Зоопланктон
| Часть серии о |
| Планктон |
|---|
 |
Зоопланктон — это животный (или гетеротрофный ) компонент планктонного сообщества (приставка « зоо- » происходит от древнегреческого : ζῷον , латинизированного : zôion , букв. «животное»), которому для процветания приходится потреблять другие организмы. Планктон – это водные организмы, которые не способны эффективно плавать против течения. Следовательно, они дрейфуют или уносятся течениями в океане или течениями в морях , озерах и реках .
Зоопланктону можно противопоставить фитопланктон ( цианобактерии и микроводоросли ), которые являются растительным компонентом планктонного сообщества (приставка « фито- » происходит от древнегреческого: φῠτόν , латинизированного: phuton , букв. «растение», хотя таксономически это не так ). растения ). Зоопланктон гетеротрофен (питается другими), тогда как фитопланктон автотрофен (самопитается), часто генерируя биологическую энергию и макромолекулы посредством хлорофиллической фиксации углерода с использованием солнечного света - другими словами, зоопланктон не может производить собственную пищу, а фитопланктон может. В результате зоопланктон должен получать питательные вещества другими , питаясь организмами , такими как фитопланктон, которые обычно меньше зоопланктона. Большая часть зоопланктона микроскопична, но некоторые (например, медузы ) макроскопичны , то есть их можно увидеть невооруженным глазом . [1]
Многие простейшие (одноклеточные простейшие , питающиеся другой микроскопической жизнью) представляют собой зоопланктон, включая зоофлагелляты , фораминиферы , радиолярии , некоторые динофлагелляты и морские микроживотные . Макроскопический зоопланктон включает пелагических книдарий , гребневиков , моллюсков , членистоногих и оболочников , а также планктонных стрельчатых и щетинковых червей .
Различие между автотрофией и гетеротрофией часто теряется у очень маленьких организмов. Недавние исследования морского микропланктона показали, что более половины микроскопического планктона являются миксотрофами . Миксотроф — это организм, который может вести себя иногда как растение, а иногда как животное, используя сочетание автотрофности и гетеротрофии. Многие морские микрозоопланктоны являются миксотрофными, а это означает, что их также можно отнести к фитопланктонам.
Обзор
[ редактировать ]
Зоопланктон ( / ˈ z oʊ . ə p l æ ŋ k t ən / ; [2] / ˌ z oʊ . ə ˈ p l æ ŋ k t ən / ) [3] являются гетеротрофным (иногда детритофагом ) планктоном . Слово зоопланктон происходит от древнегреческого : ζῷον , латинизированного : zôion , букв. 'животное'; и πλᾰγκτός , planktós , 'странник; бродяга». [4]
Зоопланктон — это классификация, охватывающая целый ряд организмов различных размеров, включая мелких простейших и крупных многоклеточных животных . В него входят голопланктонные организмы, полный жизненный цикл которых проходит внутри планктона, а также меропланктонные организмы, которые проводят часть своей жизни в планктоне, прежде чем перейти либо к нектону , либо к сидячему , донный образ жизни. Хотя зоопланктон в основном переносится окружающими водными течениями, многие из них передвигаются , чтобы избежать хищников (как при дневной вертикальной миграции ) или увеличить частоту встреч с добычей.
Как любой вид может быть ограничен в пределах географического региона, так и зоопланктон. Однако виды зоопланктона не рассредоточены равномерно или беспорядочно в пределах региона океана. Как и в случае с фитопланктоном, «участки» видов зоопланктона существуют по всему океану. Хотя над мезопелагией существует мало физических барьеров , определенные виды зоопланктона строго ограничены градиентами солености и температуры, в то время как другие виды могут выдерживать большие градиенты температуры и солености. [5] На пятнистость зоопланктона также могут влиять биологические факторы, а также другие физические факторы. Биологические факторы включают размножение, хищничество, концентрацию фитопланктона и вертикальную миграцию. [5] Физическим фактором, который больше всего влияет на распределение зоопланктона, является перемешивание водной толщи ( апвеллинг и даунвеллинг вдоль побережья и в открытом океане), что влияет на доступность питательных веществ и, в свою очередь, на продукцию фитопланктона. [5]
Посредством потребления и переработки фитопланктона и других источников пищи зоопланктон играет роль в водных пищевых цепях , как ресурс для потребителей на более высоких трофических уровнях (включая рыбу), а также как канал для упаковки органического материала в биологический насос . Поскольку они обычно небольшие, зоопланктон может быстро реагировать на увеличение численности фитопланктона. [ нужны разъяснения ] например, во время весеннего цветения . Зоопланктон также является ключевым звеном в биомагнификации таких загрязнителей , как ртуть . [6]
- Типичные модели с участием зоопланктона
 Внизу слева: модели размерного спектра.Эти модели также имеют временные и пространственные компоненты. [7]
Внизу слева: модели размерного спектра.Эти модели также имеют временные и пространственные компоненты. [7]

К экологически важным группам простейших зоопланктона относятся фораминиферы , радиолярии и динофлагелляты (последние из них часто миксотрофны ). Важный многоклеточный зоопланктон включает книдарий, таких как медузы и португальский военный корабль ; ракообразные, такие как ветвистоусые , копеподы , остракоды , изоподы , амфиподы , мизиды и криль ; хетогнаты (черви-стрелы); моллюски, такие как крылоногие ; и хордовые, такие как сальпы и молодь рыб. Этот широкий филогенетический диапазон включает в себя столь же широкий диапазон пищевого поведения: фильтрационное питание , хищничество и симбиоз с автотрофным фитопланктоном , как это наблюдается у кораллов. Зоопланктон питается бактериопланктоном , фитопланктоном, другим зоопланктоном (иногда каннибалистически ), детритом (или морским снегом ) и даже нектонными организмами . В результате зоопланктон в основном встречается в поверхностных водах, где в изобилии имеются пищевые ресурсы (фитопланктон или другой зоопланктон).
Зоопланктон также может выступать в качестве болезней резервуара . Было обнаружено, что зоопланктон ракообразных является домом для бактерии Vibrio cholerae , вызывающей холеру , позволяя холерным вибрионам прикрепляться к своим хитиновым экзоскелетам . Эти симбиотические отношения повышают способность бактерий выживать в водной среде, поскольку экзоскелет обеспечивает бактерию углеродом и азотом. [8]
Классификация размеров
[ редактировать ]Размер тела был определен как «основной признак» планктона, поскольку это морфологическая характеристика, общая для организмов всех таксономий, которая характеризует функции, выполняемые организмами в экосистемах. [9] [10] Это оказывает огромное влияние на рост, воспроизводство, стратегии кормления и смертность. [11] Одно из старейших проявлений биогеографии признаков было предложено более 170 лет назад, а именно правило Бергмана , согласно которому полевые наблюдения показали, что более крупные виды имеют тенденцию встречаться в более высоких и холодных широтах. [12] [13]
В океанах размер имеет решающее значение для определения трофических связей в планктонных экосистемах и, таким образом, является решающим фактором в регулировании эффективности биологического углеродного насоса . [14] Размер тела чувствителен к изменению температуры из-за термической зависимости физиологических процессов. [15] Планктон в основном состоит из эктотермных организмов, которые не выделяют достаточного метаболического тепла для повышения температуры своего тела, поэтому их метаболические процессы зависят от внешней температуры. [16] Следовательно, эктотермные животные растут медленнее и достигают зрелости при большем размере тела в более холодных условиях, что долгое время озадачивало биологов, поскольку классические теории эволюции жизненного цикла предсказывают меньшие размеры взрослых особей в средах, задерживающих рост. [17] Эта закономерность изменения размеров тела, известная как правило размера-температуры (TSR), [18] наблюдался у широкого круга эктотермных животных, включая одноклеточные и многоклеточные виды, беспозвоночных и позвоночных. [17] [19] [13]
Процессы, лежащие в основе обратной зависимости между размером тела и температурой, еще предстоит определить. [17] Несмотря на то, что температура играет важную роль в формировании широтных изменений размеров организмов, эти закономерности могут также зависеть от сложных взаимодействий между физическими, химическими и биологическими факторами. Например, подача кислорода играет центральную роль в определении величины экзотермической реакции температуры на размер, но трудно отделить относительные эффекты кислорода и температуры от полевых данных, поскольку эти две переменные часто сильно взаимосвязаны в поверхностном океане. . [20] [21] [13]
Зоопланктон можно разделить на классы размеров. [22] которые разнообразны по своей морфологии, питанию, стратегиям кормления и т. д. как внутри классов, так и между классами:
| тип зоопланктона | диапазон размеров |
|---|---|
| пикозоопланктон | 2 мкм |
| нанозоопланктон | 2–20 мкм |
| микрозоопланктон | 20–200 мкм |
| мезозоопланктон | 0,2–20 миллиметров |

Микрозоопланктон
[ редактировать ]Микрозоопланктон определяют как гетеротрофный и миксотрофный планктон. В основном они состоят из фаготрофных простейших , включая инфузорий, динофлагеллят и мезозоопланктона науплиусов . [23] Микрозоопланктон является основным хищником планктонного сообщества. Микрозоопланктон, как основные потребители морского фитопланктона, ежедневно потребляет ~ 59–75% первичной продукции моря , что намного больше, чем мезозоопланктон. Тем не менее, макрозоопланктон иногда может иметь более высокие темпы потребления в эвтрофных экосистемах, поскольку там может доминировать более крупный фитопланктон. [24] [25] Микрозоопланктон также является ключевым регенератором питательных веществ, которые питают первичное производство и являются источниками пищи для многоклеточных животных. [25] [26]
Несмотря на свою экологическую значимость, микрозоопланктон остается недостаточно изученным. Регулярные океанографические наблюдения редко позволяют отслеживать биомассу микрозоопланктона или уровень растительноядности, хотя метод разбавления, элегантный метод измерения уровня растительноядности микрозоопланктона, разрабатывался уже почти четыре десятилетия (Landry and Hassett 1982). Число наблюдений за растительноядностью микрозоопланктона в мире составляет около 1600, [27] [28] гораздо меньше, чем у первичной продуктивности (>50 000). [29] Это затрудняет проверку и оптимизацию выпасной функции микрозоопланктона в моделях экосистем океана. [26]
Мезозоопланктон
[ редактировать ]Поскольку планктон редко вылавливается, утверждается, что численность и видовой состав мезопланктона можно использовать для изучения реакции морских экосистем на изменение климата. Это связано с тем, что их жизненный цикл обычно длится менее года, а это означает, что они реагируют на изменения климата между годами. Редкая ежемесячная выборка по-прежнему будет указывать на колебания. [30]
Таксономические группы
[ редактировать ]Простейшие
[ редактировать ]Простейшие — это протисты , которые питаются органическими веществами, такими как другие микроорганизмы или органические ткани и мусор. [31] [32] Исторически простейшие считались «одноклеточными животными», поскольку они часто обладают животным поведением, например, подвижностью и хищничеством , и лишены клеточной стенки , как это наблюдается у растений и многих водорослей . [33] [34] Хотя традиционная практика группирования простейших с животными больше не считается обоснованной, этот термин продолжает использоваться в широком смысле для обозначения одноклеточных организмов, которые могут передвигаться независимо и питаться путем гетеротрофии .
К морским простейшим относятся зоофлагелляты , фораминиферы , радиолярии и некоторые динофлагелляты .
Радиолярии
[ редактировать ]

Радиолярии — это одноклеточные хищные протисты , заключенные в сложные шаровидные раковины, обычно сделанные из кремнезема и пронизанные отверстиями. Их название происходит от латинского слова «радиус». Они ловят добычу, вытягивая части своего тела через отверстия. Как и в случае с кремнеземными панцирями диатомей, раковины радиолярий могут опускаться на дно океана, когда радиолярии умирают, и сохраняются как часть океанских отложений . Эти останки в виде микрокаменелостей предоставляют ценную информацию о прошлых условиях океана. [35]
 Как и диатомеи, радиолярии бывают разных форм.
Как и диатомеи, радиолярии бывают разных форм. Раковины радиолярий, как и диатомеи, обычно состоят из силиката.
Раковины радиолярий, как и диатомеи, обычно состоят из силиката. Однако акантарианские радиолярии имеют раковины из сульфата стронция. кристаллов
Однако акантарианские радиолярии имеют раковины из сульфата стронция. кристаллов Схематическая диаграмма сферической раковины радиолярий в разрезе
Схематическая диаграмма сферической раковины радиолярий в разрезе




| Внешние видео | |
|---|---|
Фораминиферы
[ редактировать ]Как и радиолярии, фораминиферы ( сокращенно форамы ) представляют собой одноклеточные хищные протисты, также защищенные раковинами с отверстиями. Их название происходит от латинского слова «носители дыр». Их раковины, часто называемые панцирями , имеют камеры (по мере роста у них появляются дополнительные камеры). Раковины обычно состоят из кальцита, но иногда состоят из склеенных частиц осадка или хитона и (редко) кремнезема. Большинство форамов донные, но около 40 видов — планктонные. [36] Они широко исследуются с использованием хорошо известных летописей окаменелостей, которые позволяют ученым делать много выводов о окружающей среде и климате прошлого. [35]


 разрез, показывающий камеры спирального отверстия
разрез, показывающий камеры спирального отверстия Живая аммиачная тепида, струящая гранулированную эктоплазму для улавливания пищи
Живая аммиачная тепида, струящая гранулированную эктоплазму для улавливания пищи Группа планктонных форм.
Группа планктонных форм.![Египетские пирамиды были построены из известняка, содержащего нуммулиты.[37]](//upload.wikimedia.org/wikipedia/commons/thumb/a/af/All_Gizah_Pyramids.jpg/325px-All_Gizah_Pyramids.jpg)



![Египетские пирамиды были построены из известняка, содержащего нуммулиты.[37]](http://upload.wikimedia.org/wikipedia/commons/thumb/a/af/All_Gizah_Pyramids.jpg/325px-All_Gizah_Pyramids.jpg)
| Внешние видео | |
|---|---|
Амеба
[ редактировать ]

 Эскиз обнаженной амебы, показывающий пищевые вакуоли и проглоченную диатомовую водоросль.
Эскиз обнаженной амебы, показывающий пищевые вакуоли и проглоченную диатомовую водоросль.
 Ксеногенная раковинная амеба, покрытая диатомовыми водорослями
Ксеногенная раковинная амеба, покрытая диатомовыми водорослями



Инфузории
[ редактировать ]
 голофиры Яйцо
голофиры Яйцо
 Эта инфузория переваривает цианобактерии . Рот находится внизу справа.
Эта инфузория переваривает цианобактерии . Рот находится внизу справа.




Динофлагелляты
[ редактировать ]Динофлагелляты — это тип одноклеточных жгутиконосцев , насчитывающий около 2000 морских видов. [38] Некоторые динофлагелляты являются хищными и, таким образом, принадлежат к сообществу зоопланктона. Их название происходит от греческого «динос», что означает вращение , и латинского «жгутик», означающего кнут или плеть . Это относится к двум хлыстообразным приспособлениям (жгутикам), используемым для движения вперед. Большинство динофлагеллят защищены красно-коричневой целлюлозной броней. Раскопки могут быть самой базальной линией жгутиковых. [39]


 Гиродиний , одна из немногих голых динофлагеллят, лишенных панциря.
Гиродиний , одна из немногих голых динофлагеллят, лишенных панциря. Динофлагеллята Protoperidinium выдвигает большую кормовую вуаль для захвата добычи.
Динофлагеллята Protoperidinium выдвигает большую кормовую вуаль для захвата добычи. Насселлярии радиолярии могут находиться в симбиозе с динофлагеллятами.
Насселлярии радиолярии могут находиться в симбиозе с динофлагеллятами.



Динофлагелляты часто живут в симбиозе с другими организмами. Многие насселлярий радиолярии динофлагеллят . симбионты содержат в своих раковинах [40] Насселлярия обеспечивает аммонием и углекислым газом , а динофлагеллята обеспечивает насселлярию слизистой оболочкой, полезной для охоты и защиты от вредных захватчиков. динофлагелляту [41] свидетельствует Анализ ДНК о том, что симбиоз динофлагеллят с радиоляриями развился независимо от других симбиозов динофлагеллят, например, с фораминиферами . [42]
 Tripos muelleri можно узнать по рогам U-образной формы.
Tripos muelleri можно узнать по рогам U-образной формы.
 Karenia brevis вызывает красные приливы, очень токсичные для человека [44]
Karenia brevis вызывает красные приливы, очень токсичные для человека [44]




Миксотрофы
[ редактировать ]Миксотроф — это организм, который может использовать смесь различных источников энергии и углерода вместо одного трофического режима в континууме от полной автотрофии на одном конце до гетеротрофии на другом. Подсчитано, что миксотрофы составляют более половины всего микроскопического планктона. [45] Существует два типа эукариотических миксотрофов: те, которые имеют собственные хлоропласты , и те, которые имеют эндосимбионты , а также другие, которые приобретают их посредством клептопластики или путем порабощения всей фототрофной клетки. [46]
Различие между растениями и животными часто нарушается у очень маленьких организмов. Возможные комбинации: фото- и хемотрофия , лито- и органотрофия , ауто- и гетеротрофия или другие их комбинации. Миксотрофы могут быть как эукариотами , так и прокариотами . [47] Они могут использовать различные условия окружающей среды. [48]
Многие морские микрозоопланктоны являются миксотрофными, а это означает, что их также можно отнести к фитопланктонам. Недавние исследования морского микрозоопланктона показали, что 30–45% численности инфузорий являются миксотрофными, а до 65% биомассы амебоидов, форамов и радиолярий - миксотрофными. [49]
Миксотрофный зоопланктон, сочетающий фототрофию и гетеротрофию – таблица на основе Stoecker et al., 2017. [50] | ||||||
|---|---|---|---|---|---|---|
| Описание | Пример | Дальнейшие примеры | ||||
| назвал их неконститутивными миксотрофами . Mitra et al., 2016 [51] Зоопланктон, обладающий фотосинтезом: микрозоопланктон или многоклеточный зоопланктон, приобретающий фототрофию за счет удержания хлоропластов. а или поддержание эндосимбионтов водорослей. | ||||||
| Генералисты | Протисты, сохраняющие хлоропласты и редко другие органеллы многих таксонов водорослей. |  | Большинство инфузорий - олиготрихов, сохраняющих пластиды. а | |||
| Специалисты | 1. Протисты, сохраняющие хлоропласты, а иногда и другие органеллы одного вида водорослей или очень близких видов водорослей. |  | Динофиз остроконечный | Динофизис виды. Мирионекта красная | ||
| 2. Протисты или зоопланктон с водорослевыми эндосимбионтами только одного вида водорослей или очень близких видов водорослей. |  | Ноктилука сцинтилланс | Метазоопланктон с эндосимбионтами водорослей Наиболее миксотрофные ризарии ( Acantharea , Polycystinea и Foraminifera ). Зеленая Noctiluca scintillans | |||
| а Удержание хлоропластов (или пластид) = секвестрация = порабощение. Некоторые виды, сохраняющие пластиды, также сохраняют другие органеллы и цитоплазму добычи. | ||||||
Виды Phaeocystis являются эндосимбионтами акантарианских радиолярий. [52] [53] Phaeocystis — важный род водорослей, встречающийся в составе морского фитопланктона по всему миру. Он имеет полиморфный жизненный цикл: от свободноживущих клеток до крупных колоний. [54] Он обладает способностью образовывать плавающие колонии, где сотни клеток встроены в гелевую матрицу, которая может значительно увеличиваться в размерах во время цветения . [55] В результате Phaeocystis вносит важный вклад в содержание морского углерода. [56] и циклы серы . [57]
- Миксопланктон
 Тинтинниды инфузории Фавелла
Тинтинниды инфузории Фавелла Euglena mutabilis — фотосинтезирующий жгутиконосец .
Euglena mutabilis — фотосинтезирующий жгутиконосец . Зоохлореллы (зеленые), живущие внутри инфузории Stichotricha secunda .
Зоохлореллы (зеленые), живущие внутри инфузории Stichotricha secunda . Динофлагеллята Dinophys acuta.
Динофлагеллята Dinophys acuta.






Ряд форумов миксотрофны. У них есть одноклеточные водоросли как эндосимбионты , принадлежащие к различным линиям, таким как зеленые водоросли , красные водоросли , золотые водоросли , диатомовые водоросли и динофлагелляты . [36] Миксотрофные фораминиферы особенно распространены в бедных питательными веществами океанических водах. [58] Некоторые отверстия являются клептопластическими , сохраняя хлоропласты из проглоченных водорослей для проведения фотосинтеза . [59]
По трофической направленности динофлагелляты встречаются повсеместно. Известно, что некоторые динофлагелляты фотосинтезируют , но большая часть из них на самом деле миксотрофны , сочетая фотосинтез с поеданием добычи ( фаготрофия ). [60] Некоторые виды являются эндосимбионтами морских животных и других протистов и играют важную роль в биологии коралловых рифов . Другие предшествуют другим простейшим, а некоторые формы являются паразитическими. Многие динофлагелляты миксотрофны и их также можно отнести к фитопланктону. Токсичная динофлагеллята Dinophys acuta приобретает хлоропласты от своей добычи. «Он не может поймать криптофиты сам по себе и вместо этого полагается на заглатывание инфузорий, таких как красный Myrionecta Rubra , которые изолируют свои хлоропласты от определенной клады криптофитов (Geminigera/Plagioselmis/Teleaulax)». [50]
Многоклеточные животные (животные)
[ редактировать ]
Свободноживущие виды класса ракообразных Copepoda обычно имеют длину от 1 до 2 мм и тело каплевидной формы. Как и у всех ракообразных, их тело разделено на три отдела: голову, грудь и брюшко с двумя парами усиков; первая пара часто длинная и выступающая. У них прочный экзоскелет из карбоната кальция и обычно один красный глаз в центре прозрачной головы. [61] Известно около 13 000 видов копепод, из них около 10 200 морских. [62] [63] Обычно они являются одними из наиболее доминирующих представителей зоопланктона. [64]
Помимо копепод, планктонные представители имеются также в классах ракообразных остракоды , жаберные моллюски и малакостраканы . Моллюски планктонны только на личиночной стадии. [65]
- Многоклеточный зоопланктон
 Веслоногие ракообразные с яйцами
Веслоногие ракообразные с яйцами








Голопланктон и меропланктон
[ редактировать ]Ихтиопланктон
[ редактировать ]Ихтиопланктон — это яйца и личинки рыб («ихтио» происходит от греческого слова « рыба »). Они являются планктонными, потому что не могут эффективно плавать своим ходом, а должны дрейфовать по океанским течениям. Икра рыб вообще не умеет плавать и однозначно является планктоном. Личинки ранних стадий плавают плохо, но личинки более поздних стадий плавают лучше и перестают быть планктонными по мере того, как вырастают в молодь рыб . Личинки рыб являются частью зоопланктона, который поедает более мелкий планктон, а икра рыб несет в себе собственный запас пищи. И яйца, и личинки поедаются более крупными животными. [66] [67]
 Молодые планктонные кальмары
Молодые планктонные кальмары Личинки морской солнечной рыбы (2,7 мм)
Личинки морской солнечной рыбы (2,7 мм) самцового рыбы Личинка
самцового рыбы Личинка



Желатиновый зоопланктон
[ редактировать ]студенистый зоопланктон включает гребневики , медузы , сальпы и Chaetognatha В прибрежных водах . Медузы — медленные пловцы, и большинство видов составляют часть планктона. Традиционно медузы рассматривались как трофические тупики, второстепенные участники морской пищевой цепи , студенистые организмы, строение тела которых в основном основано на воде, которая не представляет особой питательной ценности или интереса для других организмов, за исключением нескольких специализированных хищников, таких как океанская солнечная рыба и кожистая морская черепаха . [68] [69]
Недавно эта точка зрения была оспорена. Медузы и студенистый зоопланктон в целом, в который входят сальпы и гребневики , очень разнообразны, хрупкие, без твердых частей, их трудно увидеть и контролировать, они подвержены быстрым колебаниям численности и часто живут неудобно далеко от берега или глубоко в океане. Ученым сложно обнаружить и проанализировать медуз в кишках хищников, поскольку при поедании они превращаются в кашу и быстро перевариваются. [68] Но медузы цветут в огромных количествах, и было доказано, что они составляют основные компоненты рациона тунца , морской рыбы и рыбы-меч , а также различных птиц и беспозвоночных, таких как осьминоги , морские огурцы , крабы и амфиподы . [70] [69] «Несмотря на низкую энергетическую плотность, вклад медуз в энергетический бюджет хищников может быть намного больше, чем предполагалось, из-за быстрого пищеварения, низких затрат на отлов, доступности и избирательного питания более богатыми энергией компонентами. морские хищники, восприимчивые к проглатыванию пластика». [69] Согласно исследованию 2017 года, наркомедузы потребляют наибольшее разнообразие мезопелагических жертв, за ними следуют физонектовые сифонофоры , гребневики и головоногие моллюски . [71]

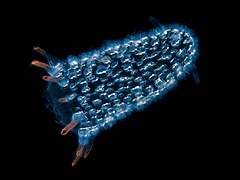 Эта свободно плавающая пиросома состоит из сотен отдельных биолюминесцентных оболочников .
Эта свободно плавающая пиросома состоит из сотен отдельных биолюминесцентных оболочников . Сальповая цепь
Сальповая цепь



Важность так называемой «желейной паутины» только начинает пониматься, но кажется, что медузы, гребневики и сифонофоры могут быть ключевыми хищниками в глубоких пелагических пищевых цепях с экологическим воздействием, аналогичным воздействию хищных рыб и кальмаров. Традиционно студенистые хищники считались неэффективными поставщиками морских трофических путей, но, по-видимому, они играют существенную и неотъемлемую роль в глубоких пелагических пищевых сетях . [71]
Роль в пищевых сетях
[ редактировать ]Выпас одноклеточного зоопланктона является причиной большей части органического углерода потерь в результате первичной морской продукции . [72] Однако выпас зоопланктона остается одной из ключевых неизвестных в глобальных прогнозных моделях потока углерода, структуры морской пищевой сети и характеристик экосистем, поскольку эмпирические измерения выпаса редки, что приводит к плохой параметризации функций выпаса. [73] [74] Чтобы преодолеть этот критический пробел в знаниях, было предложено сосредоточить усилия на разработке приборов, которые смогут связать изменения в биомассе или оптических свойствах фитопланктона с выпасом. [72]
Выпас скота является центральным процессом, определяющим темпы развития океанских экосистем и движущей силой морских биогеохимических циклов . [75] Во всех океанских экосистемах выпас гетеротрофных протистов является крупнейшим фактором потерь первичной морской продукции и изменяет распределение частиц по размерам. [76] Выпас влияет на все пути экспортного производства, делая выпас важным как для поверхностных, так и для углерода . глубинных процессов [77] Прогнозирование основных парадигм функционирования экосистем океана, включая реакцию на изменение окружающей среды, требует точного представления выпаса скота в глобальных биогеохимических, экосистемных и межбиомных моделях сравнения. [73] Несколько крупномасштабных анализов пришли к выводу, что потери фитопланктона, в которых преобладает выпас, являются предполагаемым объяснением годовых циклов биомассы фитопланктона, темпов накопления и экспортного производства. [78] [79] [74] [72]
- Пелагическая пищевая сеть
![Пелагическая пищевая сеть и биологический насос. Связи между биологическим насосом океана и пелагической пищевой сетью, а также возможность удаленного отбора проб этих компонентов с кораблей, спутников и автономных транспортных средств. Голубые воды представляют собой эвфотическую зону, а более темные голубые воды представляют собой сумеречную зону.[80]](//upload.wikimedia.org/wikipedia/commons/thumb/1/13/Export_Processes_in_the_Ocean_from_Remote_Sensing.jpg/627px-Export_Processes_in_the_Ocean_from_Remote_Sensing.jpg) Пелагическая пищевая сеть и биологический насос . Связи между биологическим насосом океана и пелагической пищевой сетью, а также возможность удаленного отбора проб этих компонентов с кораблей, спутников и автономных транспортных средств. Светло-голубые воды представляют собой эвфотическую зону , а более темные голубые воды представляют собой сумеречную зону . [80]
Пелагическая пищевая сеть и биологический насос . Связи между биологическим насосом океана и пелагической пищевой сетью, а также возможность удаленного отбора проб этих компонентов с кораблей, спутников и автономных транспортных средств. Светло-голубые воды представляют собой эвфотическую зону , а более темные голубые воды представляют собой сумеречную зону . [80]
![Пелагическая пищевая сеть и биологический насос. Связи между биологическим насосом океана и пелагической пищевой сетью, а также возможность удаленного отбора проб этих компонентов с кораблей, спутников и автономных транспортных средств. Голубые воды представляют собой эвфотическую зону, а более темные голубые воды представляют собой сумеречную зону.[80]](http://upload.wikimedia.org/wikipedia/commons/thumb/1/13/Export_Processes_in_the_Ocean_from_Remote_Sensing.jpg/627px-Export_Processes_in_the_Ocean_from_Remote_Sensing.jpg)

Роль в биогеохимии
[ редактировать ]Помимо связи первичных продуцентов с более высокими трофическими уровнями в морских пищевых цепях , зоопланктон также играет важную роль в качестве «переработчика» углерода и других питательных веществ, которые существенно влияют на морские биогеохимические циклы , включая биологический насос . Это особенно важно в олиготрофных водах открытого океана. В результате небрежного питания, выделения, выведения и выщелачивания фекальных гранул зоопланктон высвобождает растворенное органическое вещество (РОВ), которое контролирует круговорот РОВ и поддерживает микробный цикл . Эффективность поглощения, дыхание и размер добычи еще больше усложняют то, как зоопланктон способен преобразовывать и доставлять углерод в глубины океана . [76]
Небрежная подача и высвобождение DOM
[ редактировать ]
DOC = растворенный органический углерод
POC = твердый органический углерод
Адаптировано из Møller et al. (2005), [81]
Саба и др. (2009) [82] и Стейнберг и др. (2017). [76]
Экскреция и небрежное питание (физическое расщепление источника пищи) составляют 80% и 20% высвобождения РОВ ракообразными, опосредованного зоопланктоном, соответственно. [83] В том же исследовании было обнаружено, что выщелачивание фекальных гранул вносит незначительный вклад. У простейших, питающихся травоядными, РОВ выделяется в основном посредством экскреции и пищеварения, а студенистый зоопланктон также может выделять РОВ посредством выработки слизи. Выщелачивание фекальных гранул может продолжаться от нескольких часов до нескольких дней после первоначального выделения, и его последствия могут варьироваться в зависимости от концентрации и качества пищи. [84] [85] Различные факторы могут влиять на то, сколько РОВ высвобождается отдельными особями или популяциями зоопланктона. Эффективность поглощения (AE) — это доля пищи, поглощаемая планктоном, которая определяет, насколько доступны потребляемые органические материалы для удовлетворения необходимых физиологических потребностей. [76] В зависимости от скорости кормления и состава добычи, изменения AE могут привести к изменениям в производстве фекальных гранул и, таким образом, регулировать количество органического материала, перерабатываемого обратно в морскую среду. Низкая скорость подачи обычно приводит к высокой AE и получению мелких и плотных гранул, тогда как высокая скорость подачи обычно приводит к низкой AE и более крупным гранулам с большим содержанием органических веществ. Еще одним фактором, способствующим высвобождению РОВ, является частота дыхания. Физические факторы, такие как доступность кислорода, pH и условия освещенности, могут влиять на общее потребление кислорода и на то, сколько углерода теряется из зоопланктона в виде вдыхаемого CO 2 . Относительные размеры зоопланктона и добычи также влияют на то, сколько углерода выделяется при неаккуратном кормлении. Более мелкая добыча проглатывается целиком, тогда как более крупная добыча может питаться более «небрежно», то есть в результате неэффективного потребления выделяется больше биоматериала. [86] [87] Есть также доказательства того, что состав рациона может влиять на высвобождение питательных веществ: в рационе плотоядных животных выделяется больше растворенного органического углерода (DOC) и аммония, чем в рационе всеядных животных. [84]

Естественно обогащено железом

Высокое содержание питательных веществ, низкое содержание хлорофилла
Экспорт углерода
[ редактировать ]океана Зоопланктон играет решающую роль в поддержке биологического насоса посредством различных форм экспорта углерода , включая производство фекальных гранул, слизистых кормовых сетей, линьки и туш. По оценкам, фекальные гранулы вносят большой вклад в этот экспорт, причем ожидается, что размер веслоногих раков, а не их численность, будет определять, сколько углерода фактически достигнет дна океана. Важность фекальных гранул может варьироваться как в зависимости от времени, так и от места. Например, цветение зоопланктона может привести к образованию большего количества фекальных гранул, что приведет к увеличению экспорта углерода. Кроме того, когда фекальные гранулы тонут, они перерабатываются микробами в толще воды, что, таким образом, может изменить углеродный состав гранул. Это влияет на то, сколько углерода перерабатывается в эвфотической зоне и сколько достигает глубины. Вклад фекальных гранул в экспорт углерода, вероятно, недооценен; однако в настоящее время разрабатываются новые достижения в количественной оценке этого производства, включая использование изотопных характеристик аминокислот для определения того, сколько углерода экспортируется при производстве фекальных гранул зоопланктона. [89] Тушки также получают признание как важный источник экспорта углерода. Желепадение – массовое затопление студенистых туш зоопланктона – происходит по всему миру в результате сильного цветения. Ожидается, что из-за своего большого размера этот студенистый зоопланктон будет содержать большее количество углерода, что делает их тонущие туши потенциально важным источником пищи для донных организмов . [76]
См. также
[ редактировать ]- Учет морского зоопланктона
- Дильская вертикальная миграция
- Закисление океана
- Первичное производство
- Тонкие слои (океанография)
Ссылки
[ редактировать ]- ^ Сардет, Кристиан (5 июня 2015 г.). Планктон: чудеса дрейфующего мира . Издательство Чикагского университета. ISBN 978-0-226-26534-6 .
- ^ «зоопланктон» . Lexico Британский словарь английского языка . Издательство Оксфордского университета . Архивировано из оригинала 1 марта 2020 года.
- ^ «зоопланктон» . Словарь Merriam-Webster.com .
- ^ Турман, Х.В. (1997). Вводная океанография . Нью-Джерси, США: Колледж Прентис Холл. ISBN 978-0-13-262072-7 .
- ^ Перейти обратно: а б с Лалли, К.М. и Парсонс, Т.Р. (1993). Биологическая океанография. Введение . Берлингтон, Массачусетс: Elsevier. п. 314. ИСБН 978-0-7506-3384-0 .
- ^ «Как мы работаем в IISD-ELA: исследование Меркурия» . МИСР . 05.04.2017 . Проверено 6 июля 2020 г.
- ^ Эверетт, Дж. Д., Бэрд, М. Е., Бьюкенен, П., Балман, К., Дэвис, К., Дауни, Р., Гриффитс, К., Хенеган, Р., Клозер, Р. Дж., Лайоло, Л. и Лара- Лопес, А. (2017) «Моделирование того, что мы отбираем, и отбор проб, что мы моделируем: проблемы оценки модели зоопланктона». Границы морской науки , 4 : 77. два : 10.3389/fmars.2017.00077 .
 Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 . - ^ Джуд, бакалавр; Кирн, Ти Джей; Тейлор Р.К. (2005). «Фактор колонизации связывает выживание Vibrio cholerae в окружающей среде и заражение человека». Природа . 438 (7069): 863–6. Бибкод : 2005Natur.438..863K . дои : 10.1038/nature04249 . ПМИД 16341015 . S2CID 1964530 .
- ^ Личман, Елена; Оман, Марк Д.; Кьёрбо, Томас (2013). «Подходы к сообществам зоопланктона на основе признаков» . Журнал исследований планктона . 35 (3): 473–484. дои : 10.1093/plankt/fbt019 .
- ^ Кьёрбо, Томас; Херст, Эндрю Г. (2014). «Сдвиги в массовом масштабировании дыхания, питания и темпов роста при переходе форм жизни в морских пелагических организмах» . Американский натуралист . 183 (4): Е118–Е130. дои : 10.1086/675241 . ПМИД 24642502 . S2CID 15891709 .
- ^ Андерсен, К.Х.; Берге, Т.; Гонсалвес, Р.Дж.; Хартвиг, М.; Хойшель, Дж.; Хайландер, С.; Якобсен, Н.С.; Линдеманн, К.; Мартенс, Э.А.; Нойхаймер, AB; Олссон, К.; Палач, А.; Проу, АЭФ; Сэнмонт, Дж.; Трэвинг, С.Дж.; Виссер, AW; Вадхва, Н.; Кьёрбо, Т. (2016). «Характерные размеры жизни в океанах, от бактерий до китов» (PDF) . Ежегодный обзор морской науки . 8 : 217–241. Бибкод : 2016ARMS....8..217A . doi : 10.1146/annurev-marine-122414-034144 . hdl : 11336/52445 . ПМИД 26163011 .
- ^ Бергманн, Карл (1847). «О связи теплового хозяйства животных с их размерами» . Геттингенские исследования . 3 (1): 595–708.
- ^ Перейти обратно: а б с Брандао, Маноэла К.; и др. (2021). «Макромасштабные закономерности состава и размерной структуры океанического зоопланктона» . Научные отчеты . 11 (1): 15714. Бибкод : 2021NatSR..1115714B . дои : 10.1038/s41598-021-94615-5 . ПМЦ 8333327 . ПМИД 34344925 . Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Вудсон, К. Брок; Шрамски, Джон Р.; Джой, Саманта Б. (2018). «Объединяющая теория структуры экосистемы океана с тяжелыми верхушками» . Природные коммуникации . 9 (1): 23. Бибкод : 2018NatCo...9...23W . дои : 10.1038/s41467-017-02450-y . ПМК 5750233 . ПМИД 29295998 .
- ^ Браун, Джеймс Х.; Гиллули, Джеймс Ф.; Аллен, Эндрю П.; Сэвидж, Ван М.; Уэст, Джеффри Б. (2004). «К метаболической теории экологии». Экология . 85 (7): 1771–1789. Бибкод : 2004Ecol...85.1771B . дои : 10.1890/03-9000 .
- ^ Гарднер, Джанет Л.; Питерс, Энн; Кирни, Майкл Р.; Джозеф, Лев; Хейнсон, Роберт (2011). «Уменьшение размера тела: третий универсальный ответ на потепление?». Тенденции в экологии и эволюции . 26 (6): 285–291. Бибкод : 2011TEcoE..26..285G . дои : 10.1016/j.tree.2011.03.005 . ПМИД 21470708 .
- ^ Перейти обратно: а б с Анджилетта, MJ; Стейри, Т.Д.; Сирс, М.В. (2004). «Температура, скорость роста и размер тела у эктотермных животных: части головоломки истории жизни» . Интегративная и сравнительная биология . 44 (6): 498–509. дои : 10.1093/icb/44.6.498 . ПМИД 21676736 .
- ^ Аткинсон, Д. (1994). Температура и размер организма — биологический закон для эктотермных животных? . Достижения экологических исследований. Том. 25. стр. 1–58. дои : 10.1016/S0065-2504(08)60212-3 . ISBN 9780120139255 .
- ^ Аткинсон, Дэвид; Сибли, Ричард М. (1997). «Почему организмы обычно крупнее в более холодных условиях? Разбираемся в загадке истории жизни». Тенденции в экологии и эволюции . 12 (6): 235–239. Бибкод : 1997TEcoE..12..235A . дои : 10.1016/S0169-5347(97)01058-6 . ПМИД 21238056 .
- ^ Сунагава, С.; и др. (2015). «Структура и функции микробиома глобального океана» (PDF) . Наука . 348 (6237). дои : 10.1126/science.1261359 . hdl : 10261/117712 . ПМИД 25999513 . S2CID 206562917 .
- ^ Аудиёните, Аста; Барнеш, Диего Р.; Бодрон, Алан Р.; Белмейкер, Джонатан; Кларк, Тимоти Д.; Маршалл, К. Тара; Морронджиелло, Джон Р.; Ван Рейн, Итай (2019). «Является ли ограничение кислорода в нагревающихся водах действенным механизмом, объясняющим уменьшение размеров тела у водных эктотермных животных?» . Глобальная экология и биогеография . 28 (2): 64–77. Бибкод : 2019GloEB..28...64A . дои : 10.1111/geb.12847 . hdl : 10536/DRO/DU:30117155 . S2CID 92601781 .
- ^ Стейнберг, Дебора К.; Лэндри, Майкл Р. (3 января 2017 г.). «Зоопланктон и углеродный цикл океана» . Ежегодный обзор морской науки . 9 (1): 413–444. Бибкод : 2017ARMS....9..413S . doi : 10.1146/annurev-marine-010814-015924 . ISSN 1941-1405 . ПМИД 27814033 .
- ^ Зибурт, Джон МакН. ; Сметачек, Виктор; Ленц, Юрген (1978). «Структура пелагической экосистемы: гетеротрофные отделы планктона и их соотношение с размерными фракциями планктона 1» . Лимнология и океанография . 23 (6): 1256–1263. Бибкод : 1978LimOc..23.1256S . дои : 10.4319/lo.1978.23.6.1256 . S2CID 85568208 .
- ^ Кальбет, Альберт; Лэндри, Майкл Р. (2004). «Рост фитопланктона, выпас микрозоопланктона и круговорот углерода в морских системах». Лимнология и океанография . 49 (1): 51–57. Бибкод : 2004LimOc..49...51C . дои : 10.4319/lo.2004.49.1.0051 . hdl : 10261/134985 . S2CID 22995996 .
- ^ Перейти обратно: а б Кальбет, Альберт (2008). «Трофическая роль микрозоопланктона в морских системах» . Журнал морских наук ICES . 65 (3): 325–331. doi : 10.1093/icesjms/fsn013 .
- ^ Перейти обратно: а б Лю, Кайлин; Чен, Бинчжан; Чжэн, Липин; Су, Сухонг; Хуан, Банцинь; Чен, Мианрун; Лю, Хунбин (2021). «Что контролирует биомассу микрозоопланктона и уровень растительноядности в окраинных морях Китая?» . Лимнология и океанография . 66 (1): 61–75. Бибкод : 2021LimOc..66...61L . дои : 10.1002/lno.11588 . ISSN 0024-3590 . S2CID 224916151 . Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Чен, Бинчжан; Лэндри, Майкл Р.; Хуан, Банцинь; Лю, Хунбин (2012). «Усиливает ли потепление влияние выпаса микрозоопланктона на морской фитопланктон в океане?». Лимнология и океанография . 57 (2): 519–526. Бибкод : 2012LimOc..57..519C . дои : 10.4319/lo.2012.57.2.0519 .
- ^ Шмокер, Клэр; Эрнандес-Леон, Сантьяго; Кальбет, Альберт (2013). «Микрозоопланктон, выпас в океанах: последствия, изменчивость данных, пробелы в знаниях и будущие направления» . Журнал исследований планктона . 35 (4): 691–706. дои : 10.1093/plankt/fbt023 .
- ^ Бютенхейс, Эрик Т.; Хасиока, Такето; Кере, Корин Ле (2013). «Комбинированные ограничения на глобальную первичную продукцию океана с использованием наблюдений и моделей» . Глобальные биогеохимические циклы . 27 (3): 847–858. Бибкод : 2013GBioC..27..847B . дои : 10.1002/gbc.20074 . S2CID 140628035 .
- ^ Маккас, Дэвид Л.; Богранд, Грегори (10 февраля 2010 г.). «Сравнение временных рядов зоопланктона» . Журнал морских систем . Влияние изменчивости климата на морские экосистемы: сравнительный подход. 79 (3): 286–304. Бибкод : 2010JMS....79..286M . дои : 10.1016/j.jmarsys.2008.11.030 . ISSN 0924-7963 .
- ^ Панно, Джозеф (14 мая 2014 г.). Клетка: эволюция первого организма . Издательство информационной базы. ISBN 9780816067367 .
- ^ Бертран, Жан-Клод; Кометт, Пьер; Лебарон, Филипп; Мэтерон, Роберт; Норманд, Филипп; Симе-Нгандо, Телесфор (26 января 2015 г.). Экологическая микробиология: основы и приложения: Микробная экология . Спрингер. ISBN 9789401791182 .
- ^ Мэдиган, Майкл Т. (2012). Брок Биология микроорганизмов . Бенджамин Каммингс. ISBN 9780321649638 .
- ^ Йегер, Роберт Г. (1996). Простейшие: строение, классификация, рост и развитие . НКБИ. ISBN 9780963117212 . ПМИД 21413323 . Проверено 23 марта 2018 г.
- ^ Перейти обратно: а б Васильев, Мэгги (2006) «Планктон – животный планктон» , Те Ара – Энциклопедия Новой Зеландии . Доступ: 2 ноября 2019 г.
- ^ Перейти обратно: а б Хемлебен, К.; Андерсон, Орегон; Шпиндлер, М. (1989). Современные планктонные фораминиферы . Спрингер-Верлаг. ISBN 978-3-540-96815-3 .
- ^ Фораминиферы: История изучения , Университетский колледж Лондона . Проверено: 18 ноября 2019 г.
- ^ Гомес Ф (2012). «Контрольный список и классификация живых динофлагеллят (Dinoflagellata, Alveolata)» . СИКИМАР Океаниды . 27 (1): 65–140. doi : 10.37543/oceanides.v27i1.111 .
- ^ Доусон, Скотт С; Паредес, Александр Р. (2013). «Альтернативные цитоскелетные ландшафты: новизна цитоскелета и эволюция простейших с базальными выемками» . Современное мнение в области клеточной биологии . 25 (1): 134–141. дои : 10.1016/j.ceb.2012.11.005 . ПМЦ 4927265 . ПМИД 23312067 .
- ^ Болтовской, Деметрио; Андерсон, О. Роджер; Корреа, Нэнси М. (2017). Справочник протистов . Спрингер, Чам. стр. 731–763. дои : 10.1007/978-3-319-28149-0_19 . ISBN 9783319281476 .
- ^ Андерсон, Орегон (1983). Радиолярии . Springer Science & Business Media.
- ^ Гаст, Р.Дж.; Кэрон, Д.А. (1 ноября 1996 г.). «Молекулярная филогения симбиотических динофлагеллят планктонных фораминифер и радиолярий». Молекулярная биология и эволюция . 13 (9): 1192–1197. doi : 10.1093/oxfordjournals.molbev.a025684 . ISSN 0737-4038 . ПМИД 8896371 .
- ^ «Простейшие, поражающие жабры и кожу» . Ветеринарное руководство компании Merck . Архивировано из оригинала 3 марта 2016 года . Проверено 4 ноября 2019 г.
- ^ Брэнд, Ларри Э.; Кэмпбелл, Лиза; Бреснан, Эйлин (2012). « Карения : Биология и экология ядовитого рода» . Вредные водоросли . 14 : 156–178. Бибкод : 2012HAlga..14..156B . дои : 10.1016/j.hal.2011.10.020 . ПМЦ 9891709 . ПМИД 36733478 .
- ^ Коллинз, Ричард (14 ноября 2016 г.). «Остерегайтесь миксотрофов – они могут уничтожить целые экосистемы «за считанные часы» » . Ирландский эксперт .
- ^ Университет Суонси. «Микроскопические похитители тел наводнили наши океаны» . физ.орг .
- ^ Эйлер А. (декабрь 2006 г.). «Доказательства повсеместного распространения миксотрофных бактерий в верхних слоях океана: последствия и последствия» . Appl Environ Microbiol . 72 (12): 7431–7. Бибкод : 2006ApEnM..72.7431E . дои : 10.1128/АЕМ.01559-06 . ПМК 1694265 . ПМИД 17028233 .
- ^ Катечакис А., Стибор Х. (июль 2006 г.). «Миксотроф Ochromonas tuberculata может вторгаться и подавлять специализированные фаго- и фототрофные планктонные сообщества в зависимости от условий питания». Экология . 148 (4): 692–701. Бибкод : 2006Oecol.148..692K . дои : 10.1007/s00442-006-0413-4 . ПМИД 16568278 . S2CID 22837754 .
- ^ Лелес, С.Г.; Митра, А.; Флинн, К.Дж.; Стокер, ДК; Хансен, П.Дж.; Кальбет, А.; Макманус, Великобритания; Сандерс, RW; Кэрон, Д.А.; Нет, Ф.; Халлеграефф, генеральный директор (2017). «Океанические протисты с различными формами приобретенной фототрофии демонстрируют контрастную биогеографию и численность» . Труды Королевского общества B: Биологические науки . 284 (1860): 20170664. doi : 10.1098/rspb.2017.0664 . ПМЦ 5563798 . ПМИД 28768886 .
- ^ Перейти обратно: а б Стокер, ДК; Хансен, П.Дж.; Кэрон, Д.А.; Митра, А. (2017). «Миксотрофия морского планктона» (PDF) . Ежегодный обзор морской науки . 9 : 311–335. Бибкод : 2017ARMS....9..311S . doi : 10.1146/annurev-marine-010816-060617 . ПМИД 27483121 . S2CID 25579538 . Архивировано из оригинала (PDF) 27 февраля 2019 г.
- ^ Митра, А; Флинн, К.Дж.; Тильманн, Ю; Рэйвен, Дж; Кэрон, Д; и др. (2016). «Определение функциональных групп планктонных протистов по механизмам получения энергии и питательных веществ; включение разнообразных миксотрофных стратегий» . Протист . 167 (2): 106–20. дои : 10.1016/j.protis.2016.01.003 . hdl : 10261/131722 . ПМИД 26927496 .
- ^ Десель, Йохан; Саймон, Рафаэль; Галли, Марти; Варгас, Колумбан; Колен, Себастьян; Дедевиз, Ив; Биттнер, Люси; Проберт, Ян; Нет, Фабрис (30 октября 2012 г.). «Оригинальный способ симбиоза планктона открытого океана» . Труды Национальной академии наук . 109 (44): 18000–18005. Бибкод : 2012PNAS..10918000D . дои : 10.1073/pnas.1212303109 . ISSN 0027-8424 . ПМЦ 3497740 . ПМИД 23071304 .
- ^ Марс Брисбин, Маргарет; Гроссманн, Мэри М.; Месроп, Лиза Ю.; Митарай, Сатоши (2018). «Разнообразие симбионтов внутри хозяина и расширенное поддержание симбионтов в фотосимбиотических акантареях (клада F)» . Границы микробиологии . 9 : 1998. doi : 10.3389/fmicb.2018.01998 . ISSN 1664-302X . ПМК 6120437 . ПМИД 30210473 .
- ^ Шеманн, Вероника; Беквор, Сильви; Стефельс, Жаклин; Руссо, Вероника; Ланселот, Кристиана (1 января 2005 г.). «Цветение Phaeocystis в мировом океане и механизмы его контроля: обзор». Журнал морских исследований . Ресурсы железа и океанические питательные вещества – развитие глобального экологического моделирования. 53 (1–2): 43–66. Бибкод : 2005JSR....53...43S . CiteSeerX 10.1.1.319.9563 . дои : 10.1016/j.seares.2004.01.008 .
- ^ «Добро пожаловать на домашнюю страницу проекта секвенирования генома Phaeocystis antarctica» . Архивировано из оригинала 20 ноября 2015 г. Проверено 23 августа 2020 г.
- ^ ДиТуллио, Греция; Гребмайер, Дж. М .; Арриго, КР; Лизотт, член парламента; Робинсон, Д.Х.; Левентер, А.; Барри, JP; ВанВорт, ML; Данбар, РБ (2000). «Быстрый и ранний экспорт цветения Phaeocystis antarctica в море Росса, Антарктида». Природа . 404 (6778): 595–598. Бибкод : 2000Natur.404..595D . дои : 10.1038/35007061 . ПМИД 10766240 . S2CID 4409009 .
- ^ Дж, Стефельс; Л, Дейхуизен; WWC, Гискес (20 июля 1995 г.). «Активность DMSP-лиазы в весеннем цветении фитопланктона у голландского побережья, связанная с численностью Phaeocystis sp.» (PDF) . Серия «Прогресс в области морской экологии» . 123 : 235–243. Бибкод : 1995MEPS..123..235S . дои : 10.3354/meps123235 .
- ^ Маршалл, КК (11 ноября 2013 г.). Достижения микробной экологии . Springer Science & Business Media. ISBN 978-1-4684-7612-5 .
- ^ Бернхард, Дж. М.; Баузер, С.М. (1999). «Бентосные фораминиферы дизоксичных отложений: секвестрация хлоропластов и функциональная морфология». Обзоры наук о Земле . 46 (1): 149–165. Бибкод : 1999ESRv...46..149B . дои : 10.1016/S0012-8252(99)00017-3 .
- ^ Стокер Д.К. (1999). «Миксотрофия среди динофлагеллят». Журнал эукариотической микробиологии . 46 (4): 397–401. дои : 10.1111/j.1550-7408.1999.tb04619.x . S2CID 83885629 .
- ^ Роберт Д. Барнс (1982). Зоология беспозвоночных . Филадельфия, Пенсильвания : Холт-Сондерс Интернэшнл. стр. 683–692. ISBN 978-0-03-056747-6 .
- ^ «WoRMS — Всемирный реестр морских видов — Copepoda» . www.marinespecies.org . Архивировано из оригинала 30 июня 2019 г. Проверено 28 июня 2019 г.
- ^ Джефф А. Боксхолл; Даниэль Дефай (2008). «Глобальное разнообразие копепод (Crustacea: Copepoda) в пресной воде». Гидробиология . 595 (1): 195–207. дои : 10.1007/s10750-007-9014-4 . S2CID 31727589 .
- ^ Йоханнес Дюрбаум; Торстен Кюннеманн (5 ноября 1997 г.). «Биология копепод: Введение» . Ольденбургский университет Карла фон Осецкого . Архивировано из оригинала 26 мая 2010 года . Проверено 8 декабря 2009 г.
- ^ Трактат по зоологии - анатомия, таксономия, биология. Ракообразные
- ^ «Что такое ихтиопланктон?» . Юго-западный научный центр рыболовства . 3 сентября 2007 г. Архивировано из оригинала 18 февраля 2018 г. Проверено 22 июля 2011 г.
- ^ Аллен, доктор Ларри Г.; Хорн, доктор Майкл Х. (15 февраля 2006 г.). Экология морских рыб: Калифорния и прилегающие воды . Издательство Калифорнийского университета. стр. 269–319. ISBN 9780520932470 .
- ^ Перейти обратно: а б Гамильтон, Г. (2016) «Тайная жизнь медуз: медузы, долгое время считавшиеся второстепенными игроками в экологии океана, на самом деле являются важной частью морской пищевой сети» . Природа , 531 (7595): 432–435. дои : 10.1038/531432a
- ^ Перейти обратно: а б с Хейс, Г.К. , Дойл, Т.К. и Хоутон, Дж.Д. (2018) «Сдвиг парадигмы в трофической важности медуз?» Тенденции в экологии и эволюции , 33 (11): 874–884. дои : 10.1016/j.tree.2018.09.001
- ^ Кардона, Л., Де Кеведо, И.А., Боррелл, А. и Агилар, А. (2012) «Массовое потребление студенистого планктона высшими хищниками Средиземноморья». ПЛОС ОДИН , 7 (3): e31329. дои : 10.1371/journal.pone.0031329
- ^ Перейти обратно: а б Чой, Калифорния, Хэддок, С.Х. и Робисон, Б.Х. (2017) «Структура глубокой пелагической пищевой сети, выявленная наблюдениями за питанием in situ ». Труды Королевского общества B: Биологические науки , 284 (1868): 2017–2116. дои : 10.1098/rspb.2017.2116 . Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Перейти обратно: а б с д Менден-Дойер, Сюзанна ; Слэйд, Уэйн Гомер; Дирсен, Хайди (2021). «Содействие разработке инструментов для новых направлений исследований в науке об океане: открытие черного ящика выпаса» . Границы морской науки . 8 . дои : 10.3389/fmars.2021.695938 . Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Перейти обратно: а б Сток, Чарльз; Данн, Джон (2010). «Контроль за соотношением производства мезозоопланктона и первичной продукции в морских экосистемах». Глубоководные исследования. Часть I: Статьи океанографических исследований . 57 (1): 95–112. Бибкод : 2010DSRI...57...95S . дои : 10.1016/j.dsr.2009.10.006 .
- ^ Перейти обратно: а б Биссон, Келси; Сигел, Дэвид А.; Деврис, Тимоти (2020). «Диагностика механизмов экспорта углерода из океана в спутниковой модели пищевой сети» . Границы морской науки . 7 . дои : 10.3389/fmars.2020.00505 .
- ^ Уорден, Аризона; Следует, MJ; Джованнони, С.Дж.; Уилкен, С.; Циммерман, А.Е.; Килинг, Пи Джей (2015). «Переосмысление морского углеродного цикла: учет разнообразия образа жизни микробов» . Наука . 347 (6223). дои : 10.1126/science.1257594 . ПМИД 25678667 . S2CID 206560125 .
- ^ Перейти обратно: а б с д и Стейнберг, Дебора К.; Лэндри, Майкл Р. (2017). «Зоопланктон и углеродный цикл океана». Ежегодный обзор морской науки . 9 : 413–444. Бибкод : 2017ARMS....9..413S . doi : 10.1146/annurev-marine-010814-015924 . ПМИД 27814033 .
- ^ Мариани, Патрицио; Андерсен, Кен Х.; Виссер, Андре В.; Бартон, Эндрю Д.; Кьёрбо, Томас (2013). «Контроль сезонной смены планктона путем адаптивного выпаса» . Лимнология и океанография . 58 (1): 173–184. Бибкод : 2013LimOc..58..173M . дои : 10.4319/lo.2013.58.1.0173 .
- ^ Беренфельд, Майкл Дж. (2010). «Отказ от гипотезы критической глубины Свердрупа о цветении фитопланктона». Экология . 91 (4): 977–989. Бибкод : 2010Экол...91..977Б . дои : 10.1890/09-1207.1 . ПМИД 20462113 .
- ^ Миньо, А.; Феррари, Р.; Клаустр, Х. (2018). «Поплавки с биооптическими датчиками показывают, какие процессы вызывают цветение Северной Атлантики» . Природные коммуникации . 9 (1): 190. Бибкод : 2018NatCo...9..190M . дои : 10.1038/s41467-017-02143-6 . ПМК 5768750 . ПМИД 29335403 .
- ^ Сигел, Дэвид А.; Бюсселер, Кен О.; Беренфельд, Майкл Дж.; Бенитес-Нельсон, Клаудия Р.; Босс, Эммануэль; Бжезинский, Марк А.; Бурд, Адриан; Карлсон, Крейг А.; д'Асаро, Эрик А.; Дони, Скотт С.; Перри, Мэри Дж.; Стэнли, Рэйчел HR; Стейнберг, Дебора К. (2016). «Прогнозирование экспорта и судьбы чистой первичной продукции мирового океана: научный план экспорта» . Границы морской науки . 3 . дои : 10.3389/fmars.2016.00022 . Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Мёллер, Э.Ф.; Тор, П.; Нильсен, Т.Г. (2003). «Производство DOC Calanus finmarchicus , C. Glacialis и C. Hyperboreus в результате небрежного кормления и утечки фекальных гранул» . Серия «Прогресс в области морской экологии» . 262 : 185–191. Бибкод : 2003MEPS..262..185M . дои : 10.3354/meps262185 .
- ^ Саба, ГК; Стейнберг, ДК; Бронк, Д.А. (2009). «Влияние диеты на высвобождение растворенных органических и неорганических питательных веществ копеподом Acartiatonsa » . Серия «Прогресс в области морской экологии» . 386 : 147–161. Бибкод : 2009MEPS..386..147S . дои : 10.3354/meps08070 .
- ^ Саба, Грейс К.; Стейнберг, Дебора К.; Бронк, Дебора А. (2011). «Относительная важность небрежного кормления, выделения и выщелачивания фекальных шариков в выделении растворенного углерода и азота веслоногими ракообразными Acartiatonsa». Журнал экспериментальной морской биологии и экологии . 404 (1–2): 47–56. Бибкод : 2011JEMBE.404...47S . дои : 10.1016/j.jembe.2011.04.013 .
- ^ Перейти обратно: а б Тор, П.; Плотина, ХГ; Роджерс, ДР (2003). «Судьба органического углерода, выделяющегося из разлагающихся фекальных гранул веслоногих ракообразных, в связи с производством бактерий и эктоферментативной активностью» . Водная микробная экология . 33 : 279–288. дои : 10.3354/ame033279 .
- ^ Ханселл, Деннис А.; Карлсон, Крейг А. (2 октября 2014 г.). Биогеохимия морских растворенных органических веществ . Академическая пресса. ISBN 9780124071537 .
- ^ Моллер, Э.Ф. (2004). «Небрежное питание морских копепод: производство растворенного органического углерода в зависимости от размера добычи» . Журнал исследований планктона . 27 : 27–35. дои : 10.1093/plankt/fbh147 .
- ^ Мюллер, Ева Фриис (2007). «Производство растворенного органического углерода в результате неаккуратного питания копеподами Acartia tonsa, Centropages typicus и Temora longicornis» . Лимнология и океанография . 52 (1): 79–84. Бибкод : 2007LimOc..52...79M . дои : 10.4319/lo.2007.52.1.0079 .
- ^ Перейти обратно: а б с Халфтер, Свеня; Каван, Эмма Л.; Свадлинг, Керри М.; Эриксен, Рут С.; Бойд, Филип В. (2020). «Роль зоопланктона в установлении режимов экспорта углерода в Южном океане - сравнение двух репрезентативных тематических исследований в субантарктическом регионе» . Границы морской науки . 7 . дои : 10.3389/fmars.2020.567917 . S2CID 222003883 . Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Доэрти, С.; Маас, А.Е.; Стейнберг, ДК; Попп, Б.Н.; Клоуз, Х.Г. (2019). «Изотопный анализ фекальных гранул зоопланктона: понимание диетических и трофических процессов и характеристика фекальных гранул как конечного члена органического вещества». Американский геофизический союз, тезисы осеннего собрания 2019 г. аннотация № PP42C–12. Бибкод : 2019AGUFMPP42C..12D .
Внешние ссылки
[ редактировать ]- SAHFOS Фонд сэра Алистера Харди по наукам об океане
- Ocean Drifters Короткометражный фильм, рассказанный Дэвидом Аттенборо, о разнообразной роли планктона.
- Слайд-шоу Sea Drifters BBC Audio
- Plankton Chronicles Короткометражные документальные фильмы и фотографии
- COPEPOD: Глобальная база данных планктона . Глобальная база данных биомассы и численности зоопланктона.
- Путеводитель по морскому зоопланктону юго-восточной Австралии , Тасманский институт аквакультуры и рыболовства
- Австралийский проект непрерывного регистратора планктона, заархивировано 1 декабря 2008 г. в Wayback Machine.
- Основанный на изображениях ключ к зоопланктону Северной Америки



| Базы данных органов управления : Национальные |
|---|