Сифонофоры
| Сифонофоры | |
|---|---|
 | |
| (A) Rhizophysa eysenhardtii Масштабная линейка = 1 см, (B) Bathyphysa conifera 2 см, (C) Hippopodius hippopus 5 мм, (D) Kephyes hiulcus 2 мм (E) Desmophyes haematogaster 5 мм (F) Sphaeronectes christiansonae 2 мм, ( G) Praya dubia 40 м (130 футов), (H) Apolemia sp. 1 см, (I) Lychnagalma utricularia 1 см, (J) Nanomia sp. 1 см, (К) Physophora Hydrostatica 5 мм. | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Книдарийцы |
| Сорт: | Гидрозоа |
| Подкласс: | Гидроидолин |
| Заказ: | Сифонофоры Эшшольц , 1829 г. |
| Подзаказы [1] | |
| Синонимы | |
| |
Сифонофоры (от греческого siphōn «трубка» + pherein «нести»). [2] ) — порядок в составе Hydrozoa , который представляет собой класс морских организмов типа Cnidaria . По данным Всемирного реестра морских видов , отряд насчитывает 175 описанных на данный момент видов. [3]
Сифонофоры — высокополиморфные и сложные организмы. [4] Хотя они могут показаться отдельными организмами, на самом деле каждый экземпляр представляет собой колониальный организм, состоящий из медузоидных и полипоидных зооидов , которые морфологически и функционально специализированы. [5] Зооиды — это многоклеточные единицы, которые развиваются из одной оплодотворенной яйцеклетки и объединяются в функциональные колонии, способные размножаться, переваривать пищу, плавать, поддерживать положение тела и использовать реактивное движение для передвижения. [6] Большинство колоний — длинные, тонкие, прозрачные поплавки, обитающие в пелагиали . [7]
Как и другие гидрозойные , некоторые сифонофоры излучают свет, чтобы привлечь добычу и атаковать ее. В то время как многие морские животные производят синюю и зеленую биолюминесценцию , сифонофор из рода Erenna был лишь второй формой жизни, излучающей красный свет (первой была бесчешуйная рыба-дракон Chirostomias pliopterus ). [8] [9]
Анатомия и морфология
[ редактировать ]Характеристики колонии
[ редактировать ]Сифонофоры - это колониальные гидрозойные растения, которые не демонстрируют чередования поколений , а вместо этого размножаются бесполым путем путем почкования. [10] Зооиды — это многоклеточные единицы, которые строят колонии. Одиночная почка, называемая пропочкой, инициирует рост колонии путем деления. [7] Каждый зооид создается генетически идентичным; однако мутации могут изменить их функции и увеличить разнообразие зооидов внутри колонии. [7] Сифонофоры уникальны тем, что пропочка инициирует производство разнообразных зооидов с определенными функциями. [7] Функции и организация зооидов в колониях у разных видов сильно различаются; однако большинство колоний расположены двусторонне, дорсальной и вентральной сторонами стебля. [7] Стебель — это вертикальная ветвь в центре колонии, к которой прикрепляются зооиды. [7] Зооиды обычно выполняют особые функции и, таким образом, принимают определенные пространственные закономерности вдоль стебля. [7]
Общая морфология
[ редактировать ]Сифонофоры обычно демонстрируют один из трех стандартных планов тела, соответствующих подотрядам: Cystonectae , Physonectae и Calycophorae . [11] Цистонекты имеют длинный стебель с прикрепленными зооидами. [11] В каждой группе зооидов есть гастрозооид. [11] Гастрозооид имеет щупальца, используемые для захвата и переваривания пищи. [11] У групп также есть гонофоры, специализирующиеся на размножении. [11] Они используют пневматофор — газонаполненный поплавок на переднем конце и дрейфуют у поверхности воды или остаются на плаву в глубоком море. [11] У физонектов есть пневматофор и нектосома, в которых находятся нектофоры, используемые для реактивного движения. [11] Нектофоры перекачивают воду назад, чтобы двигаться вперед. [11] Каликофораны отличаются от цистонектов и фисонектов тем, что у них есть два нектофора и нет пневматофора. [11] Вместо этого у них часто есть заполненные маслом железы, которые, вероятно, помогают плавучести. [12]
Сифонофоры обладают несколькими типами зооидов. [13] Ученые определили две возможные эволюционные гипотезы для этого наблюдения: 1. С течением времени количество типов зооидов увеличилось. [13] 2. У последнего общего предка было много типов зооидов, и наблюдаемое сегодня разнообразие обусловлено утратой типов зооидов. [13] Исследования не выявили доказательств, подтверждающих первую гипотезу, но выявили некоторые доказательства в поддержку второй. [13]
Зооиды
[ редактировать ]- Сифонофоры могут иметь зооиды, представляющие собой полипы или медузы. [14] Однако зооиды уникальны и могут выполнять разные функции. [14]
Нектофоры
[ редактировать ]- Нектофоры — это медузы, которые способствуют приведению в движение некоторых сифонофоров в воде. [6] Они характерны для физонект и каликофоров. Нектофоры этих организмов расположены в нектосомах, где они могут координировать плавание колоний. [6] Также было замечено, что нектофоры работают вместе с репродуктивными структурами, обеспечивая движение во время отделения колонии. [6]
Прицветники
[ редактировать ]- Прицветники — это зооиды, уникальные для отряда сифонофоры. Они выполняют функцию защиты и поддержания нейтральной плавучести. [6] Однако прицветники имеются не у всех видов сифонофоров. [6]
Гастрозооиды
[ редактировать ]- Гастрозооиды — это полипы, у которых появилась функция питания сифонофоров. [13]
Пальпоны
[ редактировать ]- Пальпоны — это модифицированные гастрозооиды, которые участвуют в пищеварении, регулируя циркуляцию гастроваскулярной жидкости. [6]
Гонофоры
[ редактировать ]- Гонофоры – зооиды, участвующие в репродуктивных процессах сифонофоров. [6]
Пневматофоры
[ редактировать ]- Наличие пневматофоров характеризует подгруппы Cystonectae и Physonectae. [15] Это газонаполненные поплавки, расположенные у этих видов на переднем конце колоний. [6] Они помогают колониям сохранять ориентацию в воде. [6] В подгруппе цистонектов пневматофоры выполняют дополнительную функцию, помогая организмам плавать. [6] Сифонофоры, обладающие этой особенностью, развивают структуру на раннем этапе личиночного развития за счет инвагинаций уплощенной структуры планулы. [6] Дальнейшие наблюдения за видами сифонофоров Nanomia bijuga показывают, что функция пневматофора потенциально также функционирует, реагируя на изменения давления и регулируя хемотаксис у некоторых видов. [16]
Распространение и среда обитания
[ редактировать ]В настоящее время Всемирный реестр морских видов (WoRMS) идентифицирует 175 видов сифонофоров. [11] Они могут сильно различаться по размеру и форме, что во многом отражает среду, в которой они обитают. [11] Сифонофоры чаще всего являются пелагическими организмами, однако уровневые виды являются донными . [11] Меньшие тепловодные сифонофоры обычно обитают в эпипелагиали и используют свои щупальца для ловли зоопланктона и веслоногих ракообразных . [11] Сифонофоры большего размера живут в более глубоких водах, поскольку они обычно длиннее и хрупкие и должны избегать сильных течений. В основном они питаются более крупной добычей. [11] Большинство сифонофоров обитают в глубоководных районах и их можно встретить во всех океанах. [11] Виды сифонофоров редко обитают только в одном месте. [11] Однако некоторые виды могут обитать в определенном диапазоне глубин и/или районе океана. [11]
Поведение
[ редактировать ]Движение
[ редактировать ]Сифонофоры используют метод передвижения , аналогичный реактивному движению. Сифонофор представляет собой сложную совокупную колонию, состоящую из множества нектофоров, которые представляют собой клональные особи, образующиеся путем почкования и генетически идентичные. [17] В зависимости от того, где внутри сифонофора расположен каждый отдельный нектофор, их функции различаются. [17] Колониальное движение определяется отдельными нектофорами всех стадий развития. Более мелкие особи концентрируются к вершине сифонофора, их функция — поворот и корректировка ориентации колонии. [17] Чем старше они становятся, тем больше становятся особи. Более крупные особи располагаются у основания колонии, и их основная функция — толчковое движение. [17] Эти более крупные особи важны для достижения максимальной скорости колонии. [17] Каждый человек играет ключевую роль в движении совокупной колонии, и понимание их организации может позволить нам добиться успехов в наших собственных многореактивных двигательных установках. [17] Колониальная организация сифонофоров, особенно у Nanomia bijuga, дает эволюционные преимущества. [17] Большое количество сконцентрированных особей допускает резервирование. [17] Это означает, что даже если некоторые отдельные нектофоры становятся функционально нарушенными, их роль игнорируется, поэтому это не оказывает негативного влияния на колонию в целом. [17] Велум, тонкая полоска ткани, окружающая отверстие струи, также играет роль в способах плавания, что показано, в частности, в исследованиях, проведенных на ранее упомянутом виде N. bijuga. [18] Велюм становится меньше и более круглым во время движения вперед по сравнению с большим велумом, который наблюдается во время периодов наполнения. [18] Кроме того, положение паруса меняется в зависимости от поведения при плавании; велум изогнут вниз во время выброса струи, но во время наполнения велум перемещается обратно в нектофор. [18] Сифонофор Namonia bijuga также практикует дневную вертикальную миграцию , поскольку днем он остается на глубине моря, а ночью поднимается вверх. [17]
Хищничество и кормление
[ редактировать ]Сифонофоры — хищные хищники . [5] Их рацион состоит из множества веслоногих ракообразных, других мелких ракообразных и мелкой рыбы. [5] Как правило, рацион сильноплавающих сифонофоров состоит из более мелкой добычи, а рацион слабоплавающих сифонофоров состоит из более крупной добычи. [19] У большинства сифонофоров есть гастрозооиды с характерным щупальцем, прикрепленным к основанию зооида. Эта структурная особенность помогает организмам ловить добычу. [6] Виды с крупными гастрозооидами способны поедать добычу самых разных размеров. [19] Подобно многим другим организмам типа Cnidaria , многие виды сифонофоров имеют стрекательные капсулы нематоцист на ветвях своих щупалец, называемых тентиллами. [6] Нематоцисты расположены плотными батареями по бокам тентиллы. [6] Когда сифонофор встречает потенциальную добычу, их щупальца реагируют на то место, где щупальца размером 30–50 см (12–20 дюймов) создают сеть, изменяя свою форму вокруг добычи. [5] [20] [21] Нематоцисты затем стреляют миллионами [19] парализующих, а иногда и смертельных молекул токсина в захваченной добыче, которая затем переносится в нужное место для переваривания. [5] Некоторые виды сифонофоров используют агрессивную мимикрию, используя биолюминесцентный свет, поэтому добыча не может правильно идентифицировать хищника. [21]
В сифонофорной тентилле различают четыре типа нематоцист: гетеронемы, гаплонемы, десмонемы и ропалонемы. [21] Гетеронемы — самые крупные нематоцисты, представляющие собой шипы на стержне, расположенные рядом с канальцами, прикрепленными к центру сифонофора. [21] Гаплонемы имеют канальцы с открытыми кончиками и шипами, но без отчетливого стержня. [21] Это наиболее распространенная нематоциста среди сифонофоров. [21] У десмонем нет шипов, но вместо этого на трубочках есть клейкие свойства, позволяющие удерживать добычу. [21] Ропалонемы — нематоцисты с широкими трубочками для добычи. [21]
Из-за нехватки пищи в глубоководной среде большинство видов сифонофоров действуют по тактике «сиди и ожидай» в поисках еды. [22] Студенистое строение тела обеспечивает гибкость при ловле добычи, но студенистая адаптация зависит от среды обитания. [23] Они плавают, ожидая, пока их длинные щупальца наткнутся на добычу. Кроме того, сифонофоры из группы, обозначенной как Erenna, обладают способностью генерировать биолюминесценцию и красную флуоресценцию, в то время как их щупальцы подергиваются, имитируя движения мелких ракообразных и копепод. [8] Эти действия побуждают добычу приблизиться к сифонофору, позволяя ей поймать и переварить ее. [8]
Воспроизведение
[ редактировать ]Способы размножения сифонофоров различаются у разных видов, и по сей день некоторые способы остаются неизвестными. Обычно из одной зиготы начинается формирование колонии зооидов. [5] Оплодотворенная яйцеклетка созревает в простейшего, что запускает процесс почкования и создания нового зооида. [5] Этот процесс повторяется до тех пор, пока вокруг центрального стебля не образуется колония зооидов. [5] Напротив, некоторые виды размножаются с помощью полипов . Полипы могут содержать яйцеклетки и/или сперматозоиды и могут выделяться в воду из заднего конца сифонофора. [5] Полипы затем могут быть оплодотворены вне организма. [5]
Сифонофоры используют гонофоры для создания репродуктивных гамет . [13] Гонофоры бывают мужскими или женскими; однако типы гонофоров в колонии могут различаться у разных видов. [13] Виды характеризуются как однодомные или раздельнополые в зависимости от их гонофоров. [13] Однодомные виды содержат мужские и женские гонофоры в одной колонии зооидов, тогда как двудомные виды содержат мужские и женские гонофоры отдельно в разных колониях зооидов. [13]
Биолюминесценция
[ редактировать ]Почти все сифонофоры обладают биолюминесцентными способностями. Поскольку эти организмы чрезвычайно хрупкие, их редко можно увидеть живыми. [8] Считалось, что биолюминесценция сифонофоров развилась как защитный механизм. [8] сифонофоры глубоководного рода Erenna ( Считается, что находятся на глубинах 1600–2300 метров или 5200–7500 футов) также используют свою биолюминесцентную способность для нападения, в качестве приманки для привлечения рыбы. [8] Этот род — один из немногих, которые питаются рыбой, а не ракообразными. [8] Биолюминесцентные органы, называемые тентиллами , у этих незрячих особей излучают красную флуоресценцию вместе с ритмичным движением, что привлекает добычу, поскольку она напоминает более мелкие организмы, такие как зоопланктон и копеподы . Таким образом, был сделан вывод, что они используют свечение как приманку для привлечения добычи. [8] Некоторые исследования показывают, что глубоководные организмы не могут обнаруживать длинные волны, а красный свет имеет длину волны 680 нм. Если это так, то Эренна не приманивает рыбу , и должно быть другое объяснение. Однако глубоководные районы остаются в значительной степени неисследованными, и чувствительность к красному свету таких рыб, как циклотон и глубоководные рыбы -миктофиды . не следует сбрасывать со счетов [8]
Биолюминесцентные приманки встречаются во многих видах сифонофоров и используются по разным причинам. Такие виды, как Agalma okeni , Atorybia rosacea , Atorybia lucida , и Lychnafalma utricularia используют свои приманки как средство мимикрии для привлечения добычи. [11] A. rosacea имитирует личинки рыб, A. lucida , как полагают, имитирует домики личинок, а L. utricularia имитирует гидромедузу. [11] Вид Resomia ornicephala использует свои зелено-синие флуоресцирующие щупальца для привлечения криля, помогая им вытеснять другие организмы, охотящиеся за той же добычей. [11] Сифонофоры рода Erenna используют биолюминесцентные приманки, окруженные красной флуоресценцией, для привлечения добычи и, возможно, имитируют рыбу из рода Cyclotone . [11] Их жертва заманивается с помощью уникального движения, свойственного тентилле. [8] В молодом возрасте щупальцы организмов рода Erenna содержат только биолюминесцентную ткань, но по мере старения организма в этих тканях также присутствует красный флуоресцентный материал. [8]
Таксономия
[ редактировать ]
Организмы отряда Siphonophorae подразделяются на тип Cnidaria и класс Hydrozoa. [3] Филогенетические взаимоотношения сифонофоров представляют большой интерес в связи с высокой изменчивостью организации их колоний полипов и медуз. [24] [13] Когда-то считавшиеся отдельной группой, сходство личинок и морфологические особенности заставили исследователей поверить в то, что сифонофоры произошли от более простых колониальных гидрозоев, подобных таковым в отрядах Anthoathecata и Leptothecata . [15] Следовательно, теперь они объединены с ними в подкласс Hydroidolina .
Ранний анализ разделил сифонофоры на три основные подгруппы в зависимости от наличия или отсутствия двух разных признаков: плавательные колокольчики (нектофоры) и поплавки (пневматофоры). [15] Подгруппы состояли из Cystonectae, Physonectae и Calycorphores. У Cystonectae были пневматофоры, у Calycophores - нектофоры, а у Physonectae - и то, и другое. [15]
Эукариотический ядерный рибосомальный ген малой субъединицы 18S, эукариотический митохондриальный рибосомальный ген большой субъединицы 16S и анализ транскриптома дополнительно подтверждают филогенетическое разделение Siphonophorae на две основные клады: Cystonectae и Codonophora. Подотряды внутри Codonophora включают Physonectae (состоящие из клад Calycophorae и Euphysonectae), Pyrostephidae и Apolemiidae. [6] [13]
- Подотряд Calycophorae
- Абилиды Агассис, 1862 г.
- Клаусофииды Тоттон, 1965 г.
- Diphyidae Quoy & Gaimard, 1827 г.
- Hippopodiidae Кёлликер, 1853 г.
- Прайиды Кёлликер, 1853 г.
- Сфаэронектиды Хаксли, 1859 г.
- Tottonophyidae Pugh, Dunn & Haddock, 2018 г.
- Подотряд Cystonectae
- Физалии Брандт, 1835 г.
- Ризофизиды Брандт, 1835 г.
- Подотряд Фисонекты
- Агалматиды Брандт, 1834 г.
- Аполемиииды Хаксли, 1859 г.
- Кордагалматиды Пью, 2016 г.
- Эренниды Пью, 2001 г.
- Форскалииды Геккель, 1888 г.
- Physophoridae Эшшольц, 1829 г.
- Пиростефиды Мозер, 1925 г.
- Резомииды Пью, 2006 г.
- Родалииды Геккель, 1888 г.
- Стефаномииды Хаксли, 1859 г.
История
[ редактировать ]Открытие
[ редактировать ]Карл Линней описал первый сифонофор, португальский военный корабль , в 1758 году. [11] Скорость открытия видов сифонофоров в 18 веке была медленной, поскольку было обнаружено только четыре дополнительных вида. [11] В XIX веке благодаря исследовательским рейсам, проведенным европейскими державами, было обнаружено 56 новых видов. [11] Большинство новых видов, обнаруженных в этот период времени, были собраны в прибрежных, поверхностных водах. [11] Во время HMS Challenger экспедиции были собраны различные виды сифонофоров. Эрнст Геккель попытался провести описание всех видов сифонофоров, собранных в этой экспедиции. Он представил 46 «новых видов»; однако его работа подверглась резкой критике, поскольку в конечном итоге выяснилось, что некоторые из идентифицированных им видов не являются сифонофорами. [11] Тем не менее, некоторые его описания и рисунки (на фото ниже) современные биологи считают полезными. В течение 20-го века наблюдалась скорость открытия около 10 новых видов за десятилетие. [11] А. К. Тоттон, считающийся самым важным исследователем сифонофоров, в середине 20 века представил 23 новых вида сифонофоров. [11]
6 апреля 2020 года Институт океана Шмидта объявил об открытии гигантского сифонофора Apolemia в подводных каньонах недалеко от побережья Нингалу , диаметром 15 м (49 футов) и кольцом длиной примерно 47 м (154 фута), возможно, самого большого сифонофора, и самое длинное животное из когда-либо зарегистрированных. [25] [26]
нет Окаменелостей сифонофоров , хотя они развивались и адаптировались в течение длительного периода времени. Их тип, Cnidaria , представляет собой древнюю линию, восходящую к ок. 640 миллионов лет назад. [11]
Сифонофоры Геккеля
[ редактировать ]Эрнст Геккель описал многочисленные сифонофоры, и на нескольких пластинах из его «Kunstformen der Natur» (1904) изображены представители таксона : [27]
 Табличка 7
Табличка 7 Табличка 37
Табличка 37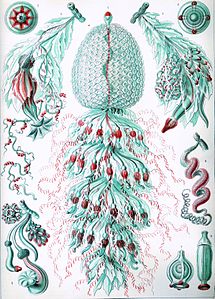 Табличка 59
Табличка 59 Табличка 77
Табличка 77




Ссылки
[ редактировать ]- ^ Шухерт, П. (2019). «Сифонофоры» . Мировая база данных гидрозоа . Получено 27 января 2019 г. - из Всемирного реестра морских видов.
- ^ «Сифонофора» . Лексико . Архивировано из оригинала 22 октября 2020 г. Проверено 10 марта 2020 г.
- ^ Перейти обратно: а б «Сифонофоры» . Всемирный регистр морских видов (2018). Проверено 8 января 2018 г.
- ^ Маки, ГО; Пью, PR; Перселл, Дж. Э. (1 января 1988 г.), Блакстер, JHS; Саутвард, А.Дж. (ред.), Биология сифонофоров , Достижения в морской биологии, том. 24, Academic Press, стр. 97–262, номер документа : 10.1016/s0065-2881(08)60074-7 , ISBN. 9780120261246 , получено 8 апреля 2023 г.
- ^ Перейти обратно: а б с д и ж г час я дж Тихий океан, Аквариум. «Пелагический сифонофор» . www.aquariumofpacific.org . Проверено 10 марта 2020 г.
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п Манро, Катриона; Зиберт, Стефан; Сапата, Фелипе; Хоуисон, Марк; Дамиан Серрано, Алехандро; Черч, Сэмюэл Х.; Гетц, Фрейя Э.; Пью, Филип Р.; Хэддок, Стивен HD; Данн, Кейси В. (20 января 2018 г.). «Улучшение филогенетического разрешения сифонофоры (Cnidaria) с последствиями для эволюции признаков» . биоRxiv . дои : 10.1101/251116 .
- ^ Перейти обратно: а б с д и ж г Данн, Кейси В. (декабрь 2005 г.). «Сложная организация на уровне колонии глубоководного сифонофора Bargmannia elongata (Cnidaria, Hydrozoa) направленно асимметрична и возникает в результате подразделения пропочек» . Динамика развития . 234 (4): 835–845. дои : 10.1002/dvdy.20483 . ПМИД 15986453 . S2CID 8644671 .
- ^ Перейти обратно: а б с д и ж г час я дж к Хэддок С.Х., Данн К.В., Пью П.Р., Шницлер К.Э. (июль 2005 г.). «Биолюминесцентные и красно-флуоресцентные приманки в глубоководном сифонофоре». Наука . 309 (5732): 263. CiteSeerX 10.1.1.384.7904 . дои : 10.1126/science.1110441 . ПМИД 16002609 . S2CID 29284690 .
- ^ «Сифонофоры | Смитсоновский океан» . Ocean.si.edu . Проверено 18 июня 2021 г.
- ^ Пью, Филип Р. (2014). «Сифонофора» . Доступ к науке . дои : 10.1036/1097-8542.625800 .
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п д р с т в v В х и С аа аб и Мапстоун, Джиллиан М. (06 февраля 2014 г.). «Глобальное разнообразие и обзор сифонофоров (Cnidaria: Hydrozoa)» . ПЛОС ОДИН . 9 (2): e87737. Бибкод : 2014PLoSO...987737M . дои : 10.1371/journal.pone.0087737 . ISSN 1932-6203 . ПМЦ 3916360 . ПМИД 24516560 .
- ^ Медуза: Естественная история
- ^ Перейти обратно: а б с д и ж г час я дж к Данн, Кейси В.; Пью, Филип Р.; Хэддок, Стивен HD (1 декабря 2005 г.). Нэйлор, Гэвин (ред.). «Молекулярная филогенетика сифонофоры (Cnidaria) с последствиями для эволюции функциональной специализации». Систематическая биология . 54 (6): 916–935. дои : 10.1080/10635150500354837 . ISSN 1076-836X . ПМИД 16338764 .
- ^ Перейти обратно: а б Данн, Кейси (2009). «Сифонофоры» . Современная биология . 19 (6): Р233-4. дои : 10.1016/j.cub.2009.02.009 . ПМИД 19321136 . Проверено 10 марта 2020 г.
- ^ Перейти обратно: а б с д Коллинз, Аллен Г. (30 апреля 2002 г.). «Филогения Medusozoa и эволюция жизненных циклов книдарий» . Журнал эволюционной биологии . 15 (3): 418–432. дои : 10.1046/j.1420-9101.2002.00403.x . S2CID 11108911 .
- ^ Черч, Сэмюэл Х.; Зиберт, Стефан; Бхаттачарья, Патикрит; Данн, Кейси В. (июль 2015 г.). «Гистология Nanomia bijuga (Hydrozoa: Siphonophora)» . Журнал экспериментальной зоологии, часть B: Молекулярная эволюция и эволюция развития . 324 (5): 435–449. дои : 10.1002/jez.b.22629 . ПМК 5032985 . ПМИД 26036693 .
- ^ Перейти обратно: а б с д и ж г час я дж Костелло, Джон Х.; Колин, Шон П.; Геммелл, Брэд Дж.; Дабири, Джон О.; Сазерленд, Келли Р. (ноябрь 2015 г.). «Многореактивное движение, организованное путем клонального развития в колониальном сифонофоре» . Природные коммуникации . 6 (1): 8158. Бибкод : 2015NatCo...6.8158C . дои : 10.1038/ncomms9158 . ISSN 2041-1723 . ПМЦ 4569723 . ПМИД 26327286 .
- ^ Перейти обратно: а б с Сазерленд, Келли Р.; Геммелл, Брэд Дж.; Колин, Шон П.; Костелло, Джон Х. (15 марта 2019 г.). «Принципы пропульсивной конструкции многоструйного сифонофора» . Журнал экспериментальной биологии . 222 (6): jeb198242. дои : 10.1242/jeb.198242 . hdl : 1912/23794 . ISSN 0022-0949 . ПМИД 30814298 . S2CID 73512609 .
- ^ Перейти обратно: а б с Цитирование: Перселл, Дженнифер Э. (1980). Влияние поведения сифонофоров на их естественный рацион: доказательства агрессивной мимикрии. Наука, том. 209, стр. 1045-1047. DOI: 10.1126/science.209.4460.1045.
- ^ Дамиан-Серрано, Алехандро; Хэддок, Стивен HD; Данн, Кейси В. (2 апреля 2020 г.). «Созданы, чтобы убивать: эволюция сифонофора-тентиллы для специализированного захвата добычи в открытом океане» . bioRxiv : 653345. doi : 10.1101/653345 . S2CID 215404157 .
- ^ Перейти обратно: а б с д и ж г час Дамиан-Серрано, Алехандро; Хэддок, Стивен HD; Данн, Кейси В. (12 июня 2019 г.). «Созданы, чтобы убивать: эволюция сифонофора-тентиллы для специализированного захвата добычи в открытом океане» . биоRxiv . дои : 10.1101/653345 . S2CID 215404157 .
- ^ Данн, Кейси (2005). «Сифонофоры» . Проверено 8 июля 2008 г.
- ^ Мадинанд, LP; Харбисон, GR (01 января 2001 г.), «Студенистый зоопланктон *» , в Стиле, Джоне Х. (ред.), Энциклопедия наук об океане (второе издание) , Оксфорд: Academic Press, стр. 9–19, doi : 10.1016/b978-012374473-9.00198-3 , ISBN 978-0-12-374473-9 , получено 31 октября 2020 г.
- ^ Ваггонер, Бен (21 июля 1995 г.). «Hydrozoa: Еще о морфологии» . Музей палеонтологии Калифорнийского университета . Проверено 10 марта 2020 г.
- ^ «Самое длинное гигантское жилистое морское существо, когда-либо зарегистрированное, похоже, обитает в космическом пространстве» . Интересный инжиниринг.com . 09.04.2020 . Проверено 10 апреля 2020 г.
- ^ Океанический институт Шмидта (9 апреля 2020 г.). «Новые виды обнаружены во время исследования глубоководных морских каньонов у Нингалу» . ЭврекАлерт! . Проверено 12 апреля 2020 г.
- ^ Константино, Грейс. «Формы искусства в природе: морские виды от Эрнста Геккеля» . Смитсоновский океан . Смитсоновский институт . Проверено 10 марта 2020 г.
Дальнейшее чтение
[ редактировать ]- Мапстоун, Джиллиан М. (2009). Сифонофора (Cnidaria, Hydrozoa) канадских вод Тихого океана . Оттава: NRC Research Press. ISBN 978-0-660-19843-9 .
- PinkTentacle.com (2008): Сифонофор: глубоководный суперорганизм (видео) . Проверено 23 мая 2009 г.
Внешние ссылки
[ редактировать ]- Данн, Кейси (nd). «Сифонофоры» . Современная биология . 19 (6). н/д: Р233-4. дои : 10.1016/j.cub.2009.02.009 . ПМИД 19321136 . Проверено 19 сентября 2014 г.
- Scubamedia.de (30 августа 2013 г.). «Таухен в Норвегии — Квас-фьорд» . Ютуб . scubamedia.de. Архивировано из оригинала 21 декабря 2021 г. Проверено 19 сентября 2014 г.
- Pinktentacle3 (22 декабря 2008 г.). «Сифонофор» . Ютуб . Архивировано из оригинала 21 декабря 2021 г. Проверено 19 сентября 2014 г.
{{cite web}}: CS1 maint: числовые имена: список авторов ( ссылка ) - «Потрясающее наблюдение за сифонофором» . Nautilus Live: исследуйте океан в прямом эфире вместе с доктором Робертом Баллардом и Исследовательским корпусом . Фонд исследования океана. 27 июня 2014 года . Проверено 18 сентября 2014 г.
- «Глубоководный сифонофор» (10 апреля 2017 г.) YouTube. Снимок сделан NOAA Okeanos Explorer 14 марта 2017 года на высоте 1560 метров к западу от комплекса Уинслоу Риф. Проверено 28 января 2018 г.