Байесовский вывод в филогении
| Классификация | Эволюционная биология |
|---|---|
| Подклассификация | Молекулярная филогенетика |
| Оптимальные критерии поиска | Байесовский вывод |
Байесовский вывод филогении объединяет априорную информацию и информацию о правдоподобии данных для создания так называемой апостериорной вероятности деревьев, которая представляет собой вероятность того, что дерево правильное с учетом данных, априорной модели и модели правдоподобия. Байесовский вывод был введен в молекулярную филогенетику в 1990-х годах тремя независимыми группами: Брюсом Ранналой и Зихэн Янгом в Беркли, [1] [2] Боб Мау в Мэдисоне, [3] и Шуин Ли из Университета Айовы, [4] последние двое в то время были аспирантами. Этот подход стал очень популярным с момента выпуска программного обеспечения MrBayes в 2001 году. [5] и в настоящее время является одним из самых популярных методов молекулярной филогенетики.
Байесовский вывод об основах и основах филогении
[ редактировать ]
Байесовский вывод относится к вероятностному методу, разработанному преподобным Томасом Байесом на основе теоремы Байеса . Опубликованная посмертно в 1763 году, она стала первым выражением обратной вероятности и основой байесовского вывода. Независимо, не зная о работе Байеса, Пьер-Симон Лаплас разработал теорему Байеса в 1774 году. [6]
Байесовский вывод или метод обратной вероятности был стандартным подходом в статистическом мышлении до начала 1900-х годов, пока Р. А. Фишер не разработал то, что сейчас известно как классический/частотный/фишеровский вывод. Вычислительные трудности и философские возражения препятствовали широкому внедрению байесовского подхода до 1990-х годов, когда алгоритмы цепи Маркова Монте-Карло (MCMC) произвели революцию в байесовских вычислениях.
Байесовский подход к филогенетической реконструкции объединяет априорную вероятность дерева P(A) с правдоподобием данных (B) для создания апостериорного распределения вероятностей на деревьях P(A|B). [7] Апостериорная вероятность дерева — это вероятность того, что дерево правильное с учетом априорных данных и правильности модели правдоподобия.
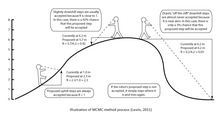
Методы MCMC можно описать в три этапа: сначала с помощью стохастического механизма новое состояние цепи Маркова предлагается . Во-вторых, рассчитывается вероятность того, что это новое состояние будет правильным. В-третьих, предлагается новая случайная величина (0,1). Если это новое значение меньше вероятности принятия, новое состояние принимается и состояние цепочки обновляется. Этот процесс выполняется тысячи или миллионы раз. Количество посещений одного дерева в ходе цепочки является аппроксимацией его апостериорной вероятности. Некоторые из наиболее распространенных алгоритмов, используемых в методах MCMC, включают алгоритмы Метрополиса – Гастингса, MCMC с соединением Метрополиса (MC³) и ЛОКАЛЬНЫЙ алгоритм Ларже и Саймона.
Алгоритм Метрополиса – Гастингса
[ редактировать ]Одним из наиболее распространенных используемых методов MCMC является алгоритм Метрополиса-Гастингса . [8] модифицированная версия оригинального алгоритма Метрополиса. [9] Это широко используемый метод случайной выборки из сложных и многомерных вероятностей распределения. Алгоритм Метрополиса описывается следующими шагами: [10] [11]
- Исходное дерево Ti выбирается случайным образом.
- Соседнее дерево T j выбирается из набора деревьев.
- Отношение R вероятностей (или функций плотности вероятности) T j и T i вычисляется следующим образом: R = f(T j )/f(T i )
- Если R ≥ 1, T j принимается в качестве текущего дерева.
- Если R < 1, T j принимается в качестве текущего дерева с вероятностью R, в противном случае T i сохраняется.
- На этом этапе процесс повторяется с шага 2 N раз.
Алгоритм продолжает работать до тех пор, пока не достигнет равновесного распределения. Также предполагается, что вероятность предложения нового дерева Tj , когда мы находимся в состоянии старого дерева Ti , равна той же вероятности предложения Ti , когда мы находимся в состоянии Tj . В противном случае применяются поправки Гастингса.Целью алгоритма Метрополиса-Гастингса является создание набора состояний с определенным распределением до тех пор, пока марковский процесс не достигнет стационарного распределения. Алгоритм состоит из двух компонентов:
- Потенциальный переход из одного состояния в другое (i → j) с использованием функции вероятности перехода q i,j
- Переход цепочки в состояние j с вероятностью α i,j и пребывание в состоянии i с вероятностью 1 – α i,j . [2]
MCMC, связанный с Метрополисом
[ редактировать ]Алгоритм MCMC, связанный с Метрополисом (MC³) [12] было предложено решить практическую проблему, связанную с перемещением цепи Маркова через пики, когда известно, что целевое распределение имеет несколько локальных пиков, разделенных низкими впадинами, которые существуют в древовидном пространстве. Это имеет место при эвристическом поиске по дереву по критериям максимальной экономии (MP), максимального правдоподобия (ML) и минимальной эволюции (ME), и того же можно ожидать для стохастического поиска по дереву с использованием MCMC. Эта проблема приведет к тому, что образцы не будут правильно аппроксимироваться апостериорной плотностью. (MC³) улучшает смешивание цепей Маркова при наличии нескольких локальных пиков апостериорной плотности. Он запускает несколько (m) цепочек параллельно, каждая для n итераций и с разными стационарными распределениями. , , где первый, - целевая плотность, а , выбраны для улучшения смешивания. Например, можно выбрать поэтапный нагрев вида:





![{\displaystyle \pi _{j}(\theta)=\pi (\theta)^{1/[1+\lambda (j-1)]},\ \ \lambda >0,}](https://wikimedia.org/api/rest_v1/media/math/render/svg/2e9d8c159f93b004aa99ba2ce2249578daf6db1e)
так, чтобы первая цепь представляла собой холодовую цепь с правильной целевой плотностью, а цепи это нагретые цепи. Обратите внимание, что увеличение плотности к власти с имеет эффект выравнивания распределения, аналогичный нагреву металла. В таком распределении легче перемещаться между пиками (разделенными впадинами), чем в исходном распределении. После каждой итерации предлагается обмен состояниями между двумя случайно выбранными цепочками посредством шага типа Метрополиса. Позволять быть текущим состоянием в цепочке , . Обмен между состояниями цепочек и принимается с вероятностью:








В конце цикла используется продукция только холодовой цепи, а продукция горячей цепи отбрасывается. С эвристической точки зрения горячие цепи будут довольно легко посещать локальные пики, а смена состояний между цепями позволит холодной цепи время от времени преодолевать долины, что приведет к лучшему перемешиванию. Однако, если нестабильна, предлагаемые обмены редко будут приняты. Это причина использования нескольких цепочек, которые отличаются лишь постепенно.

Очевидным недостатком алгоритма является то, что цепочки запускаются, и для вывода используется только одна цепочка. По этой причине, идеально подходит для реализации на параллельных машинах, поскольку каждая цепочка, как правило, требует одинакового объема вычислений на итерацию.


ЛОКАЛЬНЫЙ алгоритм Ларже и Саймона
[ редактировать ]ЛОКАЛЬНЫЕ алгоритмы [13] предлагает вычислительное преимущество по сравнению с предыдущими методами и демонстрирует, что байесовский подход способен оценить неопределенность с помощью вычислений, практичных в больших деревьях. Алгоритм LOCAL является усовершенствованием алгоритма GLOBAL, представленного Мау, Ньютоном и Ларже (1999). [14] в котором длины всех ветвей изменяются в каждом цикле. Алгоритмы LOCAL модифицируют дерево, выбирая внутреннюю ветвь дерева случайным образом. Каждый узел на концах этой ветви соединен с двумя другими ветвями. Из каждой пары случайным образом выбирается один. Представьте себе, что вы берете эти три выбранных края и натягиваете их, как бельевую веревку, слева направо, причем направление (лево/право) также выбирается случайным образом. Две конечные точки первой выбранной ветви будут иметь поддерево, свисающее, как кусок одежды, привязанный к линии. Алгоритм умножает три выбранные ветви на общую случайную величину, что похоже на растяжение или сжатие бельевой веревки. Наконец, самое левое из двух висящих поддеревьев отсоединяется и снова прикрепляется к бельевой веревке в случайно выбранном месте. Это будет дерево кандидатов.
Предположим, мы начали с выбора внутренней ветви длиной который разделяет таксоны и от остального. Предположим также, что у нас есть (случайным образом) выбранные ветви длиной и с каждой стороны, и что мы ориентировали эти ветки. Позволять , — текущая длина бельевой веревки. Мы выбираем новую длину, которая будет , где является однородной случайной величиной на . Тогда для алгоритма LOCAL вероятность принятия может быть вычислена как:










Оценка конвергенции
[ редактировать ]Чтобы оценить длину ветки 2-таксонного дерева под JC, в котором сайты однообразны и являются переменными, предполагают экспоненциальное априорное распределение со скоростью . Плотность . Вероятности возможных шаблонов сайта:






для однотипных сайтов и

Таким образом, ненормализованное апостериорное распределение имеет вид:

или, альтернативно,

Обновите длину ветки, выбирая новое значение равномерно случайным образом из окна половинной ширины. центрировано по текущему значению:


где равномерно распределяется между и . Принятиевероятность это:



Пример: , . Мы сравним результаты для двух значений , и . В каждом случае мы начнем с начальной длины и обновите длину раз.






Максимальная экономия и максимальная вероятность
[ редактировать ]

Существует множество подходов к реконструкции филогенетических деревьев, каждый из которых имеет свои преимущества и недостатки, и нет однозначного ответа на вопрос: «Какой метод лучше?». Максимальная экономия (MP) и максимальное правдоподобие (ML) — это традиционные методы, широко используемые для оценки филогении, и оба напрямую используют информацию о признаках, как это делают байесовские методы.
Максимальная экономия восстанавливает одно или несколько оптимальных деревьев на основе матрицы дискретных признаков для определенной группы таксонов и не требует модели эволюционных изменений. MP дает наиболее простое объяснение заданному набору данных, реконструируя филогенетическое дерево, которое включает в себя как можно меньше изменений в последовательностях. Поддержка ветвей дерева представлена процентом начальной загрузки . По той же причине, по которой он широко использовался, а также из-за его простоты, MP также подвергся критике и был отодвинут на второй план ML и байесовскими методами. MP представляет несколько проблем и ограничений. Как показал Фельзенштейн (1978), MP может быть статистически непоследовательным, [15] Это означает, что по мере накопления все большего количества данных (например, длины последовательности) результаты могут сойтись на неправильном дереве и привести к притяжению длинных ветвей - филогенетическому феномену, при котором таксоны с длинными ветвями (многочисленные изменения состояния признаков) имеют тенденцию казаться более тесно связанными в филогении, чем они есть на самом деле. Что касается морфологических данных, недавние исследования моделирования показывают, что экономия может быть менее точной, чем деревья, построенные с использованием байесовских подходов. [16] возможно, из-за чрезмерной точности, [17] хотя это оспаривается. [18] Исследования с использованием новых методов моделирования показали, что различия между методами вывода являются результатом используемой стратегии поиска и метода консенсуса, а не используемой оптимизации. [19]
Как и в случае максимальной экономии, метод максимального правдоподобия будет оценивать альтернативные деревья. Однако он учитывает вероятность того, что каждое дерево объясняет данные данные на основе модели эволюции. В этом случае дерево с наибольшей вероятностью объяснения данных выбирается среди других. [20] Другими словами, он сравнивает, как разные деревья предсказывают наблюдаемые данные. Введение модели эволюции в анализ ML дает преимущество перед MP, поскольку учитываются вероятность нуклеотидных замен и скорость этих замен, что позволяет более реалистично объяснить филогенетические связи таксонов. Важным фактором этого метода является длина ветки, которую игнорирует экономия, поскольку изменения с большей вероятностью произойдут на длинных ветвях, чем на коротких. Этот подход может устранить привлекательность длинных ветвей и объяснить большую согласованность ML по сравнению с MP. Хотя многие считают, что ML является лучшим подходом к выводу о филогениях с теоретической точки зрения, ML требует больших вычислительных ресурсов, и почти невозможно изучить все деревья, поскольку их слишком много. Байесовский вывод также включает в себя модель эволюции, и его основные преимущества перед MP и ML заключаются в том, что он более эффективен в вычислительном отношении, чем традиционные методы, он количественно определяет и устраняет источник неопределенности и способен включать сложные модели эволюции.
Подводные камни и споры
[ редактировать ]- Бутстрап-значения против апостериорных вероятностей. Было замечено, что значения бутстреп-поддержки, рассчитанные с учетом экономии или максимального правдоподобия, имеют тенденцию быть ниже, чем апостериорные вероятности, полученные с помощью байесовского вывода. [21] [22] [23] [24] [25] Это приводит к ряду вопросов, таких как: Приводят ли апостериорные вероятности к чрезмерной уверенности в результатах? [26] Являются ли значения начальной загрузки более надежными, чем апостериорные вероятности? Одним из фактов, лежащих в основе этого противоречия, является то, что все данные используются во время байесовского анализа и расчета апостериорных вероятностей, в то время как природа начальной загрузки означает, что в большинстве повторов начальной загрузки будут отсутствовать некоторые исходные данные. В результате биразделы (ветви), поддерживаемые относительно небольшим количеством символов в наборе данных, могут получать очень высокие апостериорные вероятности, но умеренную или даже низкую поддержку начальной загрузки, поскольку многие реплики начальной загрузки не содержат достаточного количества критических символов для получения бираздела.
- Споры об использовании априорных вероятностей. Использование априорных вероятностей для байесовского анализа многими рассматривается как преимущество, поскольку оно обеспечивает способ включения информации из источников, отличных от анализируемых данных. Однако когда такая внешняя информация отсутствует, приходится использовать априорную информацию, даже если невозможно использовать статистическое распределение для отражения полного незнания. Также вызывает беспокойство то, что байесовские апостериорные вероятности могут отражать субъективные мнения, когда априорные вероятности произвольны и субъективны.
- Выбор модели. Результаты байесовского анализа филогении напрямую коррелируют с выбранной моделью эволюции, поэтому важно выбрать модель, которая соответствует наблюдаемым данным, иначе выводы в филогении будут ошибочными. Многие ученые поднимали вопросы об интерпретации байесовского вывода, когда модель неизвестна или неверна. Например, чрезмерно упрощенная модель может дать более высокие апостериорные вероятности. [21] [27]
Программное обеспечение MrBayes
[ редактировать ]MrBayes — это бесплатный программный инструмент, который выполняет байесовский вывод филогении. Первоначально он был написан Джоном П. Хюльзенбеком и Фредериком Ронквистом в 2001 году. [28] По мере роста популярности байесовских методов MrBayes стал одним из предпочтительных программ для многих специалистов по молекулярной филогенетике. Он предлагается для операционных систем Macintosh, Windows и UNIX и имеет интерфейс командной строки. Программа использует стандартный алгоритм MCMC, а также вариант MCMC, связанный с Metropolis. MrBayes считывает выровненные матрицы последовательностей (ДНК или аминокислот) в стандартном формате NEXUS . [29]
MrBayes использует MCMC для аппроксимации апостериорных вероятностей деревьев. [9] Пользователь может изменить предположения о модели замещения, априорные данные и детали анализа MC³. Это также позволяет пользователю удалять и добавлять таксоны и символы для анализа. Среди нескольких нуклеотидных моделей программа включает в себя наиболее стандартную модель замены ДНК, 4x4, также называемую JC69, которая предполагает, что изменения между нуклеотидами происходят с равной вероятностью. [30] Он также реализует ряд моделей замены аминокислот 20x20 и моделей замены кодонов ДНК. Он предлагает различные методы смягчения предположения об одинаковой скорости замен в сайтах нуклеотидов. [31] Г-н Байес также может делать выводы о наследственных состояниях, учитывающих неопределенность филогенетического дерева и параметров модели.
Мистер Байес 3 [32] был полностью реорганизованной и реструктурированной версией оригинального MrBayes. Главной новинкой стала способность программного обеспечения учитывать неоднородность наборов данных. Эта новая структура позволяет пользователю смешивать модели и использовать преимущества эффективности байесовского анализа MCMC при работе с различными типами данных (например, белковыми, нуклеотидными и морфологическими). По умолчанию он использует Metropolis-Coupling MCMC.
MrBayes 3.2 был выпущен в 2012 году. [33] Новая версия позволяет пользователям выполнять несколько анализов параллельно. Он также обеспечивает более быстрые вычисления правдоподобия и позволяет делегировать эти вычисления графическим процессорам (GPU). Версия 3.2 предоставляет более широкие возможности вывода, совместимые с FigTree и другими просмотрщиками деревьев.
Список программного обеспечения для филогенетики
[ редактировать ]В эту таблицу включены некоторые наиболее распространенные филогенетические программы, используемые для вывода филогений в рамках байесовской модели. Некоторые из них не используют исключительно байесовские методы.
| Имя | Описание | Метод | Автор | Ссылка на сайт |
|---|---|---|---|---|
| г-н Байес | Филогенетический вывод | Программа для байесовского вывода и выбора моделей из широкого спектра филогенетических и эволюционных моделей. | Занг, Хюльзенбек, Дер Марк, Ронквист и Тесленко | https://nbisweden.github.io/MrBayes/ |
| ЗВЕРЬ | Деревья выборки байесовского эволюционного анализа | Байесовский вывод, расслабленные молекулярные часы, демографическая история | Эй Джей Драммонд, А. Рамбо и М. А. Сушард [34] | https://beast.community |
| ЗВЕРЬ 2 | Программная платформа для байесовского эволюционного анализа | Байесовский вывод, пакеты , множественные модели | Р. Букерт, Дж. Хелед, Д. Кюнерт, Т. Вон, Ч. Ву, Д. Се, М. А. Сушард, А. Рамбо, А. Дж. Драммонд. [35] | http://www.beast2.org |
| ФилоБайес / ФилоБайес MPI | Пробоотборник байесовской цепи Маркова Монте-Карло (MCMC) для филогенетической реконструкции. | Непараметрические методы моделирования межсайтовых вариаций склонностей нуклеотидов или аминокислот. | Н. Лартильо, Н. Родриг, Д. Стаббс, Дж. Ричер [36] | http://www.atgc-montpellier.fr/phylobayes/ |
| Бали-Фи | Одновременный байесовский вывод о выравнивании и филогении | Байесовский вывод, выравнивание, а также поиск по дереву | Сушард М.А., Ределингс Б.Д. [37] | http://www.bali-phy.org |
| БАКИ | Байесовское соответствие генных деревьев | Байесовское согласие с использованием модифицированного жадного консенсуса неукорененных квартетов | К. Ане , Б. Ларже, Д. А. Баум, С. Д. Смит, А. Рокас и Б. Ларже, С. К. Кота, Ч. Н. Дьюи, К. Ане [38] | http://www.stat.wisc.edu/~ane/bucky/ |
| БАТВИНГ | Байесовский анализ деревьев с генерацией внутренних узлов | Байесовский вывод, демографическая история, разделение населения | Ай Джей Уилсон, Д. Уил, Д. Болдинг [39] | http://www.maths.abdn.ac.uk/˜ijw [ постоянная мертвая ссылка ] |
| Байесовская филогения | Байесовский вывод деревьев с использованием методов Монте-Карло для цепей Маркова | Байесовский вывод, множественные модели, смешанная модель (автоматическое разделение) | М. Пейджел, А. Мид [40] | http://www.evolution.rdg.ac.uk/BayesPhy.html Архивировано 19 февраля 2020 г. в Wayback Machine. |
| Платформа рабочих процессов Armadillo | Платформа рабочих процессов, посвященная филогенетическому и общему биоинформатическому анализу | Обертка графического интерфейса вокруг MrBayes | Э. Лорд, М. Леклерк, А. Бок, А.Б. Диалло и В. Макаренков [41] | https://github.com/armadilloUQAM/armadillo2/ |
| Гениальный (плагин MrBayes) | Geneious предоставляет инструменты для исследования генома и протеома | Обертка графического интерфейса вокруг MrBayes | A. J. Drummond,M.Suchard,V.Lefort et al. | http://www.geneious.com |
| TOPALi | Филогенетический вывод | Обертка графического интерфейса вокруг MrBayes | И. Милн, Д. Линднер и др. [42] | http://www.topali.org |
Приложения
[ редактировать ]Байесовский вывод широко использовался молекулярными филогенетиками для множества приложений. Некоторые из них включают в себя:

- Вывод о филогениях. [43] [44]
- Вывод и оценка неопределенности филогений. [45]
- Вывод об эволюции состояния наследственного характера. [46] [47]
- Выводы о родовых ареалах. [48]
- Молекулярный датирующий анализ. [49] [50]
- Модельная динамика диверсификации и исчезновения видов [51]
- Выясните закономерности распространения возбудителей. [52]
- Вывод об эволюции фенотипических признаков. [53] [54]
Ссылки
[ редактировать ]- ^ Раннала, Брюс; Ян, Цзыхэн (сентябрь 1996 г.). «Распределение вероятностей молекулярных эволюционных деревьев: новый метод филогенетического вывода». Журнал молекулярной эволюции . 43 (3): 304–311. Бибкод : 1996JMolE..43..304R . дои : 10.1007/BF02338839 . ПМИД 8703097 . S2CID 8269826 .
- ^ Перейти обратно: а б Ян, З.; Раннала, Б. (1 июля 1997 г.). «Байесовский филогенетический вывод с использованием последовательностей ДНК: метод Монте-Карло цепи Маркова» . Молекулярная биология и эволюция . 14 (7): 717–724. doi : 10.1093/oxfordjournals.molbev.a025811 . ПМИД 9214744 .
- ^ Мау, Боб; Ньютон, Майкл А.; Ларге, Брет (март 1999 г.). «Байесовский филогенетический вывод с помощью методов Монте-Карло для цепей Маркова» . Биометрия . 55 (1): 1–12. дои : 10.1111/j.0006-341x.1999.00001.x . JSTOR 2533889 . ПМИД 11318142 . S2CID 932887 .
- ^ Ли, Шуйин; Перл, Деннис К.; Досс, Хани (июнь 2000 г.). «Построение филогенетического дерева с использованием цепи Маркова Монте-Карло» . Журнал Американской статистической ассоциации . 95 (450): 493–508. дои : 10.1080/01621459.2000.10474227 . JSTOR 2669394 . S2CID 122459537 .
- ^ Хюльзенбек, JP; Ронквист, Ф. (1 августа 2001 г.). «MRBAYES: Байесовский вывод филогенетических деревьев» . Биоинформатика . 17 (8): 754–755. дои : 10.1093/биоинформатика/17.8.754 . ПМИД 11524383 .
- ^ Лаплас П. (1774 г.). «Память о вероятности причин по событиям». Королевская академия наук . 6 :621–656. НАИД 10010866843 . английский перевод Стиглер С.М. (1986). «Воспоминания о вероятности причин событий» . Статистическая наука . 1 (3): 359–378. дои : 10.1214/ss/1177013620 .
- ^ Насименто, Фабрисия Ф.; Рейс, Марио дос; Ян, Цзыхэн (октябрь 2017 г.). «Руководство биолога по байесовскому филогенетическому анализу» . Экология и эволюция природы . 1 (10): 1446–1454. Бибкод : 2017NatEE...1.1446N . дои : 10.1038/s41559-017-0280-x . ПМК 5624502 . ПМИД 28983516 .
- ^ Гастингс В.К. (апрель 1970 г.). «Методы выборки Монте-Карло с использованием цепей Маркова и их приложения». Биометрика . 57 (1): 97–109. Бибкод : 1970Бимка..57...97H . дои : 10.1093/biomet/57.1.97 .
- ^ Перейти обратно: а б Метрополис Н., Розенблут А.В., Розенблут М.Н., Теллер А.Х., Теллер Э. (июнь 1953 г.). «Уравнение вычислений состояния быстрых вычислительных машин». Журнал химической физики . 21 (6): 1087–92. Бибкод : 1953ЖЧФ..21.1087М . дои : 10.1063/1.1699114 . ОСТИ 4390578 . S2CID 1046577 .
- ^ Фельзенштейн Дж (2004). Выводы о филогениях . Сандерленд, Массачусетс: Sinauer Associates.
- ^ Ян Цзы (2014). Молекулярная эволюция: статистический подход . Оксфорд, Англия: Издательство Оксфордского университета.
- ^ Гейер CJ (1991). «Цепь Маркова Монте-Карло с максимальной вероятностью». Керамидас Э.М., Кауфман С.М. (ред.). Информатика и статистика: материалы 23-го симпозиума по интерфейсу . Станция Фэрфакс: Фонд Интерфейса. стр. 156–163. ОСЛК 26603816 .
- ^ Ларгет Б., Саймон Д.Л. (июнь 1999 г.). «Алгоритмы Монте-Карло с цепью Маркова для байесовского анализа филогенетических деревьев» . Молекулярная биология и эволюция . 16 (6): 750–9. doi : 10.1093/oxfordjournals.molbev.a026160 .
- ^ Мау Б., Ньютон М.А., Ларжет Б. (март 1999 г.). «Байесовский филогенетический вывод с помощью методов Монте-Карло для цепей Маркова» . Биометрия . 55 (1): 1–12. дои : 10.1111/j.0006-341x.1999.00001.x . ПМИД 11318142 . S2CID 932887 .
- ^ Фельзенштейн Дж. (декабрь 1978 г.). «Случаи, в которых методы экономии или совместимости будут вводить в заблуждение». Систематическая зоология . 27 (4): 401–10. дои : 10.1093/sysbio/27.4.401 .
- ^ Касторани М.К., Рид Д.С., Раймонди П.Т., Альберто Ф., Белл Т.В., Кавано К.К. и др. (январь 2017 г.). «Колебания рождаемости населения приводят к изменениям в демографической связности и динамике метапопуляции» . Слушания. Биологические науки . 284 (1847): 20162086. doi : 10.1098/rspb.2016.2086 . ПМК 5310032 . ПМИД 28123088 .
- ^ О'Рейли Дж. Э., Путтик М. Н., Парри Л., Таннер А. Р., Тарвер Дж. Э., Флеминг Дж., Пизани Д., Донохью ПК (апрель 2016 г.). «Байесовские методы превосходят экономность, но за счет точности оценки филогении на основе дискретных морфологических данных» . Письма по биологии . 12 (4): 20160081. doi : 10.1098/rsbl.2016.0081 . ПМЦ 4881353 . ПМИД 27095266 .
- ^ Голобов П.А., Торрес А., Ариас Дж.С. (2018). «Взвешенная экономия превосходит другие методы филогенетического вывода в моделях, подходящих для морфологии» . Кладистика . 34 (4): 407–437. дои : 10.1111/cla.12205 . hdl : 11336/57822 . ISSN 0748-3007 . PMID 34649370 .
- ^ Китинг Дж.Н., Сэнсом Р.С., Саттон, доктор медицины, Найт К.Г., Гарвуд Р.Дж. (февраль 2020 г.). «Морфологическая филогенетика оценивается с использованием нового эволюционного моделирования» . Систематическая биология . 69 (5): 897–912. дои : 10.1093/sysbio/syaa012 . ПМК 7440746 . ПМИД 32073641 .
- ^ Суоффорд Д.Л., Олсен Г.Дж., Уодделл П.Дж., Хиллис Д.М. (1996). «Филогенетический вывод». В Hillis DM, Moritz C, Mable BK (ред.). Молекулярная систематика, 2-е издание . Сандерленд, Массачусетс: Синауэр. стр. 407–514. ISBN 9780878932825 .
- ^ Перейти обратно: а б Сузуки Ю., Глазко Г.В., Ней М. (декабрь 2002 г.). «Чрезмерная достоверность молекулярной филогении, полученной с помощью байесовской филогенетики» . Труды Национальной академии наук Соединенных Штатов Америки . 99 (25): 16138–43. Бибкод : 2002PNAS...9916138S . дои : 10.1073/pnas.212646199 . ПМЦ 138578 . ПМИД 12451182 .
- ^ Альфаро М.Э., Золлер С., Лутзони Ф. (февраль 2003 г.). «Байес или бутстрап? Моделирование, сравнивающее эффективность выборки Монте-Карло байесовской цепи Маркова и бутстрепа при оценке филогенетической достоверности» . Молекулярная биология и эволюция . 20 (2): 255–66. дои : 10.1093/molbev/msg028 . ПМИД 12598693 .
- ^ Дуади С.Дж., Дельсук Ф., Баучер Ю., Дулиттл В.Ф., Дузери Э.Дж. (февраль 2003 г.). «Сравнение байесовских мер филогенетической надежности и бутстраповских мер максимального правдоподобия» . Молекулярная биология и эволюция . 20 (2): 248–54. дои : 10.1093/molbev/msg042 . ПМИД 12598692 .
- ^ Гарсия-Сандовал Р. (январь 2014 г.). «Почему некоторые клады имеют низкие бутстреп-частоты и высокие байесовские апостериорные вероятности». Израильский журнал экологии и эволюции . 60 (1): 41–4. дои : 10.1080/15659801.2014.937900 .
- ^ Ян, З. (18 апреля 2007 г.). «Парадокс справедливого баланса, парадокс звездного дерева и байесовская филогенетика» . Молекулярная биология и эволюция . 24 (8): 1639–1655. дои : 10.1093/molbev/msm081 . ПМИД 17488737 .
- ^ Ян, Цзыхэн; Чжу, Тяньци (20 февраля 2018 г.). «Байесовский выбор неправильно определенных моделей является слишком самоуверенным и может вызвать ложные апостериорные вероятности для филогенетических деревьев» . Труды Национальной академии наук . 115 (8): 1854–1859. Бибкод : 2018PNAS..115.1854Y . дои : 10.1073/pnas.1712673115 . ПМЦ 5828583 . ПМИД 29432193 .
- ^ Эриксон П., Свеннблад Б., Бриттон Т., Оксельман Б. (октябрь 2003 г.). «Надежность байесовских апостериорных вероятностей и бутстреп-частот в филогенетике» . Систематическая биология . 52 (5): 665–73. дои : 10.1080/10635150390235485 . ПМИД 14530133 .
- ^ Хюльзенбек Дж. П., Ронквист Ф. (август 2001 г.). «MRBAYES: Байесовский вывод филогенетических деревьев» . Биоинформатика . 17 (8). Оксфорд, Англия: 754–5. дои : 10.1093/биоинформатика/17.8.754 . ПМИД 11524383 .
- ^ Мэддисон Д.Р., Суоффорд Д.Л., Мэддисон В.П. (декабрь 1997 г.). «NEXUS: расширяемый формат файла для систематической информации» . Систематическая биология . 46 (4): 590–621. дои : 10.1093/sysbio/46.4.590 . ПМИД 11975335 .
- ^ Джукс Т.Х., Кантор Ч.Р. (1969). Эволюция белковых молекул . Нью-Йорк: Академическая пресса. стр. 21–132.
- ^ Ян Цзы (ноябрь 1993 г.). «Оценка максимального правдоподобия филогении по последовательностям ДНК, когда скорости замен различаются в разных сайтах» . Молекулярная биология и эволюция . 10 (6): 1396–401. doi : 10.1093/oxfordjournals.molbev.a040082 . ПМИД 8277861 .
- ^ Ронквист Ф., Хюльзенбек Дж. П. (август 2003 г.). «MrBayes 3: Байесовский филогенетический вывод в рамках смешанных моделей» . Биоинформатика . 19 (12). Оксфорд, Англия: 1572–154. doi : 10.1093/биоинформатика/btg180 . ПМИД 12912839 .
- ^ Ронквист Ф., Тесленко М., ван дер Марк П., Эйрес Д.Л., Дарлинг А., Хёна С., Ларжет Б., Лю Л., Сушард М.А., Хюльзенбек Дж.П. (май 2012 г.). «MrBayes 3.2: эффективный байесовский филогенетический вывод и выбор модели в большом модельном пространстве» . Систематическая биология . 61 (3): 539–42. дои : 10.1093/sysbio/sys029 . ПМЦ 3329765 . ПМИД 22357727 .
- ^ Драммонд А.Дж., Сушард М.А., Се Д., Рамбо А. (август 2012 г.). «Байесовская филогенетика с BEAUti и BEAST 1.7» . Молекулярная биология и эволюция . 29 (8): 1969–73. дои : 10.1093/molbev/mss075 . ПМК 3408070 . ПМИД 22367748 .
- ^ Букерт Р., Хелед Дж., Кюнерт Д., Воган Т., Ву Ч., Се Д., Сушард М.А., Рамбо А., Драммонд А.Дж. (апрель 2014 г.). «BEAST 2: программная платформа для байесовского эволюционного анализа» . PLOS Вычислительная биология . 10 (4): e1003537. Бибкод : 2014PLSCB..10E3537B . дои : 10.1371/journal.pcbi.1003537 . ПМЦ 3985171 . ПМИД 24722319 .
- ^ Лартильо Н., Филипп Х (июнь 2004 г.). «Модель байесовской смеси для неоднородностей между сайтами в процессе замены аминокислот» . Молекулярная биология и эволюция . 21 (6): 1095–109. дои : 10.1093/molbev/msh112 . ПМИД 15014145 .
- ^ Сушард М.А., Ределингс Б.Д. (август 2006 г.). «BAli-Phy: одновременный байесовский вывод о выравнивании и филогении» . Биоинформатика . 22 (16): 2047–8. doi : 10.1093/биоинформатика/btl175 . ПМИД 16679334 .
- ^ Ане С. , Ларге Б., Баум Д.А., Смит С.Д., Рокас А. (февраль 2007 г.). «Байесовская оценка согласованности генных деревьев» . Молекулярная биология и эволюция . 24 (2): 412–26. дои : 10.1093/molbev/msl170 . ПМИД 17095535 .
- ^ Уилсон И.Дж., Уил М.Э., Балдинг-ди-джей (июнь 2003 г.). «Выводы на основе данных ДНК: истории популяций, эволюционные процессы и вероятности совпадения судебно-медицинской экспертизы» . Журнал Королевского статистического общества, серия A (Статистика в обществе) . 166 (2): 155–88. дои : 10.1111/1467-985X.00264 .
- ^ Пейгель М., Мид А. (июнь 2006 г.). «Байесовский анализ коррелированной эволюции дискретных символов с помощью цепи Маркова с обратимым скачком Монте-Карло». Американский натуралист . 167 (6): 808–25. дои : 10.1086/503444 . ПМИД 16685633 . S2CID 205984494 .
- ^ Лорд Э., Леклерк М., Бок А., Диалло А.Б., Макаренков В. (2012). «Armadillo 1.1: оригинальная рабочая платформа для проектирования и проведения филогенетического анализа и моделирования» . ПЛОС ОДИН . 7 (1): e29903. Бибкод : 2012PLoSO...729903L . дои : 10.1371/journal.pone.0029903 . ПМК 3256230 . ПМИД 22253821 .
- ^ Милн И., Линднер Д., Байер М., Хусмайер Д., Макгуайр Дж., Маршалл Д.Ф., Райт Ф. (январь 2009 г.). «TOPALi v2: богатый графический интерфейс для эволюционного анализа множественных согласований в кластерах HPC и многоядерных настольных компьютерах» . Биоинформатика . 25 (1): 126–7. doi : 10.1093/биоинформатика/btn575 . ПМЦ 2638937 . ПМИД 18984599 .
- ^ Алонсо Р., Кроуфорд А.Дж., Бермингем Э. (март 2012 г.). «Молекулярная филогения эндемической радиации кубинских жаб (Bufonidae: Peltophryne) на основе митохондриальных и ядерных генов». Журнал биогеографии . 39 (3): 434–51. Бибкод : 2012JBiog..39..434A . дои : 10.1111/j.1365-2699.2011.02594.x . S2CID 4152245 .
- ^ Антонелли А., Санмартин I (октябрь 2011 г.). «Массовое вымирание, постепенное похолодание или быстрая радиация? Реконструкция пространственно-временной эволюции древнего рода покрытосеменных Hedyosmum (Chloranthaceae) с использованием эмпирических и смоделированных подходов» . Систематическая биология . 60 (5): 596–615. дои : 10.1093/sysbio/syr062 . hdl : 10261/34829 . ПМИД 21856636 .
- ^ де Вилемрей П., Уэллс Дж. А., Эдвардс Р. Д., Бломберг С. П. (июнь 2012 г.). «Байесовские модели для сравнительного анализа, учитывающие филогенетическую неопределенность» . Эволюционная биология BMC . 12 (1): 102. Бибкод : 2012BMCEE..12..102В . дои : 10.1186/1471-2148-12-102 . ПМЦ 3582467 . ПМИД 22741602 .
- ^ Ронквист Ф. (сентябрь 2004 г.). «Байесовский вывод эволюции характера». Тенденции в экологии и эволюции . 19 (9): 475–81. дои : 10.1016/j.tree.2004.07.002 . ПМИД 16701310 .
- ^ Шеффер С., Коблмюллер С., Пфингстль Т., Штурмбауэр С., Криспер Г. (август 2010 г.). «Реконструкция предкового состояния выявляет множественную независимую эволюцию диагностических морфологических признаков у «Высших Орибатид» (Акари), что противоречит современным классификационным схемам» . Эволюционная биология BMC . 10 (1): 246. Бибкод : 2010BMCEE..10..246S . дои : 10.1186/1471-2148-10-246 . ПМК 2930640 . ПМИД 20701742 .
- ^ Филипович Н., Реннер СС (июль 2012 г.). «Брунфельсия (Пасленовые): род, поровну разделенный между Южной Америкой и радиацией на Кубе и других Антильских островах». Молекулярная филогенетика и эволюция . 64 (1): 1–11. дои : 10.1016/j.ympev.2012.02.026 . ПМИД 22425729 .
- ^ Бэкон CD, Бейкер WJ, член парламента Симмонса (май 2012 г.). «Рассеяние миоцена приводит к островной радиации в пальмовом трибе Trachycarpeae (Arecaceae)» . Систематическая биология . 61 (3): 426–42. дои : 10.1093/sysbio/syr123 . ПМИД 22223444 .
- ^ Саркинен Т., Бос Л., Олмстед Р.Г., Кнапп С. (сентябрь 2013 г.). «Филогенетическая основа эволюционного изучения пасленовых (Solanaceae): датированное дерево с 1000 кончиками» . Эволюционная биология BMC . 13 (1): 214. Бибкод : 2013BMCEE..13..214S . дои : 10.1186/1471-2148-13-214 . ПМЦ 3850475 . ПМИД 24283922 .
- ^ Сильвестро Д., Шницлер Дж., Лиоу Л.Х., Антонелли А., Саламин Н. (май 2014 г.). «Байесовская оценка видообразования и вымирания на основе неполных данных о местонахождении ископаемых» . Систематическая биология . 63 (3): 349–67. дои : 10.1093/sysbio/syu006 . ПМК 4361715 . ПМИД 24510972 .
- ^ Леми П., Рамбо А., Драммонд А.Дж., Сушард М.А. (сентябрь 2009 г.). «Байесовская филогеография находит свои корни» . PLOS Вычислительная биология . 5 (9): е1000520. Бибкод : 2009PLSCB...5E0520L . дои : 10.1371/journal.pcbi.1000520 . ПМК 2740835 . ПМИД 19779555 .
- ^ Сайбис Дж., Зиншаймер Дж., Бедфорд Т., Мазер А., Леми П., Сушард М.А. (2015). «Оценка фенотипической корреляции с помощью многомерной филогенетической модели скрытой ответственности» . Анналы прикладной статистики . 9 (2): 969–991. дои : 10.1214/15-AOAS821 . ISSN 1932-6157 . ПМК 4820077 . ПМИД 27053974 .
- ^ Толкофф М., Альфаро М., Бээле Г., Леми П., Сушард М.А. (2018). «Филогенетический факторный анализ» . Систематическая биология . 67 (3): 384–399. дои : 10.1093/sysbio/syx066 . ISSN 1063-5157 . ПМЦ 5920329 . ПМИД 28950376 .
Внешние ссылки
[ редактировать ]| Соответствующие поля | ||
|---|---|---|
| Основные понятия | ||
| Методы вывода | ||
| Текущие темы | ||
| Групповые черты | ||
| Типы групп | ||
| Номенклатура | ||