761 . ПМК 2197202 . ПМИД 17254986 .
- ^ «Sagittoidea Клаус и Гроббен, 1905 год» . Интегрированная система таксономической информации . Проверено 8 февраля 2012 г.
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п д р с т в v В х и С аа аб и объявление но из в ах есть также Перес, Иван; Мюллер, Карстен Х.Г.; Харцш, Штеффен (2021). «Глава 15: Четогната» . В Шируотере, Бернд; ДеСалле, Роб (ред.). Зоология беспозвоночных: подход «Древо жизни» . ЦРК Пресс. п. 231. ИСБН 978-1-4822-3582-1 . Проверено 14 августа 2023 г.
- ^ Перейти обратно: а б Боун Кью, Капп Х, Пьеро-Бультс AC, ред. (1991). Биология хетогнатов . Лондон: Издательство Оксфордского университета . ISBN 978-0-19-857715-7 .
- ^ Перейти обратно: а б с д и ж г час Поли, Дэниел; Лян, Цуй; Сиань, Вэйвэй; Чу, Элейн; Байи, Николя (2021). «Размеры, рост и размножение стрелковых червей (Chaetognatha) в свете теории ограничения жаберного кислорода (GOLT)» . Журнал морской науки и техники . 9 (12): 1397. doi : 10.3390/jmse9121397 .
- ^ Перейти обратно: а б с д и ж г Барнс, Роберт Д. (1982). Зоология беспозвоночных . Филадельфия, Пенсильвания: Холт-Сондерс Интернэшнл. стр. 1046–1050. ISBN 978-0-03-056747-6 .
- ^ Григор, Джордан Дж.; Шмид, Мориц С.; Фортье, Луи (01 ноября 2017 г.). «Рост и размножение щеток Eukrohnia hamata и Parasagitta elegans в канадском Северном Ледовитом океане: капитальное разведение против прибыльного разведения» . Журнал исследований планктона . 39 (6): 910–929. дои : 10.1093/plankt/fbx045 . ISSN 0142-7873 .
- ^ Понд, Дэвид В. (1 июня 2012 г.). «Физические свойства липидов и их роль в контроле распространения зоопланктона в океанах» . Журнал исследований планктона . 34 (6): 443–453. дои : 10.1093/plankt/fbs027 . ISSN 0142-7873 .
- ^ Королевства и домены: Иллюстрированный путеводитель по типам жизни на Земле
- ^ «Фоторецепция». Британская энциклопедия из Британской энциклопедии 2006, DVD Ultimate Reference Suite . 2009.
- ^ Хэддок С.Х., Дело Дж.Ф. (20 января 1994 г.). «Биолюминесцентный хетогнат». Природа . 367 (6460): 225–226. Бибкод : 1994Natur.367..225H . дои : 10.1038/367225a0 . S2CID 4362422 .
- ^ Туесен Е.В., Гетц Ф.Е., Хэддок С.Х. (октябрь 2010 г.). «Биолюминесцентные органы двух глубоководных червей-стрелок, Eukrohnia fowleri и Caecosagitta macrocephala, с дальнейшими наблюдениями за биолюминесценцией у щетогенных». Биологический вестник . 219 (2): 100–11. дои : 10.1086/BBLv219n2p100 . ПМИД 20972255 . S2CID 14502203 .
- ^ Джордан CE (1992). «Модель быстрого старта плавания при промежуточном числе Рейнольдса: волнообразное передвижение четогната Sagitta elegans » . Журнал экспериментальной биологии . 163 (1): 119–137. дои : 10.1242/jeb.163.1.119 .
- ^ Перейти обратно: а б с д и ж г Болл, Элдон Э.; Миллер, Дэвид Дж. (2006). «Филогения: продолжающаяся классификационная загадка четогнатов» . Современная биология . 16 (15): 593–596 р. Бибкод : 2006CBio...16.R593B . дои : 10.1016/j.cub.2006.07.006 . hdl : 1885/26305 . ПМИД 16890517 . S2CID 18793650 .
- ^ Теразаки, М.; Миллер, CB (1 ноября 1982 г.). «Размножение мезо- и батипелагических щеток рода Eucrohnia». Морская биология . 71 (2): 193–196. Бибкод : 1982MarBi..71..193T . дои : 10.1007/BF00394629 . ISSN 1432-1793 . S2CID 84519862 .
- ^ Перейти обратно: а б Дапонте, MC; Капитанио, Флорида; Нахабедиан, Делавэр; Виньяс, доктор медицины; Негри, РМ (2004). «Sagitta friderici Ritter-Záhony (Chaetognatha) из вод Южной Атлантики: численность, структура популяции и жизненный цикл» . Журнал морских наук ICES . 61 (4): 680–686. Бибкод : 2004ICJMS..61..680D . doi : 10.1016/j.icesjms.2004.03.006 .
- ^ Кособокова Ксения Н.; Хопкрофт, Рассел Р. (2010). «Разнообразие и вертикальное распределение мезозоопланктона в арктическом канадском бассейне». Глубоководные исследования, часть II: Актуальные исследования в океанографии . 57 (1–2): 96–110. Бибкод : 2010DSRII..57...96K . дои : 10.1016/j.dsr2.2009.08.009 .
- ^ Перейти обратно: а б с д и ж Чу, Сохви; Чон, Ман-Ки; Со, Хо Ён (2022). «Таксономическая переоценка щетогенных (Chaetognatha, Sagittoidea, Aphragmophora) из корейских вод» . ZooKeys (1106): 165–211. Бибкод : 2022ZooK.1106..165C . дои : 10.3897/zookeys.1106.80184 . ПМЦ 9848744 . ПМИД 36760822 .
- ^ Салливан, Барбара К. (1980). «Пищевое поведение Sagitta elegans и Eukrohnia hamata (Chaetognatha) in situ в зависимости от вертикального распределения и численности добычи на океанской станции «P»1». Лимнология и океанография . 25 (2): 317–326. Бибкод : 1980LimOc..25..317S . дои : 10.4319/lo.1980.25.2.0317 .
- ^ Григор, Джордан Дж.; Шмид, Мориц С.; Кауэтт, Марианна; Сен-Онж, Вики; Браун, Томас А.; Бартелеми, Роксана-М. (01.07.2020). «Неплотоядное питание арктических щекочелюстных» . Прогресс в океанографии . 186 : 102388. Бибкод : 2020Proce.18602388G . дои : 10.1016/j.pocean.2020.102388 . ISSN 0079-6611 . S2CID 225636394 .
- ^ Туесен Э.В. (1991). «Яд тетродотоксина хетогнатов». В Bone Q, Капп Х, Пьеро-Бультс AC (ред.). Биология щетогнатов . Издательство Оксфордского университета. стр. 55–60. ISBN 978-0-19-857715-7 .
- ^ Перейти обратно: а б с д Папийон, Дэниел; Перес, Иван; Каби, Ксавье; Ле Парко, Янник (2004). «Идентификация четогнатов как протостомов подтверждается анализом их митохондриального генома» . Молекулярная биология и эволюция . 21 (11): 2122–2129. дои : 10.1093/molbev/msh229 . ПМИД 15306659 .
- ^ Формаджиони, Алессандро; Лучетти, Андреа; Плацци, Федерико (2021). «Митохондриальный геномный ландшафт: портрет митохондриального генома через 40 лет после первой полной последовательности» . Жизнь . 11 (7): 663. Бибкод : 2021Life...11..663F . дои : 10.3390/life11070663 . ПМЦ 8303319 . ПМИД 34357035 .
- ^ Хелфенбейн, Кевин Г.; Фуркад, Х.Мэтью; Ванджани, Рохит Г.; Бур, Джеффри Л. (2004). «Митохондриальный геном Paraspadella gotoi сильно редуцирован и указывает на то, что хетогнаты являются сестринской группой протостомов» . Труды Национальной академии наук . 101 (29): 10639–10643. Бибкод : 2004PNAS..10110639H . дои : 10.1073/pnas.0400941101 . ПМЦ 489987 . ПМИД 15249679 .
- ^ Марлетаз, Фердинанд; Ле Парко, Янник; Лю, Шэнлинь; Пейненбург, Катя ТСА (2017). «Чрезвычайная митогеномная изменчивость в природных популяциях щетинистых» . Геномная биология и эволюция . 9 (6): 1374–1384. дои : 10.1093/gbe/evx090 . ПМК 5470650 . ПМИД 28854623 .
- ^ Беккуш, Николя; Гонсиоровски, Людвик (2022). «Тщательное изменение наборов морфологических данных улучшает филогенетические рамки: переоценка размещения ископаемого Amiskwia sagittiformis » (PDF) . Журнал систематической палеонтологии . 20 : 1–14. дои : 10.1080/14772019.2022.2109217 .
- ↑ Таксон Systema Naturae 2000: Phylum Chaetognatha per Margulis and Schwartz. Архивировано 27 ноября 2005 г. в Wayback Machine (выберите Margulis & Schwartz в разделе «Классификация») - последнее получение 25 ноября 2006 г.
- ^ Марлетаз, Фердинанд; Пейненбург, Катя ТСА; Гото, Тайчиро; Сато, Нориюки; Рохсар, Дэниел С. (2019). «Новая спиральная филогения относит загадочных стреловидных червей к гнатиферам» (PDF) . Современная биология . 29 (2): 312–318.е3. Бибкод : 2019CBio...29E.312M . дои : 10.1016/j.cub.2018.11.042 . ПМИД 30639106 .
- ^ Винтер, Якоб; Парри, Люк А. (2019). «Двусторонние элементы челюсти Amiskwia sagittiformis устраняют морфологический разрыв между челюстно-челюстными и щетогнатными» (PDF) . Современная биология . 29 (5): 881–888.e1. Бибкод : 2019CBio...29E.881V . дои : 10.1016/j.cub.2019.01.052 . ПМИД 30799238 .
- ^ Фрёбиус, Андреас К.; Фанч, Питер (2017). «Гены Rotiferan Hox дают новое представление об эволюции строения многоклеточных животных» . Природные коммуникации . 8 (1): 9. Бибкод : 2017NatCo...8....9F . дои : 10.1038/s41467-017-00020-w . ПМК 5431905 . ПМИД 28377584 .
- ^ Лаумер, Кристофер Э.; Беккуш, Николя; Кербл, Александра; Гетц, Фрейя; Невес, Рикардо К.; Соренсен, Мартин В.; Кристенсен, Рейнхардт М.; Хейнол, Андреас; Данн, Кейси В.; Гирибет, Гонсало; Ворсаа, Катрин (2015). «Спиральная филогения определяет эволюцию микроскопических линий» . Современная биология . 25 (15): 2000–2006. Бибкод : 2015CBio...25.2000L . дои : 10.1016/j.cub.2015.06.068 . ПМИД 26212884 .
- ^ Силафф, Мальта; Шмидт, Ханно; Удар, Торстен Х.; Розенкранц, Дэвид; Марк Уэлч, Дэвид Б.; Ханкельн, Томас; Херлин, Хольгер (2016). «Филогения Syndermata (син. Rotifera): порядок митохондриальных генов подтверждает, что эпизойные Seisonidea являются сестрой эндопаразитических скребней в пределах монофилетических Hemirotifera». Молекулярная филогенетика и эволюция . 96 : 79–92. Бибкод : 2016МОЛПЭ..96...79С . дои : 10.1016/j.ympev.2015.11.017 . ПМИД 26702959 .
- ↑ Таксон Systema Naturae 2000: Phylum Chaetognatha per Cavalier-Smith. Архивировано 27 ноября 2005 г. в Wayback Machine (выберите Cavalier-Smith в разделе «Классификация») - последнее получение 25 ноября 2006 г.
- ^ Матус Д.К., Копли Р.Р., Данн К.В., Хейнол А., Экклстон Х., Халанич К.М., Мартиндейл М.К., Телфорд М.Дж. (август 2006 г.). «Широкая выборка таксонов и генов указывает на то, что щетинкочелюстные являются протостомами» . Современная биология . 16 (15): Р575-6. Бибкод : 2006CBio...16.R575M . дои : 10.1016/j.cub.2006.07.017 . ПМИД 16890509 . S2CID 11284138 .
- ^ Марлетаз Ф., Мартин Э., Перес Ю., Папийон Д., Кабит Икс, Лоу С.Дж., Фриман Б., Фазано Л., Доссат С., Винкер П., Вайссенбах Дж., Ле Парко Ю. (август 2006 г.). «Филогеномика Chaetognath: протостом с вторичноротым развитием» . Современная биология . 16 (15): Р577-8. Бибкод : 2006CBio...16.R577M . дои : 10.1016/j.cub.2006.07.016 . ПМИД 16890510 . S2CID 18339954 .
- ^ Папийон Д., Перес Ю., Кабит Х., Ле Парко Ю. (ноябрь 2004 г.). «Идентификация четогнатов как протостомов подтверждается анализом их митохондриального генома» . Молекулярная биология и эволюция . 21 (11): 2122–9. дои : 10.1093/molbev/msh229 . ПМИД 15306659 .
- ^ Ригер В., Перес И., Мюллер Ч., Липке Е., Сомбке А., Ханссон Б.С., Харцш С. (февраль 2010 г.). «Иммуногистохимический анализ и 3D-реконструкция головной нервной системы Chaetognatha: понимание эволюции раннего билатерального мозга?». Биология беспозвоночных . 129 (1): 77–104. дои : 10.1111/j.1744-7410.2010.00189.x .
- ^ Харцш С., Мюллер CH (май 2007 г.). «Новый взгляд на вентральный нервный центр Sagitta: значение для филогенетического положения Chaetognatha (стрелковых червей) и эволюции двусторонней нервной системы» . Границы в зоологии . 4:14 . дои : 10.1186/1742-9994-4-14 . ПМЦ 1885248 . ПМИД 17511857 .
- ^ Фрёбиус А.С., Фунч П. (апрель 2017 г.). «Гены Rotiferan Hox дают новое представление об эволюции строения многоклеточных животных» . Природные коммуникации . 8 (1): 9. Бибкод : 2017NatCo...8....9F . дои : 10.1038/s41467-017-00020-w . ПМК 5431905 . ПМИД 28377584 .
- ^ Марлетас Ф., Пейненбург К.Т., Гото Т., Сато Н., Рохсар Д.С. (январь 2019 г.). «Новая спиральная филогения помещает загадочных червей-стрелок среди гнатиферов» (PDF) . Современная биология . 29 (2): 312–318.e3. Бибкод : 2019CBio...29E.312M . дои : 10.1016/j.cub.2018.11.042 . ПМИД 30639106 .
- ^ Чен Цзюй, Хуан Ди (октябрь 2002 г.). «Возможный нижнекембрийский четогнат (стреляной червь)». Наука . 298 (5591): 187. doi : 10.1126/science.1075059 . ПМИД 12364798 . S2CID 544495 .
- ^ Ху SX (2005). «Тафономия и палеоэкология раннекембрийской биоты Чэнцзян из Восточного Юньнани, Китай». Berliner Paläobiologische Abhandlungen . 7 : 1–197.
- ^ Санявский Х (2005). «Кембрийские четогнаты обнаружены в окаменелостях в сланцах Берджесс» (PDF) . Acta Palaeontologica Polonica . 50 (1): 1–8. Архивировано (PDF) из оригинала 9 октября 2022 г.
- ^ Пак Тэ-Юн С.; Нильсен, Мортен Лунде; Парри, Люк А.; Соренсен, Мартин Винтер; Ли, Миринаэ; Ким, Джи-Хун; Ан, Инхе; Парк, Чанкунь; Де Виво, Джачинто; Смит, М. Пол; Харпер, Дэвид А.Т.; Нильсен, Арне Т.; Винтер, Якоб (2024). «Гигантский хетогнат стеблевой группы» . Достижения науки . 10 (1): eadi6678. Бибкод : 2024SciA...10I6678P . дои : 10.1126/sciadv.adi6678 . ПМЦ 10796117 . ПМИД 38170772 .
- ^ Санявский Х (2002). «Новые доказательства протоконодонтного происхождения щетинистых» (PDF) . Acta Palaeontologica Polonica . 47 (3): 405–419. Архивировано (PDF) из оригинала 9 октября 2022 г.
- ^ Ваннье, Дж.; Штайнер, М.; Ренвуаз, Э.; Ху, С.-Х; Казанова, Ж.-П (2007). «Раннекембрийское происхождение современных пищевых сетей: данные по хищным червям-стрелам» . Труды Королевского общества B: Биологические науки . 274 (1610): 627–633. дои : 10.1098/rspb.2006.3761 . ПМК 2197202 . ПМИД 17254986 .
- ^ Слаббер, Мартинус (1778). Физические удовольствия, содержащие микроскопические наблюдения за домашними и зарубежными водными и наземными животными . Харлем: Дж. Бош. стр. 46–48.
- ^ Куой, JRC; Гаймар, Ж. П. «Зоологические наблюдения, сделанные на борту «Астролябии» в мае 1826 года в Гибралтарском проливе (продолжение и конец). Описание родов Biphore, Carinaire, Hyale, Flèche, Cléodore, Anatife и Briarée». Энн. наук. Нат. 1827, 10, 225–239.
- ^ Дарвин, К. «Наблюдения за структурой и распространением рода Sagitta». Энн. Маг. Нат. Хист. 1844, 13, 1–6.
- ^ Крон, Массачусетс (1845 г.). «XXXI.— Анатомо-физиологические наблюдения над Sagitta bipunctata» . Анналы и журнал естественной истории . 16 (106): 289–304. дои : 10.1080/037454809496523 .
- ^ Лейкарт, Р. Дополнения и исправления к первому тому «Справочника по зоологии» Дж. ван дер Хувена. Систематизированный обзор важнейших достижений последнего времени: по зоологии беспозвоночных; Л. Восс: Лейпциг, Германия, 1856 г. (на немецком языке).
- ^ Риттер-Захони Р. (1911) «Редакция Chaetognathan». Немецкая южнополярная экспедиция 1901–1903 гг . Том 13, Зоология 5-я глава 1: 1–72.
- ^ Токиока, Такаси (1965). «Таксономический очерк Chaetognatha» . Публикации Сетоской морской биологической лаборатории . 12 (5): 335–357. дои : 10.5134/175381 . S2CID 81053929 .
- ^ Биери, Роберт. «Систематика Chaetognatha». в Биологии щетогнатов (1991): 122-136.
- ^ Шинн Г.Л., Буллард Б.Л. (19 сентября 2018 г.). Сан-Мартин С (ред.). «Ультраструктура Meelsvirus: ядерный вирус стрелковых червей (тип Chaetognatha), продуцирующий гигантские «хвостые» вирионы» . ПЛОС ОДИН . 13 (9): e0203282. Бибкод : 2018PLoSO..1303282S . дои : 10.1371/journal.pone.0203282 . ПМК 6145532 . ПМИД 30231047 .
- ^ Хорридж Г.А., Бултон П.С. (14 ноября 1967 г.). «Обнаружение добычи Chaetognatha с помощью чувства вибрации». Труды Лондонского королевского общества. Серия Б. Биологические науки . 168 (1013): 413–419. Бибкод : 1967RSPSB.168..413H . дои : 10.1098/rspb.1967.0072 . S2CID 86422882 .
- ^ Казанова Дж. П., Дювер М., Гото Т. (октябрь 2003 г.). «Ультраструктурное исследование и онтогенез придатков и связанной с ними мускулатуры Paraspadella (Chaetognatha)». Ткани и клетки . 35 (5): 339–51. дои : 10.1016/S0040-8166(03)00055-7 . ПМИД 14517101 .
- ^ Роксана-Мари Бартелеми, Эрик Фор, Тайчиро Гото: случайное открытие у морских беспозвоночных (тип Chaetognatha) самых длинных гигантских вирусов, о которых сообщалось до настоящего времени . В: Биология , 2019, Аннотация
Внешние ссылки
[ редактировать ]- Изображение газеллы Pseudosagitta с крилем в кишечнике, сделанное Тасманским институтом аквакультуры и рыболовства.
- Chaetognatha of the World - последнее посещение 13 декабря 2006 г.
- Эрик Форе, Роксана-Мари Бартелеми: Специфические митохондриальные оц-тРНК в типе Chaetognatha . В: Журнал энтомологии и зоологических исследований 7 (3), апрель 2019 г., стр. 304–315. hal-02130653

| Базы данных органов управления : Национальные |
|---|