Бделлоидея
| Бделлоидные коловратки Временной диапазон: | |
|---|---|
 | |
| СЭМ, показывающая морфологические вариации бделлоидных коловраток и их челюстей. | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | коловратка |
| Суперкласс: | Евротатор |
| Сорт: | Бделлоидея Гудзон , 1884 год. |
Bdelloidea / ˈ d ɛ l ɔɪ d i ə / ( греч. βδέλλα , bdella , «пиявка») — класс коловраток , встречающихся в пресноводных средах обитания по всему миру. Описано более 450 видов бделлоидных коловраток (или бделлоидов). [1] отличаются друг от друга главным образом на основании морфологии . [2] Основными характеристиками, отличающими бделлоидов от родственных групп коловраток, являются исключительно партеногенетическое размножение и способность выживать в сухих, суровых условиях путем перехода в состояние вызванного высушиванием покоя ( ангидробиоза ) на любом этапе жизни. [3] Их часто называют «древними асексуалами» из-за их уникальной бесполой истории, которая, согласно ископаемым свидетельствам, насчитывает более 25 миллионов лет назад. [4] Бделлоидные коловратки представляют собой микроскопические организмы, обычно длиной от 150 до 700 мкм. [3] Большинство из них слишком малы, чтобы их можно было увидеть невооруженным глазом, но выглядят как крошечные белые точки даже через слабую ручную линзу , особенно при ярком свете. В июне 2021 года биологи сообщили о восстановлении бделлоидных коловраток после 24 000-летней заморозки в сибирской вечной мерзлоте . [5]
отношения Эволюционные
Тип класса Rotifera традиционно включал три : Bdelloidea, Monogononta и Seisonidea . [6] До 1990 года филогенетические исследования, основанные на морфологии, по-видимому, указывали на то, что сестринской группой бделлоидных коловраток была Monogononta, а сейсонидные коловратки были ранней дивергентной внешней группой . [7]

Современные молекулярно-филогенетические исследования показывают, что это классическое понимание «коловраток» является неполным ( парафилетическим ), поскольку в нем не учитывается четвертая клада близкородственных организмов: скребни , или черви с шипами. [10] Первоначально классифицированные как отдельный тип , молекулярные и морфологические данные, накопленные в период с 1994 по 2014 год, указывают на то, что скребни образуют монофилетическую группу с Bdelloidea, Monogononta и Seisonidea. [8] [11] Чтобы учесть этот вывод, некоторые авторы расширяют термин «коловратки», включив в него сильно модифицированные паразитические «скребниевые коловратки» наряду с бделлоидными, моногононтными и сейзонидными коловратками. [12] Другие называют группу четырех таксонов Syndermata, термин, полученный из их общего синцитиального эпидермиса. [11]
Положение Bdelloidea внутри Syndermata (или Rotifera) не совсем ясно. Альтернативные возможные филогенетические отношения внутри клады иллюстрируются прилагаемыми кладограммами. По состоянию на 2014 год «наиболее полный филогеномный анализ взаимоотношений синдерматанов» был основан на данных транскриптома всех четырех групп: [8] и предоставили «сильную поддержку» гипотезе, проиллюстрированной в левом нижнем углу рисунка, согласно которой Seisonidea и скребни являются сестринскими таксонами. Исследование также показало, что сестринской группой этого таксона является Bdelloidea, тогда как Monogononta является внешней группой для всех трех. Это означало бы, что ближайшими ныне живущими родственниками бделлоидных коловраток являются не моногононтные коловратки, как считалось ранее, а сейзонидные коловратки и скребни, несмотря на их сильно модифицированную морфологию.
Классификация и идентификация [ править ]
Bdelloidea — класс типа коловраток , состоящий из трёх отрядов: Philodinavida, Philodinida и Adinetida. [13] Эти отряды разделены на четыре семейства и около 450 видов . [14] Поскольку эти организмы бесполы, обычное определение вида как группы организмов, способных создавать плодовитое потомство, неприменимо, поэтому концепция вида у этих организмов вместо этого основана на смеси морфологических и молекулярных данных. Исследования ДНК показывают, что разнообразие намного больше, чем предполагают первоначальные морфологические классификации. [15] [16]
Бделлоидов можно идентифицировать только на глаз, пока они живы, поскольку многие из характеристик, важных для классификации, связаны с питанием и ползанием; однако генетическая идентификация бделлоидов возможна на мертвых особях. После сохранения особи сжимаются в «капли», что ограничивает анализ. [17] В настоящее время существуют три методологии морфологической идентификации, две из которых считаются устаревшими: Бартош (1951). [18] и Доннер (1965). [14] Третий метод представляет собой диагностический ключ, разработанный в 1995 году Шилом. [17]
Морфология [ править ]
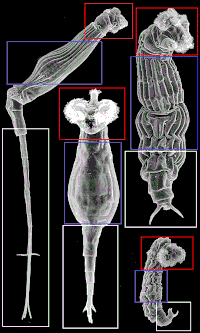
Выделяют три основных отдела тела бделлоидов: голову, туловище и стопу. На соседнем изображении изображена каждая область, чтобы показать, как части тела могут сильно различаться, хотя они называются одинаково в зависимости от вида. Бделлоиды обычно имеют хорошо развитую коронку, разделенную на две части, на втягивающейся головке.
Некоторые отличительные особенности бделлоидов включают:
- Хорошо развитые железы стопы [17]
- Ротовое отверстие с длинным пищеводом [17]
- Крепкие зубы (обозначаются индексом зуба) [17]
- Много ресничек [17]
- Видоспецифическая форма верхней губы [17]
- Тип короны в зависимости от заказа [3]
- Филодиниды состоят из двух ресничных дисков.
- Адинетиды состоят из вентрального мерцательного поля.
- У Philodinavida маленькая коронка.
Бделлоидные пищеварительная и репродуктивная системы можно найти в туловищных частях тела, при этом желудок является наиболее заметным из органов . У некоторых родов ( Habrotrocha , Otostephanos и Scepanotrocha ) бделлоид можно идентифицировать по появлению в желудке отдельных сферических гранул, которые выделяются в виде фекалий. Эти гранулы являются отличительной характеристикой, поскольку все остальные виды выделяют фекалии в виде сыпучего материала. [3]
Большинство бделлоидов втягивают ногу во время еды, но есть четыре рода, у которых нога отсутствует: Adineta , Bradyscela , Henoceros и Philodinavus . Это влияет не только на то, как они питаются, но и на то, как они ползают; например, Adineta и Bradyscela скользят, тогда как представители других родов петляют. [3]
Поведение [ править ]
Поведение бделлоидов можно разделить на четыре категории: питание, передвижение, размножение и поведение, вызванное стрессом.
Кормление [ править ]
Специфическое пищевое поведение бделлоидов разнообразно, но большинство из них используют кольца ресничек в короночном органе для создания потоков воды, которые переносят пищу через рот к мастаксному органу , приспособленному специально для измельчения пищи. [19] В пищу входят взвешенные бактерии, водоросли, детрит и другие вещества.
Передвижение [ править ]
По-видимому, существует три основных способа передвижения: свободное плавание , ползание по субстрату и сидячее положение . Ползание (или ползание) предполагает попеременные шаги головой и хвостом, как это делают некоторые пиявки , что и дало группе их название ( греческое βδέλλα или bdella , что означает пиявка). В этом видео показано, как бделлоиды передвигаются в трех различных ситуациях: передвижение и питание бделлоидных коловраток .
Репродукция [ править ]
Бделлоиды представляют интерес для изучения эволюции пола , поскольку самцов никогда не наблюдалось. [20] а самки размножаются исключительно путем партеногенеза , формы бесполого размножения, при которой эмбрионы растут и развиваются без необходимости оплодотворения; это похоже на апомиксис , наблюдаемый у некоторых растений. [21] У каждой особи имеются парные гонады . Несмотря на то, что они были бесполыми в течение миллионов лет, они разделились на более чем 450 видов и очень похожи на другие виды коловраток, размножающихся половым путем.
Однако новое исследование предоставило доказательства межиндивидуального генетического обмена и рекомбинации у Adineta vaga , вида, который ранее считался бесполым. [22]
Adineta vaga способна осуществлять репарацию ДНК путем нередукционного мейоза . [23] Восстановление зародышевой ДНК происходит в определенный период оогенеза, во время которого гомологичные хромосомы принимают мейоподобную конфигурацию. [23] Такое восстановление ДНК зародышевой линии приводит к точному восстановлению генетического материала, передаваемого потомству.
Эволюция партеногенетического облигатного размножения
В 2003 г. способ бесполого размножения бделлоидных коловраток был совершенно неизвестен. [24] Одна из теорий возникновения облигатного партеногенеза у бделлоидных коловраток заключалась в том, что партеногенные линии утратили способность реагировать на сигнал, индуцирующий пол, поэтому эти линии сохранили свою асексуальность. [25] Облигатные партеногенетические штаммы бделлоидных коловраток производят сигнал, индуцирующий пол, но утратили способность реагировать на этот сигнал. Позже было обнаружено, что неспособность реагировать на сигналы, индуцирующие пол, у облигатных партеногенов вызвана простым менделевским наследованием гена op. [26]
стрессом вызванное , Поведение
Бделлоиды способны выдерживать стрессы окружающей среды, переходя в состояние покоя, известное как ангидробиоз , которое позволяет организму быстро обезвоживаться и, таким образом, противостоять высыханию. При подготовке к этому состоянию покоя многие метаболические процессы корректируются, чтобы соответствовать изменению состояния; например, производство защитных химикатов. [27] Бделлоиды могут оставаться в этом состоянии, известном как «ксеросома», до тех пор, пока не вернется достаточное количество воды, после чего они регидратируются и становятся активными в течение нескольких часов. Вылупление детенышей произойдет только тогда, когда условия будут наиболее благоприятными. [28] Эти формы покоя также известны как криптобиоз или покой. Известно, что бделлоиды выживают в этом состоянии до 9 лет, ожидая возвращения благоприятных условий. [28] В дополнение к выживанию при высыхании за счет ангидробиоза, стресс от высыхания у двух видов бделлоидов фактически помог сохранить физическую форму и даже улучшить плодовитость их вида . [29] Коловушки, которых постоянно содержали гидратированными, жили хуже, чем те, которые были высушены и регидратированы. [30]
Bdelloidea разработали уникальный механизм, помогающий преодолеть одну из главных опасностей бесполого размножения. Согласно Красной Королевы гипотезе коэволюции , облигатные асексуалы будут вымерты из-за быстро меняющихся паразитов и патогенов, потому что они не смогут достаточно быстро изменить свои генотипы , чтобы не отставать в этой бесконечной гонке. Однако в популяциях бделлоидных коловраток многие паразиты уничтожаются в периоды длительного высыхания. [31] Более того, высушенные бделлоидные коловратки легко уносятся ветром из зараженных паразитами мест обитания и создают новые, здоровые популяции в других местах, что позволяет им избежать Красной Королевы, перемещаясь во времени и пространстве вместо того, чтобы использовать секс для изменения своего генотипа. [32]
Когда эти существа восстанавливаются после высыхания, было показано, что они подвергаются потенциально уникальному генетическому процессу, при котором горизонтальный перенос генов. происходит [ нужна ссылка ] , в результате чего значительная часть бделлоидного генома, до 10%, была получена путем горизонтального переноса генов от бактерий, грибов и растений. [33] Как и почему происходит горизонтальный перенос генов у бделлоидов, в настоящее время вызывает много споров; особенно в отношении возможных связей между чужеродными генами и процессом высыхания, а также возможных связей с древней асексуальностью бделлоидов.
Когда они полностью высыхают, их ДНК распадается на множество частей. А когда они возвращаются к жизни после регидратации, это создает возможность для фрагментов чужеродной ДНК проникнуть в их геном. Этот процесс был усовершенствован 60 миллионов лет назад, когда они таким образом захватили бактериальный ген, что дало им новую систему регуляции генов. Новая система использовалась для транспозонов . контроля [34]
Бделлоидные коловратки чрезвычайно устойчивы к повреждениям от ионизирующей радиации благодаря тем же адаптациям, сохраняющим ДНК, которые используются для выживания в состоянии покоя. [35] Эти адаптации включают чрезвычайно эффективный механизм восстановления двухцепочечных разрывов ДНК. [36] Этот механизм репарации был изучен у двух видов Bdelloidea: Adineta vaga , [36] и Филодина розеола . [37] и, по-видимому, включает митотическую рекомбинацию между гомологичными участками ДНК внутри каждого вида.
генов Горизонтальный перенос
Крупномасштабный горизонтальный перенос генов бактерий, растений и грибов в бделлоидные коловратки [38] был задокументирован и может представлять собой важный фактор в эволюции бделлоидов.
Галерея [ править ]
 Боковой вид бделлоида.
Боковой вид бделлоида. Фронтальный вид короны бделлоида.
Фронтальный вид короны бделлоида. Боковой вид бделлоида.
Боковой вид бделлоида. Боковой вид бделлоида.
Боковой вид бделлоида. Вид сбоку на бделлоида в воде, богатой водорослями.
Вид сбоку на бделлоида в воде, богатой водорослями. Образец рода Philodina.
Образец рода Philodina.






Ссылки [ править ]
- ^ Доннер, Джозеф (1965). «Отряд Bdelloidea (Rotatoria, коловратки)». Опознавательные книги по почвенной фауне Европы, том 6 . Берлин: Издательство Академии. ОСЛК 6733231 . и Сегерс, Хендрик (2007). «Аннотированный контрольный список коловраток (Phylum Rotifera) с примечаниями по номенклатуре, таксономии и распространению». Зоотакса . 1590 : 3–104. Абстрактный .
- ^ Сегерс, Хендрик (2007). Аннотированный контрольный список коловраток (Phylum Rotifera) с примечаниями по номенклатуре, таксономии и распространению (PDF) . Окленд: Магнолия Пресс. ISBN 978-1-86977-129-4 .
- ↑ Перейти обратно: Перейти обратно: а б с д и Риччи, Клаудия (2000). «Определитель родов бделлоидных коловраток». Гидробиология . 418 : 73–80. дои : 10.1023/A:1003840216827 . S2CID 44054669 .
- ^ Пойнар-младший, ГО; Риччи, Клаудия (1992). «Бделлоидные коловратки в доминиканском янтаре: доказательства партеногенетической преемственности». Эксперименты . 48 (4): 408–410. дои : 10.1007/BF01923444 . S2CID 13098228 .
- ^ Рено, Марион (7 июня 2021 г.). «Это крошечное существо пережило 24 000 лет заморозки в вечной мерзлоте Сибири. Микроскопические животные были заморожены, когда шерстистые мамонты еще бродили по планете, но были восстановлены, как будто время не прошло» . «Нью-Йорк Таймс» . Проверено 7 июня 2021 г.
- ^ Кинг, Чарльз Э.; Риччи, Клаудия; Шонфельд, Джастин; Серра, Мануэль (сентябрь 2005 г.). «Эволюционная динамика моделей жизненного цикла бделлоидов и моногононтных коловраток». Гидробиология . 546 (1): 55–70. CiteSeerX 10.1.1.455.6499 . дои : 10.1007/s10750-005-4102-9 . S2CID 25975998 .
- ^ Уоллес, Роберт Ли; Колберн, Ребекка Арлин (декабрь 1989 г.). «Филогенетические взаимоотношения внутри типа Rotifera: отряды и род Notholca». Гидробиология . 186–187 (1): 311–318. дои : 10.1007/BF00048926 . S2CID 20809514 .
- ↑ Перейти обратно: Перейти обратно: а б с Вей-Фабрициус, Александра Р.; Херлин, Хольгер; Ригер, Бенджамин; Розенкранц, Дэвид; Витек, Александр; Уэлч, Дэвид Б. Марк; Эберсбергер, Инго; Ханкельн, Томас; Шмитц, Юрген (10 февраля 2014 г.). «Данные транскриптома раскрывают взаимоотношения синдерматана и предполагают эволюцию эндопаразитизма у скребней через эпизойную стадию» . ПЛОС ОДИН . 9 (2): e88618. Бибкод : 2014PLoSO...988618W . дои : 10.1371/journal.pone.0088618 . ПМЦ 3919803 . ПМИД 24520404 .
- ^ Ласек-Нессельквист, Эрика (23 августа 2012 г.). «Митогеномная переоценка филогении бделлоидов и взаимоотношений между синдерматами» . ПЛОС ОДИН . 7 (8): е43554. Бибкод : 2012PLoSO...743554L . дои : 10.1371/journal.pone.0043554 . ПМЦ 3426538 . ПМИД 22927990 .
- ^ Уэлч, Дэвид Б. Марк (2001). «Ранний вклад молекулярной филогенетики в понимание эволюции коловраток». Гидробиология . 446 : 315–322. дои : 10.1023/A:1017502923286 . S2CID 28895228 .
- ↑ Перейти обратно: Перейти обратно: а б Альрикс, Вилко Х. (1997). «Эпидермальная ультраструктура Seison nebaliae и Seison annulatus , а также сравнение эпидермальных структур Gnathifera». Зооморфология . 117 (1): 41–48. дои : 10.1007/s004350050028 . S2CID 24719056 .
- ^ Нильсен, Клаус (2012). Эволюция животных: взаимоотношения живых типов (3-е изд.). Оксфорд: Издательство Оксфордского университета. ISBN 978-0199606030 .
- ^ Мелоне, Джулио и Риччи, Клаудия (1995). «Вращательный аппарат у бделлоидов». Гидробиология . 313 (1): 91–98. дои : 10.1007/BF00025935 . S2CID 35937088 .
- ↑ Перейти обратно: Перейти обратно: а б Доннер, Джозеф (1965). Заказать Бделлоидея . Издательство Академии. п. 297. ИСБН 9789031908851 .
- ^ Кая, Мурат; Эрниу, Элизабет А.; Барракло, Тимоти Г. и Фонтането, Диего (2009). «Несогласованные оценки разнообразия традиционной таксономии и ДНК-таксономии бделлоидных коловраток» . Разнообразие и эволюция организмов . 9 (1): 3–12. дои : 10.1016/j.ode.2008.10.002 .
- ^ Фонтането, Диего; Кая, Мурат; Эрниу, Элизабет А. и Барракло, Тимоти Г. (2009). «Чрезвычайные уровни скрытого разнообразия микроскопических животных (Rotifera), выявленные с помощью таксономии ДНК». Молекулярная филогенетика и эволюция . 53 (1): 182–189. дои : 10.1016/j.ympev.2009.04.011 . ПМИД 19398026 .
- ↑ Перейти обратно: Перейти обратно: а б с д и ж г Шил, Рассел Дж. (1995). Руководство по идентификации коловраток, кладоцер и веслоногих ракообразных из внутренних вод Австралии . Австралия: Кооперативный исследовательский центр экологии пресной воды. ISBN 978-0-646-22410-7 .
- ^ Бартош, Эмануэль (1951). «Чехословацкая ротатория ордена Bdelloidea». Мемуары Чехословацкого зоологического общества в Праге . 15 : 241–500.
- ^ Клюземанн, Дж.; Кляйнов, В.; Питерс, В. (1990). «Твердые части (трофусы) мастакса коловратки действительно содержат хитин: данные исследований Brachionus plicatilis». Гистохим. Клеточная Биол . 94 (3): 277–283. дои : 10.1007/bf00266628 . ПМИД 2401635 . S2CID 26501940 .
- ^ Джадсон, Оливия П.; Нормарк, Бенджамин Б. (1996). «Древние асексуальные скандалы». Тенденции в экологии и эволюции . 11 (2): 41–46. дои : 10.1016/0169-5347(96)81040-8 . ПМИД 21237759 .
- ^ Милиус, Сьюзен (1 ноября 2002 г.). «Бделлоиды: секса нет уже более 40 миллионов лет» . Новости науки . Проверено 6 ноября 2016 г.
- ^ Вахрушева О.А.; Мнацаканова Е.А.; Галимов Ю.Р. (18 декабря 2020 г.). «Геномные признаки рекомбинации в природной популяции бделлоидной коловратки Adineta vaga» . Природа . 11 (1): 6421. Бибкод : 2020NatCo..11.6421V . дои : 10.1038/s41467-020-19614-y . ПМЦ 7749112 . ПМИД 33339818 .
- ↑ Перейти обратно: Перейти обратно: а б Тервангне, М.; Николас, Э.; Хеспелс, Б.; Гертер, Л.; Дева, Дж.; Демази, К.; Хойскин, AC; Халлет, Б.; Ван Донинк, К. (2022). «Репарация ДНК во время нередукционного мейоза у бесполой коловратки Adineta vaga» . Достижения науки . 8 (48): eadc8829. Бибкод : 2022SciA....8C8829T . дои : 10.1126/sciadv.adc8829 . ПМЦ 9710870 . ПМИД 36449626 .
- ^ Симон, Жан-Кристоф; Дельмотт, Франсуа; Риспе, Клод; Криз, Тереза (2003). «Филогенетические отношения между партеногенами и их половыми родственниками: возможные пути партеногенеза у животных» . Биологический журнал Линнеевского общества . 79 (1): 151–163. дои : 10.1046/j.1095-8312.2003.00175.x .
- ^ Стельцер, Клаус-Питер (2008). «Облигатный асексуал у коловратки и роль половых сигналов» . Журнал эволюционной биологии . 21 (1): 287–293. дои : 10.1111/j.1420-9101.2007.01437.x . ПМИД 17995949 .
- ^ Штельцер, Клаус-Петер; Шмидт, Джоанна; Видлройтер, Аннелизе; Рисс, Симона (20 сентября 2010 г.). «Потеря полового размножения и карликовость у мелких многоклеточных животных» . ПЛОС ОДИН . 5 (9): e12854. Бибкод : 2010PLoSO...512854S . дои : 10.1371/journal.pone.0012854 . ПМЦ 2942836 . ПМИД 20862222 .
- ^ Кроу, Джон Х. (1971). «Ангидробиоз: нерешенная проблема». Американский натуралист . 105 (946): 563–573. дои : 10.1086/282745 . JSTOR 2459752 . S2CID 84512992 .
- ↑ Перейти обратно: Перейти обратно: а б Гуидетти, Роберто и Йонссон, К. Ингемар (2002). «Долгосрочная ангидробиотическая выживаемость полуназемных микрометазоа» . Журнал зоологии . 257 (2): 181–187. CiteSeerX 10.1.1.630.9839 . дои : 10.1017/s095283690200078x .
- ^ Риччи, Клаудия и Фонтането, Диего (2009). «Важность быть бделлоидом: экологические и эволюционные последствия покоя» . Итальянский журнал зоологии . 76 (3): 240–249. дои : 10.1080/11250000902773484 .
- ^ Риччи, Клаудия; Каприоли, Мануэла; Фонтането, Диего (2007). «Стресс и приспособленность партеногенов: является ли покой ключевой особенностью бделлоидных коловраток?» . Эволюционная биология BMC . 7 (Дополнение 2): S9. дои : 10.1186/1471-2148-7-S2-S9 . ЧВК 1963474 . ПМИД 17767737 .
- ^ Уилсон, Кристофер Г.; Шерман, Пол В. (29 января 2010 г.). «Древние бесполые бделлоидные коловратки спасаются от смертельных грибковых паразитов, высыхая и сдуваясь ветром». Наука . 327 (5965): 574–576. Бибкод : 2010Sci...327..574W . дои : 10.1126/science.1179252 . ПМИД 20110504 . S2CID 43898914 .
- ^ Уилсон, Кристофер Г.; Шерман, Пол В. (22 августа 2013 г.). «Пространственное и временное спасение от грибкового паразитизма в природных сообществах древних бесполых бделлоидных коловраток» . Труды Королевского общества Б. 280 (1765): 20131255. doi : 10.1098/rspb.2013.1255 . ПМЦ 3712457 . ПМИД 23825214 .
- ^ Боскетти, Кьяра; Пушкина-Станчева Наталья; Хоффманн, Пиа и Таннаклифф, Алан (2011). «Чужие гены и новые гены гидрофильных белков участвуют в реакции высыхания бделлоидной коловратки Adineta ricciae » . Журнал экспериментальной биологии . 214 (1): 59–68. дои : 10.1242/jeb.050328 . ПМИД 21147969 .
- ^ 60-летние бактерии проливают свет на «новую» систему модификации ДНК у животных
- ^ Гладышев, Евгений и Мезельсон, Мэтью (1 апреля 2008 г.). «Чрезвычайная устойчивость бделлоидных коловраток к ионизирующей радиации» . Труды Национальной академии наук . 105 (13): 5139–5144. Бибкод : 2008PNAS..105.5139G . дои : 10.1073/pnas.0800966105 . ПМК 2278216 . ПМИД 18362355 .
- ↑ Перейти обратно: Перейти обратно: а б Хеспилс Б., Кнапен М., Хано-Мамбрес Д., Хойскин А.С., Пино Ф., ЛУКАС С., Кошул Р., Ван Донинк К. (июль 2014 г.). «Ворота к генетическому обмену? Двухцепочечные разрывы ДНК у бделлоидной коловратки Adineta vaga, подвергшейся высыханию» (PDF) . Дж. Эвол. Биол . 27 (7): 1334–45. дои : 10.1111/jeb.12326 . ПМИД 25105197 .
- ^ Уэлч, Дэвид Б. Марк; Уэлч, Джессика Л. Марк и Мезельсон, Мэтью (1 апреля 2008 г.). «Доказательства вырожденной тетраплоидии у бделлоидных коловраток» . Труды Национальной академии наук . 105 (13): 5145–9. Бибкод : 2008PNAS..105.5145M . дои : 10.1073/pnas.0800972105 . ПМЦ 2278229 . ПМИД 18362354 .
- ^ Гладышев Евгений А.; Мезельсон, Мэтью; Архипова Ирина Р. (30 мая 2008 г.). «Массовый горизонтальный перенос генов у бделлоидных коловраток» . Наука . 320 (5880): 1210–1213. Бибкод : 2008Sci...320.1210G . дои : 10.1126/science.1156407 . ISSN 1095-9203 . ПМИД 18511688 . S2CID 11862013 .
Внешние ссылки [ править ]
- Знакомство с коловратками
- Мировой каталог Rotifer, авторы Jersabek CD и Leitner MF.
- The Weird Sisters. Архивировано 1 октября 2015 г. в Wayback Machine.
- Бделлоиды: секса нет уже более 40 миллионов лет. Архивировано 23 апреля 2008 г. в Wayback Machine.
- Эволюционный скандал из журнала Harvard Magazine.
- Кому вообще нужен секс (или мужчины)?
- Крошечное существо возвращается к жизни после 24 000 лет пребывания в сибирских морозах. Архивировано 28 июня 2021 г. в Wayback Machine.