Внешняя группа (кладистика)
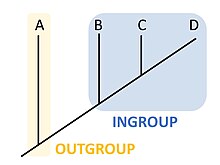
В кладистике или филогенетике группа внешняя [1] — более отдаленно родственная группа организмов, служащая референтной группой при определении эволюционных взаимоотношений внутренней группы, совокупности изучаемых организмов, и отличающаяся от социологических аутгрупп . Внешняя группа используется как точка сравнения внутренней группы и, в частности, позволяет филогению укоренить . Поскольку полярность (направление) изменения признаков может быть определена только на основе укоренившейся филогении, выбор внешней группы важен для понимания эволюции признаков на протяжении филогении. [2]
История
[ редактировать ]Хотя концепция внешних групп использовалась с самых первых дней кладистики, считается, что термин «внешняя группа» был придуман в начале 1970-х годов в Американском музее естественной истории . [3] До появления этого термина биологи-эволюционисты использовали различные другие термины, в том числе «бывшая группа», «родственная группа» и «внешние группы». [3]
Выбор внешней группы
[ редактировать ]Предполагается, что выбранная внешняя группа менее тесно связана с внутренней группой, чем внутренняя группа связана с самой собой. Эволюционный вывод из этих отношений заключается в том, что виды чужой группы имеют общего предка с внутренней группой, который старше, чем общий предок внутренней группы. Выбор внешней группы может изменить топологию филогении. [4] Поэтому филогенетики обычно используют более одной внешней группы в кладистическом анализе. Использование нескольких внешних групп предпочтительнее, поскольку оно обеспечивает более надежную филогению, защиту от плохих кандидатов в внешние группы и проверку гипотетической монофилии внутренней группы. [3] [5] [6]
Чтобы квалифицироваться как аутгруппа, таксон должен удовлетворять следующим двум характеристикам:
- Он не должен быть членом внутренней группы.
- Он должен быть связан с внутренней группой достаточно тесно, чтобы можно было проводить значимые сравнения с внутренней группой.
Следовательно, соответствующая внешняя группа должна однозначно находиться за пределами клады , представляющей интерес для филогенетического исследования. Внешняя группа, вложенная во внутреннюю группу, при использовании для укоренения филогении приведет к неверным выводам о филогенетических отношениях и эволюции признаков. [7] Однако оптимальный уровень родства внешней группы с внутренней зависит от глубины филогенетического анализа. Выбор близкородственной внешней группы по отношению к внутренней группе более полезен при рассмотрении тонких различий, в то время как выбор чрезмерно удаленной внешней группы может привести к ошибочному принятию конвергентной эволюции за прямую эволюционную связь из-за общего предка . [8] [9] Для поверхностной филогенетики — например, для выяснения эволюционных отношений клады внутри рода — подходящей внешней группой может быть член сестринской клады. [10] Однако для более глубокого филогенетического анализа можно использовать менее близкие таксоны. Например, Джарвис и др. (2014) использовали людей и крокодилов в качестве внешних групп при исследовании ранних ветвей филогении птиц. [11] В молекулярной филогенетике удовлетворение второго требования обычно означает, что последовательности ДНК или белков из внешней группы могут быть успешно сопоставлены с последовательностями из внутренней группы. Хотя существуют алгоритмические подходы к выявлению аутгрупп с максимальной глобальной экономией, они часто ограничены из-за неспособности отразить непрерывную количественную природу определенных состояний персонажей. [12] Состояния характера — это черты, наследственные или производные, которые влияют на построение моделей ветвления в филогенетическом дереве. [13]
Примеры
[ редактировать ]| Ингруппа | Внешняя группа |
|---|---|
| Великие обезьяны [14] | Гиббонс |
| Плацентарные млекопитающие [15] | Сумчатые |
| Хордовые [16] | Амбулакрарианец |
| покрытосеменные растения [17] | Голосеменные растения |
В каждом примере филогения организмов внутренней группы может быть основана на оценке одинаковых состояний признаков для одного или нескольких членов внешней группы.
См. также
[ редактировать ]- Апоморфия — производный признак организма.
- Сестринская группа — группа, которая может быть тесно связана с внутренней группой.
- Плезиоморфия — наследственный признак организма.
- Примитивный (филогенетика) — термин, обозначающий наследственные черты.
Ссылки
[ редактировать ]- ^ Гримальди, Дэвид; Энгель, Майкл С.; Энгель, Майкл С. (16 мая 2005 г.). Эволюция насекомых . Издательство Кембриджского университета. ISBN 9780521821490 .
- ^ Фаррис, Дж. С. (1982). «Аутгруппы и бережливость». Систематическая биология . 31 (3): 328–334. дои : 10.1093/sysbio/31.3.328 . ISSN 1063-5157 .
- ↑ Перейти обратно: Перейти обратно: а б с Никсон, Кевин; Карпентер, Джеймс (декабрь 1993 г.). «Об аутгруппах». Кладистика . 9 (4): 413–426. дои : 10.1111/j.1096-0031.1993.tb00234.x . S2CID 221577454 .
- ^ Гирибет, Г.; Рибера, К. (июнь 1998 г.). «Положение членистоногих в животном мире: поиск надежной внешней группы для внутренней филогении членистоногих». Молекулярная филогенетика и эволюция . 9 (3): 481–488. дои : 10.1006/mpev.1998.0494 . ПМИД 9667996 .
- ^ Барриэль, В.; Тасси, П. (июнь 1998 г.). «Укоренение нескольких внешних групп: консенсус против бережливости» . Кладистика . 14 (2): 193–200. дои : 10.1111/j.1096-0031.1998.tb00332.x . S2CID 84759858 .
- ^ де ла Торре-Барсена, Хосе Эдуардо; Колокотронис, С.О.; Ли, Эрнест; Стивенсон, Деннис; Бреннер, Эрик; Катари, Манприт; Коруцци, Глория; ДеСалле, Роб (2009). «Влияние выбора аутгруппы и недостающих данных на филогенетику основных семенных растений с использованием общегеномных данных EST» . ПЛОС ОДИН . 4 (6): е5764. Бибкод : 2009PLoSO...4.5764D . дои : 10.1371/journal.pone.0005764 . ПМК 2685480 . ПМИД 19503618 .
- ^ Мэддисон, Уэйн; и др. (1984). «Анализ аутгрупп и экономия» (PDF) . Систематическая зоология . 33 (1): 83–103. дои : 10.2307/2413134 . JSTOR 2413134 .
- ^ Уилберг, Эрик В. (01 июля 2015 г.). «Что такое внешняя группа? Влияние выбора внешней группы на филогенетическое положение Thalattosuria (Crocodylomorpha) и происхождение Crocodyliformes» . Систематическая биология . 64 (4): 621–637. дои : 10.1093/sysbio/syv020 . ISSN 1063-5157 . ПМИД 25840332 .
- ^ О'БРАЙЕН, МАЙКЛ Дж.; ЛАЙМАН, Р.ЛИ; СААБ, ЮСЕФ; СААБ, ЭЛИАС; ДАРВЕНТ, ДЖОН; ГЛОВЕР, ДЭНИЭЛ С. (2002). «Два вопроса археологической филогенетики: построение таксонов и отбор внешней группы». Журнал теоретической биологии . 215 (2): 133–150. дои : 10.1006/jtbi.2002.2548 . ПМИД 12051970 .
- ^ Дэвид А. Баум; Стейси Д. Смит (2013). Древовидное мышление: введение в филогенетическую биологию . Робертс. п. 175. ИСБН 978-1-936221-16-5 .
- ^ Джарвис, Э.; и др. (декабрь 2014 г.). «Анализ всего генома позволяет выявить ранние ветви древа жизни современных птиц» . Наука . 346 (6215): 1320–1331. Бибкод : 2014Sci...346.1320J . дои : 10.1126/science.1253451 . ПМЦ 4405904 . ПМИД 25504713 .
- ^ Стивенс, П.Ф. (1991). «Состояния характера, морфологические вариации и филогенетический анализ: обзор». Систематическая ботаника . 16 (3): 553–583. дои : 10.2307/2419343 . JSTOR 2419343 .
- ^ Рино, Валентин; Гранд, Анаис; Сарагета, Рене; Лорен, Мишель (1 мая 2015 г.). «Экспериментальная систематика: чувствительность кладистических методов к схемам поляризации и упорядочения символов» . Вклад в зоологию . 84 (2): 129–148. дои : 10.1163/18759866-08402003 .
- ^ Прадо-Мартинес, Хавьер; Маркес-Боне, Томас (2013). «Генетическое разнообразие и история популяций великих обезьян» . Природа . 499 (7459): 471–475. Бибкод : 2013Natur.499..471P . дои : 10.1038/nature12228 . ПМЦ 3822165 . ПМИД 23823723 .
- ^ Мерфи, Уильям; Прингл, Томас; Крайдер, Тесс; Спрингер, Марк; Миллер, Уэбб (2007). «Использование геномных данных для раскрытия корней филогении плацентарных млекопитающих» . Геномные исследования . 17 (4): 413–421. дои : 10.1101/гр.5918807 . ПМК 1832088 . ПМИД 17322288 .
- ^ Кэмерон, Крис; Гэри, Джеймс; Свалла, Билли (2000). «Эволюция строения тела хордовых: новые данные из филогенетического анализа типов вторичноротых» . ПНАС . 97 (9): 4469–4474. дои : 10.1073/pnas.97.9.4469 . ЧВК 18258 . ПМИД 10781046 .
- ^ Мэтьюз, Сара; Донохью, Майкл (1999). «Корень филогении покрытосеменных растений, выведенный из дублирующих генов фитохромов». Наука . 286 (5441): 947–950. дои : 10.1126/science.286.5441.947 . ПМИД 10542147 .
| Соответствующие поля | ||
|---|---|---|
| Основные понятия | ||
| Методы вывода | ||
| Текущие темы | ||
| Групповые черты | ||
| Типы групп | ||
| Номенклатура | ||