Медуза
| Медуза | |
|---|---|
 | |
| Крапива тихоокеанская морская ( Chrysaora fuscescens ) | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Книдарийцы |
| Подтип: | Медузозоа |
| Группы включены | |
| Кладистически включенные, но традиционно исключенные таксоны | |
Медузы , также известные как морские медузы , представляют собой фазу медузы некоторых студенистых представителей подтипа Medusozoa , который является основной частью типа Cnidaria .
Медузы — это в основном свободно плавающие морские животные с зонтиковидными колокольчиками и висячими щупальцами , хотя некоторые из них прикрепляются к морскому дну стеблями, а не передвигаются. Колокол может пульсировать, обеспечивая толчок для эффективного передвижения . Щупальца вооружены стрекательными клетками и могут использоваться для захвата добычи и защиты от хищников. Медузы имеют сложный жизненный цикл . Медуза обычно находится в половой фазе, в результате которой образуются планулы личинки . Затем они широко расходятся и вступают в фазу сидячих полипов , которая может включать бесполое почкование до достижения половой зрелости.
Медузы встречаются по всему миру, от поверхностных вод до морских глубин. Сцифозои («настоящие медузы») исключительно морские , но некоторые гидрозойные со схожим внешним видом обитают и в пресных водах . Большие, часто красочные медузы распространены в прибрежных зонах по всему миру. Медузы большинства видов быстро растут и созревают в течение нескольких месяцев, а затем умирают вскоре после размножения, но стадия полипа, прикрепленного к морскому дну, может быть гораздо более продолжительной. Медузы существуют уже по меньшей мере 500 миллионов лет. [1] и, возможно, 700 миллионов лет или больше, что делает их древнейшей группой животных с множеством органов. [2]
медуз едят люди В некоторых культурах . Их считают деликатесом в некоторых азиатских странах, где виды отряда прессуют Rhizostomeae и солят, чтобы удалить лишнюю воду. Австралийские исследователи описали их как «идеальную пищу»: экологически чистую и богатую белком, но с относительно низкой калорийностью . [3]
Они также используются в исследованиях в области клеточной и молекулярной биологии , особенно зеленый флуоресцентный белок, используемый некоторыми видами для биолюминесценции . Этот белок был адаптирован в качестве флуоресцентного репортера для встроенных генов и оказал большое влияние на флуоресцентную микроскопию .
Жалящие клетки, используемые медузами для подчинения своей добычи, могут нанести вред человеку. Ежегодно ужалены тысячи пловцов по всему миру, последствия которых варьируются от легкого дискомфорта до серьезных травм или даже смерти. При благоприятных условиях медузы могут образовывать огромные стаи, которые могут повреждать рыболовные снасти, заполняя рыболовные сети, а иногда и засорять системы охлаждения электростанций и опреснительных установок, которые забирают воду из моря.
Имена
Название «медуза», используемое с 1796 года, [4] традиционно применялся к медузам и всем подобным животным, включая гребневиков ( гребневики , другой тип). [5] [6] Термин «желе» или «морские медузы» появился совсем недавно: он был введен в общественные аквариумы, чтобы избежать использования слова «рыба» с его современным значением животного с позвоночником, хотя моллюски , каракатицы и морские звезды также не являются позвоночными. [7] [8] В научной литературе слова «желе» и «медуза» используются как синонимы. [9] [10] называют только сцифозоев . Во многих источниках «настоящими медузами» [11]
Группа медуз называется «привкус». [12] или «привкус». [13]
Картирование таксономических групп

Филогения
Определение
Термин «медуза» в целом соответствует медузам. [4] то есть стадия жизненного цикла у Medusozoa . Американский биолог-эволюционист Полин Картрайт дает следующее общее определение:
Обычно медузозойных книдарий наблюдается пелагическая медузы стадия хищной в жизненном цикле ; staurozoans являются исключением [поскольку они преследуются]. [14]
Словарь Мерриама-Вебстера определяет медузу следующим образом:
Свободно плавающая морская кишечнополостная , представляющая собой размножающуюся половым путем форму гидрозоя или сцифозоя, имеющая почти прозрачное тело в форме блюдца и растяжимые краевые щупальца, усеянные стрекательными клетками. [15]
Учитывая, что «медуза» — общее название, его соответствие биологическим группам неточно. Некоторые авторитеты называют гребневики [16] и некоторые сальпы [16] медузы, хотя другие авторитетные источники заявляют, что ни один из них не является медузой, и, по их мнению, это должно быть ограничено определенными группами внутри медузозоа. [17] [18]
Немедузозойные клады, называемые некоторыми, но не всеми авторитетами, медузами (в каждом случае приводятся как согласные, так и несогласные цитаты), обозначены знаком « ??? » на следующей кладограмме животного царства:
| животное |
| ||||||||||||||||||



Медузозойная медуза
Медузы не являются кладой , поскольку они включают в себя большую часть Медузозоа, за исключением некоторых Гидрозоа. [19] [20] Группы медузозойных, включенные авторитетными источниками, обозначены на следующем филогенетическом дереве наличием цитат. Названия включенных медуз (по возможности на английском языке) выделены жирным шрифтом; Наличие названного и цитируемого примера указывает на то, что по крайней мере этот вид в его группе назывался медузой.
| Книдарийцы |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||







Таксономия
Подтип Medusozoa включает всех книдарий, находящихся на стадии медузы в жизненном цикле. Основной цикл — яйцо, личинка планулы , полип, медуза, причем медуза является половой стадией. Стадия полипа иногда утрачивается вторично. Подтип включает основные таксоны: Scyphozoa (большие медузы), Cubozoa (коробчатые медузы) и Hydrozoa (маленькие медузы), за исключением Anthozoa (кораллы и морские анемоны). [25] Это позволяет предположить, что форма медузы развилась после полипов. [26] Медузозойные животные обладают тетрамерной симметрией, с частями, состоящими из четырех или кратных четырем. [25]
Четыре основных класса медузозойных Cnidaria:
- Сцифозоа иногда называют настоящими медузами, хотя они не более настоящие медузы, чем другие, перечисленные здесь. Они обладают тетрарадиальной симметрией. У большинства из них есть щупальца по внешнему краю чашеобразного колокола и длинные ротовые ручки вокруг рта в центре субзонтика. [25]
- Кубозоа (коробчатая медуза) имеют (округленный) коробчатый колокол, а велариум помогает им плавать быстрее. Кубомедузы могут быть более тесно связаны с медузами-сцифозами, чем с гидрозоа. [26]
- Hydrozoa medusae также обладает тетрарадиальной симметрией, почти всегда имеет велум (диафрагму, используемую при плавании), прикрепленную сразу к краю колокола, не имеет ротовых рук, но имеет гораздо меньшую центральную стебельчатую структуру, рукоятку, с конечным ротовым отверстием. , и отличаются отсутствием клеток в мезоглее. Hydrozoa демонстрируют большое разнообразие образа жизни; некоторые виды сохраняют форму полипа на протяжении всей своей жизни и вообще не образуют медуз (например, гидра , которая, следовательно, не считается медузой), а некоторые являются полностью медузальными и не имеют формы полипа. [25]
- Staurozoa (медузы со стебельками) характеризуются формой медузы, которая обычно сидячая, ориентирована вверх ногами и со стеблем, выходящим из вершины «чашечки» (колокола), который прикрепляется к субстрату. По крайней мере, у некоторых Staurozoa также есть форма полипа, которая чередуется с медузоидной частью жизненного цикла. До недавнего времени Staurozoa относили к Scyphozoa. [25]
Существует более 200 видов Scyphozoa, около 50 видов Staurozoa, около 50 видов Cubozoa, а Hydrozoa включает около 1000–1500 видов, образующих медуз, но гораздо больше видов, которые этого не делают. [27] [28]
Ископаемая история



Поскольку у медуз нет твердых частей, окаменелости встречаются редко. Самая старая однозначная окаменелость свободно плавающей медузы - это Burgessomedusa из среднекембрийских сланцев Берджесс в Канаде, которая, вероятно, представляет собой либо стволовую группу кубомедуз (Cubozoa), либо Acraspeda (клада, включающая Staurozoa, Cubozoa и Scyphozoa). Другие заявленные находки из кембрия Китая и Юты в Соединенных Штатах сомнительны и, возможно, вместо этого представляют собой гребневики . [29]
Анатомия

Главной особенностью настоящей медузы является зонтиковидный колокол. Это полая структура, состоящая из массы прозрачного желеобразного вещества, известного как мезоглея , образующего гидростатический скелет животного. [25] Мезоглея на 95% и более состоит из воды. [30] а также содержит коллаген и другие волокнистые белки, а также блуждающие амебоциты , которые могут поглощать мусор и бактерии. Мезоглоея ограничена эпидермисом снаружи и гастродермой изнутри. Край колокола часто делится на закругленные лепестки, известные как выступы , которые позволяют колоколу изгибаться. В промежутках или нишах между клапанами свисают рудиментарные органы чувств, известные как ропалии , а на краю колокольчика часто имеются щупальца. [25]

На нижней стороне колокола находится рукоятка, стеблеобразная структура, свисающая из центра, на кончике которой находится рот, который также выполняет функцию ануса. К рукоятке часто присоединяются четыре ротовых рукава, стекающие в воду внизу. [31] Рот открывается в гастроваскулярную полость , где происходит пищеварение и всасывание питательных веществ. Он разделен четырьмя толстыми перегородками на центральный желудок и четыре желудочных кармана. Четыре пары гонад прикреплены к перегородкам, а рядом с ними четыре перегородочные воронки открываются наружу, возможно, обеспечивая хорошее снабжение гонад кислородом. Около свободных краев перегородок желудочные нити впадают в полость желудка; они вооружены нематоцистами и клетками, продуцирующими ферменты, и играют роль в подчинении и переваривании добычи. У некоторых сцифозоев полость желудка соединена с радиальными каналами, которые широко разветвляются и могут присоединяться к краевому кольцевому каналу. Реснички в этих каналах циркулируют жидкость в правильном направлении. [25]
Кубомедуза во многом схожа по строению. У него квадратный колокольчик в форме коробки. С каждого из четырех нижних углов свисает короткая педаль или стебель. К каждой педали прикреплены одно или несколько длинных тонких щупалец. [32] Край колокола загнут внутрь, образуя полку, известную как велариум, которая ограничивает отверстие колокола и создает мощную струю, когда колокол пульсирует, позволяя коробчатым медузам плавать быстрее, чем настоящим медузам. [25] Гидрозои также похожи, обычно имеют всего четыре щупальца на краю колокола, хотя многие гидрозои являются колониальными и могут не иметь свободноживущей медузальной стадии. У некоторых видов образуется неотделяющийся бутон, известный как гонофор , который содержит гонаду, но лишен многих других медузальных особенностей, таких как щупальца и ропалии. [25] Стебельковые медузы прикреплены к твердой поверхности базальным диском и напоминают полип, ротовой конец которого частично развился в медузу с лопастями, несущими щупальца, и центральной рукояткой с четырехгранным ртом. [25]
Большинство медуз не имеют специализированных систем осморегуляции , дыхания и кровообращения , а также не имеют центральной нервной системы . Нематоцисты, доставляющие жало, расположены преимущественно на щупальцах; У настоящих медуз они также есть вокруг рта и желудка. [33] Медузам не нужна дыхательная система, поскольку через эпидермис диффундирует достаточное количество кислорода. Они ограниченно контролируют свои движения, но могут ориентироваться с помощью пульсаций колоколообразного тела; некоторые виды большую часть времени являются активными пловцами, тогда как другие в основном дрейфуют. [34] Ропалии содержат рудиментарные органы чувств, которые способны обнаруживать свет, вибрации воды, запах и ориентацию. [25] рыхлая сеть нервов, называемая « нервной сетью расположена В эпидермисе » . [35] [36] Хотя традиционно считается, что у них нет центральной нервной системы , концентрацию нервных сетей и ганглиоподобные структуры можно считать составляющими ее у большинства видов. [37] Медуза обнаруживает раздражители и передает импульсы как по всей нервной сети, так и вокруг кольцевого нервного кольца другим нервным клеткам. Ропалиальные ганглии содержат пейсмекерные нейроны, которые контролируют скорость и направление плавания. [25]
У многих видов медуз ропалии включают глазки — светочувствительные органы , способные отличать свет от тьмы. Обычно это глазки с пигментными пятнами, некоторые клетки которых пигментированы. Ропалии подвешены на стеблях с тяжелыми кристаллами на одном конце, действуя как гироскопы, направляя взгляд в небо. Некоторые медузы смотрят вверх на полог мангровых зарослей, совершая ежедневную миграцию из мангровых болот в открытую лагуну, где они кормятся, и обратно. [2]
У кубомедуз более развитое зрение, чем у других групп. У каждого человека есть 24 глаза, два из которых способны различать цвета, и четыре параллельные области обработки информации, которые действуют в конкуренции. [38] предположительно, это делает их одними из немногих видов животных, которые имеют обзор окружающей среды на 360 градусов. [39]
Глаз коробчатой медузы
Изучение эволюции глаз медуз является промежуточным звеном в лучшем понимании того, как развивались зрительные системы на Земле. [40] Медузы демонстрируют огромные различия в зрительных системах: от участков фоторецепторных клеток, наблюдаемых в простых фоторецепторных системах, до более сложных глаз, наблюдаемых у кубомедуз. [40] Основные темы исследований зрительной системы медуз (с акцентом на кубомедуз) включают: эволюцию зрения медуз от простых к сложным зрительным системам), морфологию глаз и молекулярную структуру кубомедуз (включая сравнение с глазами позвоночных), а также различные варианты использования. видения, включая поведение, ориентированное на выполнение задач, и нишевую специализацию.
Эволюция
Экспериментальные данные о светочувствительности и фоторецепции у книдарий появились еще в середине 1900-х годов, и с тех пор обширный объем исследований посвящен эволюции зрительных систем медуз. [41] Зрительные системы медуз варьируются от простых фоторецепторных клеток до сложных глаз, формирующих изображения. Более древние зрительные системы включают экстраокулярное зрение (зрение без глаз), которое включает в себя многочисленные рецепторы, отвечающие за однофункциональное поведение. Более производные зрительные системы включают восприятие, способное к множеству действий, ориентированных на выполнение задач.
Хотя у книдарий отсутствует настоящий мозг, они имеют «кольцевую» нервную систему , которая играет значительную роль в двигательной и сенсорной активности. Эта сеть нервов отвечает за сокращение и движение мышц и приводит к появлению светочувствительных структур. [40] У Cnidaria существуют большие различия в системах, лежащих в основе светочувствительности. Светочувствительные структуры варьируются от неспециализированных групп клеток до более «обычных» глаз, похожих на глаза позвоночных . [41] Общие эволюционные шаги по развитию сложного зрения включают (от более предкового к более производным состояниям): ненаправленную фоторецепцию, направленную фоторецепцию, зрение с низким разрешением и зрение с высоким разрешением. [40] Увеличение сложности среды обитания и задач способствовало развитию зрительных систем высокого разрешения, характерных для производных книдарий, таких как кубомедузы . [40]
Базальные зрительные системы, наблюдаемые у различных книдарий, демонстрируют фоточувствительность, характерную для одной задачи или поведения. Экстраокулярная фоторецепция (форма ненаправленной фоторецепции) является наиболее основной формой светочувствительности и определяет разнообразие поведения книдарий. Он может регулировать циркадный ритм (как это видно у безглазых гидрозоев ) и другие светоориентированные модели поведения, реагирующие на интенсивность и спектр света. Экстраокулярная фоторецепция может функционировать дополнительно при положительном фототаксисе (у личинок планулы гидрозойных), [41] а также во избежание вредного количества УФ-излучения посредством отрицательного фототаксиса . Направленная фоторецепция (способность воспринимать направление падающего света) обеспечивает более сложные фототаксические реакции на свет и, вероятно, развивается посредством укладки мембран . [40] Результирующие поведенческие реакции могут варьироваться от управляемого нереста, приуроченного к лунному свету, до теневых реакций, направленных на избегание потенциальных хищников. [41] [42] Светоориентированное поведение наблюдается у многих сцифозоев, в том числе у обыкновенной лунной медузы Aurelia aurita , которая мигрирует в ответ на изменения окружающего освещения и положения Солнца, хотя у них нет собственных глаз. [41]
Зрительная система кубомедуз с низким разрешением является более развитой, чем направленная фоторецепция, и, таким образом, зрение коробчатых медуз представляет собой самую базовую форму истинного зрения, при которой несколько направленных фоторецепторов объединяются для создания первого изображения и пространственного разрешения . Это отличается от зрения с высоким разрешением, которое наблюдается в камере или сложных глазах позвоночных и головоногих моллюсков , которые полагаются на фокусирующую оптику . [41] Крайне важно, что зрительные системы коробчатых медуз отвечают за управление несколькими задачами или поведением, в отличие от менее развитых зрительных систем других медуз, которые управляют отдельными поведенческими функциями. Это поведение включает в себя фототаксис, основанный на солнечном свете (положительный) или тенях (отрицательный), избегании препятствий и контроле частоты плавательного пульса. [43]
Кубомедузы обладают «правильными глазами» (похожими на позвоночных), которые позволяют им обитать в такой среде, которую не могут сделать медузы более низкого уровня. Фактически, они считаются единственным классом в кладе Medusozoa , поведение которого требует пространственного разрешения и подлинного зрения. [41] Однако хрусталик в их глазах функционально более похож на чашеобразные глаза, наблюдаемые у организмов с низким разрешением, и практически не обладает способностью к фокусировке. [44] [43] Отсутствие способности фокусироваться связано с тем, что фокусное расстояние превышает расстояние до сетчатки , что приводит к созданию несфокусированных изображений и ограничению пространственного разрешения. [41] Зрительная система кубомедузы по-прежнему достаточна для создания изображения, которое помогает решать такие задачи, как избегание объектов.
Полезность как модельный организм
Глаза кубомедузы представляют собой сложную во многих отношениях зрительную систему. Эти сложности включают в себя значительные различия в морфологии глаз кубомедуз (включая их задачи и характеристики поведения), а также молекулярный состав их глаз, включая фоторецепторы, опсины , линзы и синапсы . [41] Сравнение этих атрибутов с более производными зрительными системами может позволить лучше понять, как могла происходить эволюция более производных зрительных систем, и дает представление о том, как коробчатая медуза может играть роль модели эволюции/развития для всех зрительных систем. . [45]
Характеристики
Зрительные системы кубомедуз одновременно разнообразны и сложны и включают в себя несколько фотосистем . [41] Вероятно, существуют значительные различия в визуальных свойствах между видами кубомедуз, учитывая значительные межвидовые морфологические и физиологические различия. Глаза, как правило, различаются по размеру и форме, а также количеству рецепторов (включая опсины ) и физиологии у разных видов кубомедуз. [41]
У кубомедуз есть ряд сложных глаз с линзами, которые похожи на глаза более продвинутых многоклеточных организмов, таких как позвоночные. Их 24 глаза относятся к четырем различным морфологическим категориям. [46] Эти категории состоят из двух больших, морфологически различных медиальных глаз (нижний и верхний глаз с линзами), содержащих сферические линзы, латеральной пары глаз с пигментными щелями и латеральной пары глаз с пигментными ямками. [43] Глаза расположены на ропалиях (небольших сенсорных структурах), выполняющих сенсорные функции кубомедузы, и выходят из полостей эксумбреллы (поверхности тела) на боковой стороне колокольчиков медузы. [41] Два больших глаза расположены на средней линии булавы и считаются сложными, поскольку содержат линзы. Четыре оставшихся глаза лежат по бокам по обе стороны каждой ропалии и считаются простыми. Простые глаза представляют собой небольшие вдавленные чашечки эпителия с развитой пигментацией . [47] Более крупный из сложных глаз содержит клеточную роговицу, состоящую из монореснитчатого эпителия, клеточный хрусталик, гомогенную капсулу хрусталика, стекловидное тело с призматическими элементами и сетчатку из пигментированных клеток. Меньший из сложных глаз считается немного менее сложным, поскольку у него нет капсулы, но в остальном он имеет ту же структуру, что и больший глаз. [47]
У кубомедуз есть несколько фотосистем, включающих разные пары глаз. [41] Доказательства включают иммуноцитохимические и молекулярные данные, которые показывают различия в фотопигментах между различными морфологическими типами глаз, а также физиологические эксперименты, проведенные на кубомедузах, чтобы предположить поведенческие различия между фотосистемами. Каждый отдельный тип глаз представляет собой фотосистему, которая работает коллективно, контролируя визуально управляемое поведение. [41]
Глаза кубомедузы в основном используют c-PRC (реснитчатые фоторецепторные клетки), аналогичные глазам позвоночных. Эти клетки подвергаются каскадам фототрансдукции (процесс поглощения света фоторецепторами), которые запускаются к-опсинами. [48] Доступные последовательности опсинов позволяют предположить, что существует два типа опсинов, которыми обладают все книдарии, включая древний филогенетический опсин и сестринский цилиарный опсин из группы к-опсинов. Кубомедузы могут иметь как ресничные, так и книдопсы (книдарийные опсины), которые ранее не считались возможными в одной и той же сетчатке. [41] Тем не менее, не совсем очевидно, обладают ли книдарии множественными опсинами, способными обладать отличительной спектральной чувствительностью . [41]
Сравнение с другими организмами
Сравнительные исследования генетического и молекулярного состава глаз коробчатых медуз по сравнению с более производными глазами, наблюдаемыми у позвоночных и головоногих моллюсков, сосредоточены на: хрусталике и кристаллинов составе , синапсах и генах Pax , а также их предполагаемых доказательствах наличия общих изначальных (предковых) генов в эволюции глаз. [49]
Говорят, что глаза кубомедузы представляют собой эволюционную модель всех глаз, основанную на эволюционном наборе кристаллинов и генов Pax. [45] Исследования, проведенные на коробчатых медузах, в том числе Tripedalia Cysophora, показали, что они обладают единственным геном Pax, PaxB. PaxB функционирует путем связывания с промоторами кристаллинов и их активации. Гибридизация PaxB in situ привела к экспрессии PaxB в хрусталике, сетчатке и статоцистах . [45] Эти результаты и отказ от предыдущей гипотезы о том, что Pax6 был предковым геном Pax в глазах, привели к выводу, что PaxB был изначальным геном в эволюции глаз и что глаза всех организмов, вероятно, имели общего предка. [45]
Структура линз кубомедузы очень похожа на структуру хрусталика других организмов, но кристаллины отличаются как по функциям, так и по внешнему виду. [49] Слабые реакции наблюдались в сыворотке, и наблюдалось очень слабое сходство последовательностей кристаллинов хрусталиков позвоночных и беспозвоночных. [49] Вероятно, это связано с различиями в белках с более низкой молекулярной массой и последующим отсутствием иммунологических реакций с антисыворотками , которые проявляются в линзах других организмов. [49]
Все четыре зрительные системы видов кубомедуз, подробно исследованных ( Carybdea marsupialis , Chiropsalmus Quadrumanus , Tamoya haplonema и Tripedalia Cysophora ), имеют инвагинированные синапсы, но только в верхних и нижних линзах глаза. Различная плотность была обнаружена между верхними и нижними линзами, а также между видами. [46] В ропалиях были обнаружены четыре типа химических синапсов, которые могут помочь в понимании нейронной организации, в том числе: четкие однонаправленные, однонаправленные с плотным ядром, четкие двунаправленные и двунаправленные с четким и плотным ядром. Синапсы хрустальных глаз могут быть полезны в качестве маркеров, позволяющих узнать больше о нейронных цепях в областях сетчатки кубомедуз. [46]
Эволюция как ответ на естественные стимулы
Основные адаптивные реакции на изменения окружающей среды, наблюдаемые в глазах кубомедуз, включают скорость сужения зрачков в ответ на световую среду, а также настройку фоторецепторов и адаптацию хрусталика , чтобы лучше реагировать на изменения между светлой средой и темнотой. Интересно, что глаза некоторых видов кубомедуз, по-видимому, развили более сфокусированное зрение в зависимости от среды их обитания. [50]
Сокращение зрачков, по-видимому, развилось в ответ на изменение освещенности в экологических нишах трех видов кубомедуз ( Chironex fleckeri , Chiropsella bronzie и Carukia barnesi ). Поведенческие исследования показывают, что более высокая скорость сокращения зрачков позволяет лучше избегать объектов. [50] Фактически, виды с более сложной средой обитания демонстрируют более высокие темпы роста. Ч. Bronzie обитают на неглубоких пляжах с плохой видимостью и очень небольшим количеством препятствий, поэтому более быстрое сужение зрачков в ответ на объекты в их среде не имеет значения. ок. барнези и Ч. fleckeri встречаются в более сложной трехмерной среде, например в мангровых зарослях , с обилием естественных препятствий, где более быстрое сокращение зрачков является более адаптивным. [50] Поведенческие исследования подтверждают идею о том, что более высокая скорость сокращения зрачков помогает избегать препятствий, а также регулировать глубину в ответ на различную интенсивность света.
Адаптация к свету и темноте посредством зрачковых световых рефлексов является дополнительной формой эволюционного ответа на световую среду. Это связано с реакцией зрачка на изменение интенсивности света (обычно от солнечного света к темноте). В процессе адаптации к свету/темноте верхняя и нижняя линза глаза у разных видов кубомедуз различаются по специфическим функциям. [43] Нижние хрусталиковые глаза содержат пигментированные фоторецепторы и длинные пигментные клетки с темными пигментами, которые мигрируют при адаптации к свету/темне, тогда как верхние хрусталиковые глаза играют концентрированную роль в направлении света и фототаксисе, поскольку они обращены вверх, к поверхности воды (в сторону солнце или луна). [43] Верхняя линза Ch. bronzie не проявляет значительной оптической силы, тогда как Tr. цистофора (вид коробчатых медуз, обычно обитающий в мангровых зарослях). Способность использовать свет для визуального руководства поведением не имеет для Ч. большого значения. bronzie , как и вид в среде с большим количеством препятствий. [43] Различия в зрительно-ориентированном поведении служат свидетельством того, что виды, имеющие одинаковое количество и структуру глаз, могут демонстрировать различия в том, как они контролируют поведение.
Самый большой и самый маленький
Медузы имеют высоту и диаметр колокола примерно от одного миллиметра. [51] почти 2 метра ( 6 + 1 ⁄ ; фута) высоты и диаметра колокола щупальца и ротовая часть обычно выходят за пределы этого размера колокола. [25]
Самые маленькие медузы — своеобразные ползучие медузы из родов Staurocladia и Eleutheria , имеющие колоколообразные диски из От 0,5 миллиметров ( 1 ⁄ 32 дюйма) до нескольких миллиметров в диаметре, с короткими щупальцами, выходящими за пределы этого диаметра, которые эти медузы используют для перемещения по поверхности морских водорослей или дну каменистых водоемов; [51] Многие из этих крошечных ползающих медуз невозможно увидеть в полевых условиях без ручной линзы или микроскопа. Они могут размножаться бесполым путем делением (разделением пополам). Другие очень мелкие медузы, имеющие колокольчики около одного миллиметра, представляют собой гидромедузы многих видов, только что вышедшие из родительских полипов; [52] некоторые из них живут всего несколько минут, прежде чем отбросить свои гаметы в планктон и затем умереть, в то время как другие будут расти в планктоне в течение недель или месяцев. Hydromedusae Cladonema radiatum и Cladonema Californicum также очень маленькие, живут месяцами, но никогда не вырастают выше нескольких мм в высоту и диаметр колокола. [53]

Медуза со львиной гривой , Cyanea capillata , долгое время считалась самой крупной медузой и, возможно, самым длинным животным в мире, с тонкими нитевидными щупальцами, которые могут достигать 36,5 м (119 футов 9 дюймов) в длину (хотя большинство далеко не такие большие). [54] [55] У них умеренно болезненный, но редко смертельный укус. [56] Все более распространенная гигантская медуза Номуры , Nemopilema nomurai , встречающаяся в некоторые, но не во все годы в водах Японии , Кореи и Китая летом и осенью, является еще одним кандидатом на звание «крупнейшей медузы», по диаметру и весу, поскольку самая большая Медуза Номура поздней осенью может достигать 2 м (6 футов 7 дюймов) в диаметре колокола (тела) и весить около 200 кг (440 фунтов), при этом средние экземпляры часто достигают 0,9 м (2 фута 11 дюймов) в диаметре колокола и около Вес 150 кг (330 фунтов). [57] [58] Большая колоколообразная масса гигантской медузы Номуры. [59] может затмить ныряльщика и почти всегда намного больше Львиной Гривы, диаметр колокола которой может достигать 1 м (3 фута 3 дюйма). [60]
Редко встречающаяся глубоководная медуза Stygiomedusa gigantea — еще один кандидат на звание «крупнейшей медузы» с толстым массивным колоколом шириной до 100 см (3 фута 3 дюйма) и четырьмя толстыми, «ремешковыми» ротовыми ветвями, простирающимися до 6 м ( 19 + 1 ⁄ фута) в длину, сильно отличается от типичных тонких нитевидных щупалец , которые окаймляют зонтик более типичных на вид медуз, включая Львиную гриву. [61]
Desmonema glaciale , обитающая в Антарктическом регионе, может достигать очень больших размеров (несколько метров). [62] [63] Желе с пурпурными полосами ( Chrysaora colorata ) также может быть очень длинным (до 15 футов). [64]
История жизни и поведение

1–3 личинки ищут сайт
4–8 полипов растут
9–11 Полип стробилизирует
12–14 Медуза растет
Жизненный цикл
Медузы имеют сложный жизненный цикл, который включает как сексуальную, так и бесполую фазы, причем в большинстве случаев половой стадией является медуза. Сперматозоиды оплодотворяют яйца, которые развиваются в личиночные планулы, становятся полипами, отпочковываются в эфиры и затем превращаются во взрослых медуз. У некоторых видов определенные стадии могут быть пропущены. [65]
Достигнув взрослых размеров, медузы регулярно нерестятся, если имеется достаточный запас пищи. У большинства видов нерест контролируется светом, при этом все особи нерестятся примерно в одно и то же время суток; во многих случаях это происходит на рассвете или в сумерках. [66] Медузы обычно бывают мужского или женского пола (иногда встречаются гермафродиты ). В большинстве случаев взрослые особи выделяют сперму и яйца в окружающую воду, где незащищенные яйца оплодотворяются и развиваются в личинки. У некоторых видов сперма попадает в рот самки, оплодотворяя яйцеклетки внутри ее тела, где они остаются на ранних стадиях развития. У лунных медуз яйца откладываются в ямках на ротовых ветвях, которые образуют временную расплодную камеру для развивающихся личинок планулы . [67]
Планула — маленькая личинка , покрытая ресничками . При достаточном развитии он оседает на твердую поверхность и развивается в полип . Полип обычно состоит из небольшого стебля, увенчанного ртом, окруженным обращенными вверх щупальцами. Полипы напоминают полипы близкородственных антозоев , таких как морские анемоны и кораллы . Полип медузы может быть сидячим , обитающим на дне корпуса лодки или других субстратах, или свободно плавающим или прикрепленным к крошечным кусочкам свободноживущего планктона. [68] или редко, рыба [69] [70] или другие беспозвоночные. Полипы могут быть одиночными или колониальными. [71] Большинство полипов имеют диаметр всего несколько миллиметров и постоянно питаются. Стадия полипа может длиться годами. [25]
По прошествии определенного периода времени, стимулируемый сезонными или гормональными изменениями, полип может начать бесполое размножение путем почкования и у Scyphozoa называется сегментирующим полипом или сцифистомой. Почкование дает больше сцифистом, а также эфиров. [25] Места почкования различаются в зависимости от вида; из луковиц щупалец , рукоятки (над ртом) или гонад гидромедуз. [68] В ходе процесса, известного как стробилация , щупальца полипа рассасываются, и тело начинает сужаться, образуя поперечные сужения в нескольких местах вблизи верхней конечности полипа. Они углубляются по мере того, как места сужения перемещаются вниз по телу и отделяются отдельные сегменты, известные как эфиры. Это свободноплавающие предшественники стадии взрослой медузы, которую обычно называют медузой. [25] [72] Эфиры, обычно размером всего лишь один-два миллиметра в поперечнике, отплывают от полипа и растут. Полипы Limnomedusae могут бесполым путем образовывать ползучие личиночные панцири , которые уползают, прежде чем превратиться в другой полип. [25] Некоторые виды могут производить новых медуз путем отпочкования непосредственно на стадии медузы. Некоторые гидромедузы размножаются делением. [68]
Продолжительность жизни
Об истории жизни многих медуз известно мало, поскольку не обнаружено мест на морском дне, где обитают донные формы этих видов. Однако форма стробилы, размножающаяся бесполым путем, иногда может жить несколько лет, производя каждый год новых медуз (личинок эфиры). [73]
Необычный вид Turritopsis dohrnii , ранее классифицировавшийся как Turritopsis nutricula . [74] может быть фактически бессмертным из-за его способности при определенных обстоятельствах превращаться из медузы обратно в стадию полипа, тем самым избегая смерти, которая обычно ожидает медуз после размножения, если они иным образом не были съедены каким-либо другим организмом. До сих пор этот обратный процесс наблюдался только в лаборатории. [75]
Передвижение

На примере лунного желе Aurelia aurita было доказано, что медузы являются наиболее энергоэффективными пловцами из всех животных. [76] Они движутся в воде, радиально расширяя и сжимая свои колоколообразные тела, толкая воду за собой. Они делают паузу между фазами сжатия и расширения, создавая два вихревых кольца. Для сокращения тела используются мышцы, создающие первый вихрь и толкающие животное вперед, но мезоглея настолько эластична, что расширение происходит исключительно за счет расслабления колокола, высвобождающего энергию, запасенную от сокращения. Тем временем второе вихревое кольцо начинает вращаться быстрее, засасывая воду в колокол и прижимаясь к центру тела, давая вторичный и «свободный» толчок вперед. Механизм, называемый пассивным возвратом энергии, работает только с относительно небольшими медузами, движущимися с низкой скоростью, позволяя животному путешествовать на 30 процентов дальше за каждый цикл плавания. Медузы достигли на 48 процентов более низких затрат на транспортировку (потребление пищи и кислорода по сравнению с энергией, затраченной на движение), чем другие животные в аналогичных исследованиях. Одна из причин этого заключается в том, что большая часть студенистой ткани колокола неактивна и не использует энергию во время плавания. [77]
Экология
Диета
Медузы, как и другие книдарии, обычно плотоядны (или паразитичны). [78] питаются планктонными организмами, ракообразными, мелкой рыбой, икрой и личинками рыб, а также другими медузами, заглатывая пищу и выводя непереваренные отходы через рот. Они охотятся пассивно, используя свои щупальца в качестве дрейфа, или тонут в воде, широко раскинув щупальца; щупальца, содержащие нематоцисты, оглушающие или убивающие добычу , затем могут сгибаться, помогая поднести ее ко рту. [25] Их техника плавания также помогает им ловить добычу; когда их колокол расширяется, он всасывает воду, что приводит к тому, что в пределах досягаемости щупалец появляется больше потенциальной добычи. [79]
Некоторые виды, такие как Aglaura hemistoma , всеядны и питаются микропланктоном, который представляет собой смесь зоопланктона и фитопланктона (микроскопических растений), таких как динофлагелляты . [80] Другие содержат мутуалистические водоросли ( зооксантеллы ); в своих тканях [25] Типична из них пятнистая медуза ( Mastigias papua ), получающая часть своего питания за счет продуктов фотосинтеза , а часть за счет пойманного зоопланктона. [81] [82] ( Перевернутая медуза Cassiopea andromeda ) также имеет симбиотические отношения с микроводорослями , но ловит крошечных животных, чтобы дополнить свой рацион. Это делается путем высвобождения крошечных шариков живых клеток, состоящих из мезоглеи . Они используют реснички, чтобы провести себя через воду, и стрекательные клетки, которые оглушают добычу. Кажется, что капли также обладают пищеварительными способностями. [83]
Хищничество
Другие виды медуз являются одними из наиболее распространенных и важных хищников медуз. Морские анемоны могут поедать медуз, попадающих в их ареал. Среди других хищников — тунцы , акулы, рыба-меч , морские черепахи и пингвины. [84] [85] Медуз, выброшенных на берег, поедают лисы, другие наземные млекопитающие и птицы. [86] Однако в целом немногие животные охотятся на медуз; в целом их можно считать главными хищниками в пищевой цепи. Как только медузы стали доминировать в экосистеме, например, в результате чрезмерного вылова рыбы, который уничтожает хищников, питающихся личинками медуз, может не оказаться очевидного способа восстановить прежний баланс: они поедают рыбную икру и молодь рыб и конкурируют с рыбами за пищу, предотвращая рыбные запасы от восстановления. [87]
Симбиоз
Некоторые мелкие рыбы невосприимчивы к укусам медузы и живут среди щупалец, служа приманкой в ловушке для рыбы; они в безопасности от потенциальных хищников и могут поделиться рыбой, пойманной медузой. [88] Медуза -пушечное ядро находится в симбиотических отношениях с десятью различными видами рыб, а также с длинноносым крабом-пауком , который живет внутри колокола, делясь пищей с медузой и покусывая ее ткани. [89]
Цветет

Круги представляют записи данных; большие кружки обозначают более высокую достоверность результатов.
Медузы образуют большие массы или цветут в определенных условиях окружающей среды: океанских течениях , питательных веществах , солнечном свете, температуре, сезоне, наличии добычи, уменьшении хищничества и концентрации кислорода . Течения собирают медуз вместе, особенно в годы с необычно высокой популяцией. Медузы могут обнаруживать морские течения и плыть против течения, собираясь в цветки. [91] [92] Медузы лучше выживают в богатой питательными веществами и бедной кислородом воде, чем их конкуренты, и поэтому могут питаться планктоном без конкуренции. Медузам также может быть полезна более соленая вода, поскольку более соленая вода содержит больше йода , который необходим полипам для превращения в медуз. Повышение температуры моря, вызванное изменением климата , также может способствовать цветению медуз, поскольку многие виды медуз способны выживать в более теплых водах. [93] Увеличение количества питательных веществ из сельскохозяйственных или городских стоков с питательными веществами, включая соединения азота и фосфора, увеличивает рост фитопланктона, вызывая эвтрофикацию и цветение водорослей . Когда фитопланктон умирает, он может создавать мертвые зоны , называемые так потому, что они гипоксичны (с низким содержанием кислорода). Это, в свою очередь, убивает рыб и других животных, но не медуз. [94] позволяя им цвести. [95] [96] Популяции медуз могут расширяться по всему миру в результате поверхностного стока и чрезмерного вылова их естественных хищников . [97] [98] Медузы имеют все шансы извлечь выгоду из нарушения морских экосистем. Они быстро размножаются; они охотятся на многие виды, тогда как на них охотятся лишь немногие виды; и они питаются посредством прикосновения, а не визуально, поэтому могут эффективно питаться ночью и в мутной воде. [99] [100] может быть трудно Рыбным запасам восстановиться в морских экосистемах после того, как в них начнут доминировать медузы, поскольку медузы питаются планктоном, который включает рыбную икру и личинки . [101] [102] [96]

Как и предполагалось на рубеже этого столетия, [107] [108] цветение медуз становится все более частым. В период с 2013 по 2020 год Средиземноморская научная комиссия еженедельно отслеживала частоту таких вспышек в прибрежных водах от Марокко до Черного моря, обнаруживая относительно высокую частоту этих цветений почти круглый год, при этом пики наблюдались с марта по июль, а зачастую и с марта по июль. снова осенью. Цветение вызвано разными видами медуз в зависимости от их локализации в пределах бассейна: наблюдается явное доминирование вспышек Pelagia noctiluca и Velella velella в западном Средиземноморье, вспышек Rhizostoma pulmo и Rhopilema nomadica в восточном Средиземноморье, а также Aurelia aurita. и Mnemiopsis leidyi в Черном море. [109]
Некоторые популяции медуз, которые продемонстрировали явный рост за последние несколько десятилетий, являются инвазивными видами , недавно прибывшими из других мест обитания: примеры включают Черное море , Каспийское море , Балтийское море , центральное и восточное Средиземноморье , Гавайи, а также тропические и субтропические части Запада. Атлантический (включая Карибский бассейн , Мексиканский залив и Бразилию). [105] [106]
Цветение медуз может оказать существенное влияние на структуру сообщества. Некоторые хищные виды медуз питаются зоопланктоном, тогда как другие питаются первичными продуцентами. [110] Сокращение количества зоопланктона и ихтиопланктона из-за цветения медуз может затронуть трофические уровни. Популяции медуз с высокой плотностью могут вытеснить других хищников и сократить пополнение рыбы. [111] Увеличение выпаса медузами первичных производителей также может прервать передачу энергии на более высокие трофические уровни. [112]
Во время цветения медузы значительно изменяют доступность питательных веществ в окружающей среде. Для роста цветения требуется большое количество доступных органических питательных веществ в толще воды, что ограничивает доступность для других организмов. [113] Некоторые медузы имеют симбиотические отношения с одноклеточными динофлагеллятами, что позволяет им ассимилировать неорганический углерод, фосфор и азот, создавая конкуренцию фитопланктону. [113] Их большая биомасса делает их важным источником растворенных и твердых органических веществ для микробных сообществ посредством выделения, производства слизи и разложения. [90] [114] Микробы расщепляют органические вещества на неорганический аммоний и фосфат. Однако низкая доступность углерода смещает процесс с производства на дыхание, создавая области с низким содержанием кислорода, что делает растворенный неорганический азот и фосфор практически недоступными для первичного производства.
Эти цветения оказывают весьма реальное влияние на промышленность. Медузы могут превзойти рыбу, используя открытые ниши в условиях чрезмерного вылова рыбы. [115] Улов медуз может привести к перегрузке рыболовных снастей и привести к расходам, связанным с повреждением снастей. Электростанции были остановлены из-за медуз, блокирующих поток охлаждающей воды. [116] Цветение также нанесло ущерб туризму, вызывая рост числа укусов, а иногда и закрытие пляжей. [117]
Медузы являются компонентом желепадения — явления, когда студенистый зоопланктон падает на морское дно, обеспечивая там пищу донным организмам. [118] В умеренных и приполярных регионах желепады обычно следуют сразу после цветения. [119]
Места обитания

Большинство медуз — морские животные, хотя несколько гидромедуз обитают в пресных водах . Самый известный пресноводный пример — космополитная гидрозойная медуза Craspedacusta sowerbii . Он меньше дюйма (2,5 см) в диаметре, бесцветен и не жалит. [120] Некоторые популяции медуз стали обитать в прибрежных соленых озерах, таких как озеро Медуз в Палау. [121] Озеро медуз — это морское озеро , в котором ежедневно горизонтально мигрируют миллионы золотых медуз ( виды Mastigias ). [82]
Хотя большинство медуз живут далеко от дна океана и составляют часть планктона, некоторые виды большую часть своей жизни тесно связаны со дном и могут считаться бентосными . Перевернутые медузы рода Cassiopea обычно лежат на дне неглубоких лагун, где они иногда мягко пульсируют, повернув верхушкой зонтика вниз. Даже некоторые глубоководные виды гидромедуз и сцифомедуз обычно собираются на дне или вблизи него. Все стауромедузы прикреплены либо к морским водорослям, либо к каменистому или другому твердому материалу на дне. [122]
Некоторые виды явно адаптируются к приливным потокам. В заливе Роско медузы плывут по течению во время отлива, пока не ударяются о гравийную косу , а затем опускаются ниже течения. Они остаются в стоячих водах до тех пор, пока не поднимется прилив, поднимаясь и позволяя ему унести их обратно в залив. Они также активно избегают пресной воды из талых горных снегов, ныряя до тех пор, пока не найдут достаточно соли. [2]
Паразиты
Медузы являются хозяевами самых разных паразитических организмов. Они выступают в качестве промежуточных хозяев эндопаразитических гельминтов , при этом инфекция передается окончательному хозяину-рыбе после нападения хищников . Некоторые дигенеевые трематоды , особенно виды семейства Lepocreadiidae , используют медуз в качестве вторых промежуточных хозяев. Рыбы заражаются трематодами, когда питаются зараженными медузами. [123] [124]
Отношение к людям

Рыболовство
Медуз уже давно едят в некоторых частях мира. [3] Рыболовство начало добычу американской медузы-пушечника Stomolophus meleagris вдоль южного атлантического побережья США и в Мексиканском заливе для экспорта в Азию. [126]
Медуз также добывают для получения коллагена , который исследуется для использования в различных целях, включая лечение ревматоидного артрита . [127]
Аквакультура и рыболовство других видов часто несут серьезные потери – и, следовательно, потери продуктивности – из-за медуз. [128] [129]
Продукты

Аристотель заявил в « Частях животных» IV, 6, что медуз (морскую крапиву) ели зимой в рыбном рагу. [130]
В некоторых странах, включая Китай, Японию и Корею, медузы являются деликатесом. Медузу сушат, чтобы она не испортилась.Лишь около 12 видов сцифозных медуз, принадлежащих к отряду Rhizostomeae, добываются в пищу, в основном в Юго-Восточной Азии. [131] Корневища, особенно Rhopilema esculentum в Китае ( 海蜇 hǎizhé , «морские жала») и Stomolophus meleagris (медуза-пушечное ядро) в Соединенных Штатах, пользуются предпочтением из-за их более крупных и жестких тел, а также потому, что их токсины безвредны для человека. [126]
Традиционные методы обработки, осуществляемые мастером-медузой, предполагают многофазную процедуру длительностью от 20 до 40 дней, при которой после удаления половых желез и слизистых оболочек зонтик и ротовые дужки обрабатываются смесью поваренной соли и квасцов ; и сжатый. Обработка делает медузу более сухой и кислой, создавая хрустящую текстуру. Приготовленные таким образом медузы сохраняют 7–10% первоначального веса, а переработанный продукт состоит примерно на 94% из воды и на 6% из белка. Свежеобработанная медуза имеет белый, кремовый цвет и при длительном хранении желтеет или коричневеет. [126]
В Китае обработанных медуз опресняют, замачивая на ночь в воде, и едят в вареном или сыром виде. Блюдо часто подают натертым с заправкой из масла, соевого соуса, уксуса и сахара или в виде салата с овощами. В Японии вяленую медузу ополаскивают, нарезают соломкой и подают с уксусом в качестве закуски. [126] [132] Также доступны обессоленные, готовые к употреблению продукты. [126]
Биотехнология

Плиний Старший сообщил в своей «Естественной истории » , что слизь медузы Pulmo marinus излучает свет, если ее потереть о трость. [133]
В 1961 году Осаму Шимомура извлек зеленый флуоресцентный белок (GFP) и другой биолюминесцентный белок, названный экворином , из крупной и многочисленной гидромедузы Aequorea victoria , одновременно изучая фотопротеины , вызывающие биолюминесценцию у этого вида. [134] Три десятилетия спустя Дуглас Прашер секвенировал и клонировал ген GFP. [135] Мартин Чалфи придумал, как использовать GFP в качестве флуоресцентного маркера генов, встроенных в другие клетки или организмы. [136] Позже Роджер Цзянь химически манипулировал GFP, чтобы получить другие флуоресцентные цвета для использования в качестве маркеров. В 2008 году Шимомура, Чалфи и Цьен получили Нобелевскую премию по химии за работу с GFP. [134] Искусственный GFP стал широко использоваться в качестве флуоресцентной метки , чтобы показать, какие клетки или ткани экспрессируют определенные гены. Метод генной инженерии объединяет ген интересующий с геном GFP. Затем слитую ДНК помещают в клетку для создания либо клеточной линии, либо (с помощью методов ЭКО ) всего животного, несущего этот ген. В клетке или животном искусственный ген включается в тех же тканях и в то же время, что и нормальный ген, образуя слияние нормального белка с GFP, прикрепленным к концу, освещение животного или клетки показывает, какие ткани экспрессируют этот белок — или на какой стадии развития. Флуоресценция показывает, где экспрессируется ген. [137]
Аквариумный дисплей

Медузы представлены во многих общественных аквариумах . Часто фон аквариума синий, а животные освещаются боковым светом, что увеличивает контраст между животным и фоном. В естественных условиях многие желе настолько прозрачны, что их практически не видно. [138] Медузы не приспособлены к жизни в закрытых помещениях. Они зависят от течений, которые переносят их с места на место. Профессиональные экспонаты, такие как аквариум Монтерей-Бей, имеют точные потоки воды, обычно в круглых резервуарах, чтобы избежать застревания особей в углах. Отток распределяется по большой площади поверхности, а приток входит в виде слоя воды перед оттоком, поэтому медузы не засасываются в него. [139] С 2009 года медузы стали популярны в домашних аквариумах, где для них требуется аналогичное оборудование. [140]
Укусы
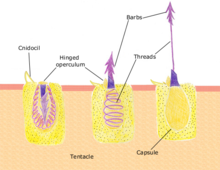
Медузы вооружены нематоцистами — разновидностью специализированных стрекательных клеток. [141] Контакт с щупальцами медузы может спровоцировать появление миллионов нематоцист, которые проткнут кожу и впрыскат яд . [142] но яд только некоторых видов вызывает у человека неблагоприятную реакцию. [143] В исследовании, опубликованном в журнале Communications Biology , исследователи обнаружили вид медуз под названием Cassiopea xamachana , который при срабатывании выделяет крошечные шарики клеток, которые плавают вокруг медузы, жаля все на своем пути. Исследователи описали их как «самодвижущиеся микроскопические гранаты» и назвали кассиосомами. [144]
Последствия укусов варьируются от легкого дискомфорта до сильной боли и смерти. [145] [146] Укусы большинства медуз не смертельны, но укусы некоторых кубомедуз ( медуз Ируканджи ), например морской осы , могут быть смертельными. Укусы могут вызвать анафилаксию (разновидность шока), которая может привести к летальному исходу. Только на Филиппинах медузы убивают от 20 до 40 человек в год. В 2006 году Испанский Красный Крест оказал помощь 19 000 ужаленным пловцам на побережье Коста-Брава . [146] [147]
Уксус (3–10% водный раствор уксусной кислоты ) может помочь при кубомедуз . укусах [148] [149] но не укусы португальского военного корабля . [148] Очистка области от желе и щупалец уменьшает количество нематоцист. [150] Соскоб пораженной кожи, например, краем кредитной карты, может удалить оставшиеся нематоцисты. [151] После очистки кожи от нематоцист местное применение крема с гидрокортизоном уменьшает боль и воспаление. [152] Антигистаминные препараты могут помочь контролировать зуд . [151] Иммунные противоядия используются при серьезных укусах кубомедуз. [153] [154]
На островах Эльба и Корсике жители и туристы теперь используют dittrichia viscosa для лечения укусов медуз, пчел и ос путем прижатия свежих листьев к коже, что приводит к быстрым результатам.
 Кубомедузы маленькие и ядовитые.
Кубомедузы маленькие и ядовитые. Медузный дерматит
Медузный дерматит Флаг на пляже Сарауса, предупреждающий о наличии в воде медуз.
Флаг на пляже Сарауса, предупреждающий о наличии в воде медуз.



Механические проблемы
Медузы в больших количествах могут заполнить и расколоть рыболовные сети и раздавить пойманную рыбу. [155] Они могут засорить холодильное оборудование, выведя из строя электростанции в ряде стран; медузы стали причиной каскадного отключения электроэнергии на Филиппинах в 1999 году. [146] а также повреждение электростанции Диабло-Каньон в Калифорнии в 2008 году. [156] Они также могут остановить опреснительные установки и судовые двигатели. [155] [157]
См. также
- Медузный дерматит
- Список доисторических медузозойных
- Океанская солнечная рыба , крупный хищник медуз.
- Гребневик
Примечания
Ссылки
- ^ «Летопись окаменелостей выявила неуловимую медузу возрастом более 500 миллионов лет» . ScienceDaily . Проверено 10 февраля 2023 г.
- ^ Jump up to: Перейти обратно: а б с Анжер, Натали (6 июня 2011 г.). «Намного больше, чем плазма и яд» . Нью-Йорк Таймс . Архивировано из оригинала 18 мая 2013 года . Проверено 2 декабря 2011 г.
- ^ Jump up to: Перейти обратно: а б Изабель Родд (20 октября 2020 г.). «Почему медуза может быть «идеальной едой» » . Новости Би-би-си . Проверено 7 мая 2023 г.
- ^ Jump up to: Перейти обратно: а б «медуза» . Интернет-словарь этимологии . Проверено 9 июня 2018 г.
- ^ Келман, Джанет Харви; Преподобный Теодор Вуд (1910). Берег моря, показанный детям . Лондон: TC и EC Джек. п. 146. ОЛ 7043926М .
- ^ Каплан, Юджин Х.; Каплан, Сьюзен Л.; Петерсон, Роджер Тори (август 1999 г.). Полевой путеводитель по коралловым рифам: Карибский бассейн и Флорида . Бостон: Хоутон Миффлин. п. 55. ИСБН 978-0-618-00211-5 .
- ^ «Цветочная шляпка желе» . 6 апреля 2009 года. Архивировано из оригинала 6 апреля 2009 года . Проверено 10 февраля 2023 г.
- ^ «Что такое рыба?» . Энциклопедия жизни. Архивировано из оригинала 24 марта 2018 года . Проверено 13 октября 2018 г.
И большинству людей известно, что миноги, акулы, скаты, угри, морские коньки и другие странного вида водные существа — это рыбы, а моллюски, каракатицы, морские звезды, раки и медузы (несмотря на их названия) — не рыбы.
- ^ Бротц, Лукас. Изменение популяций медуз: тенденции в крупных морских экосистемах. Архивировано 16 апреля 2013 года в Wayback Machine . 2011. стр.1.
- ^ Куломб, Дебора А. (14 февраля 1990 г.). Приморский натуралист: Путеводитель по обучению на берегу моря . Саймон и Шустер . п. 60. ИСБН 978-0-671-76503-3 . Архивировано из оригинала 31 декабря 2013 года . Проверено 20 марта 2013 г.
- ^ Клаппенбах, Лаура. «Десять фактов о медузах» . Архивировано из оригинала 26 февраля 2009 года . Проверено 24 января 2010 г.
- ^ Липтон, Джеймс (1991). Возвышение жаворонков . Викинг. ISBN 978-0-670-30044-0 .
- ^ Макивер, Ангус (2004). Первая помощь на английском языке . Ходдер Гибсон (Хачетт). ISBN 978-1-444-19376-3 .
- ^ Картрайт, Полин; Хальгедал, Сьюзен Л .; Хендрикс, Джонатан Р.; и др. (2007). Хамфрис, Стюарт (ред.). «Исключительно сохранившиеся медузы среднего кембрия» . ПЛОС ОДИН . 2 (10): е1121. Бибкод : 2007PLoSO...2.1121C . дои : 10.1371/journal.pone.0001121 . ПМК 2040521 . ПМИД 17971881 .
- ^ «Медуза» . Мерриам-Вебстер . 1 сентября 2018 года . Проверено 11 сентября 2018 г.
- ^ Jump up to: Перейти обратно: а б с д и ж г час я «Обнаружение медуз | Виды медуз» . Политически ориентированные морские экологические исследования в южноевропейских морях (PERSEUS) . Проверено 28 августа 2018 г.
- ^ Jump up to: Перейти обратно: а б Миллс, CE (8 ноября 2010 г.). «Гребневики» . Университет Вашингтона . Проверено 28 августа 2018 г.
- ^ Jump up to: Перейти обратно: а б «Наши желеобразные родственники: Распространенные заблуждения о сальпах» . Программа Нерей . Проверено 28 августа 2018 г.
- ^ Jump up to: Перейти обратно: а б Сапата, Фелипе; Гетц, Фрейя Э.; Смит, Стивен А.; и др. (2015). «Филогеномный анализ поддерживает традиционные отношения внутри Cnidaria» . ПЛОС ОДИН . 10 (10): e0139068. Бибкод : 2015PLoSO..1039068Z . дои : 10.1371/journal.pone.0139068 . ПМЦ 4605497 . ПМИД 26465609 .
- ^ Каял, Эхсан; Бентлаге, Бастиан; Сабрина Пэнки, М.; и др. (2018). «Филогеномика обеспечивает надежную топологию основных линий книдарий и дает представление о происхождении ключевых особенностей организма» . Эволюционная биология BMC . 18 (1): 68. Бибкод : 2018BMCEE..18...68K . дои : 10.1186/s12862-018-1142-0 . ПМЦ 5932825 .
- ^ «STAUROMEDUSAE UK Онлайн-путеводитель по стебельчатым медузам (Stauromedusae), обитающим в прибрежных водах Соединенного Королевства и Ирландии. Включает примечания по их идентификации, а также о том, где и как их найти. НАЗАД Великобритания Контрольный список для стебельчатых медуз (Stauromedusae)» . Ставуромедузы Великобритания . Проверено 28 августа 2018 г.
- ^ Шируотер, Бернд; Хелм, Ребекка Р.; Данн, Кейси В. (2017). «Индолы вызывают метаморфоз у широкого спектра медуз, но не у медуз (Coronatae)» . ПЛОС ОДИН . 12 (12): e0188601. Бибкод : 2017PLoSO..1288601H . дои : 10.1371/journal.pone.0188601 . ПМЦ 5744923 . ПМИД 29281657 .
- ^ Осборн, KJ (2014). «Медуза-красный бумажный фонарь» . Смитсоновский институт . Проверено 13 октября 2018 г.
- ^ Дейли, Джейсон (1 марта 2017 г.). «Взгляните на завораживающую «космическую медузу» » . Смитсоновский институт . Проверено 28 августа 2018 г.
- ^ Jump up to: Перейти обратно: а б с д и ж г час я дж к л м н тот п д р с т Руперт, Эдвард Э.; Фокс, Ричард, С.; Барнс, Роберт Д. (2004). Зоология беспозвоночных, 7-е издание . Cengage Обучение. стр. 148–174. ISBN 978-81-315-0104-7 .
{{cite book}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Jump up to: Перейти обратно: а б Cnidaria. Архивировано 21 сентября 2012 г. в Wayback Machine , Древо жизни.
- ^ Маркес, AC; А.Г. Коллинз (2004). «Кладистический анализ Medusozoa и эволюции книдарий». Биология беспозвоночных . 123 : 23–42. дои : 10.1111/j.1744-7410.2004.tb00139.x . S2CID 28342963 .
- ^ Крамп, PL (1961). «Краткий обзор Медуз мира». Журнал Морской биологической ассоциации Соединенного Королевства . 40 : 1–469. Бибкод : 1961JMBUK..40....7K . дои : 10.1017/s0025315400007347 .
- ^ Мун, Джастин; Карон, Жан-Бернар; Мойсюк, Иосиф (9 августа 2023 г.). «Макроскопическая свободно плавающая медуза из среднекембрийских сланцев Берджесс». Труды Королевского общества B: Биологические науки . 290 (2004). дои : 10.1098/rspb.2022.2490 . ISSN 0962-8452 . PMC 10394413. PMID 37528711 .
- ^ Се, Юн-Хва; Радлоу, Джек (1994). «Потенциал использования медуз в пищу в западных странах». Тенденции в пищевой науке и технологиях . 5 (7): 225–229. дои : 10.1016/0924-2244(94)90253-4 .
- ^ «Медуза — Иллюстрированный словарь» . infovisual.info . Проверено 10 февраля 2023 г.
- ^ Ваггонер, Бен; Коллинз, Аллен Г. «Кубозоа: больше о морфологии» . Музей палеонтологии Калифорнийского университета . Проверено 6 января 2019 г.
- ^ «Нематоцисты» . Железона . 2 апреля 2015 года. Архивировано из оригинала 2 апреля 2015 года . Проверено 29 марта 2014 г.
- ^ Кир, Уильям (2012). «Многообразие гидростатических скелетов» . Журнал экспериментальной биологии . 215 (Часть 8): 1247–1257. дои : 10.1242/jeb.056549 . ПМИД 22442361 .
- ^ Саттерли, РА (2002). «Нейрональный контроль плавания медуз: сравнительная история» . Канадский журнал зоологии . 80 (10): 1654–1669. дои : 10.1139/z02-138 . Архивировано из оригинала (PDF) 12 июля 2013 года.
- ^ Кацуки, Такео; Гринспен, Ральф Дж. (2013). «Нервная система медузы» . Современная биология . 23 (14): R592–R594. Бибкод : 2013CBio...23.R592K . дои : 10.1016/j.cub.2013.03.057 . ПМИД 23885868 .
- ^ Саттерли, Ричард А. (2011). «Есть ли у медуз центральная нервная система?» . Журнал экспериментальной биологии . 214 (8): 1215–1223. дои : 10.1242/jeb.043687 . ПМИД 21430196 .
- ^ Венер, Р. (2005). «Сенсорная физиология: безмозглые глаза» (PDF) . Природа . 435 (7039): 157–159. Бибкод : 2005Natur.435..157W . дои : 10.1038/435157a . ПМИД 15889076 . S2CID 4408533 . Архивировано (PDF) из оригинала 29 июля 2013 года.
- ^ «Многоглазая медуза помогает решить загадку Дарвина» . Новый учёный . Проверено 10 февраля 2023 г.
- ^ Jump up to: Перейти обратно: а б с д и ж Нильссон, Делавэр (2013). «Эволюция глаза и его функциональная основа» . Визуальная нейронаука . 30 (1–2): 5–20. дои : 10.1017/S0952523813000035 . ПМЦ 3632888 . ПМИД 23578808 .
- ^ Jump up to: Перейти обратно: а б с д и ж г час я дж к л м н тот п Гарм, Андерс; Экстрем, Питер (2010). «Доказательства существования множественных фотосистем у медуз». Глава 2 – Доказательства существования множественных фотосистем у медуз . Том. 280. Академическая пресса. стр. 41–78. дои : 10.1016/S1937-6448(10)80002-4 . ISBN 9780123812605 . ISSN 1937-6448 . ПМИД 20797681 .
{{cite book}}:|journal=игнорируется ( помогите ) - ^ Шуга Х., Чопп П., Грациусси Д.Ф. и др. (10 августа 2010 г.). «Гибкое использование генов Pax в развитии глаз на ранней стадии эволюции животных продемонстрировано исследованиями на гидрозойных медузах» . ПНАС . 107 (32): 14263–14268. Бибкод : 2010PNAS..10714263S . дои : 10.1073/pnas.1008389107 . ПМЦ 2922549 . ПМИД 20660753 .
- ^ Jump up to: Перейти обратно: а б с д и ж О'Коннор М., Гарм А., Нильссон Д.Э. (2009). «Строение и оптика глаз кубомедузы Chiropsella Bronzie». Журнал сравнительной физиологии А. 195 (6): 557–569. дои : 10.1007/s00359-009-0431-x . ПМИД 19347342 . S2CID 9563849 .
- ^ Нильссон Д.Э., Гислен Л., Коутс М. и др. (2005). «Передовая оптика в глазу медузы». Природа . 435 (7039): 201–205. Бибкод : 2005Natur.435..201N . дои : 10.1038/nature03484 . ПМИД 15889091 . S2CID 4418085 .
- ^ Jump up to: Перейти обратно: а б с д Пятигорский Ю., Козьмик З. (2004). «Кубозойская медуза: модель Evo/Devo для глаз и других сенсорных систем» . Межд. Дж. Дев. Биол . 48 (8–9): 719–729. doi : 10.1387/ijdb.041851jp . ПМИД 15558464 .
- ^ Jump up to: Перейти обратно: а б с Грей Г.К., Мартин В.Дж., Саттерли Р.А. (август 2009 г.). «Ультраструктура синапсов сетчатки кубозойных» (PDF) . Биол Булл . 217 (1): 35–49. дои : 10.1086/BBLv217n1p35 . ПМИД 19679721 . S2CID 24400231 .
- ^ Jump up to: Перейти обратно: а б Бергер, Эдвард В. (1898). «Гистологическое строение глаз кубомедуз» . Журнал сравнительной неврологии . 8 (3): 223–230. дои : 10.1002/cne.910080317 . S2CID 85422599 .
- ^ Суга, Хироши; Шмид, Волкер; Геринг, Уолтер Дж. (2008). «Эволюция и функциональное разнообразие опсинов медуз» . Современная биология . 18 (1): 51–55. Бибкод : 2008CBio...18...51S . дои : 10.1016/j.cub.2007.11.059 . ISSN 0960-9822 . ПМИД 18160295 . S2CID 13344739 .
- ^ Jump up to: Перейти обратно: а б с д Пятигорский Дж., Хорвиц Дж., Кувабара Т., Катресс С. (1989). «Клеточная глазная линза и кристаллины кубомедузанских медуз». Журнал сравнительной физиологии А. 164 (5): 577–587. дои : 10.1007/bf00614500 . ПМИД 2565398 . S2CID 19797109 .
- ^ Jump up to: Перейти обратно: а б с Сеймур, Джейми Э.; О'Хара, Эмили П. (2020). «Реакция зрачков на свет у трех видов Cubozoa (коробчатых медуз)» . Исследования планктона и бентоса . 15 (2): 73–77. дои : 10.3800/pbr.15.73 . S2CID 219759193 .
- ^ Jump up to: Перейти обратно: а б Миллс, CE; Хирано, Ю.М. (2007). Энциклопедия приливных озер и скалистых берегов: Hydromedusae . Издательство Калифорнийского университета. стр. 286–288. ISBN 978-0-520-25118-2 .
- ^ Миллс, CE (1976). « Podocoryne selena , новый вид гидроидов из Мексиканского залива, и сравнение с Hydractinia echinata » . Биологический бюллетень . 151 (1): 214–224. дои : 10.2307/1540715 . JSTOR 1540715 .
- ^ Костелло, Дж. (1988). «Лабораторное культивирование и кормление гидромедузы Cladonema californicum Hyman (Anthomedusa: Cladonemidae)». Журнал экспериментальной морской биологии и экологии . 123 (2): 177–188. Бибкод : 1988JEMBE.123..177C . дои : 10.1016/0022-0981(88)90168-2 .
- ^ «Редкое появление медузы со львиной гривой в заливе Трамор» . Уотерфорд сегодня. 1 августа 2007 года. Архивировано из оригинала 30 мая 2010 года . Проверено 18 октября 2010 г.
- ^ «Медуза Львиная грива - Справочная библиотека» . красныйОрбита. 12 июня 2003 года. Архивировано из оригинала 30 июля 2010 года . Проверено 18 октября 2010 г.
- ^ «150 медуз ужалили на Рай-Бич» . Wmur.com. 21 июля 2010 года. Архивировано из оригинала 14 октября 2011 года . Проверено 11 июня 2018 г.
- ^ Омори, Макото; Китамура, Минору (2004). «Таксономический обзор трех японских видов съедобных медуз (Scyphozoa: Rhizostomeae)» (PDF) . Планктонная биология и экология . 51 (1): 36–51. Архивировано (PDF) из оригинала 23 марта 2012 года.
- ^ Уе, Син-Ичи (2008). «Цветение гигантской медузы Nemopilema nomurai : угроза устойчивости рыболовства в восточноазиатских окраинных морях» (PDF) . Исследования планктона и бентоса . 3 (Приложение): 125–131. дои : 10.3800/pbr.3.125 . Архивировано (PDF) из оригинала 16 мая 2013 года.
- ^ «Гигантская медуза Этидзен у берегов Японии» . Би-би-си. 30 ноября 2009 г. Архивировано из оригинала 1 января 2011 г.
- ^ Крамп, PL (1961). «Краткий обзор медуз мира». Журнал Морской биологической ассоциации Соединенного Королевства . 40 : 1–469. Бибкод : 1961JMBUK..40....7K . дои : 10.1017/s0025315400007347 .
- ^ Бертон, Джоди (23 апреля 2010 г.). «Гигантская глубоководная медуза снята в Мексиканском заливе» . BBC Новости Земли . Архивировано из оригинала 5 июля 2010 года.
- ^ «Фотографии антарктической гигантской медузы (Desmonema glaciale) • iNaturalist» .
- ^ Лига, Майкл (11 октября 2011 г.). «Как завершить погружение» . ПоларТРЕК . Станция Мак-Мёрдо , Антарктида. Архивировано из оригинала 13 июня 2023 года . Проверено 13 июня 2023 г.
- ^ «Ныряние под воду с гигантскими медузами» . 26 апреля 2010 г.
- ^ «Как размножаются медузы? Какое влияние оказывает их жало на человека? В чем разница между красными и полупрозрачными медузами?» . Научный американец . 15 октября 2013 года. Архивировано из оригинала 23 октября 2013 года . Проверено 22 октября 2013 г.
- ^ Миллс, Клаудия (1983). «Вертикальная миграция и суточная активность гидромедуз: исследования в большом аквариуме». Журнал исследований планктона . 5 (5): 619–635. дои : 10.1093/планкт/5.5.619 . S2CID 13914997 .
- ^ Епископ, Андрей. «Лунная желе ( Aurelia aurita )» . Морские беспозвоночные Бермудских островов . Проверено 11 июня 2018 г.
- ^ Jump up to: Перейти обратно: а б с Миллс, CE (1987). Ж. Бульон; Ф. Боэро; Ф. Чиконья; ПФС Корнелиус (ред.). Исследования живых гидромедуз и гидроидов на месте и на корабле: предварительные наблюдения за адаптацией жизненного цикла к открытому океану . Кларендон Пресс. ISBN 978-0-19-857190-2 .
{{cite book}}:|work=игнорируется ( помогите ) - ^ Фьюкс, Дж. Уолтер (1887). «Гидроид, паразитирующий на рыбе» . Природа . 36 (939): 604–605. Бибкод : 1887Natur..36..604F . дои : 10.1038/036604b0 . S2CID 4078889 .
- ^ Шухерт, Питер. «Гидрозоа» . Архивировано из оригинала 4 февраля 2010 года . Проверено 24 января 2010 г.
- ^ «Как растут медузы: от яиц до полипов и медуз» . МысльКо . Проверено 10 февраля 2023 г.
- ^ Хьюз, Клэр. «Жизненный цикл коробчатой медузы» . Artforlibraries.org . Архивировано из оригинала 4 марта 2016 года . Проверено 2 января 2016 г.
- ^ Бруска, Ричард (2016). Беспозвоночные . Синауэр Ассошиэйтс. п. 310. ИСБН 978-1-60535-375-3 .
- ^ Миглиетта, член парламента; Пираино, С.; Кубота, С.; Шухерт, П. (2007). «Виды рода Turritopsis (Cnidaria, Hydrozoa): молекулярная оценка» . Журнал зоологической систематики и эволюционных исследований . 45 (1): 11–19. дои : 10.1111/j.1439-0469.2006.00379.x .
- ^ Пираино, С.; Боэро, Ф.; Эшбах, Б.; Шмид, В. (1996). «Обращение жизненного цикла: медузы превращаются в полипы и трансдифференцировка клеток у Turritopsis nutricula (Cnidaria, Hydrozoa)». Биологический бюллетень . 190 (3): 302–312. дои : 10.2307/1543022 . JSTOR 1543022 . ПМИД 29227703 . S2CID 3956265 .
- ^ Рати, Акшат (15 мая 2014 г.). «Медузы — самые энергоэффективные пловцы, подтверждает новый показатель» . Арс Техника . Архивировано из оригинала 3 ноября 2014 года . Проверено 3 декабря 2014 г.
- ^ Джеммелл, Би Джей; Костелло, Дж. Х.; Колин, СП; и др. (2013). «Пассивный возврат энергии медузами способствует их движущему преимуществу перед другими многоклеточными животными» . Труды Национальной академии наук . 110 (44): 17904–17909. Бибкод : 2013PNAS..11017904G . дои : 10.1073/pnas.1306983110 . ПМЦ 3816424 . ПМИД 24101461 .
- ^ Бруска, Ричард (2016). Беспозвоночные . Синауэр Ассошиэйтс. п. 296. ИСБН 978-1-60535-375-3 .
Все книдарии — хищники (или паразиты). Обычно питающиеся щупальца, наполненные нематоцистами, захватывают добычу животного и несут ее в область рта, где она проглатывается целиком.
- ^ «Большие медузы, унаследовавшие океан, показали исследования» . msnbc.com . 15 сентября 2011 года. Архивировано из оригинала 14 июля 2014 года . Проверено 3 декабря 2014 г.
- ^ Дэвис, Швейцария; Слотвински, А.С. «Австралийский морской зоопланктон-медузы, кладоцеры» (PDF) . Институт морских и антарктических исследований Университета Тасмании . Проверено 12 октября 2018 г.
- ^ Доусон, Майкл Н. (2000). «Пестрые мезокосмы как альтернатива береговым планктонкрейселям: заметки о разведении медуз из морских озер» . Журнал исследований планктона . 22 (9): 1673–1682. дои : 10.1093/планкт/22.9.1673 .
- ^ Jump up to: Перейти обратно: а б Доусон, Майк Н.; Мартин, Лаура Э.; Лолита К., Лолита К.; Пенленд (май 2001 г.). Стаи медуз, туристы и Младенец Христос . Том. 451. Спрингер. стр. 131–144. дои : 10.1023/А:1011868925383 . ISBN 978-0-7923-6964-6 . S2CID 1679552 .
{{cite book}}:|journal=игнорируется ( помогите ) - ^ Немо, Лесли (13 февраля 2020 г.). «Ядовитые сопли помогают этим медузам жалить без щупалец» . Обнаружить . Проверено 11 июля 2021 г.
- ^ Инь, Стеф (29 сентября 2017 г.). «Кто ест медуз? Пингвины, вот кто» . Нью-Йорк Таймс . Архивировано из оригинала 1 октября 2017 года . Проверено 4 октября 2017 г.
- ^ Тибот, Жан-Батист; Арнульд, Джон П.И.; Гомес-Лайх, Агустина; и др. (2017). «Медузы и другие виды джелаты как пища для четырех видов пингвинов – выводы из видеороликов, снятых хищниками». Границы в экологии и окружающей среде . 15 (8): 437–441. Бибкод : 2017FrEE...15..437T . дои : 10.1002/плата.1529 . S2CID 90152409 .
- ^ Гершвин, Лиза-Энн (2016). Медуза: естественная история . Издательство Чикагского университета. п. 140. ИСБН 978-0-226-28767-6 .
- ^ Гершвин, Лиза-Энн (2013). Ужалено!: О цветении медуз и будущем океана . Издательство Чикагского университета. стр. 274–. ISBN 978-0-226-02010-5 . Архивировано из оригинала 6 августа 2016 года.
- ^ колуго7 (2006). «Медуза» . Веб-проект «Древо жизни» . Архивировано из оригинала 12 июня 2018 года . Проверено 7 июня 2018 г.
{{cite web}}: CS1 maint: числовые имена: список авторов ( ссылка ) - ^ Гриффин, ДюБоз Б.; Мерфи, Томас М. «Медуза-пушечное ядро» (PDF) . Департамент природных ресурсов Южной Каролины. Архивировано из оригинала (PDF) 19 января 2008 года . Проверено 7 июня 2018 г.
- ^ Jump up to: Перейти обратно: а б Бротц, Лукас; Чунг, Уильям В.Л.; Кляйснер, Кристин; и др. (2012). «Увеличение популяции медуз: тенденции в крупных морских экосистемах» . Гидробиология . 688 : 3–20. дои : 10.1007/s10750-012-1039-7 .
- ^ Джилл, Виктория. «Медуза «чувствует океанские течения» » . Новости Би-би-си. Архивировано из оригинала 26 января 2015 года . Проверено 26 января 2015 г.
- ^ Хейс, Грэм К. (2017). «Океанские течения и морская жизнь» . Современная биология . 27 (11): 470–473 р. Бибкод : 2017CBio...27.R470H . дои : 10.1016/j.cub.2017.01.044 . hdl : 10536/DRO/DU:30100274 . ПМИД 28586681 .
- ^ Шубин, Кристи (10 декабря 2008 г.). «Антропогенные факторы, связанные с цветением медуз – окончательный вариант II» . Курсы тропических полей: Западная программа: Университет Майами. Архивировано из оригинала 14 июня 2010 года . Проверено 19 ноября 2009 г.
- ^ «Что такое мертвая зона?» . Национальная океаническая служба . Проверено 3 октября 2018 г.
- ^ Йонг, Эд (6 июня 2011 г.). «Медузы меняют пищевые сети океана, питая бактерии слизью и экскрементами» . Откройте для себя журнал. Архивировано из оригинала 6 ноября 2018 года . Проверено 3 октября 2018 г.
- ^ Jump up to: Перейти обратно: а б «Цветение медуз может быть признаком больного моря» . www.eurocbc.org . Проверено 10 февраля 2023 г.
- ^ Хейс, GC; Бастиан, Т.; Дойл, ТК; и др. (2011). «Высокая активность и поиск Леви: медузы могут искать в толще воды, как рыбы» (PDF) . Труды Королевского общества Б. 279 (1728): 465–473. дои : 10.1098/rspb.2011.0978 . ПМК 3234559 . ПМИД 21752825 . Архивировано (PDF) из оригинала 25 декабря 2012 года.
- ^ Поли, Д.; Кристенсен, В.; Далсгаард, Дж.; и др. (1998). «Лов рыбы в морских пищевых сетях» (PDF) . Наука . 279 (5352): 860–863. Бибкод : 1998Sci...279..860P . дои : 10.1126/science.279.5352.860 . ПМИД 9452385 . Архивировано (PDF) из оригинала 10 июля 2012 года.
- ^ Ричардсон, Эй Джей; Бакун А.; Хейс, GC; Гиббонс, MJ (2009). «Поездка на медузе: причины, последствия и реакция руководства на более студенистое будущее» (PDF) . Тенденции в экологии и эволюции . 24 (6): 312–322. Бибкод : 2009TEcoE..24..312R . дои : 10.1016/j.tree.2009.01.010 . ПМИД 19324452 . [ постоянная мертвая ссылка ]
- ^ Акснес, Д.Л.; Нейстгаард, Дж.; Сэдберг, Э.; Сёрнес, Т. (2004). «Оптический контроль популяций рыб и зоопланктона» . Лимнология и океанография . 49 (1): 233–238. Бибкод : 2004LimOc..49..233A . дои : 10.4319/lo.2004.49.1.0233 .
- ^ Линам, CP; Гиббонс, MJ; Аксельсен, Бельгия; и др. (2006). «Медузы обгоняют рыбу в экосистеме, где активно ведется рыбная ловля» (PDF) . Современная биология . 16 (13): 492–493. Бибкод : 2006CBio...16.R492L . дои : 10.1016/j.cub.2006.06.018 . ПМИД 16824906 . S2CID 62793057 . Архивировано (PDF) из оригинала 23 марта 2011 года.
- ^ Поли, Д.; Грэм, В.; Либралато, С.; и др. (2009). «Медузы в экосистемах, онлайн-базы данных и модели экосистем» . Гидробиология . 616 : 67–85. дои : 10.1007/s10750-008-9583-x . S2CID 12415790 . Архивировано из оригинала (PDF) 12 июля 2013 года.
- ^ Доусон, Миннесота; Сен Гупта, А.; Англия, Миннесота (2005). «Совместная биофизическая модель глобального океана и молекулярно-генетический анализ позволяют выявить многочисленные интродукции криптогенных видов» . Учеб. Натл. акад. наук. США . 102 (34): 11968–73. Бибкод : 2005PNAS..10211968D . дои : 10.1073/pnas.0503811102 . ПМК 1189321 . ПМИД 16103373 .
- ^ Доусон, Миннесота (2003). «Макроморфологические вариации загадочных видов лунной медузы Aurelia (Cnidaria: Scyphozoa)». Морская биология . 143 (2): 369–79. Бибкод : 2003МарБи.143..369D . дои : 10.1007/s00227-003-1070-3 . S2CID 189820003 .
- ^ Jump up to: Перейти обратно: а б Ринат, Зафрир (15 июня 2009 г.). «Самая агрессивная медуза в мире распространяется вдоль побережья Израиля» . Гаарец . Проверено 13 октября 2018 г.
- ^ Бриан, Фредерик; Боэро, Фердинандо (2001). «Вспышки студенистого зоопланктона – обзор цветения медуз» . Монографии ЦИЕСМ . 14 :5-17.
- ^ Миллс, CE (2001). «Цветение медуз: увеличивается ли популяция во всем мире в ответ на изменение условий океана?» (PDF) . Гидробиология . 451 : 55–68. дои : 10.1023/А:1011888006302 . S2CID 10927442 . Архивировано (PDF) из оригинала 3 марта 2016 г.
- ^ «ЦИЭСМ ГИС» . www.ciesm.org . Проверено 10 февраля 2023 г.
- ^ Перселл, Дж.; Арай, М. (2001). Перселл, Дж. Э; Грэм, WM; Дюмон, HJ (ред.). «Взаимодействие пелагических книдарий и гребневиков с рыбами: обзор». Гидробиология . 541 : 27–44. дои : 10.1007/978-94-010-0722-1 . ISBN 978-94-010-3835-5 . S2CID 27615539 .
- ^ Бродер, Ричард Д.; Линк, Джейсон С.; Смит, Б.Э.; и др. (2016). «Экологические и экономические последствия игнорирования медуз: призыв к усилению мониторинга экосистем». Рыболовство . 41 (11): 630–637. Бибкод : 2016Рыба...41..630Б . дои : 10.1080/03632415.2016.1232964 .
- ^ Ружичка, Джей Джей; Бродер, Р.Д.; Эммет, РЛ; и др. (2012). «Межгодовая изменчивость структуры пищевой сети Северной Калифорнии: изменения в путях потоков энергии и роль кормовой рыбы, эвфаузиид и медуз». Прогресс в океанографии . 102 : 19–41. Бибкод : 2012Proce.102...19R . дои : 10.1016/j.pocean.2012.02.002 .
- ^ Jump up to: Перейти обратно: а б Питт, Кайли; Валлийский, Дэвид; Кондон, Роберт (январь 2009 г.). «Влияние цветения медуз на круговорот углерода, азота и фосфора и производство планктона». Гидробиология . 616 : 133–149. дои : 10.1007/s10750-008-9584-9 . S2CID 22838905 .
- ^ Кондон, Роберт Х.; Дуарте, Карлос М.; Питт, Кайли А.; и др. (15 января 2013 г.). «Периодическое цветение медуз является следствием глобальных колебаний» . Труды Национальной академии наук . 110 (3): 1000–1005. Бибкод : 2013PNAS..110.1000C . дои : 10.1073/pnas.1210920110 . ПМК 3549082 . ПМИД 23277544 .
- ^ Линам, Кристофер П.; Гиббонс, Марк Дж.; Аксельсен, Бьёрн Э.; и др. (11 июля 2006 г.). «Медузы догоняют рыбу в экосистеме, где активно ведется рыбная ловля» . Современная биология . 16 (13): 492–493 рэндов. Бибкод : 2006CBio...16.R492L . дои : 10.1016/j.cub.2006.06.018 . ПМИД 16824906 . S2CID 62793057 .
- ^ Масиламани, Дж; Джесудосс, К; Канавиллил, Нандакумар; и др. (10 сентября 2000 г.). «Проникновение медуз: угроза бесперебойной работе береговых электростанций» . Современная наука . 79 : 567–569.
- ^ Перселл, Дженнифер Э.; Уе, Синъити; Ло, Вен-Ценг (22 ноября 2007 г.). «Антропогенные причины цветения медуз и их прямые последствия для человека: обзор» . Серия «Прогресс в области морской экологии» . 350 : 153–174. Бибкод : 2007MEPS..350..153P . дои : 10.3354/meps07093 .
- ^ Свитман, Эндрю К.; Смит, Крейг Р.; Дейл, Трина; Джонс, Дэниел О.Б. (2014). «Быстрое очищение туш медуз показывает важность студенистого материала для глубоководных пищевых сетей» . Труды Королевского общества B: Биологические науки . 281 (1796): 20142210. doi : 10.1098/rspb.2014.2210 . ПМЦ 4213659 . ПМИД 25320167 .
- ^ Лебрато, Марио; Пахлоу, Маркус; Ошлис, Андреас; и др. (2011). «Снижение экспорта органических веществ на глубину, связанное с падением желе» (PDF) . Лимнология и океанография . 56 (5): 1917–1928. Бибкод : 2011LimOc..56.1917L . дои : 10.4319/lo.2011.56.5.1917 . hdl : 10072/43275 . S2CID 3693276 .
- ^ Диджюлис, Викторас. «Информационный бюллетень об инвазивных чужеродных видах: Craspedacusta sowerbyi » (PDF) . НОБАНИС. Архивировано (PDF) из оригинала 17 мая 2014 года . Проверено 16 июня 2016 г.
- ^ Доусон, Майк Н.; Мартин, Лаура Э.; Пенленд, Лолита К. (2001). Стаи медуз, туристы и Младенец Христос . Том. 451. стр. 131–144. дои : 10.1023/А:1011868925383 . ISBN 978-0-7923-6964-6 . S2CID 1679552 .
{{cite book}}:|journal=игнорируется ( помогите ) - ^ Миллс, CE; Хирано, Ю.М. (2007). «Стауромедузы». Энциклопедия приливных озер и скалистых берегов : 541–543.
- ^ Кондо, Юсуке; Оцука, Сусуму; Хирабаяси, Такеши; и др. (2016). «Сезонные изменения в инфекции видами трематод, использующими медуз в качестве хозяев: свидетельства передачи окончательному хозяину-рыбе через медузивных животных» . Паразит . 23:16 . doi : 10.1051/parasite/2016016 . ПМЦ 4824873 . ПМИД 27055563 . Архивировано из оригинала 4 ноября 2017 года.
- ^ Люнг, Томми (26 мая 2016 г.). « Опечона олссони » . Блог: Паразит дня. Архивировано из оригинала 30 июня 2016 года . Проверено 1 июня 2016 г.
- ^ «ФАОСТАТ» . www.фао.орг . Проверено 10 февраля 2023 г.
- ^ Jump up to: Перейти обратно: а б с д и Се, Ю.Х. Пегги; Леонг, Фуй-Мин; Радлоу, Джек (2001). «Медуза как еда». Гидробиология . 451 (1–3): 11–17. дои : 10.1023/А:1011875720415 . S2CID 20719121 .
- ^ Джордж, Алета (1 ноября 2012 г.). «Желе в центре внимания» . Эндокринные новости . Эндокринное общество . Проверено 16 июня 2018 г.
- ^ Клинтон, Мораг; Ферье, Дэвид ЭК; Мартин, Сэмюэл AM; Бриерли, Эндрю С. (2 апреля 2021 г.). Байрон, Кэрри (ред.). «Воздействие медуз на морскую садковую аквакультуру: обзор существующих знаний и проблем для здоровья рыб». Журнал морских наук ICES . 78 (5). Международный совет по исследованию моря ( ОУП ): 1557–1573 гг. doi : 10.1093/icesjms/fsaa254 . hdl : 10023/25136 . ISSN 1054-3139 .
- ^ Бош-Бельмар, Мар; Милисенда, Джакомо; Бассо, Лорена; и др. (3 сентября 2020 г.). «Влияние медуз на морскую аквакультуру и рыболовство» . Обзоры в области рыболовства и аквакультуры . 29 (2). Тейлор и Фрэнсис : 242–259. дои : 10.1080/23308249.2020.1806201 . HDL : 10447/531140 . ISSN 2330-8249 . S2CID 225330785 .
- ^ Аристотель ; Уильям Огл (пер.) (2018). Части животных . Том. IV. п. 6. ISBN 9782378989842 . [ постоянная мертвая ссылка ]
- ^ Омори, М.; Накано, Э. (2001). «Промысел медуз в Юго-Восточной Азии». Гидробиология . 451 : 19–26. дои : 10.1023/А:1011879821323 . S2CID 6518460 .
- ^ Ферт, FE (1969). Энциклопедия морских ресурсов . Ван Ностранд Рейнхольд. ISBN 978-0-442-22399-1 .
- ^ «Как желе засияло» . Американский музей естественной истории . Архивировано из оригинала 12 июня 2018 года . Проверено 11 июня 2018 г.
- ^ Jump up to: Перейти обратно: а б Шимомура, О.; Джонсон, Ф.Х.; Сайга, Ю. (1962). «Извлечение, очистка и свойства экворина, биолюминесцентного белка из светящегося гидромедузана Aequorea ». Журнал клеточной и сравнительной физиологии . 59 (3): 223–39. дои : 10.1002/jcp.1030590302 . ПМИД 13911999 .
- ^ Прашер, округ Колумбия; Экенроде, В.К.; Уорд, WW; и др. (1992). «Первичная структура зеленого флуоресцентного белка Aequorea victoria». Джин . 111 (2): 229–33. дои : 10.1016/0378-1119(92)90691-H . ПМИД 1347277 .
- ^ Чалфи, М.; Ту, Ю.; Ойскирхен, Г.; и др. (февраль 1994 г.). «Зеленый флуоресцентный белок как маркер экспрессии генов». Наука . 263 (5148): 802–5. Бибкод : 1994Sci...263..802C . дои : 10.1126/science.8303295 . ПМИД 8303295 . S2CID 9043327 .
- ^ Пиерибоне, В.; Грубер, Д.Ф. (2006). Сияние в темноте: революционная наука биофлуоресценции . Издательство Гарвардского университета. ISBN 978-0-674-02413-7 .
- ^ Сельдь, Питер (2002). Биология глубокого океана Издательство Оксфордского университета . Издательство Оксфордского университета. стр. 190–191 . ISBN 978-0-19-854956-7 .
- ^ «Патент США на аквариум с медузами» . Архивировано из оригинала 20 февраля 2015 года.
- ^ Рихтель, Мэтт (14 марта 2009 г.). «Как избежать разжижения медузы» . Нью-Йорк Таймс . Архивировано из оригинала 26 марта 2010 года . Проверено 6 мая 2010 г.
- ^ МакГи, Ричард Г.; Вебстер, Анджела С .; Льюис, Шэрон Р.; Уэлсфорд, Мишель (5 июня 2023 г.). «Вмешательства при симптомах и признаках укусов медуз» . Кокрановская база данных систематических обзоров . 2023 (6): CD009688. дои : 10.1002/14651858.CD009688.pub3 . ISSN 1469-493X . ПМК 10240560 . ПМИД 37272501 .
- ^ Первес, ВК; Садава, Д.; Орианс, Г.Х.; Хеллер, ХК 1998. Жизнь. Наука биологии. Часть 4: Эволюция разнообразия. Глава 31
- ^ «Аквариумы с медузами и живые домашние медузы на продажу в Jellyfish Art — покупайте медуз и аквариумы с медузами» . jellyfishart.com . Архивировано из оригинала 2 марта 2012 года . Проверено 3 декабря 2014 г.
- ^ Джаймо, Кара (13 февраля 2020 г.). «Вы не трогали этих медуз, но они могут ужалить вас крошечными гранатами» . Нью-Йорк Таймс . ISSN 0362-4331 . Проверено 27 февраля 2020 г.
- ^ Махон, Эндрю; Мэллинсон, Том Э (2020). «Укус медузы из львиной гривы». Международная парамедицинская практика . 10 (2): 46–48. дои : 10.12968/ippr.2020.10.2.46 . ISSN 2052-4889 . S2CID 225837493 .
- ^ Jump up to: Перейти обратно: а б с Такер, Эбигейл (июль 2010 г.). «Новый король моря». Смитсоновский институт . 54 (4): 540–561. дои : 10.1177/1363461517722869 . ПМИД 28752797 . S2CID 12532183 .
- ^ Адамс, Джули (13 августа 2016 г.). «Коробчатые медузы: почему они такие смертоносные?» . Наша прекрасная планета. Архивировано из оригинала 10 сентября 2016 года . Проверено 17 сентября 2016 г.
- ^ Jump up to: Перейти обратно: а б Феннер, П.; Уильямсон, Дж.; Бернетт, Дж.; Рифкин, Дж. (1993). «Первая помощь при укусах медуз в Австралии. Реакция на новый дифференцированный вид». Медицинский журнал Австралии . 158 (7): 498–501. дои : 10.5694/j.1326-5377.1993.tb137588.x . ПМИД 8469205 . S2CID 42453046 .
- ^ Карри, Б.; Хо, С.; Олдерслейд, П. (1993). «Коробочка-медуза, кока-кола и старое вино». Медицинский журнал Австралии . 158 (12): 868. doi : 10.5694/j.1326-5377.1993.tb137688.x . ПМИД 8100984 . S2CID 19857333 .
- ^ Хартвик, Р.; Калланан, В.; Уильямсон, Дж. (1980). «Обезвреживание коробомедузы: ингибирование нематоциста у Chironex fleckeri ». Медицинский журнал Австралии . 1 (1): 15–20. дои : 10.5694/j.1326-5377.1980.tb134566.x . ПМИД 6102347 . S2CID 204054168 .
- ^ Jump up to: Перейти обратно: а б Перкинс, Р.; Морган, С. (2004). «Отравление, отравление и травмы морских существ». Американский семейный врач . 69 (4): 885–90. ПМИД 14989575 .
- ^ Симмонс, Брайан Дж.; Гриффит, Роберт Д.; Фальто-Айзпуруа, Лейр А.; Нури, Кейван (2015). «Укусы лунной медузы» . JAMA Дерматология . 151 (4): 454–6. дои : 10.1001/jamadermatol.2014.4644 . ПМИД 25517656 .
- ^ Бакстер, Э.Х.; Марр, годовое общее собрание акционеров (май 1974 г.). «Противовенен морской осы ( Chironex fleckeri ): нейтрализующая эффективность против яда трех других видов медуз». Токсикон . 12 (3): 223–225. Бибкод : 1974Txcn...12..223B . дои : 10.1016/0041-0101(74)90062-2 . ПМИД 4156430 .
- ^ «Укусы медуз: лечение и лекарства» . Клиника Мэйо . Фонд Мэйо медицинского образования и исследований. 1 сентября 2011 года. Архивировано из оригинала 20 мая 2013 года . Проверено 15 апреля 2013 г.
- ^ Jump up to: Перейти обратно: а б «Медуза одичала — только текст» . Nsf.gov. Архивировано из оригинала 12 июля 2010 года . Проверено 18 октября 2010 г.
- ^ «Отчет об уведомлении о текущих событиях» . НРК. 22 октября 2008 г. Архивировано из оригинала 5 июня 2011 г. Проверено 14 июля 2010 г.
- ^ Райалл, Джулиан (2 ноября 2009 г.). «Японский рыболовный траулер, потопленный гигантской медузой» . Лондон: Telegraph.co.uk. Архивировано из оригинала 4 ноября 2009 года.
Дальнейшее чтение
- Джули Бервальд (2017). Бесхребетный: наука о медузах и искусство выращивания позвоночника . Книги Риверхеда. ISBN 978-0-7352-1126-1 .
Внешние ссылки
- Медузы и гребневики - Смитсоновский океанический портал
- Факты о медузах – информация о медузах и безопасности медуз
- « Медузы не существует » с MBARI . YouTube-канала
- «Порочные красавицы – Медузы» – документальный фильм о медузах.
- Они захватывают власть! nybooks.com, 26 сентября 2013 г. Тим Флэннери.
- Фотографии
Существующие Cnidaria классы |
|---|
