Эволюционная биология развития
Эволюционная биология развития (неформально, эво-дево ) — это область биологических исследований , которая сравнивает процессы развития различных организмов , чтобы сделать вывод о том, как развивались процессы развития .
Эта область развивалась с начала XIX века, когда эмбриология столкнулась с загадкой: зоологи не знали, как эмбриональное развитие контролируется на молекулярном уровне. Чарльз Дарвин отметил, что наличие схожих эмбрионов предполагает общее происхождение, но до 1970-х годов прогресс был незначительным. Затем технология рекомбинантной ДНК наконец объединила эмбриологию с молекулярной генетикой . Ключевым ранним открытием были гомеозисные гены , которые регулируют развитие у широкого круга эукариот .
Эта область состоит из множества основных эволюционных концепций. Одним из них является глубокая гомология : открытие того, что разные органы, такие как глаза насекомых , позвоночных и головоногих моллюсков, которые, как долгое время считалось, развивались отдельно, контролируются сходными генами, такими как pax-6 , из набора генов evo-devo . Эти гены являются древними и высококонсервативными среди типов ; они создают закономерности во времени и пространстве, которые формируют эмбрион и в конечном итоге формируют план тела организма. Во-вторых, виды не сильно различаются по своим структурным генам, например, по генам, кодирующим ферменты ; Что действительно отличается, так это то, как экспрессия генов регулируется набором генов . Эти гены многократно используются в неизмененном виде в разных частях эмбриона и на разных стадиях развития, образуя сложный каскад контроля, включающий и выключающий другие регуляторные гены, а также структурные гены по точной схеме. Это многократное плейотропное повторное использование объясняет, почему эти гены высоко консервативны, поскольку любое изменение будет иметь множество неблагоприятных последствий, которые естественный отбор будет против.
Новые морфологические особенности и, в конечном итоге, новые виды возникают в результате изменений в наборе инструментов, либо когда гены экспрессируются в новом паттерне, либо когда гены набора инструментов приобретают дополнительные функции. Другой возможностью является неоламаркианская теория о том, что эпигенетические изменения позже закрепляются на уровне генов , что могло иметь важное значение на ранних этапах истории многоклеточной жизни.
История [ править ]
Ранние теории
Философы начали задумываться о том, как животные приобретали форму еще в утробе матери еще в классической античности . Аристотель утверждает в своем трактате по физике , что, по мнению Эмпедокла , порядок «спонтанно» возникает в развивающемся зародыше. В своем трактате «Части животных » он утверждает, что теория Эмпедокла ошибочна. В рассказе Аристотеля Эмпедокл утверждал, что позвоночный столб делится на позвонки, потому что при этом эмбрион скручивается и разрывает позвоночник на части. Вместо этого Аристотель утверждает, что у этого процесса есть заранее определенная цель: что «семя», из которого развивается эмбрион, изначально имело встроенный «потенциал» стать определенными частями тела, такими как позвонки. Далее, каждый вид животных порождает животных себе подобных: у человека рождаются только человеческие детеныши. [1]
Перепросмотр [ править ]
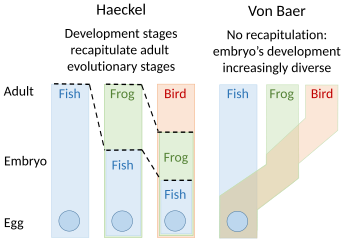
Теория перепросмотра эволюционного развития была предложена Этьеном Серром в 1824–1826 годах, перекликаясь с идеями 1808 года Иоганна Фридриха Меккеля . Они утверждали, что эмбрионы «высших» животных прошли или повторили ряд стадий, каждая из которых напоминала животное, стоящее ниже в великой цепи бытия . Например, мозг человеческого эмбриона сначала выглядел как мозг рыбы , затем, в свою очередь, как мозг рептилии , птицы и млекопитающего , прежде чем стать явно человеческим . Эмбриолог Карл Эрнст фон Бэр выступил против этого, утверждая в 1828 году, что не существует линейной последовательности, как в великой цепи бытия, основанной на едином плане тела , а есть процесс эпигенеза , в котором дифференцируются структуры. Вместо этого фон Баер выделил четыре различных строения тела животных : лучистое, как у морской звезды ; моллюск, похожий на моллюсков ; членораздельно, как омары ; и позвоночные, такие как рыбы. Зоологи тогда в значительной степени отказались от перепросмотра, хотя Эрнст Геккель возродил его в 1866 году. [3] [4] [5] [6] [7]
Эволюционная морфология [ править ]

С начала 19 века и на протяжении большей части 20 века эмбриология сталкивалась с загадкой. Было замечено, что животные развивались во взрослых особей с совершенно разным строением тела , часто проходя схожие стадии, из яйца, но зоологи почти ничего не знали о том, как эмбриональное развитие контролируется на молекулярном уровне , и, следовательно, столь же мало о том, как процессы развития . развивались [8] Чарльз Дарвин утверждал, что общая эмбриональная структура предполагает наличие общего предка. Например, Дарвин цитировал в своей книге 1859 года «Происхождение видов» , сидячие ракушки креветкообразную личинку совсем взрослые которой особи не были похожи на других членистоногих ; Линней и Кювье отнесли их к моллюскам . [9] [10] Дарвин также отметил Александра Ковалевского открытие о том, что оболочник тоже не был моллюском, но на личиночной стадии имел хорду и глоточные щели, которые развились из тех же зародышевых листков, что и эквивалентные структуры у позвоночных , и поэтому их следует отнести к группе они как хордовые . [9] [11]
Таким образом, зоология XIX века превратила эмбриологию в эволюционную науку, связав филогению с гомологиями между зародышевыми листками эмбрионов. Зоологи, в том числе Фриц Мюллер, предложили использовать эмбриологию для обнаружения филогенетических связей между таксонами. Мюллер продемонстрировал, что ракообразные имеют общую личинку Nauplius , идентифицировав несколько видов паразитов, которые не были признаны ракообразными. Мюллер также признал, что естественный отбор должен действовать на личинок так же, как и на взрослых особей, опровергая идею рекапитуляции, которая требует защиты личиночных форм от естественного отбора. [9] Две другие идеи Геккеля об эволюции развития оказались более успешными, чем перепросмотр: в 1870-х годах он утверждал, что изменения во времени ( гетерохрония ) и изменения в расположении внутри тела ( гетеротопия ) аспектов эмбрионального развития будут стимулировать эволюцию, изменяя форма тела потомка по сравнению с формой тела предка. Прошло столетие, прежде чем эти идеи оказались верными. [12] [13] [14]
В 1917 году Д'Арси Томпсон написал книгу о формах животных , показав с помощью простой математики , как небольшие изменения параметров , таких как углы спиральной раковины брюхоногих моллюсков , могут радикально изменить форму животного , хотя он предпочитал механическую форму. эволюционному объяснению. [15] [16] Но без молекулярных доказательств прогресс застопорился. [9]

В 1952 году Алан Тьюринг опубликовал свою статью « Химическая основа морфогенеза », посвященную развитию закономерностей в телах животных. Он предположил, что морфогенез можно объяснить системой реакции-диффузии , системой реагирующих химических веществ, способных диффундировать по телу. [17] Он смоделировал катализируемые химические реакции, используя уравнения в частных производных , показав, что закономерности возникают, когда в результате химической реакции образуется как катализатор ( А), так и ингибитор (В), которые замедляют производство А. Если А и В затем диффундируют с разной скоростью, А доминирует. в некоторых местах и B в других. Российский биохимик Борис Белоусов проводил эксперименты с аналогичными результатами, но не смог их опубликовать, поскольку в то время ученые считали, что создание видимого порядка нарушает второй закон термодинамики . [18]
Современный синтез начала 20 века [ править ]
В так называемом современном синтезе начала 20-го века, между 1918 и 1930 годами, Рональд Фишер объединил теорию эволюции Дарвина с ее упором на естественный отбор, наследственность и изменчивость и Грегора Менделя в законы генетики последовательную структуру. для эволюционной биологии . Биологи предположили, что организм является прямым отражением составляющих его генов: генов, кодирующих белки, которые строят тело организма. Биохимические пути (и, как они предполагали, новые виды) возникли в результате мутаций в этих генах. Это была простая, ясная и почти полная картина, но она не объясняла эмбриологию. [9] [19] Шон Б. Кэрролл заметил, что если бы идеи эво-дево были доступны, эмбриология, безусловно, сыграла бы центральную роль в синтезе. [20]
Эволюционный эмбриолог Гэвин де Бир предвосхитил эволюционную биологию развития в своей книге 1930 года «Эмбрионы и предки» . [21] показав, что эволюция может происходить гетерохронно , [22] например, при сохранении ювенильных черт у взрослых . [12] Это, как утверждал де Бир, могло вызвать внезапные изменения в летописи окаменелостей , поскольку эмбрионы окаменевают плохо. Поскольку пробелы в летописи окаменелостей использовались в качестве аргумента против постепенной эволюции Дарвина, объяснение де Бира поддержало дарвиновскую позицию. [23] Однако, несмотря на де Бира, современный синтез в значительной степени игнорировал эмбриональное развитие для объяснения формы организмов, поскольку популяционная генетика оказалась адекватным объяснением того, как развивались формы. [24] [25] [а]
Лак-оперон [ править ]

1 : РНК-полимераза , 2 : репрессор , 3 : промотор , 4 : оператор, 5 : лактоза , 6–8 : гены, кодирующие белок , контролируемые переключателем, которые вызывают переваривание лактозы.
В 1961 году Жак Моно , Жан-Пьер Шанже и Франсуа Жакоб открыли лак-оперон у бактерии Escherichia coli . Это был кластер генов , организованный в петлю управления с обратной связью , так что его продукты производились только при «включении» стимулом окружающей среды. Одним из таких продуктов был фермент, расщепляющий сахар , лактозу; а сама лактоза была стимулом, который включил гены. Это было открытием, поскольку оно впервые показало, что гены, даже в таких маленьких организмах, как бактерия, подлежат точному контролю. Подразумевалось, что многие другие гены также тщательно регулируются. [27]
Рождение эво-дево и второй синтез [ править ]
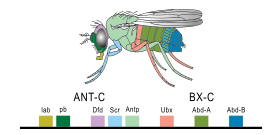
В 1977 году началась революция в представлении об эволюции и биологии развития с появлением технологии рекомбинантной ДНК в генетике , книги Онтогенез и филогения» « Стивена Дж. Гулда и статьи «Эволюция и мастерство». [28] Франсуа Жакоб . Гулд положил конец интерпретации Геккеля эволюционной эмбриологии, а Джейкоб изложил альтернативную теорию. [9] Это привело ко второму синтезу , [29] [30] наконец, включая эмбриологию, а также молекулярную генетику , филогению и эволюционную биологию, чтобы сформировать эво-дево. [31] [32] В 1978 году Эдвард Б. Льюис обнаружил гомеозисные гены, регулирующие эмбриональное развитие у -дрозофил плодовых мух , которые, как и все насекомые, являются членистоногими , одним из основных типов беспозвоночных животных. [33] Билл МакГиннис быстро обнаружил последовательности гомеотических генов, гомеобоксы , у животных других типов, у позвоночных , таких как лягушки , птицы и млекопитающие ; позже они были также обнаружены в грибах, таких как дрожжи , и в растениях . [34] [35] Очевидно, существовало сильное сходство в генах, которые контролировали развитие у всех эукариот . [36] В 1980 году Кристиана Нюсляйн-Фольхард и Эрик Вишаус описали гены пробелов , которые помогают создать структуру сегментации у эмбрионов плодовых мух ; [37] [38] они и Льюис получили Нобелевскую премию за свою работу в 1995 году. [34] [39]
Позже были обнаружены более конкретные сходства: например, в 1989 году было обнаружено, что ген Distal-less участвует в развитии придатков или конечностей у плодовых мух. [40] плавники рыб, крылья кур, параподии морских кольчатых червей, ампулы и сифоны оболочников и трубчатые ножки морских ежей . Было очевидно, что этот ген должен быть древним и восходить к последнему общему предку двусторонних животных (до эдиакарского периода, который начался около 635 миллионов лет назад). Эво-Дево начал открывать способы построения тел животных в процессе развития. [41] [42]
Контроль структуры тела [ править ]
Глубокая гомология [ править ]
Примерно сферические яйца разных животных дают начало уникальной морфологии: от медуз до омаров, от бабочек до слонов. Многие из этих организмов имеют одни и те же структурные гены для белков бодибилдинга, таких как коллаген и ферменты, но биологи ожидали, что у каждой группы животных будут свои собственные правила развития. Неожиданностью эво-дево является то, что формирование тела контролируется довольно небольшим процентом генов и что эти регуляторные гены являются древними и общими для всех животных. У жирафа нет гена длинной шеи, как и у слона нет гена большого тела. В их телах действует система переключений, благодаря которой развитие различных признаков начинается раньше или позже, происходит в той или иной части эмбриона и продолжается большее или меньшее время. [8]
Загадку того, как контролируется эмбриональное развитие, начали решать, используя плодовую мушку Drosophila melanogaster в качестве модельного организма . Поэтапный контроль его эмбриогенеза был визуализирован путем прикрепления флуоресцентных красителей разного цвета к определенным типам белков, вырабатываемых генами, экспрессируемыми в эмбрионе. [8] Краситель, такой как зеленый флуоресцентный белок , полученный из медузы , обычно присоединялся к антителу, специфичному к белку плодовой мухи, образуя точный индикатор того, где и когда этот белок появился в живом эмбрионе. [43]

Используя такой метод, в 1994 году Уолтер Геринг обнаружил, что ген pax-6 , жизненно важный для формирования глаз плодовых мух, в точности соответствует гену, формирующему глаза у мышей и людей. Тот же ген был быстро обнаружен у многих других групп животных, таких как кальмары , головоногие моллюски . Биологи, в том числе Эрнст Майр, считали, что глаза возникали в животном мире как минимум 40 раз, поскольку анатомия разных типов глаз сильно различается. [8] Например, сложный глаз плодовой мухи состоит из сотен мелких линзообразных структур ( омматидиев ); человеческий глаз имеет слепое пятно там, где зрительный нерв входит в глаз, а нервные волокна проходят по поверхности сетчатки , поэтому свет должен пройти через слой нервных волокон, прежде чем достичь детекторных клеток сетчатки, поэтому структура фактически «перевернут»; Напротив, у головоногих глаз есть сетчатка, затем слой нервных волокон, а затем стенка глаза «правильно». [44] Однако свидетельством существования pax-6 было то, что одни и те же гены контролировали развитие глаз всех этих животных, что позволяет предположить, что все они произошли от общего предка. [8] Древние гены сохранялись на протяжении миллионов лет эволюции , создавая разнородные структуры для схожих функций, демонстрируя глубокую гомологию между структурами, которые когда-то считались чисто аналогичными. [45] [46] Позднее это представление было распространено на эволюцию эмбриогенеза. [47] и вызвало радикальный пересмотр значения гомологии в эволюционной биологии. [45] [46] [20]
Генный инструментарий [ править ]
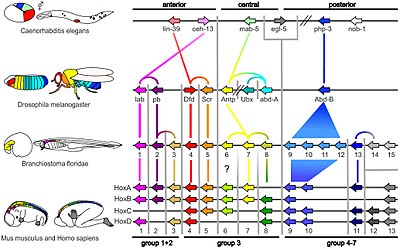
Небольшая часть генов в геноме организма контролирует развитие организма. Эти гены называются набором генетических инструментов развития. Они высоко консервативны среди типов , а это означает, что они древние и очень похожи в широко разделенных группах животных. Различия в использовании набора генов влияют на строение тела, а также на количество, идентичность и структуру частей тела. Большинство генов набора инструментов являются частью сигнальных путей : они кодируют факторы транскрипции , клеточной адгезии белки , белки рецепторов клеточной поверхности и сигнальные лиганды , которые связываются с ними, а также секретируемые морфогены , которые диффундируют через эмбрион. Все это помогает определить судьбу недифференцированных клеток эмбриона. Вместе они создают закономерности во времени и пространстве, которые формируют эмбрион и в конечном итоге формируют план тела организма. Среди наиболее важных генов инструментария являются Hox -гены . Эти факторы транскрипции содержат мотив ДНК, связывающий гомеобоксный белок, который также встречается в других генах, и создают базовый рисунок тела вдоль его оси спереди назад. [20] где повторяющиеся части, такие как многочисленные позвонки змей Hox-гены определяют , , будут расти в развивающемся эмбрионе или личинке. [8] Pax-6 , уже упомянутый, представляет собой классический набор генов. [48] растения участвуют и другие гены Хотя в формировании плана тела , [49] Гены гомеобокса также обнаружены в растениях, а это означает, что они являются общими для всех эукариот . [50] [51] [52]
эмбриона Регуляторные сети
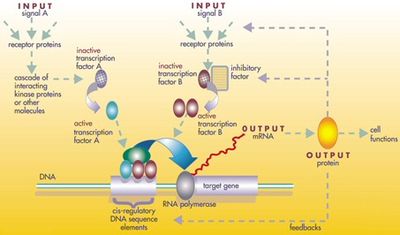
Белковые продукты регуляторного инструментария повторно используются не путем дублирования и модификации, а посредством сложной мозаики плейотропии , применяемой в неизмененном виде во многих независимых процессах развития, создавая структуру для многих несходных структур тела. [20] Локусы этих плейотропных генов имеют большие, сложные и модульные цис-регуляторные элементы . Например, в то время как ген неплейотропного родопсина у плодовой мухи имеет цис-регуляторный элемент длиной всего в несколько сотен пар оснований , плейотропная безглазая цис-регуляторная область содержит 6 цис-регуляторных элементов в более чем 7000 пар оснований. [20] зачастую Задействованные регулирующие сети очень велики. Каждый регуляторный белок контролирует «сотни» цис-регуляторных элементов. Например, 67 факторов транскрипции плодовых мушек контролировали в среднем 124 гена-мишени каждый. [20] Вся эта сложность позволяет генам, участвующим в развитии эмбриона, включаться и выключаться точно в нужное время и в нужных местах. Некоторые из этих генов являются структурными, непосредственно образующими ферменты, ткани и органы эмбриона. Но многие другие сами по себе являются регуляторными генами, поэтому зачастую включается точно рассчитанный по времени каскад переключений, включающий в себя запуск одного процесса развития за другим в развивающемся эмбрионе. [20]
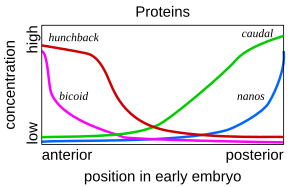
Такая каскадная регуляторная сеть была подробно изучена при развитии эмбриона плодовой мухи . Молодой эмбрион имеет овальную форму, похожую на мяч для регби . Небольшое количество генов продуцирует информационные РНК , которые создают градиенты концентрации вдоль длинной оси эмбриона. У раннего эмбриона гены bicoid и hunchback находятся в высокой концентрации вблизи переднего конца и определяют структуру будущей головы и грудной клетки; гены caudal и nanos находятся в высокой концентрации вблизи заднего конца и придают структуру самым задним сегментам брюшка. Эффекты этих генов взаимодействуют; например, белок Bicoid блокирует трансляцию информационной РНК каудального отдела , поэтому концентрация каудального белка на переднем конце становится низкой. Хвостовой позже включает гены, которые создают самые задние сегменты мухи, но только на заднем конце, где они наиболее сконцентрированы. [53] [54]

Белки Bicoid, Hunchback и Caudal, в свою очередь, регулируют транскрипцию генов пробелов, таких как гигантский , книрпс , Круппель и бесхвостый в полосатом узоре, создавая первый уровень структур, которые станут сегментами. [37] Белки из них, в свою очередь, контролируют гены правила пар , которые на следующем этапе создают 7 полос поперек длинной оси эмбриона. Наконец, гены полярности сегментов, такие как engrailed, разделяют каждую из 7 полос на две, создавая 14 будущих сегментов. [53] [54]
Этот процесс объясняет точную консервацию последовательностей генов набора инструментов, что привело к глубокой гомологии и функциональной эквивалентности белков набора инструментов у разных животных (что наблюдается, например, когда белок мыши контролирует развитие плодовых мух). Взаимодействия транскрипционных факторов и цис-регуляторных элементов или сигнальных белков и рецепторов блокируются в результате многократного использования, что делает почти любую мутацию вредной и, следовательно, устраняет естественный отбор. [20]
Механизм, который устанавливает ось перед-зад у каждого животного , один и тот же, что подразумевает общего предка. Аналогичный механизм существует для оси спины и брюшка у двусторонних он обратный животных, но у членистоногих и позвоночных . [55] Другой процесс, гаструляция эмбриона, управляется молекулярными моторами миозина II , которые не консервативны у разных видов. Этот процесс мог быть запущен движением морской воды в окружающей среде, а затем сменился эволюцией движений тканей эмбриона. [56] [57]
Истоки новизны [ править ]
Среди наиболее удивительных и, возможно, парадоксальных (с неодарвинистской точки зрения) результатов недавних исследований в области эволюционной биологии развития можно отметить то, что разнообразие тела строения и морфологии у организмов многих типов не обязательно отражается в разнообразии на уровне последовательности генов, в том числе из набора генетических инструментов развития и других генов, участвующих в развитии. Действительно, как заметили Джон Герхарт и Марк Киршнер, существует очевидный парадокс: «там, где мы больше всего ожидаем найти вариации, мы обнаруживаем консервацию, отсутствие изменений». [58] Итак, если наблюдаемая морфологическая новизна между разными кладами не возникает из-за изменений в последовательностях генов (например, в результате мутаций ), то откуда она берется? Новизна может возникнуть в результате мутационных изменений в регуляции генов . [20] [59] [60] [61]
Вариации в наборе инструментов [ править ]


Вариации в наборе инструментов, возможно, повлияли на значительную часть морфологической эволюции животных. Этот набор инструментов может стимулировать эволюцию двумя способами. Ген набора инструментов может экспрессироваться по-другому, например, когда клюв большого вьюрка Дарвина был увеличен за счет гена BMP . [62] или когда змеи лишились ног, так как дистально-менее выраженные стали недовыраженными или вовсе не выраженными в тех местах, где у других рептилий продолжали формироваться конечности. [63] Или ген из набора инструментов может приобрести новую функцию, как видно из многих функций того же гена, distal-less , который контролирует такие разнообразные структуры, как нижняя челюсть у позвоночных, [64] [65] ноги и усики у плодовой мухи, [66] и узор глазных пятен на бабочки крыльях . [67] Учитывая, что небольшие изменения в генах инструментария могут вызвать значительные изменения в структурах тела, они часто активируют одну и ту же функцию конвергентно или параллельно . distal-less генерирует узоры крыльев у бабочек Heliconius Erato и Heliconius melpomene , которые являются имитаторами Мюллера . В так называемой облегченной вариации [68] их модели крыльев возникли в результате разных эволюционных событий, но контролируются одними и теми же генами. [69] Изменения в развитии могут непосредственно способствовать видообразованию . [70]
Консолидация эпигенетических изменений
Эволюционные инновации могут иногда начинаться в стиле Ламарка с эпигенетических изменений регуляции генов или генерации фенотипов , которые впоследствии закрепляются изменениями на уровне генов . Эпигенетические изменения включают модификацию ДНК путем обратимого метилирования. [71] а также непрограммируемое переустройство организма под воздействием физических и других воздействий окружающей среды вследствие присущей пластичности . механизмам развития [72] Биологи Стюарт А. Ньюман и Герд Б. Мюллер предположили, что организмы на ранних этапах истории многоклеточной жизни были более восприимчивы к этой второй категории эпигенетической детерминации, чем современные организмы, что обеспечивает основу для ранних макроэволюционных изменений. [73]
развития Предвзятость

Развитие в определенных линиях может быть смещено либо в положительную сторону, в сторону заданной траектории или фенотипа, либо в сторону определенной траектории или фенотипа. [б] или отрицательно, от производства определенных типов изменений; либо может быть абсолютным (изменение происходит всегда или никогда), либо относительным. Однако доказательства любого такого направления эволюции трудно получить, и они также могут быть результатом ограничений развития, которые ограничивают диверсификацию. [75] Например, у брюхоногих моллюсков раковина всегда построена в виде трубки, растущей как в длину, так и в диаметре; В рамках этих ограничений отбор создал большое разнообразие форм оболочек, таких как плоские спирали, каури и высокие спирали турели. Среди многоножек Lithobiomorpha во взрослом возрасте всегда имеют 15 сегментов туловища, что, вероятно, является результатом отклонения в развитии в сторону нечетного числа сегментов туловища. В другом отряде многоножек, Geophilomorpha , количество сегментов варьируется у разных видов от 27 до 191, но число всегда нечетное, что делает это абсолютным ограничением; почти все нечетные номера в этом диапазоне заняты тем или иным видом. [74] [76] [77]
биология эволюционная развития Экологическая
Экологическая эволюционная биология развития [с] объединяет исследования в области биологии развития и экологии , чтобы изучить их связь с эволюционной теорией. [78] Исследователи изучают такие концепции и механизмы, как пластичность развития , эпигенетическое наследование , генетическая ассимиляция , построение ниш и симбиоз . [79] [80]
См. также [ править ]
- Проблема с головой членистоногого
- Передача сигналов ячейки
- Эволюция и развитие (журнал)
- Эволюционная биология развития человека
- Просто такие истории (с точки зрения биологов-эволюционистов)
- Эволюционная биология развития растений
- Теория перепросмотра
Примечания [ править ]
- ↑ Хотя Ч. Х. Уоддингтон в своей статье 1953 года «Эпигенетика и эволюция» призывал к добавлению эмбриологии к синтезу. [26]
- ^ Позитивную предвзятость иногда называют стремлением к развитию. [74]
- ^ В испанской Википедии есть статья на эту тему: eco-evo-devo .
Ссылки [ править ]
- ^ Леруа, Арман Мари (2014). Лагуна: как Аристотель изобрел науку . Блумсбери. стр. 181–182. ISBN 978-1-4088-3622-4 .
- ^ Ричардсон; Кеук (2002). «Азбука эволюции и развития Геккеля». Биологические обзоры Кембриджского философского общества . 77 (4): 495–528. CiteSeerX 10.1.1.578.2749 . дои : 10.1017/s1464793102005948 . ПМИД 12475051 . S2CID 23494485 . стр.516
- ^ О'Коннелл, Линдси (10 июля 2013 г.). «Концепция рекапитуляции Меккеля-Серра» . Энциклопедия проекта «Эмбрион» . Проверено 10 октября 2016 г.
- ^ Десмонд, Адриан Дж . (1989). Политика эволюции: морфология, медицина и реформы в радикальном Лондоне . Чикаго: Издательство Чикагского университета. стр. 53–53, 86–88, 337–340, 490–491 . ISBN 978-0-226-14374-3 .
- ^ Секорд 2003 , с. 252–253
- ^ Боулер, Питер Дж. (2003). Эволюция: история идеи . Беркли: Издательство Калифорнийского университета. стр. 120–128, 190–191, 208 . ISBN 978-0-520-23693-6 .
- ^ Секорд 2003 , с. 424, 512
- ↑ Перейти обратно: Перейти обратно: а б с д и ж Кэрролл, Шон Б. «Происхождение формы» . Естественная история . Проверено 9 октября 2016 г.
Биологи могли бы с уверенностью сказать, что формы меняются и что естественный отбор является важной силой, способствующей изменениям. Однако они ничего не могли сказать о том, как осуществляется это изменение. Как изменяются тела или части тела или как возникают новые структуры, оставалось полной загадкой.
- ↑ Перейти обратно: Перейти обратно: а б с д и ж Гилберт, Скотт Ф. (2003). «Морфогенез эволюционной биологии развития» (PDF) . Международный журнал биологии развития . 47 (7–8): 467–477. ПМИД 14756322 .
- ^ Дарвин, Чарльз (1859). О происхождении видов . Лондон: Джон Мюррей. стр. 439–440. ISBN 978-0-8014-1319-3 .
Усикопеды представляют собой хороший пример этого: даже прославленный Кювье не осознавал, что ракушка — это ракообразное, а это действительно так и есть; но взгляд на личинку безошибочно показывает, что это так.
- ^ Ричмонд, Марша (январь 2007 г.). «Исследование Дарвином Цирипедии» . Дарвин онлайн . Проверено 9 октября 2016 г.
- ↑ Перейти обратно: Перейти обратно: а б Холл, БК (2003). «Эво-Дево: эволюционные механизмы развития». Международный журнал биологии развития . 47 (7–8): 491–495. ПМИД 14756324 .
- ^ Ридли, Марк (2003). Эволюция . Уайли-Блэквелл. ISBN 978-1-4051-0345-9 .
- ^ Гулд 1977 , стр. 221–222.
- ^ Болл, Филип (7 февраля 2013 г.). «Оглядываясь назад: о росте и форме» . Природа . 494 (32–33): 32–33. Бибкод : 2013Natur.494...32B . дои : 10.1038/494032а . S2CID 205076253 .
- ^ Шализи, Косма. «Обзор: Гобелен Филипа Болла, сделанный своими руками» . Мичиганский университет . Проверено 14 октября 2016 г.
- ^ Тьюринг, Алан М. (14 августа 1952 г.). «Химические основы морфогенеза». Философские труды Лондонского королевского общества Б. 237 (641): 37–72. Бибкод : 1952РСПТБ.237...37Т . дои : 10.1098/rstb.1952.0012 . S2CID 120437796 .
- ^ Гриббин, Джон (2004). Глубокая простота . Случайный дом. п. 126.
- ^ Бок, Уолтер Дж. (июль 1981 г.). «Рецензируемая работа: Эволюционный синтез. Перспективы унификации биологии ». Аук . 98 (3): 644–646. JSTOR 4086148 .
- ↑ Перейти обратно: Перейти обратно: а б с д и ж г час я Кэрролл, Шон Б. (2008). «Эво-Дево и расширяющийся эволюционный синтез: генетическая теория морфологической эволюции» . Клетка . 134 (1): 25–36. дои : 10.1016/j.cell.2008.06.030 . ПМИД 18614008 . S2CID 2513041 .
- ^ Хелд, Льюис И. (2014). Как Змея потеряла ноги. Любопытные истории с границ Эво-Дево . Издательство Кембриджского университета . п. 67. ИСБН 978-1-107-62139-8 .
- ^ Гулд 1977 , стр. 221–222.
- ^ Бригандт, Инго (2006). «Гомология и гетерохрония: эволюционный эмбриолог Гэвин Райландс де Бир (1899-1972)» (PDF) . Журнал экспериментальной зоологии . 306Б (4): 317–328. Бибкод : 2006JEZB..306..317B . дои : 10.1002/jez.b.21100 . ПМИД 16506229 .
- ^ Гилберт, Сан-Франциско; Опиц, Дж. М.; Рафф, РА (1996). «Ресинтез эволюционной биологии и биологии развития» . Биология развития . 173 (2): 357–372. дои : 10.1006/dbio.1996.0032 . ПМИД 8605997 .
- ^ Адамс, М. (1991). «Зазеркалье: Эволюция советского дарвинизма». В Уоррене, Л.; Копровски, Х. (ред.). Новые перспективы эволюции . Лисс/Уайли. стр. 37–63.
- ^ Смоковит 1996 , с. 153
- ^ Моно, Жак ; Чангу, Япония; Джейкоб, Франсуа (1963). «Аллостерические белки и системы клеточного контроля». Журнал молекулярной биологии . 6 (4): 306–329. дои : 10.1016/S0022-2836(63)80091-1 . ПМИД 13936070 .
- ^ Жакоб, Франсуа (10 июня 1977 г.). «Эволюция и мастерство». Наука . 196 (4295): 1161–1166. Бибкод : 1977Sci...196.1161J . дои : 10.1126/science.860134 . ПМИД 860134 .
- ^ Гилберт, Сан-Франциско; Опиц, Дж. М.; Рафф, РА (1996). «Ресинтез эволюционной биологии и биологии развития» . Биология развития . 173 (2): 357–372. дои : 10.1006/dbio.1996.0032 . ПМИД 8605997 .
- ^ Мюллер, Великобритания (2007). «Эво-дево: расширение эволюционного синтеза». Обзоры природы Генетика . 8 (12): 943–949. дои : 10.1038/nrg2219 . ПМИД 17984972 . S2CID 19264907 .
- ^ Гудман, CS; Кофлин, Британская Колумбия (2000). Гудман, CS; Кафлин Б.С. (ред.). «Специальный репортаж: Эволюция биологии эво-дево» . Труды Национальной академии наук . 97 (9): 4424–4456. Бибкод : 2000PNAS...97.4424G . дои : 10.1073/pnas.97.9.4424 . ПМК 18255 . ПМИД 10781035 .
- ^ Мюллер ГБ и Ньюман С.А. (ред.) (2005). «Спецвыпуск: Эволюционные инновации и морфологические новизны» . Журнал экспериментальной зоологии, часть B. 304Б (6): 485–631. дои : 10.1002/jez.b.21080 . ПМИД 16252267 . Архивировано из оригинала 11 декабря 2012 года.
- ^ Палмер, Р.А. (2004). «Нарушение симметрии и эволюция развития». Наука . 306 (5697): 828–833. Бибкод : 2004Sci...306..828P . CiteSeerX 10.1.1.631.4256 . дои : 10.1126/science.1103707 . ПМИД 15514148 . S2CID 32054147 .
- ↑ Перейти обратно: Перейти обратно: а б Винчестер, Гил (2004). «Эдвард Б. Льюис 1918–2004» (PDF) . Современная биология . 14 (18) (опубликовано 21 сентября 2004 г.): R740–742. дои : 10.1016/j.cub.2004.09.007 . ПМИД 15380080 . S2CID 32648995 .
- ^ Бюрглин, Томас Р. «Страница Homeobox» . Каролинский институт . Проверено 13 октября 2016 г.
- ^ Голландия, PW (2013). «Эволюция генов гомеобокса». Wiley Interdiscip Rev Dev Biol . 2 (1): 31–45. дои : 10.1002/wdev.78 . ПМИД 23799629 . S2CID 44396110 .
Гены гомеобокса обнаружены почти у всех эукариот и в эволюции животных разделились на 11 классов генов и более 100 семейств генов, а также от 10 до 14 классов генов у растений.
- ↑ Перейти обратно: Перейти обратно: а б Нюсляйн-Фольхард, К.; Вишаус, Э. (октябрь 1980 г.). «Мутации, влияющие на количество и полярность сегментов у дрозофилы ». Природа . 287 (5785): 795–801. Бибкод : 1980Natur.287..795N . дои : 10.1038/287795a0 . ПМИД 6776413 . S2CID 4337658 .
- ^ Артур, Уоллес (14 февраля 2002 г.). «Новые концептуальные рамки эволюционной биологии развития». Природа . 415 (6873): 757–764. Бибкод : 2002Natur.415..757A . дои : 10.1038/415757a . ПМИД 11845200 . S2CID 4432164 .
- ^ «Эрик Вишаус и Кристиан Нюсляйн-Фольхард: сотрудничество в поиске генов развития» . iБиология. Архивировано из оригинала 13 октября 2016 года . Проверено 13 октября 2016 г.
- ^ Коэн, С.М.; Юргенс, Г. (1989). «Формирование проксимально-дистального паттерна у дрозофилы: автономная потребность клеток в бездистальной активности в развитии конечностей» . ЭМБО Дж . 8 (7): 2045–2055. дои : 10.1002/j.1460-2075.1989.tb03613.x . ПМК 401088 . ПМИД 16453891 .
- ^ Кэрролл, Шон Б. (2006) [2005]. Бесконечные самые красивые формы: новая наука Эво Дево и создание царства животных . Вайденфельд и Николсон [Нортон]. стр. 63–70. ISBN 978-0-297-85094-6 .
- ^ Панганибан, Г.; Ирвин, С.М.; Лоу, К.; Роэль, Х.; Корли, Л.С.; Шербон, Б.; Гренье, Дж. К.; Фэллон, Дж. Ф.; Кимбл, Дж.; Уокер, М.; Рэй, Джорджия; Свалла, Би Джей; Мартиндейл, MQ; Кэрролл, С.Б. (1997). «Происхождение и эволюция придатков животных» . Труды Национальной академии наук Соединенных Штатов Америки . 94 (10): 5162–5166. Бибкод : 1997PNAS...94.5162P . дои : 10.1073/pnas.94.10.5162 . ПМК 24649 . ПМИД 9144208 .
- ^ «Флуоресцентные зонды» . ТермоФишер Сайентифик. 2015 . Проверено 12 октября 2016 г.
- ^ Земля, МФ; Фернальд, Р.Д. (1992). «Эволюция глаз». Ежегодный обзор неврологии . 15 : 1–29. дои : 10.1146/annurev.ne.15.030192.000245 . ПМИД 1575438 .
- ↑ Перейти обратно: Перейти обратно: а б Томарев Станислав И.; Каллаертс, Патрик; Кос, Лидия; Зиновьева, Рина; Гальдер, Георг; Геринг, Вальтер; Пятигорский, Иорам (1997). «Кальмар Пакс-6 и развитие глаз» . Труды Национальной академии наук . 94 (6): 2421–2426. Бибкод : 1997PNAS...94.2421T . дои : 10.1073/pnas.94.6.2421 . ЧВК 20103 . ПМИД 9122210 .
- ↑ Перейти обратно: Перейти обратно: а б Пишо, Франк; Десплан, Клод (август 2002 г.). «Гены Pax и органогенез глаза». Текущее мнение в области генетики и развития . 12 (4): 430–434. дои : 10.1016/S0959-437X(02)00321-0 . ПМИД 12100888 .
- ^ Дрост, Хайк-Георг; Яница, Филипп; Гросс, Иво; Квинт, Марсель (2017). «Сравнение песочных часов развития между королевствами» . Текущее мнение в области генетики и развития . 45 : 69–75. дои : 10.1016/j.где.2017.03.003 . ПМИД 28347942 .
- ^ Сюй, ПХ; Ууу, Я.; Она, Х.; Бейер, Д.Р.; Маас, Р.Л. (1997). «Гомологам мышиной Eya гена отсутствия глаз дрозофилы требуется Pax6 для экспрессии в хрусталике и носовой плакоде». Разработка . 124 (1): 219–231. дои : 10.1242/dev.124.1.219 . ПМИД 9006082 .
- ^ Квинт, Марсель; Дрост, Хайк-Георг; Габель, Александр; Ульрих, Кристиан Карстен; Бонн, Маркус; Гроссе, Иво (4 октября 2012 г.). «Транскриптомные песочные часы в эмбриогенезе растений». Природа . 490 (7418): 98–101. Бибкод : 2012Natur.490...98Q . дои : 10.1038/nature11394 . ISSN 0028-0836 . ПМИД 22951968 . S2CID 4404460 .
- ^ Мукерджи, К.; Броккьери, Л.; Бурглин, Т.Р. (декабрь 2009 г.). «Комплексная классификация и эволюционный анализ генов гомеобокса растений» . Молекулярная биология и эволюция . 26 (12): 2775–94. дои : 10.1093/molbev/msp201 . ПМК 2775110 . ПМИД 19734295 .
- ^ Бурглин, Т.Р. (ноябрь 1997 г.). «Анализ генов гомеобокса суперкласса TALE (MEIS, PBC, KNOX, Iroquois, TGIF) выявил новый домен, консервативный у растений и животных» . Исследования нуклеиновых кислот . 25 (21): 4173–80. дои : 10.1093/нар/25.21.4173 . ПМК 147054 . ПМИД 9336443 .
- ^ Дерелл, Р.; Лопес, П.; Ле Гаядер, Х.; Мануэль, М. (2007). «Гомеодоменные белки принадлежат к древнему молекулярному набору эукариот». Эволюция и развитие . 9 (3): 212–9. дои : 10.1111/j.1525-142X.2007.00153.x . ПМИД 17501745 . S2CID 9530210 .
- ↑ Перейти обратно: Перейти обратно: а б Рассел, Питер (2010). iGenetics: молекулярный подход . Пирсон Образование. стр. 564–571. ISBN 978-0-321-56976-9 .
- ↑ Перейти обратно: Перейти обратно: а б Ривера-Помар, Роландо; Джекл, Герберт (1996). «От градиентов к полосам в эмбриогенезе дрозофилы: заполнение пробелов». Тенденции в генетике . 12 (11): 478–483. дои : 10.1016/0168-9525(96)10044-5 . ПМИД 8973159 .
- ^ Де Робертис, Эдди; Сасаи, Йошики (1996). «Общий план формирования дорсовентрального паттерна у Bilateria». Природа . 380 (6569): 37–40. Бибкод : 1996Natur.380...37D . дои : 10.1038/380037a0 . PMID 8598900 . S2CID 4355458 .
- ^ Фарж, Эммануэль (2003). «Механическая индукция скручивания передней кишки / зачатка стомодели дрозофилы» . Современная биология . 13 (16): 1365–1377. дои : 10.1016/s0960-9822(03)00576-1 . ПМИД 1293230 .
- ^ Нгуен, Нгок-Мин; Мерль, Татьяна; и др. (2022). «Механо-биохимическая морская стимуляция инверсии, гаструляции и спецификации эндомезодермы у многоклеточных эукариот» . Границы клеточной биологии и биологии развития . 10 : 992371. дои : 10.3389/fcell.2022.992371 . ПМЦ 9754125 . ПМИД 36531949 .
- ^ Герхарт, Джон; Киршнер, Марк (1997). Клетки, эмбрионы и эволюция . Блэквелл Наука. ISBN 978-0-86542-574-3 .
- ^ Кэрролл, Шон Б .; Гренье, Дженнифер К.; Уэзерби, Скотт Д. (2005). От ДНК к разнообразию: молекулярная генетика и эволюция дизайна животных — второе издание . Издательство Блэквелл. ISBN 978-1-4051-1950-4 .
- ^ Кэрролл, Шон Б. (2000). «Бесконечные формы: эволюция регуляции генов и морфологическое разнообразие» . Клетка . 101 (6): 577–80. дои : 10.1016/S0092-8674(00)80868-5 . ПМИД 10892643 . S2CID 12375030 .
- ^ Мочек, Армин П.; и др. (2015). «Значение и масштабы эволюционной биологии развития: взгляд на 21 век» (PDF) . Эволюция и развитие . 17 (3): 198–219. дои : 10.1111/ede.12125 . hdl : 2027.42/111768 . ПМИД 25963198 . S2CID 9652129 . Архивировано из оригинала (PDF) 17 апреля 2016 года . Проверено 27 ноября 2015 г.
- ^ Абжанов А.; Протас, М.; Грант, БР; Грант, PR; Табин, CJ (2004). «Bmp4 и морфологическая изменчивость клювов у зябликов Дарвина». Наука . 305 (5689): 1462–1465. Бибкод : 2004Sci...305.1462A . дои : 10.1126/science.1098095 . ПМИД 15353802 . S2CID 17226774 .
- ^ Кон, MJ; Тикл, К. (1999). «Основы развития без конечностей и осевого рисунка у змей». Природа . 399 (6735): 474–479. Бибкод : 1999Natur.399..474C . дои : 10.1038/20944 . ПМИД 10365960 . S2CID 4309833 .
- ^ Бевердам, А.; Мерло, Греция; Палеари, Л.; Мантеро, С.; Генуя, Ф.; Барбьери, О.; Жанвье, П.; Леви, Г. (август 2002 г.). «Трансформация челюстей с усилением симметрии после инактивации DLX5/DLX6: зеркало прошлого?» (PDF) . Бытие . 34 (4): 221–227. дои : 10.1002/gen.10156 . hdl : 2318/87307 . ПМИД 12434331 . S2CID 19592597 .
- ^ Депью, MJ; Луфкин, Т.; Рубинштейн, Дж. Л. (октябрь 2002 г.). «Спецификация подразделений челюсти генами DLX» . Наука . 298 (5592): 381–385. дои : 10.1126/science.1075703 . ПМИД 12193642 . S2CID 10274300 .
- ^ Панганибан, Грейс; Рубинштейн, Джон Л.Р. (2002). «Функции развития генов гомеобокса Distal-less/Dlx» . Разработка . 129 (19): 4371–4386. дои : 10.1242/dev.129.19.4371 . ПМИД 12223397 .
- ^ Бельдаде, П.; Брейкфилд, премьер-министр; Лонг, AD (2002). «Вклад дистальных элементов в количественные изменения глазных пятен бабочки». Природа . 415 (6869): 315–318. дои : 10.1038/415315a . ПМИД 11797007 . S2CID 4430563 .
- ^ Герхарт, Джон; Киршнер, Марк (2007). «Теория облегченной вариации» . Труды Национальной академии наук . 104 (приложение 1): 8582–8589. Бибкод : 2007PNAS..104.8582G . дои : 10.1073/pnas.0701035104 . ПМЦ 1876433 . ПМИД 17494755 .
- ^ Бакстер, Юго-Запад; Папа, Р.; Чемберлен, Н.; Хамфри, SJ; Джорон, М.; Моррисон, К.; французский-Констант, Р.Х.; Макмиллан, Висконсин; Джиггинс, компакт-диск (2008). «Конвергентная эволюция в генетической основе мюллеровой мимикрии у бабочек Heliconius» . Генетика . 180 (3): 1567–1577. doi : 10.1534/genetics.107.082982 . ПМК 2581958 . ПМИД 18791259 .
- ^ Пенниси, Э. (2002). «Эволюционная биология: энтузиасты Эво-Дево углубляются в детали». Наука . 298 (5595): 953–955. дои : 10.1126/science.298.5595.953 . ПМИД 12411686 . S2CID 154023266 .
- ^ Яблонка, Ева ; Лэмб, Мэрион (1995). Эпигенетическая наследственность и эволюция: ламаркианское измерение . Оксфорд, Нью-Йорк: Издательство Оксфордского университета. ISBN 978-0-19-854063-2 .
- ^ Вест-Эберхард, Мэри Джейн (2003). Пластичность развития и эволюция . Нью-Йорк: Издательство Оксфордского университета. ISBN 978-0-19-512235-0 .
- ^ Мюллер, Герд Б .; Ньюман, Стюарт А., ред. (2003). Происхождение организменной формы: за пределами гена в биологии развития и эволюции . МТИ Пресс.
- ↑ Перейти обратно: Перейти обратно: а б Артур, В. (июль 2001 г.). «Стимул развития: важный фактор, определяющий направление фенотипической эволюции». Эволюция и развитие . 3 (4): 271–278. дои : 10.1046/j.1525-142x.2001.003004271.x . ПМИД 11478524 . S2CID 41698287 .
- ^ Дрост, Хайк-Георг; Яница, Филипп; Гросс, Иво; Квинт, Марсель (2017). «Сравнение песочных часов развития между королевствами» . Текущее мнение в области генетики и развития . 45 : 69–75. дои : 10.1016/j.где.2017.03.003 . ПМИД 28347942 .
- ^ Артур, В. (октябрь 2002 г.). «Взаимодействие между отклонениями в развитии и естественным отбором: от сегментов многоножек к общей гипотезе» . Наследственность . 89 (4): 239–246. дои : 10.1038/sj.hdy.6800139 . ПМИД 12242638 .
- ^ Чипман, Ариэль Д.; Артур, Уоллес; Акам, Майкл (июль 2004 г.). «Периодичность двойных сегментов лежит в основе генерации сегментов в развитии многоножек» . Современная биология . 14 (14): 1250–1255. дои : 10.1016/j.cub.2004.07.026 . PMID 15268854 . S2CID 2371623 .
- ^ Абухейф, Э.; Фаве, М.-Ж.; Ибарраран-Винегра, AS; Лесовэй, член парламента; Рафики, AM; Раджакумар, Р. (2014). «Эко-Эво-Дево: Время пришло». В Ландри, Чехия; Обен-Хорт, Н. (ред.). Экологическая геномика: экология и эволюция генов и геномов . Достижения экспериментальной медицины и биологии. Том. 781. стр. 107–125. дои : 10.1007/978-94-007-7347-9_6 . ISBN 978-94-007-7346-2 . ПМИД 24277297 .
- ^ Шлихтинг, CD (2009). «Неравномерный путеводитель по Эко-Дево». Бионаука . 59 (11): 1000–1001. дои : 10.1525/bio.2009.59.11.12 . S2CID 116886911 .
- ^ Гилберт, Сан-Франциско ; Бош, ТКГ; Ледон-Реттиг, К. (2015). «Эко-Эво-Дево: симбиоз развития и пластичность развития как эволюционные агенты». Обзоры природы Генетика . 16 (10): 611–622. дои : 10.1038/nrg3982 . ПМИД 26370902 . S2CID 205486234 .
Источники [ править ]
- Гулд, Стивен Джей (1977). Онтогенез и филогения . Кембридж, Массачусетс: Belknap Press издательства Гарвардского университета . ISBN 978-0-674-63940-9 . LCCN 76045765 . ОСЛК 2508336 .
- Секорд, Джеймс А. (2003). Викторианская сенсация: необычайная публикация, прием и тайное авторство «Остатков естественной истории творения» . Чикаго: Издательство Чикагского университета. ISBN 978-0-226-74410-0 .
- Смоковит, Василики Бетти (1996). Объединяющая биология: эволюционный синтез и эволюционная биология . Принстон, Нью-Джерси: Издательство Принстонского университета. ISBN 978-0-691-03343-3 . OCLC 34411399 .
Внешние ссылки [ править ]
 СМИ, связанные с эволюционной биологией развития , на Викискладе?
СМИ, связанные с эволюционной биологией развития , на Викискладе?
| Базы данных органов управления : Национальные |
|---|