Переходное ископаемое
| Часть серии о |
| Палеонтология |
|---|
 |
| Палеонтологический портал Категория |
— Переходное ископаемое это любые окаменелые остатки формы жизни, которая демонстрирует черты, общие как для группы предков, так и для группы ее производных потомков. [1] Это особенно важно там, где группа потомков резко отличается по грубой анатомии и образу жизни от группы предков. Эти окаменелости служат напоминанием о том, что таксономические подразделения — это человеческие конструкции, которые задним числом были навязаны континууму вариаций. Из-за неполноты летописи окаменелостей обычно невозможно точно узнать, насколько близко переходное ископаемое находится к точке дивергенции. Следовательно, нельзя предполагать, что переходные окаменелости являются прямыми предками более поздних групп, хотя они часто используются в качестве моделей таких предков. [2]
В 1859 году, когда Чарльза Дарвина впервые была опубликована книга «Происхождение видов» , летопись окаменелостей была плохо известна. Дарвин описывал предполагаемое отсутствие переходных окаменелостей как «самое очевидное и серьезное возражение, которое можно выдвинуть против моей теории», но объяснял это, связывая это с крайним несовершенством геологических данных. [3] Он отметил ограниченность коллекций, доступных в то время, но описал имеющуюся информацию как демонстрацию закономерностей, вытекающих из его теории происхождения с модификациями посредством естественного отбора . [4] Действительно, археоптерикс был открыт всего два года спустя, в 1861 году, и представляет собой классическую переходную форму между более ранними нептичьими динозаврами и птицами . С тех пор было обнаружено гораздо больше переходных окаменелостей , и теперь имеется множество свидетельств того, как классы позвоночных , включая множество переходных окаменелостей. все связаны между собой [5] Конкретными примерами переходов на уровне классов являются: четвероногие и рыбы , птицы и динозавры , а также млекопитающие и «млекопитающие рептилии» .
Термин «недостающее звено» широко использовался в популярных трудах по эволюции человека для обозначения предполагаемого пробела в истории эволюции гоминид . Чаще всего он используется для обозначения любых новых находок переходных ископаемых. Ученые, однако, не используют этот термин, поскольку он относится к доэволюционному взгляду на природу.
и филогенетическая таксономия Эволюционная
в филогенетической номенклатуре Переходы

В эволюционной систематике, преобладающей форме таксономии на протяжении большей части 20-го века и до сих пор используемой в учебниках для неспециалистов, таксоны, основанные на морфологическом сходстве, часто изображаются как «пузыри» или «веретена», ответвляющиеся друг от друга, образующие эволюционные деревья. [6] Переходные формы рассматриваются как попадающие между различными группами с точки зрения анатомии, имеющие смесь характеристик изнутри и снаружи недавно разветвленной клады . [7]
С появлением кладистики в 1990-х годах отношения стали обычно выражаться в кладограммах , которые иллюстрируют ветвление эволюционных линий в виде палочкообразных фигур. Различные так называемые «естественные» или « монофилетические » группы образуют вложенные единицы, и только им даны филогенетические названия . Хотя в традиционной классификации четвероногие и рыбы рассматриваются как две разные группы, филогенетически четвероногие считаются ветвью рыб. Таким образом, в кладистике больше не существует перехода между устоявшимися группами, а термин «переходные окаменелости» является неправильным . Дифференциация происходит внутри групп, представленных ветвями кладограммы. [8]
В кладистическом контексте переходные организмы можно рассматривать как ранние примеры ветви, в которой не все черты, типичные для ранее известных потомков этой ветви, еще развились. [9] Таких ранних представителей группы обычно называют « базальными таксонами » или « сестринскими таксонами ». [10] в зависимости от того, принадлежит ли ископаемый организм к дочерней кладе или нет. [8]
Переходный против предкового [ править ]
Источником путаницы является представление о том, что переходная форма между двумя разными таксономическими группами должна быть прямым предком одной или обеих групп. Трудность усугубляется тем, что одной из целей эволюционной систематики является выявление таксонов, которые были предками других таксонов. Однако, поскольку эволюция представляет собой ветвящийся процесс, который создает сложную структуру кустов родственных видов, а не линейный процесс, приводящий к лестничному прогрессу, и из-за неполноты летописи окаменелостей, маловероятно, что какая-либо конкретная форма, представленная в окаменелостях, запись является прямым предком любой другой. Кладистика преуменьшает значение концепции, согласно которой одна таксономическая группа является предком другой, и вместо этого подчеркивает выявление сестринских таксонов, которые имеют более недавнего общего предка друг с другом, чем с другими группами. Есть несколько исключительных случаев, таких как некоторые морского планктона микроископаемые , когда летопись окаменелостей достаточно полна, чтобы с уверенностью предположить, что определенные окаменелости представляют собой популяцию, которая на самом деле была предком более поздней популяции другого вида. [11] Но в целом считается, что переходные окаменелости имеют особенности, которые иллюстрируют переходные анатомические особенности фактических общих предков разных таксонов, а не являются фактическими предками. [2]
Яркие примеры [ править ]
Археоптерикс [ править ]

Археоптерикс — род динозавров -теропод , тесно связанных с птицами. С конца 19-го века палеонтологи признавали его и отмечали в справочниках непрофессионалов как старейшую из известных птиц, хотя исследование 2011 года поставило под сомнение эту оценку, предположив, что это неавиальный динозавр . связанных с происхождением птиц. [12]
Он обитал на территории нынешней южной Германии в позднеюрский период около 150 миллионов лет назад, когда Европа была архипелагом в мелком теплом тропическом море, гораздо ближе к экватору, чем сейчас. По форме похожа на европейскую сороку , причем самые крупные особи, возможно, достигают размеров ворона . [13] Археоптерикс мог вырасти примерно до 0,5 метра (1,6 фута) в длину. Несмотря на небольшой размер, широкие крылья и предполагаемую способность летать или планировать, археоптерикс имеет больше общего с другими мелкими мезозойскими динозаврами, чем с современными птицами. В частности, он имеет следующие общие черты с дейнонихозаврами ( дромеозаврами и троодонтидами ): челюсти с острыми зубами, три пальца с когтями, длинный костлявый хвост, сверхразгибаемые вторые пальцы («убийственный коготь»), перья (что предполагает гомеотермию ) и различные особенности скелета. [14] Эти особенности делают археоптерикса явным кандидатом на роль переходного ископаемого между динозаврами и птицами. [15] что делает его важным для изучения как динозавров, так и происхождения птиц.
О первом полном экземпляре было объявлено в 1861 году, и археоптерикса с тех пор было найдено еще десять окаменелостей . Большинство из одиннадцати известных окаменелостей содержат отпечатки перьев — одни из старейших прямых свидетельств существования таких структур. Более того, поскольку эти перья принимают развитую форму маховых , окаменелости археоптерикса являются свидетельством того, что перья начали развиваться еще до поздней юры. [16]
Австралопитек [ править] афарский

Гоминид Australopithecus afarensis представляет собой эволюционный переход между современными двуногими людьми и их четвероногими предками- обезьянами . Ряд черт скелета A. afarensis сильно отражает прямохождение, до такой степени, что некоторые исследователи предположили, что двуногость развилась задолго до A. afarensis . [17] В общей анатомии таз гораздо больше похож на человеческий, чем на обезьяний. Лопатки подвздошных костей короткие и широкие, крестец широкий и расположен непосредственно позади тазобедренного сустава, имеется явное свидетельство сильного прикрепления разгибателей коленного сустава , что предполагает вертикальное положение. [17] : 122
Хотя таз не совсем похож на человеческий (он заметно широк или расширен, с латерально ориентированными подвздошными лопатками), эти особенности указывают на радикально реконструированную структуру, позволяющую приспособить значительную степень прямохождения . Бедренная кость наклоняется к колену от бедра . Эта черта позволяет стопе опускаться ближе к средней линии тела и явно указывает на привычное двуногое передвижение. Этой же особенностью обладают современные люди, орангутанги и обезьяны-пауки . На ступнях большие пальцы сведены , что затрудняет, а то и делает невозможным захват ветвей задними конечностями . Помимо передвижения, A. afarensis мозг был немного больше, чем у современного шимпанзе. [18] (ближайший из ныне живущих родственников человека) и имел зубы, которые больше напоминали человеческие, чем обезьяньи. [19]
, Ambulocetus Пакицетиды


Китообразные потомками (киты, дельфины и морские свиньи) являются морских млекопитающих наземных млекопитающих . Пакицетиды — семейство вымершее Indohyus копытных млекопитающих, являющихся древнейшими китами, ближайшей сестринской группой которых является из семейства Raoellidae . [20] [21] Они жили в раннем эоцене , около 53 миллионов лет назад. Их окаменелости были впервые обнаружены в Северном Пакистане в 1979 году, на реке недалеко от берегов бывшего моря Тетис . [22] [ нужна страница ] Пакицетиды могли слышать под водой, используя усиленную костную проводимость, а не барабанную перепонку, как большинство наземных млекопитающих. Такое расположение не обеспечивает направленного слуха под водой. [23]
Ambulocetus natans , живший около 49 миллионов лет назад, был обнаружен в Пакистане в 1994 году. Вероятно, он был земноводным и внешне напоминал крокодила . [24] В эоцене амбулоцетиды населяли заливы и эстуарии океана Тетис на севере Пакистана. [25] Окаменелости амбулоцетид всегда находят в прибрежных мелководных морских отложениях, связанных с обильными окаменелостями морских растений и прибрежными моллюсками . [25] Хотя они встречаются только в морских отложениях, значения их изотопов кислорода указывают на то, что они потребляли воду с разной степенью солености: у некоторых экземпляров не было обнаружено никаких признаков потребления морской воды, а у других - никаких следов потребления пресной воды в то время, когда их зубы окаменели. . Очевидно, что амбулоцетиды переносили широкий диапазон концентраций соли. [26] В их рацион, вероятно, входили наземные животные, приближавшиеся к воде для питья, или пресноводные водные организмы, обитавшие в реке. [25] Следовательно, амбулоцетиды представляют собой переходную фазу предков китообразных между пресноводной и морской средой обитания.
Тиктаалик [ править ]


Тиктаалик — род вымерших саркоптеригиев (кипастеперых рыб) позднего девона , многие черты которого сходны с четвероногими (четвероногими животными). [27] Это одна из нескольких линий древних саркоптеригов, развивших адаптацию к бедной кислородом мелководной среде обитания своего времени — адаптации, которые привели к эволюции четвероногих. [28] Хорошо сохранившиеся окаменелости были найдены в 2004 году на острове Элсмир в Нунавуте , Канада. [29]
Тиктаалик жил примерно 375 миллионов лет назад. Палеонтологи предполагают, что это типичный переход между позвоночными, не являющимися четвероногими, такими как Panderichthys , известными по окаменелостям возрастом около 380 миллионов лет, и ранними четвероногими, такими как Acanthostega и Ichthyostega , известными по окаменелостям возрастом около 365 миллионов лет. Его смесь примитивных рыб и производных характеристик четвероногих побудила одного из его первооткрывателей, Нила Шубина , охарактеризовать Тиктаалика как « рыбоногих ». [30] [31] В отличие от многих предыдущих, более похожих на рыб переходных окаменелостей, «плавники» Тиктаалика имеют основные кости запястья и простые лучи, напоминающие пальцы. Возможно, они были несущими . Как и у всех современных четвероногих, у него были ребра, подвижная шея с отдельным грудным поясом и легкие, хотя у него были жабры, чешуя и плавники рыбы. [27] Однако в статье Буасверта и соавт. 2008 г. отмечается, что Panderichthys , из-за его более развитой дистальной части, мог быть ближе к четвероногим, чем Tiktaalik , который, возможно, независимо развил сходство с четвероногими в результате конвергентной эволюции. [32]
Следы четвероногих, найденные в Польше и опубликованные в журнале Nature в январе 2010 года, были «надежно датированы» тем, что они на 10 миллионов лет старше самых старых известных элписстостегид. [33] которого является Тиктаалик ( примером ), подразумевая, что такие животные, как Тиктаалик , обладающие особенностями, которые развились около 400 миллионов лет назад, были «позднесохранившимися реликтами, а не прямыми переходными формами, и они подчеркивают, насколько мало мы знаем о самой ранней истории наземные позвоночные». [34]
Амфистиум [ править ]


Pleuronectiformes (камбалы) — отряд лучепёрых рыб . Наиболее очевидной характеристикой современной камбалы является их асимметрия: у взрослой рыбы оба глаза находятся на одной стороне головы. У некоторых семейств глаза всегда находятся на правой стороне тела (правоглазая или правоглазая камбала), а у других — всегда на левой (левосторонняя или левоглазая камбала). Примитивные колючие тюрбо включают равное количество право- и левоглазых особей и, как правило, менее асимметричны, чем другие семейства. Другими отличительными особенностями отряда являются наличие выступающих глаз, другое приспособление к жизни на морском дне ( бентосе ) и расширение спинного плавника на голову. [35]
Амфистиум — это ископаемая рыба возрастом 50 миллионов лет, идентифицированная как ранний родственник камбалы и переходная ископаемая. [36] У Amphistium переход от типичной симметричной головы позвоночного неполный, один глаз расположен ближе к центру верхней части головы. [37] Палеонтологи пришли к выводу, что «изменения происходили постепенно, в соответствии с эволюцией посредством естественного отбора , а не внезапно, как когда-то у исследователей не было другого выбора, кроме как поверить». [36]
Амфистиум входит в число многих видов ископаемых рыб, известных из Монте-Болька лагерштетте в лютетинской Италии. Heteronectes — родственное и очень похожее ископаемое из несколько более ранних слоев Франции. [37]
Рункария [ править ]

среднедевонский предшественник семенных растений В Бельгии был обнаружен , который примерно на 20 миллионов лет предшествовал самым ранним семенным растениям. Рункария , небольшая и радиально-симметричная, представляет собой интегументированный мегаспорангий, окруженный купулой . Мегаспорангий имеет нераскрытый дистальный отросток, выступающий над многолопастным покровом . Предполагается, что это расширение участвовало в анемофильном опылении . Рункария проливает новый свет на последовательность приобретения признаков, ведущую к образованию семян, обладающих всеми качествами семенных растений, за исключением твердой семенной кожуры и системы направления пыльцы к семени. [38]
Ископаемая запись [ править ]
Не каждая переходная форма присутствует в летописи окаменелостей , поскольку летопись окаменелостей не является полной. Организмы лишь в редких случаях сохраняются в виде окаменелостей и при самых благоприятных обстоятельствах, и лишь небольшая часть таких окаменелостей была обнаружена. Палеонтолог Дональд Протеро отметил, что это иллюстрируется тем фактом, что количество видов, известных по летописи окаменелостей, составляло менее 5% от числа известных живых видов, что позволяет предположить, что количество видов, известных по окаменелостям, должно быть намного меньше 1%. всех когда-либо существовавших видов. [39]
Из-за особых и редких обстоятельств, необходимых для окаменения биологической структуры, логика подсказывает, что известные окаменелости представляют собой лишь небольшой процент всех когда-либо существовавших форм жизни — и что каждое открытие представляет собой лишь снимок эволюции. Сам переход может быть проиллюстрирован и подтвержден только переходными окаменелостями, которые никогда не демонстрируют точной середины между явно расходящимися формами. [40]
Летопись окаменелостей очень неравномерна и, за немногими исключениями, сильно смещена в сторону организмов с твердыми частями, в результате чего большинство групп мягкотелых организмов практически не имеют летописи окаменелостей. [39] Группами, имеющими хорошую летопись окаменелостей, включая ряд переходных окаменелостей между традиционными группами, являются позвоночные, иглокожие , брахиоподы и некоторые группы членистоногих . [41]
История [ править ]
Постдарвиновский [ править ]


Идея о том, что виды животных и растений не постоянны, а изменяются с течением времени, была высказана еще в XVIII веке. [42] Книга Дарвина «Происхождение видов» , опубликованная в 1859 году, дала ей прочную научную основу. Однако слабостью работы Дарвина было отсутствие палеонтологических свидетельств, на что указывал сам Дарвин. Хотя легко представить, что естественный отбор порождает вариации, наблюдаемые внутри родов и семейств, трансформацию между высшими категориями представить было труднее. Впечатляющая находка лондонского экземпляра археоптерикса в 1861 году, всего через два года после публикации работы Дарвина, впервые предложила связь между классом высокоразвитых птиц и классом более базальных рептилий . [43] В письме Дарвину палеонтолог Хью Фальконер писал:
Если бы каменоломням Зольнхофена было поручено — по августейшему приказу — добыть странное существо в духе Дарвина, оно не смогло бы выполнить этот приказ лучше, чем в случае с « Археоптериксом» . [44]
Таким образом, переходные окаменелости, такие как археоптерикс, стали рассматриваться не только как подтверждение теории Дарвина, но и как самостоятельные символы эволюции. [45] Например, шведский энциклопедический словарь Nordisk familjebok 1904 года показал неточную реконструкцию окаменелости археоптерикса (см. иллюстрацию) «ett af de betydelsefullaste paleontologiska fynd, som någonsin gjorts» («одно из самых значительных палеонтологических открытий, когда-либо сделанных»). [46]
Возникновение растений [ править ]
Переходные окаменелости принадлежат не только животным. С увеличением картографирования отделов растений в начале 20 века начались поиски предка сосудистых растений . В 1917 году Роберт Кидстон и Уильям Генри Лэнг нашли останки крайне примитивного растения в кремне Райни в Абердиншире , Шотландия, и назвали его Риния . [47]
Растение ринии было небольшим и похожим на палочку, с простыми дихотомически ветвящимися стеблями без листьев, каждый из которых имел спорангий . Простая форма перекликается со спорофитом мхов , у и было показано, что риний наблюдалась смена поколений с соответствующим гаметофитом в виде скученных пучков крошечных стеблей высотой всего несколько миллиметров. [48] Таким образом, риния занимает промежуточное положение между мхами и ранними сосудистыми растениями, такими как папоротники и плауны . Из ковра мохоподобных гаметофитов более крупные спорофиты Rhynia росли во многом подобно простым плаунам, распространяясь посредством горизонтально растущих стеблей, образующих ризоиды , которые прикрепляли растение к субстрату. Необычное сочетание моховых и сосудистых черт, а также чрезвычайная структурная простота растения имели огромное значение для ботанического понимания. [49]
Недостающие ссылки [ править ]


Идея о том, что все живые существа связаны посредством какого-то процесса трансмутации, возникла еще до теории эволюции Дарвина. Жан-Батист Ламарк предполагал, что жизнь постоянно зарождается в форме простейших существ, и стремился к сложности и совершенству (т.е. человека) через прогрессивный ряд низших форм. [50] По его мнению, низшие животные были просто новичками на эволюционной сцене. [51]
После «Происхождения видов » идея «низших животных», представляющих более ранние стадии эволюции, сохранялась, как показано в Эрнста Геккеля . рисунке человеческой родословной [52] Хотя тогда считалось, что позвоночные образуют своего рода эволюционную последовательность, различные классы были различны, а неоткрытые промежуточные формы назывались «недостающими звеньями».
Этот термин впервые был использован в научном контексте Чарльзом Лайелем в третьем издании (1851 г.) его книги «Элементы геологии» в отношении недостающих частей геологической колонки , но он стал популяризирован в своем нынешнем значении, когда появился на странице xi книги. его книга «Геологические свидетельства древности человека» 1863 года. К тому времени вообще считалось, что конец последнего ледникового периода ознаменовал первое появление человечества; Лайель опирался на новые открытия в своей книге «Древность человека», чтобы заглянуть в прошлое происхождения людей. Лайель писал, что остается глубокой загадкой, как можно преодолеть огромную пропасть между человеком и животным. [53] Яркие произведения Лайеля разожгли общественное воображение, вдохновив Жюля Верна на книгу «Путешествие к центру Земли» (1864 г.) и Луи Фигье « второе издание книги La Terre avant le déluge » («Земля до потопа») 1867 г., которое включало драматические иллюстрации. дикие мужчины и женщины в звериных шкурах и с каменными топорами на месте Эдемского сада , показанного в издании 1863 года. [54]
Однако поиск окаменелости, демонстрирующей переходные черты между обезьянами и людьми, был бесплодным, пока молодой голландский геолог Эжен Дюбуа не нашел череп, коренной зуб и бедренную кость на берегу реки Соло , Ява , в 1891 году. обезьяноподобна крыша черепа , объем мозга оценивается примерно в 1000 куб.см, что находится на полпути между мозгом шимпанзе и взрослого человека. Единственный коренной зуб был больше, чем любой современный человеческий зуб, но бедренная кость была длинной и прямой, а угол колена указывал на то, что « Яванский человек » ходил прямо. [55] Получив название Pithecanthropus erectus («прямоходящий человек-обезьяна»), он стал первым в длинном списке окаменелостей эволюции человека . В то время многие называли его «недостающим звеном», что помогло установить, что этот термин в первую очередь используется для человеческих окаменелостей, хотя иногда он используется и для других промежуточных продуктов, таких как промежуточный звено динозавр-птица -археоптерикс . [56] [57]
Хотя «недостающее звено» по-прежнему является популярным термином, хорошо узнаваемым среди общественности и часто используемым в популярных средствах массовой информации, [58] этот термин избегают в научных публикациях. [5] Некоторые блоггеры назвали это «неуместным»; [59] и потому, что звенья больше не «отсутствуют», и потому, что человеческая эволюция больше не считается происходившей в рамках единой линейной прогрессии. [5] [60]
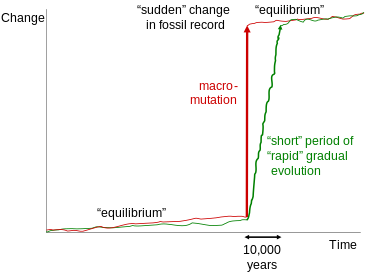
Прерывистое равновесие [ править ]
Теория прерывистого равновесия, разработанная Стивеном Джеем Гулдом и Найлсом Элдриджем и впервые представленная в 1972 году. [61] часто ошибочно вовлекается в обсуждение переходных окаменелостей. [62] Эта теория, однако, относится только к хорошо документированным переходам внутри таксонов или между близкородственными таксонами за геологически короткий период времени. Эти переходы, обычно прослеживаемые в одном и том же геологическом обнажении, часто демонстрируют небольшие скачки в морфологии между длительными периодами морфологической стабильности. Чтобы объяснить эти скачки, Гулд и Элдридж предположили сравнительно длительные периоды генетической стабильности, разделенные периодами быстрой эволюции. Гулд сделал следующее наблюдение относительно неправильного использования его работы креационистами для отрицания существования переходных окаменелостей:
Поскольку мы предложили прерывистое равновесие для объяснения тенденций, вызывает бешенство то, что креационисты снова и снова цитируют (я не знаю, намеренно или по глупости) признание того, что летопись окаменелостей не включает в себя переходных форм. Пунктуация встречается на уровне вида; Направленные тенденции (по лестничной модели) распространены на более высоком уровне переходов внутри основных групп.
См. также [ править ]
Ссылки [ править ]
- ^ Фриман и Херрон 2004 , с. 816
- ^ Jump up to: Перейти обратно: а б Протеро 2007 , стр. 133–135
- ^ Дарвин 1859 , стр. 279–280.
- ^ Дарвин 1859 , стр. 341–343.
- ^ Jump up to: Перейти обратно: а б с Протеро, Дональд Р. (1 марта 2008 г.). «Эволюция: какое недостающее звено?» . Новый учёный . 197 (2645): 35–41. дои : 10.1016/s0262-4079(08)60548-5 . ISSN 0262-4079 .
- ^ Например, см. Бентон 1997.
- ^ Протеро 2007 , с. 84 .
- ^ Jump up to: Перейти обратно: а б Казлев, М. Алан. «Земноводные, систематика и кладистика» . Палеос . Проверено 9 мая 2012 г.
- ^ Протеро 2007 , с. 127 .
- ^ Протеро 2007 , с. 263 .
- ^ Протеро, Дональд Р.; Лазарус, Дэвид Б. (июнь 1980 г.). «Планктонные микроископаемые и признание предков». Систематическая биология . 29 (2): 119–129. дои : 10.1093/sysbio/29.2.119 . ISSN 1063-5157 .
- ^ Син Сюй ; Хайлу Ю; Кай Ду; Фэнлу Хан (28 июля 2011 г.). « Археоптериксоподобный теропод из Китая и происхождение авиала». Природа . 475 (7357): 465–470. дои : 10.1038/nature10288 . ISSN 0028-0836 . ПМИД 21796204 . S2CID 205225790 .
- ^ Эриксон, Грегори М .; Раухут, Оливер В.М.; Чжунхэ Чжоу ; и др. (9 октября 2009 г.). «Была ли физиология динозавров унаследована птицами? Примиряем медленный рост археоптерикса » . ПЛОС Один . 4 (10): е7390. Бибкод : 2009PLoSO...4.7390E . дои : 10.1371/journal.pone.0007390 . ISSN 1545-7885 . ПМК 2756958 . ПМИД 19816582 .
- ^ Ялден, Дерек В. (сентябрь 1984 г.). «Какого размера был Археоптерикс ?». Зоологический журнал Линнеевского общества . 82 (1–2): 177–188. дои : 10.1111/j.1096-3642.1984.tb00541.x . ISSN 0024-4082 .
- ^ « Археоптерикс : ранняя пташка» . Музей палеонтологии Калифорнийского университета . Беркли, Калифорния: Калифорнийский университет, Беркли . Проверено 18 октября 2006 г.
- ^ Wellnhofer 2004 , стр. 282–300.
- ^ Jump up to: Перейти обратно: а б Лавджой, К. Оуэн (ноябрь 1988 г.). «Эволюция ходьбы человека» (PDF) . Научный американец . 259 (5): 82–89. Бибкод : 1988SciAm.259e.118L . дои : 10.1038/scientificamerican1188-118 . ISSN 0036-8733 . ПМИД 3212438 .
- ^ « Австралопитек афарский » . Эволюция человека . Вашингтон, округ Колумбия: Смитсоновского института Программа происхождения человека . Проверено 15 мая 2015 г.
- ^ Уайт, Тим Д .; Вор, генерал; Симпсон, Скотт; Асфау, Берхане (январь 2000 г.). «Челюсти и зубы Australopithecus afarensis из Мака, Средний Аваш, Эфиопия» Американский журнал физической антропологии . 111 (1): 45–68. doi : 10.1002/(SICI)1096-8644(200001)111:1<45::AID-AJPA4>3.0.CO;2-I . ISSN 0002-9483 . ПМИД 10618588 .
- ^ Медицинские и фармацевтические колледжи северо-восточного университета Огайо (21 декабря 2007 г.). «Киты произошли от крошечных предков, похожих на оленей» . Наука Дейли . Роквилл, доктор медицины: ScienceDaily, LLC . Проверено 15 мая 2015 г.
- ^ Джинджерич и Рассел, 1981 г.
- ^ Кастро и Хубер, 2003 г.
- ^ Нуммела, Сирпа; Тьювиссен, JGM; Баджпай, Сунил; и др. (12 августа 2004 г.). «Эоценовая эволюция слуха китов». Природа . 430 (7001): 776–778. Бибкод : 2004Natur.430..776N . дои : 10.1038/nature02720 . ISSN 0028-0836 . ПМИД 15306808 . S2CID 4372872 .
- ^ Тьювиссен, JGM; Уильямс, Эллен М.; Роу, Лоис Дж.; и др. (20 сентября 2001 г.). «Скелеты наземных китообразных и связь китов с парнокопытными». Природа . 413 (6853): 277–281. Бибкод : 2001Natur.413..277T . дои : 10.1038/35095005 . ISSN 0028-0836 . ПМИД 11565023 . S2CID 4416684 .
- ^ Jump up to: Перейти обратно: а б с Тьювиссен, JGM; Уильямс, Эллен М. (ноябрь 2002 г.). «Ранние проявления китообразных (млекопитающих): закономерности эволюции и корреляции развития». Ежегодный обзор экологии и систематики . 33 : 73–90. doi : 10.1146/annurev.ecolsys.33.020602.095426 . ISSN 1545-2069 .
- ^ Тьювиссен, JGM; Баджпай, Сунил (декабрь 2001 г.). «Происхождение китов как образец макроэволюции» . Бионаука . 51 (12): 1037–1049. doi : 10.1641/0006-3568(2001)051[1037:WOAAPC]2.0.CO;2 . ISSN 0006-3568 .
- ^ Jump up to: Перейти обратно: а б Дэшлер, Эдвард Б .; Шубин, Нил Х .; Дженкинс, Фариш А. младший (6 апреля 2006 г.). «Девонская четвероногие рыбы и эволюция строения тела четвероногих» . Природа . 440 (7085): 757–763. Бибкод : 2006Natur.440..757D . дои : 10.1038/nature04639 . ISSN 0028-0836 . ПМИД 16598249 .
- ^ Клак, Дженнифер А. (декабрь 2005 г.). «Как подняться на землю». Научный американец . 293 (6): 100–107. Бибкод : 2005SciAm.293f.100C . дои : 10.1038/scientificamerican1205-100 . ISSN 0036-8733 . ПМИД 16323697 .
- ^ Истон, Джон (23 октября 2008 г.). « Внутренняя анатомия Тиктаалика объясняет эволюционный переход от воды к суше» . Хроники Чикагского университета . 28 (3). ISSN 1095-1237 . Проверено 19 апреля 2012 г.
- ^ Уилфорд, Джон Ноубл (5 апреля 2006 г.). «Ученые называют рыбные ископаемые «недостающим звеном» » . Нью-Йорк Таймс . Проверено 17 мая 2015 г.
- ^ Shubin 2008
- ^ «Информация о грудных плавниках» . uu.diva-portal.org . Проверено 9 мая 2021 г.
- ^ Недзведский, Гжегож; Шрек, Питер; Наркевич, Катажина; и др. (7 января 2010 г.). «Следы четвероногих из раннего среднего девона Польши». Природа . 463 (7227): 43–48. Бибкод : 2010Natur.463...43N . дои : 10.1038/nature08623 . ISSN 0028-0836 . ПМИД 20054388 . S2CID 4428903 .
- ^ «Четыре фута в прошлом: следы предшествуют самым ранним окаменелостям тела» . Природа (Резюме редакции). 463 (7227). 7 января 2010 г. ISSN 0028-0836 .
- ^ Шапло и Амаока 1998 , стр. 223–226.
- ^ Jump up to: Перейти обратно: а б Минар, Энн (9 июля 2008 г.). «Находка странной рыбы противоречит аргументу разумного замысла» . Национальные географические новости . Вашингтон, округ Колумбия: Национальное географическое общество . Архивировано из оригинала 4 августа 2008 года . Проверено 17 июля 2008 г.
- ^ Jump up to: Перейти обратно: а б Фридман, Мэтт (10 июля 2008 г.). «Эволюционное происхождение асимметрии камбалы». Природа . 454 (7201): 209–212. Бибкод : 2008Natur.454..209F . дои : 10.1038/nature07108 . ISSN 0028-0836 . ПМИД 18615083 . S2CID 4311712 .
- ^ Жерриен, Филипп; Мейер-Берто, Бриджит; Фэйрон-Демаре, Мюриэль; и др. (29 октября 2004 г.). «Рункария, предшественник семенного растения среднего девона». Наука . 306 (5697): 856–858. Бибкод : 2004Sci...306..856G . дои : 10.1126/science.1102491 . ISSN 0036-8075 . ПМИД 15514154 . S2CID 34269432 .
- ^ Jump up to: Перейти обратно: а б Протеро 2007 , стр. 50–53
- ^ Исаак, Марк, изд. (5 ноября 2006 г.). «Заявление CC200: Переходные окаменелости» . Архив TalkOrigins . Хьюстон, Техас: Фонд TalkOrigins, Inc. Проверено 30 апреля 2009 г.
- ^ Донован и Пол 1998
- ^ Арчибальд, Дж. Дэвид (август 2009 г.). «Додарвиновское (1840 г.) «Древо жизни» Эдварда Хичкока » (PDF) . Журнал истории биологии . 42 (3): 561–592. CiteSeerX 10.1.1.688.7842 . дои : 10.1007/s10739-008-9163-y . ISSN 0022-5010 . ПМИД 20027787 . S2CID 16634677 .
- ^ Дарвин 1859 , Глава 10 .
- ^ Уильямс, Дэвид Б. (сентябрь 2011 г.). «Оценки: 30 сентября 1861 года: обнаружен и описан археоптерикс» . ЗЕМЛЯ . ISSN 1943-345Х . Архивировано из оригинала 7 мая 2012 года . Проверено 24 февраля 2012 г.
- ^ Веллнхофер 2009
- ^ Молоко 1904 , стр. 1379–1380 гг.
- ^ Кидстон, Роберт ; Ланг, Уильям Генри (27 февраля 1917 г.). «XXIV.— О старых растениях из красного песчаника, демонстрирующих структуру, из пласта Райни Черт, Абердиншир. Часть I. Риния Гвинн-Вогани, Кидстон и Лэнг» . Труды Королевского общества Эдинбурга . 51 (3): 761–784. дои : 10.1017/S0263593300006805 . ISSN 0080-4568 . OCLC 704166643 . S2CID 251580286 . Проверено 18 мая 2015 г.
- ^ Керп, Ганс; Тревин, Найджел Х.; Хасс, Хаген (2003). «Новые гаметофиты из раннедевонских рини-кремней». Труды Эдинбургского королевского общества: Науки о Земле . 94 (4): 411–428. дои : 10.1017/S026359330000078X . ISSN 0080-4568 . S2CID 128629425 .
- ^ Эндрюс 1967 , с. 32
- ^ Ламарк 1815–1822 гг.
- ^ Аппель, Тоби А. (осень 1980 г.). «Анри де Бленвиль и сериал о животных: цепочка бытия девятнадцатого века». Журнал истории биологии . 13 (2): 291–319. дои : 10.1007/BF00125745 . ISSN 0022-5010 . JSTOR 4330767 . S2CID 83708471 .
- ^ Геккель 2011 , с. 216.
- ^ Байнум, Уильям Ф. (лето 1984 г.). Чарльза Лайеля « Древность человека и ее критики». Журнал истории биологии . 17 (2): 153–187. дои : 10.1007/BF00143731 . ISSN 0022-5010 . JSTOR 4330890 . S2CID 84588890 .
- ^ Браун 2003 , стр. 130, 218, 515.
- ^ Свишер, Кертис и Левин, 2001 г.
- ^ Читатель 2011 г.
- ^ Бентон, Майкл Дж. (март 2001 г.). «Свидетельства эволюционных переходов» . действиебиологическая наука . Вашингтон, округ Колумбия: Американский институт биологических наук . Архивировано из оригинала 26 апреля 2012 года . Проверено 29 марта 2012 г.
- ^ Циммер, Карл (19 мая 2009 г.). «Дарвиниус: Он доставляет пиццу, удлиняется, укрепляется и находит ту тапочку, которая лежала на свободе под шезлонгом [так в оригинале] несколько недель…» The Loom (Блог). Вокеша, Висконсин: Kalmbach Publishing . Проверено 10 сентября 2011 г.
- ^ Самбрани, Наградж (10 июня 2009 г.). «Почему термин «недостающие звенья» неуместен» . Biology Times (блог) . Проверено 19 мая 2015 г.
- ^ «Недавно найденные окаменелости могут быть связаны с предком человека» . Новости ЦБК . Оттава, Онтарио, Канада: Канадская радиовещательная корпорация . 8 апреля 2010 года . Проверено 19 мая 2015 г.
Заманчиво назвать новый вид «недостающим звеном» между более ранними видами и современными людьми, но ученые говорят, что эта концепция больше не применима, учитывая новые знания об эволюции человека. [...] Исследователи теперь говорят, что эволюция человека состояла из множества разнообразных видов во многих ветвях, а не одной гладкой линии от обезьяноподобных видов к человеку.
- ^ Элдридж и Гулд 1972 , стр. 82–115.
- ^ Бейтс, Гэри (декабрь 2006 г.). «Эта цитата! — о пропавших переходных окаменелостях» . Создание . 29 (1): 12–15. ISSN 0819-1530 . Проверено 6 июля 2014 г.
- Теуниссен, Лайонел (24 июня 1997 г.). «Паттерсон неверно процитировал: история о двух «цитатах» » . Архив TalkOrigins . Хьюстон, Техас: Фонд TalkOrigins, Inc. Проверено 19 мая 2015 г.
- ^ Гулд 1980 , с. 189.
Источники [ править ]
- Эндрюс, Генри Н. младший (1967) [первоначально опубликовано в 1961 году]. Исследования по палеоботанике . Глава Чарльза Дж. Феликса о палинологии (переиздание). Нью-Йорк: Джон Уайли и сыновья . LCCN 61006768 . OCLC 12877482 .
- Бентон, Майкл Дж. (1997). Палеонтология позвоночных (2-е изд.). Лондон: Чепмен и Холл . ISBN 978-0-412-73810-4 . OCLC 37378512 .
- Браун, Джанет (2003) [первоначально опубликовано в 2002 г.]. Чарльз Дарвин: Сила места . Том. 2. Лондон: Джонатан Кейп . ISBN 978-0-7126-6837-8 . OCLC 806284755 .
- Кастро, Питер; Хубер, Майкл Э. (2003). Морская биология . Оригинальное произведение искусства Уильяма Обера и Клэр Гаррисон (4-е изд.). Нью-Йорк: МакГроу-Хилл . ISBN 978-0-07-029421-9 . LCCN 2002190248 . OCLC 49259996 .
- Шапло, Франсуа; Амаока, Кунио (1998). «Кабалки». В Пакстоне, Джон Р.; Эшмейер, Уильям М. (ред.). Энциклопедия рыб . Иллюстрации Дэвида Киршнера (2-е изд.). Сан-Диего, Калифорния: Academic Press . ISBN 978-0-12-547665-2 . LCCN 98088228 . ОСЛК 39641701 .
- Дарвин, Чарльз (1859). О происхождении видов путем естественного отбора, или о сохранении избранных рас в борьбе за жизнь (1-е изд.). Лондон: Джон Мюррей . LCCN 06017473 . OCLC 741260650 . Книга доступна в Полном собрании сочинений Чарльза Дарвина в Интернете . Проверено 13 мая 2015 г.
- Донован, Стивен К .; Пол, Кристофер Р.К., ред. (1998). Адекватность летописи окаменелостей . Чичестер; Нью-Йорк: Джон Уайли и сыновья. ISBN 978-0-471-96988-4 . LCCN 98010110 . OCLC 38281286 .
- Элдридж, Найлс ; Гулд, Стивен Джей (1972). «Периодическое равновесие: альтернатива филетическому градуализму». В Шопфе, Томас Дж. М. (ред.). Модели в палеобиологии . Сан-Франциско, Калифорния: Фриман, Купер. ISBN 978-0-87735-325-6 . LCCN 72078387 . OCLC 572084 .
- Фриман, Скотт; Херрон, Джон К. (2004). Эволюционный анализ (3-е изд.). Река Аппер-Сэддл, Нью-Джерси: Pearson Education . ISBN 978-0-13-101859-4 . LCCN 2003054833 . OCLC 52386174 .
- Джинджерич, Филип Д .; Рассел, Дональд Э. (1981). Pakicetus inachus, новый археоцет (Mammalia, Cetacea) из ранне-среднеэоценовой формации Кулдана в Кохате (Пакистан) (PDF) (отчет об исследовании). Материалы Музея палеонтологии. Том. 25. Анн-Арбор, Мичиган: Музей палеонтологии Мичиганского университета . стр. 235–246. ISSN 0097-3556 . LCCN 82621252 . OCLC 8263404 .
- Гулд, Стивен Джей (1980). Большой палец панды: больше размышлений о естествознании (1-е изд.). Нью-Йорк: WW Norton & Company . ISBN 978-0-393-01380-1 . LCCN 80015952 . OCLC 6331415 .
- Геккель, Эрнст (2011) [Первоначально опубликовано в 1912 году; Лондон: Уоттс и Ко. ]. Эволюция человека . Том. 1. Перевод с немецкого Джозефа Маккейба (5-е расширенное изд.). Гамбург, Германия: Классика Tredition. ISBN 978-3-8424-6302-8 . OCLC 830523724 .
- Ламарк, Жан-Батист (1815–1822). Естественная история животных без позвонков (на французском языке). Париж: Вердьер. LCCN 07018340 . OCLC 5269931 .
- Лавджой, Артур О. (1936). Великая цепь бытия: исследование истории идеи . Лекции Уильяма Джеймса, 1933. Кембридж, Массачусетс: Издательство Гарвардского университета . LCCN 36014264 . OCLC 192226 .
- Лече, В. (1904). «Археоптерикс». В Мейере, Бернхард (ред.). Книга о скандинавской семье (на шведском языке) (новое, переработанное и богато иллюстрированное издание). Стокгольм: Компания с ограниченной ответственностью Nordisk familjeboks förlags. LCCN 15023737 . ОСЛК 23562281 .
- Протеро, Дональд Р. (2007). Эволюция: о чем говорят окаменелости и почему это важно . Оригинальные иллюстрации Карла Бьюэлла. Нью-Йорк: Издательство Колумбийского университета . ISBN 978-0-231-13962-5 . LCCN 2007028804 . OCLC 154711166 .
- Читатель, Джон (2011). Недостающие звенья: в поисках происхождения человека . Предисловие Эндрю Хилла (Расширенное и обновленное издание). Оксфорд; Нью-Йорк: Издательство Оксфордского университета . ISBN 978-0-19-927685-1 . LCCN 2011934689 . OCLC 707267298 .
- Шубин, Нил (2008). Ваша внутренняя рыба: путешествие в 3,5-миллиардную историю человеческого тела . Нью-Йорк: Книги Пантеона . ISBN 978-0-375-42447-2 . LCCN 2007024699 . OCLC 144598195 .
- Свишер, Карл К. III; Кертис, Гарнисс Х .; Левин, Роджер (2001) [первоначально опубликовано в 2000 году]. Человек с Java: как два геолога изменили наше понимание эволюции человека . Чикаго, Иллинойс: Издательство Чикагского университета . ISBN 978-0-226-78734-3 . LCCN 2001037337 . OCLC 48066180 .
- Велнхофер, Питер (2004). «Оперение археоптерикса : перья динозавра?». В Карри, Филип Дж .; Коппельхус, Ева Б.; Шугар, Мартин А.; и др. (ред.). Пернатые драконы: исследования перехода от динозавров к птицам . Жизнь прошлого. Блумингтон, Индиана: Издательство Университета Индианы . ISBN 978-0-253-34373-4 . LCCN 2003019035 . OCLC 52942941 .
- Велнхофер, Питер (2009). Археоптерикс: икона эволюции . Перевод Фрэнка Хаазе; предисловие Луиса М. Кьяппе (переработанное английское издание 1-го немецкого изд.). Мюнхен: Верлаг доктора Фридриха Пфайля. ISBN 978-3-89937-108-6 . OCLC 501736379 .
Внешние ссылки [ править ]
- Ллойд, Робин (11 февраля 2009 г.). «Окаменелости раскрывают правду о теории Дарвина» . ЖиваяНаука . Огден ЮТ: Покупка . Проверено 19 мая 2015 г.
- Хант, Кэтлин (17 марта 1997 г.). «Часто задаваемые вопросы об окаменелостях переходных позвоночных» . Архив TalkOrigins . Хьюстон, Техас: Фонд TalkOrigins, Inc. Проверено 19 мая 2015 г.
- « Тиктаалик розовый » . Чикаго, Иллинойс: Чикагский университет . Проверено 19 мая 2015 г.
- «Киты» . Веллингтон, Новая Зеландия: Музей Новой Зеландии Те Папа Тонгарева . Проверено 19 мая 2015 г.
- Хатчинсон, Джон Р. (22 января 1998 г.). «Птицы действительно динозавры?» . ДиноБазз . Беркли, Калифорния: Музей палеонтологии Калифорнийского университета . Проверено 19 мая 2015 г.