Молекулярная палеонтология
| Часть серии о |
| Палеонтология |
|---|
 |
| Палеонтологический портал Категория |
Молекулярная палеонтология относится к восстановлению и анализу ДНК , белков , углеводов или липидов и их диагенетических продуктов из останков древнего человека, животных и растений. [1] [2] Область молекулярной палеонтологии дала важную информацию об эволюционных событиях, диаспорах видов , открытии и характеристике вымерших видов .
В последнее время достижения в области молекулярной палеонтологии позволили ученым заняться эволюционными вопросами на генетическом уровне, а не полагаться только на фенотипические вариации. Применяя методы молекулярного анализа к ДНК из недавних останков животных, можно количественно оценить уровень родства между любыми двумя организмами, ДНК которых была обнаружена. [3] Использование различных биотехнологических методов, таких как выделение ДНК , амплификация и секвенирование. [4] ученые смогли получить и расширить понимание дивергенции и истории эволюции бесчисленного количества недавно вымерших организмов. В феврале 2021 года ученые впервые сообщили о останков животных секвенировании ДНК , в данном случае мамонта , возрастом более миллиона лет, самой старой ДНК, секвенированной на сегодняшний день. [5] [6]
В давние времена неоднородность состава углеродистых останков разнообразных животных , возраст которых варьируется от неопротерозоя до недавнего времени , была связана с биологическими признаками, закодированными в современных биомолекулах посредством каскада реакций окислительного окаменения. [7] [8] [9] [10] Макромолекулярный состав углеродистых окаменелостей, примерно тонского возраста, [11] сохранять биологические сигнатуры, отражающие исходную биоминерализацию , тканей типы , метаболизм и родство ( филогенез ). [9]
История
[ редактировать ]Говорят, что изучение молекулярной палеонтологии началось с открытия Абельсоном аминокислот возрастом 360 миллионов лет, сохранившихся в ископаемых раковинах. [12] Однако Сванте Пяабо часто считают основателем области молекулярной палеонтологии. [13]
В области молекулярной палеонтологии с 1950-х годов произошло несколько крупных достижений, и эта область постоянно развивается. Ниже приводится график, показывающий заметный вклад, который был сделан.
Хронология
[ редактировать ]
середина 1950-х годов: Абельсон обнаружил сохранившиеся аминокислоты в ископаемых раковинах возрастом около 360 миллионов лет. Возникла идея сравнения последовательностей ископаемых аминокислот с существующими организмами, чтобы можно было изучить молекулярную эволюцию. [12]
1970-е: Ископаемые пептиды изучаются с помощью аминокислотного анализа . [14] Начните использовать цельные пептиды и иммунологические методы . [15]
Конец 1970-х годов: палеоботаники (можно также писать как палеоботаники) изучали молекулы хорошо сохранившихся ископаемых растений. [16]
1984: Первое успешное секвенирование ДНК вымершего вида квагги , похожего на зебру вида. [1]
1991: Опубликована статья об успешном извлечении белков из ископаемой кости динозавра, в частности сейсмозавра . [17]
2005: Ученые воскрешают вымерший вирус гриппа 1918 года . [18]
2006: Сегменты последовательности ядерной ДНК неандертальцев начинают анализироваться и публиковаться. [23]
2007: Ученые синтезируют с нуля весь вымерший эндогенный ретровирус человека (HERV-K). [19]
2010: Новый вид ранних гоминид, денисовцы , обнаружен на основе митохондриального и ядерного генома, извлеченного из кости, найденной в пещере в Сибири. Анализ показал, что денисовский экземпляр жил примерно 41 000 лет назад и имел общего предка как с современными людьми, так и с неандертальцами, жившими примерно 1 миллион лет назад в Африке. [20]
2013: Первый полный геном неандертальца успешно секвенирован. Более подробную информацию можно найти в проекте «Геном неандертальца» . [21]
2013: Образец возрастом 400 000 лет с остаточной митохондриальной ДНК секвенирован и оказался общим предком неандертальцев и денисовцев, Homo heidelbergensis . [22]
2013: Мэри Швейцер и ее коллеги предлагают первый химический механизм, объясняющий потенциальное сохранение клеток и мягких тканей позвоночных в летописи окаменелостей. Механизм предполагает, что свободные радикалы кислорода, потенциально образующиеся под действием окислительно-восстановительного железа, вызывают сшивку биомолекул. Этот механизм сшивания в некоторой степени аналогичен сшиванию, которое происходит во время гистологической фиксации ткани, например, формальдегидом. Авторы также предполагают, что источником железа является гемоглобин умершего организма. [24]
2015: ископаемых зубах возрастом 110 000 лет, содержащих ДНК денисовцев . Сообщается об [25] [26]
2018: Молекулярные палеобиологи связывают полимеры N-, O-, S-гетероциклического состава (AGEs/ALEs, как указано в цитируемой публикации, Wiemann et al. 2018) в углеродистых ископаемых остатках механически со структурными биомолекулами в исходных тканях. Посредством окислительного сшивания, процесса, подобного реакции Майяра , нуклеофильные аминокислотные остатки конденсируются с активными карбонильными группами, полученными из липидов и сахаров . [8] Процессы окаменения биомолекул, идентифицированные с помощью рамановской спектроскопии современных и ископаемых тканей, экспериментального моделирования и оценки статистических данных, включают улучшенное гликозилирование и улучшенное липоксидирование . [8]
2019: Независимая лаборатория молекулярных палеонтологов подтверждает трансформацию биомолекул посредством продвинутого гликозилирования и липоксидирования во время окаменения. [10] Авторы используют синхротронную инфракрасную спектроскопию с преобразованием Фурье .
2020: Виманн и его коллеги идентифицируют биологические признаки, отражающие первоначальную биоминерализацию , тканей типы , метаболизм и родство ( филогению ) в сохранившихся неоднородностях состава разнообразных углеродистых окаменелостей животных . [9] Это первый крупномасштабный анализ окаменелостей в возрасте от неопротерозоя до современности , а также первая опубликованная запись биологических сигналов, обнаруженных в сложном органическом веществе. [9] Авторы полагаются на статистический анализ уникально большого набора данных рамановской спектроскопии .
2021: Геохимики обнаруживают сигналы типа ткани в составе углеродистых окаменелостей, относящихся к тонианскому периоду . [11] и применить эти сигналы для идентификации эпибионтов . Авторы используют рамановскую спектроскопию .
2022: Данные рамановской спектроскопии, раскрывающие закономерности окаменения структурных биомолекул, были воспроизведены с помощью инфракрасной спектроскопии с преобразованием Фурье и множества различных рамановских инструментов, фильтров и источников возбуждения. [27]
2023: Опубликовано первое подробное химическое описание процесса окаменения оригинальных биологических клеток и тканей. Важно отметить, что исследование показывает, что гипотеза свободных радикалов кислорода (предложенная Мэри Швейцер и ее коллегами в 2013 году) во многих случаях идентична гипотезе образования AGE/ALE (предложенной Ясминой Виманн и ее коллегами в 2018 году). Комбинированные гипотезы, наряду с термическим созреванием и карбонизацией , образуют рыхлую основу для окаменения биологических клеток и тканей. [7]
Квагга
[ редактировать ]Первое успешное секвенирование ДНК вымершего вида было проведено в 1984 году на музейном экземпляре квагги, похожего на зебру, которому 150 лет. [1] Митохондриальная ДНК (также известная как мтДНК) была секвенирована из высушенных мышц квагги, и было обнаружено, что она отличается 12 заменами оснований от митохондриальной ДНК горной зебры. Был сделан вывод, что эти два вида имели общего предка 3-4 миллиона лет назад, что согласуется с известными ископаемыми свидетельствами существования этого вида. [28]
Денисовцы
[ редактировать ]Денисовцы что Евразии образец , вид гоминидов, родственный неандертальцам и людям, был обнаружен в результате секвенирования ДНК образца возрастом 41 000 лет, обнаруженного в 2008 году. Анализ митохондриальной ДНК из извлеченной кости пальца показал, генетически отличны как от людей, так и от неандертальцев. Позже выяснилось, что два зуба и кость пальца ноги принадлежали разным людям из одной и той же популяции. Анализ показывает, что и неандертальцы, и денисовцы уже присутствовали по всей Евразии, когда прибыли современные люди. [21] В ноябре 2015 года ученые сообщили об обнаружении ископаемого зуба, содержащего ДНК денисовцев, и оценили его возраст в 110 000 лет. [25] [26]
Анализ митохондриальной ДНК
[ редактировать ]
МтДНК денисовской кости пальца отличается от таковой современного человека на 385 оснований ( нуклеотидов ) в цепи мтДНК примерно из 16 500, тогда как разница между современными людьми и неандертальцами составляет около 202 оснований. Напротив, разница между шимпанзе и современными людьми составляет примерно 1462 пары оснований мтДНК. [20] Это предполагает, что время расхождения произошло около миллиона лет назад. МтДНК зуба имела большое сходство с мтДНК кости пальца, что указывает на их принадлежность к одной и той же популяции. [29] Из второго зуба была обнаружена последовательность мтДНК, которая показала неожиданно большое количество генетических различий по сравнению с последовательностью, обнаруженной в другом зубе и пальце, что указывает на высокую степень разнообразия мтДНК. Эти два человека из одной и той же пещеры продемонстрировали большее разнообразие, чем наблюдаемое среди выбранных неандертальцев со всей Евразии, и были такими же разными, как современные люди с разных континентов. [30]
Анализ ядерного генома
[ редактировать ]Выделение и секвенирование ядерной ДНК также было выполнено из кости пальца Денисовой. Этот образец показал необычную степень сохранности ДНК и низкий уровень загрязнения. Им удалось добиться почти полного секвенирования генома, что позволило провести детальное сравнение с неандертальцами и современными людьми. На основании этого анализа они пришли к выводу, что, несмотря на кажущееся расхождение митохондриальной последовательности, популяция Денисова вместе с неандертальцами имела общую ветвь линии, ведущей к современным африканским людям. По оценкам, среднее время расхождения между последовательностями денисовцев и неандертальцев составляет 640 000 лет назад, а время между ними обоими и последовательностями современных африканцев составляет 804 000 лет назад. Они предполагают, что расхождение мтДНК Денисовой является результатом либо сохранения линии, очищенной от других ветвей человечества в результате генетического дрейфа , либо интрогрессии из более старой линии гомининов. [29]
Мужчина из Гейдельберга
[ редактировать ]
Homo heidelbergensis был впервые обнаружен в 1907 году недалеко от Гейдельберга, Германия, а затем также обнаружен в других местах Европы, Африки и Азии. [31] [32] Однако только в 2013 году образец с поддающейся восстановлению ДНК был найден в бедренной кости возрастом около 400 000 лет, найденной в пещере Сима-де-лос-Уэсос в Испании. Было обнаружено, что бедренная кость содержит как мтДНК, так и ядерную ДНК. Усовершенствования в методах экстракции ДНК и подготовки библиотек позволили успешно выделить и секвенировать мтДНК, однако ядерная ДНК в наблюдаемом образце оказалась слишком деградированной, а также была загрязнена ДНК древнего пещерного медведя ( Ursus deningeri ), присутствующего в пещера. [33] Анализ мтДНК обнаружил удивительную связь между этим экземпляром и денисовцами, и это открытие вызвало много вопросов. Несколько сценариев были предложены в статье в январе 2014 года под названием «Последовательность митохондриального генома гоминина из Сима-де-лос-Уэсос», объясняющей отсутствие единого мнения в научном сообществе о том, как Homo heidelbergensis связан с другими известными группами гомининов. Один из вероятных сценариев, предложенных авторами, заключался в том, что H. heidelbergensis был предком как денисовцев, так и неандертальцев. [33] Полностью секвенированные ядерные геномы денисовцев и неандертальцев позволяют предположить, что у них был общий предок, живший примерно 700 000 лет назад, и один из ведущих исследователей в этой области, Сванте Паабо, предполагает, что, возможно, эта новая группа гомининов и является этим ранним предком. [22]
Приложения
[ редактировать ]Открытие и характеристика новых видов
[ редактировать ]Методы молекулярной палеонтологии, примененные к окаменелостям, способствовали открытию и описанию нескольких новых видов, включая денисовцев и Homo heidelbergensis . Мы смогли лучше понять путь, который прошли люди, заселив Землю, и какие виды присутствовали во время этой диаспоры .
Возрождение
[ редактировать ]
Теперь возможно возродить вымершие виды, используя методы молекулярной палеонтологии. Впервые это было достигнуто посредством клонирования в 2003 году пиренейского козла , вида диких коз, вымершего в 2000 году. Ядра клеток пиренейского козла были инъецированы в яйца коз, лишенные собственной ДНК, и имплантированы суррогатным матерям коз. [34] Потомство прожило всего семь минут после рождения из-за дефектов легких. У других клонированных животных наблюдались аналогичные дефекты легких. [35]
Многие виды вымерли в результате деятельности человека. Некоторые примеры включают дронта , большую гагарку , тасманского тигра , китайского речного дельфина и странствующего голубя . Вымерший вид можно возродить с помощью аллельной замены. [36] близкородственного вида, который все еще жив. Заменив всего лишь несколько генов в организме, вместо того, чтобы создавать геном вымершего вида с нуля, можно было бы возродить таким образом несколько видов, даже неандертальцев. [ нужна ссылка ]
Этика, связанная с реинтродукцией вымерших видов, очень противоречива. Критики возвращения к жизни вымерших видов утверждают, что это отвлечет ограниченные деньги и ресурсы от защиты текущих мировых проблем биоразнообразия . [37] Учитывая, что нынешние темпы вымирания примерно в 100–1000 раз превышают фоновые темпы вымирания, [38] есть опасения, что программа воскрешения может уменьшить обеспокоенность общественности по поводу нынешнего кризиса массового вымирания, если будет считаться, что эти виды можно просто вернуть к жизни. Как утверждают редакторы статьи Scientific American о возрождении вида: Должны ли мы вернуть шерстистого мамонта только для того, чтобы тем временем позволить слонам вымереть? [37] Основным движущим фактором вымирания большинства видов в эту эпоху (после 10 000 г. до н.э.) является потеря среды обитания, и временное возвращение вымерших видов не воссоздает среду, в которой они когда-то обитали. [39]
Сторонники воскрешения, такие как Джордж Чёрч , говорят о многих потенциальных выгодах. Возвращение вымерших ключевых видов, таких как шерстистый мамонт , может помочь восстановить баланс экосистем, которые когда-то зависели от них. Некоторые вымершие виды в случае возвращения могут принести огромную пользу окружающей среде, в которой они когда-то обитали. Например, шерстистые мамонты могут замедлить таяние российской и арктической тундры несколькими способами, например, поедая мертвую траву, чтобы новая трава могла расти и укореняться, а также периодически разбивать снег, подвергая землю внизу воздействию арктических ветров. воздух. Эти методы также можно использовать для восстановления генетического разнообразия видов, находящихся под угрозой исчезновения, или даже для введения новых генов и признаков, которые позволят животным лучше конкурировать в меняющейся среде. [40]
Исследования и технологии
[ редактировать ]Когда обнаруживается новый потенциальный образец, ученые обычно сначала анализируют сохранность клеток и тканей с помощью гистологических методов и проверяют условия выживания ДНК. Затем они попытаются выделить образец ДНК, используя метод, описанный ниже, и проведут ПЦР-амплификацию ДНК, чтобы увеличить количество ДНК, доступной для тестирования. Затем эту амплифицированную ДНК секвенируют. Особое внимание уделяется проверке соответствия последовательности филогенетическим признакам организма. [1] Когда организм умирает, метод, называемый датированием аминокислот для его старения можно использовать . Он проверяет степень рацемизации аспарагиновой кислоты , лейцина и аланина в тканях. С течением времени соотношение D/L (где «D» и «L» являются зеркальными отражениями друг друга) увеличивается с 0 до 1. [41] В образцах, в которых соотношение D/L аспарагиновой кислоты превышает 0,08, древние последовательности ДНК невозможно восстановить (по состоянию на 1996 год). [42]
Митохондриальная ДНК против ядерной ДНК
[ редактировать ]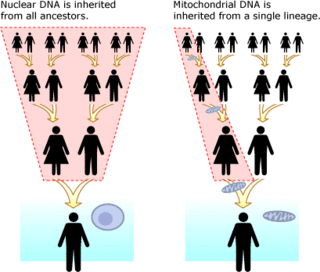
Митохондриальная ДНК (мтДНК) отделена от ядерной ДНК. Он присутствует в органеллах, называемых митохондриями, в каждой клетке . В отличие от ядерной ДНК , которая наследуется от обоих родителей и перестраивается каждое поколение, точная копия митохондриальной ДНК передается от матери к ее сыновьям и дочерям. Преимущества проведения анализа ДНК с использованием митохондриальной ДНК заключаются в том, что она имеет гораздо меньшую частоту мутаций, чем ядерная ДНК, что значительно упрощает отслеживание родословных в масштабе десятков тысяч лет. Зная базовую частоту мутаций мтДНК, [43] (у людей эта скорость также известна как митохондриальные молекулярные часы человека ) можно определить количество времени, в течение которого любые две линии были разделены. Еще одним преимуществом мтДНК является то, что в каждой клетке существуют тысячи ее копий, тогда как в каждой клетке существуют только две копии ядерной ДНК. [44] Все эукариоты (группа, в которую входят все растения, животные и грибы) имеют мтДНК. [45] Недостатком мтДНК является то, что представлена только материнская линия. Например, ребенок унаследует 1/8 своей ДНК от каждого из своих восьми прадедов, однако он унаследует точный клон мтДНК своей прабабушки по материнской линии. Это аналогично тому, как ребенок унаследовал только фамилию своего прадеда по отцовской линии, а не смесь всех восьми фамилий.
Изоляция
[ редактировать ]необходимо учитывать множество факторов При выделении вещества . Во-первых, в зависимости от того, что это такое и где оно расположено, существуют протоколы, которые необходимо соблюдать, чтобы избежать загрязнения и дальнейшей деградации образца. [4] Затем обработка материалов обычно осуществляется в физически изолированной рабочей зоне и при определенных условиях (т.е. определенной температуре, влажности и т. д.), чтобы избежать загрязнения и дальнейшей потери образца. [4]
После получения материала, в зависимости от того, что это такое, существуют разные способы его выделения и очистки. Экстракция ДНК из окаменелостей — одна из наиболее популярных практик, и для получения желаемого образца можно предпринять различные шаги. [4] , погребенных в янтаре, ДНК, извлеченную из окаменелостей можно взять из небольших образцов и смешать с различными веществами, центрифугировать , инкубировать и снова центрифугировать. [46] С другой стороны, экстракцию ДНК из насекомых можно провести путем измельчения образца, смешивания его с буфером и очистки на колонках из стекловолокна. [47] В конце концов, независимо от того, как были выделены образцы этих окаменелостей, выделенная ДНК должна быть способна подвергнуться амплификации . [4] [46] [47]
Усиление
[ редактировать ]
Область молекулярной палеонтологии получила большую выгоду от изобретения полимеразной цепной реакции (ПЦР) , которая позволяет делать миллиарды копий фрагмента ДНК всего из одной сохранившейся копии ДНК. Одной из самых больших проблем до этого момента была крайняя нехватка восстановленной ДНК из-за ее деградации с течением времени. [1]
Секвенирование
[ редактировать ]Секвенирование ДНК проводится для определения порядка нуклеотидов и генов. [48] Существует множество различных материалов, из которых можно извлечь ДНК. У животных митохондриальную хромосому можно использовать для молекулярных исследований. Хлоропласты можно изучать у растений как основной источник данных о последовательностях. [48]

В конечном итоге сгенерированные последовательности используются для построения эволюционных деревьев . [48] Методы сопоставления наборов данных включают в себя: максимальную вероятность , минимальную эволюцию (также известную как соединение соседей ), которая ищет дерево с наименьшей общей длиной, и метод максимальной экономии, который находит дерево, требующее наименьшего количества изменений состояния символов. [48] Группы видов, определенные в дереве, также можно позже оценить с помощью статистических тестов, таких как метод начальной загрузки , чтобы увидеть, действительно ли они значимы. [48]
Ограничения и проблемы
[ редактировать ]Трудно найти идеальные условия окружающей среды для сохранения ДНК там, где организм был высушен и обнаружен, а также для поддержания их состояния до анализа. Ядерная ДНК обычно быстро разлагается после смерти в результате эндогенных гидролитических процессов . [42] УФ-излучением, [1] и другие экологические стрессоры.
Кроме того, было обнаружено, что взаимодействие с продуктами органического распада окружающей почвы помогает сохранить биомолекулярные материалы. [49] Однако они также создали дополнительную проблему, связанную с возможностью разделения различных компонентов для проведения надлежащего анализа. [50] Было также обнаружено, что некоторые из этих нарушений мешают действию некоторых ферментов, используемых во время ПЦР. [49]
Наконец, одной из самых больших проблем при извлечении древней ДНК, особенно древней человеческой ДНК, является загрязнение во время ПЦР. Небольшие количества человеческой ДНК могут загрязнять реагенты, используемые для экстракции и ПЦР древней ДНК. Эти проблемы можно решить, если тщательно обращаться со всеми растворами, а также со стеклянной посудой и другими инструментами, используемыми в процессе. Также может помочь, если экстракцию будет выполнять только один человек, чтобы свести к минимуму присутствие различных типов ДНК. [42]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а б с д и ж г Марота I, Ролло Ф (январь 2002 г.). «Молекулярная палеонтология» . Клеточные и молекулярные науки о жизни . 59 (1): 97–111. дои : 10.1007/s00018-002-8408-8 . ПМИД 11846037 . S2CID 6377604 .
- ^ Швейцер М.Х. (апрель 2004 г.). «Молекулярная палеонтология: некоторые современные достижения и проблемы» . Анналы палеонтологии . 90 (2): 81–102. Бибкод : 2004AnPal..90...81H . дои : 10.1016/j.annpal.2004.02.001 . Проверено 22 апреля 2014 г.
- ^ Шапиро Б., Хофрайтер М. (январь 2014 г.). «Палеогеномный взгляд на эволюцию и функцию генов: новые идеи древней ДНК». Наука . 343 (6169): 1236573. doi : 10.1126/science.1236573 . ПМИД 24458647 . S2CID 1015148 .
- ^ Jump up to: а б с д и Вагонер Б. (2001). «Молекулярная палеонтология» (PDF) . Энциклопедия наук о жизни . Издательская группа «Природа»: 1–5.
- ^ Хант К. (17 февраля 2021 г.). «Самая старая в мире ДНК секвенирована у мамонта, жившего более миллиона лет назад» . Новости CNN . Проверено 17 февраля 2021 г.
- ^ Каллауэй Э (февраль 2021 г.). «Геномы мамонтов возрастом в миллион лет побили рекорд древнейшей древней ДНК » Природа . 590 (7847): 537–538. Бибкод : 2021Natur.590..537C . дои : 10.1038/d41586-021-00436-x . ПМИД 33597786 .
- ^ Jump up to: а б Андерсон Л. (май 2023 г.). «Химическая основа сохранения ископаемых клеток и мягких тканей позвоночных» . Обзоры наук о Земле . 240 : 104367. Бибкод : 2023ESRv..24004367A . doi : 10.1016/j.earscirev.2023.104367 . S2CID 257326012 .
- ^ Jump up to: а б с Виманн Дж., Фаббри М., Ян Т.Р., Стейн К., Сандер П.М., Норелл М.А., Бриггс Д.Э. (ноябрь 2018 г.). «Окаменение превращает белки твердых тканей позвоночных в N-гетероциклические полимеры» . Природные коммуникации . 9 (1): 4741. Бибкод : 2018NatCo...9.4741W . дои : 10.1038/s41467-018-07013-3 . ПМК 6226439 . ПМИД 30413693 .
- ^ Jump up to: а б с д Виманн Дж., Кроуфорд Дж.М., Бриггс Д.Э. (июль 2020 г.). «Филогенетические и физиологические сигналы в ископаемых биомолекулах многоклеточных животных» . Достижения науки . 6 (28): eaba6883. Бибкод : 2020SciA....6.6883W . дои : 10.1126/sciadv.aba6883 . ПМЦ 7439315 . ПМИД 32832604 .
- ^ Jump up to: а б Ботман Э.М., Гудвин М.Б., Холман Х.Н., Факра С., Чжэн В., Гронский Р., Швейцер М.Х. (октябрь 2019 г.). «Механизмы сохранения мягких тканей и белка у тираннозавра рекса» . Научные отчеты . 9 (1): 15678. Бибкод : 2019НатСР...915678Б . дои : 10.1038/s41598-019-51680-1 . ПМК 6821828 . PMID 31666554 .
- ^ Jump up to: а б Тан Ц, Пан К, Ли Г, Чен Л, Юань Икс, Сяо С (сентябрь 2021 г.). «Эпибионты возрастом один миллиард лет подчеркивают симбиотические экологические взаимодействия в ранней эволюции эукариот» . Исследования Гондваны . 97 : 22–33. Бибкод : 2021GondR..97...22T . дои : 10.1016/j.gr.2021.05.008 . ISSN 1342-937X .
- ^ Jump up to: а б с Абельсон П.Х. (1954). «Органические составляющие окаменелостей». Ежегодник Вашингтонского института Карнеги . 53 : 97–101.
- ^ Греха С. «Пресс-релиз премии Грубера по генетике 2013 г.» . Фонд Грубера.
- ^ Jump up to: а б де Йонг Э.В., Вестбрук П., Вестбрук Дж.В., Брюнинг Дж.В. (ноябрь 1974 г.). «Сохранение антигенных свойств макромолекул более 70 млн лет». Природа . 252 (5478): 63–64. Бибкод : 1974Natur.252...63D . дои : 10.1038/252063a0 . ПМИД 4139661 . S2CID 4286786 .
- ^ Jump up to: а б Вестбрук П., ван дер Мейде П.Х., ван дер Вей-Клопперс Дж.С., ван дер Слуис Р.Дж., де Леув Дж.В., де Йонг Э.В. (1979). «Ископаемые макромолекулы из панцирей головоногих моллюсков: характеристика, иммунологический ответ и диагенез». Палеобиология . 5 (2): 151–167. Бибкод : 1979Pbio....5..151W . дои : 10.1017/S0094837300006448 . S2CID 88664754 .
- ^ Jump up to: а б Нитецкий М.Х. (1982). Биохимические аспекты эволюционной биологии . Чикаго: Издательство Чикагского университета. стр. 29–91.
- ^ Jump up to: а б Герли Л.Р., Вальдес Дж.Г., Сполл В.Д., Смит Б.Ф., Джилетт Д.Д. (февраль 1991 г.). «Белки в ископаемых костях динозавра сейсмозавра». Журнал белковой химии . 10 (1): 75–90. дои : 10.1007/BF01024658 . ПМИД 2054066 . S2CID 9046093 .
- ^ Jump up to: а б Кайзер Дж. (октябрь 2005 г.). «Вирусология. Возрожденный вирус гриппа раскрывает тайны смертельной пандемии 1918 года» . Наука . 310 (5745): 28–29. дои : 10.1126/science.310.5745.28 . ПМИД 16210501 . S2CID 26252589 .
- ^ Jump up to: а б Рокфеллеровский университет (1 марта 2007 г.). «Древний ретровирус возрождается» . Наука Дейли . Проверено 9 апреля 2014 г.
- ^ Jump up to: а б с Краузе Дж., Фу К., Гуд Ю.М., Виола Б., Шунков М.В., Деревянко А.П., Паабо С. (апрель 2010 г.). «Полный геном митохондриальной ДНК неизвестного гоминина из южной Сибири» . Природа . 464 (7290): 894–897. Бибкод : 2010Natur.464..894K . doi : 10.1038/nature08976 (неактивен 21 мая 2024 г.). ПМЦ 10152974 . ПМИД 20336068 .
{{cite journal}}: CS1 maint: DOI неактивен по состоянию на май 2024 г. ( ссылка ) - ^ Jump up to: а б с Прюфер К., Расимо Ф., Паттерсон Н., Джей Ф., Санкарараман С., Сойер С. и др. (январь 2014 г.). «Полная последовательность генома неандертальца с Горного Алтая» . Природа . 505 (7481): 43–49. Бибкод : 2014Natur.505...43P . дои : 10.1038/nature12886 . ПМК 4031459 . ПМИД 24352235 .
- ^ Jump up to: а б с Каллауэй Э (декабрь 2013 г.). «ДНК гоминина ставит в тупик экспертов» . Природа . 504 (7478): 16–17. Бибкод : 2013Natur.504...16C . дои : 10.1038/504016а . ПМИД 24305130 .
- ^ Jump up to: а б Далтон Р. (май 2006 г.). «ДНК неандертальца уступает место вторжению в геном» . Природа . 441 (7091): 260–261. Бибкод : 2006Natur.441..260D . дои : 10.1038/441260b . ПМИД 16710377 .
- ^ Швейцер М.Х., Чжэн В., Клеланд Т.П., Гудвин М.Б., Ботман Э., Тиль Э., Маркус М.А., Факра СК (январь 2014 г.). «Роль химии железа и кислорода в сохранении мягких тканей, клеток и молекул в глубоком прошлом» . Труды Королевского общества B: Биологические науки . 281 (1775): 20132741. doi : 10.1098/rspb.2013.2741 . ПМЦ 3866414 . ПМИД 24285202 .
- ^ Jump up to: а б Циммер С (16 ноября 2015 г.). «В зубе ДНК некоторых очень старых родственников денисовцев» . Нью-Йорк Таймс . Проверено 16 ноября 2015 г.
- ^ Jump up to: а б Сойер С., Рено Г., Виола Б., Хаблин Дж.Дж., Гансоге М.Т., Шунков М.В. и др. (декабрь 2015 г.). «Последовательности ядерной и митохондриальной ДНК двух денисовцев» . Труды Национальной академии наук Соединенных Штатов Америки . 112 (51): 15696–15700. Бибкод : 2015PNAS..11215696S . дои : 10.1073/pnas.1519905112 . ПМЦ 4697428 . ПМИД 26630009 .
- ^ Виманн Дж., Бриггс Д.Э. (январь 2022 г.). «Раман-спектроскопия — мощный инструмент молекулярной палеобиологии: аналитический ответ Аллеону и др. (https://doi.org/10.1002/bies.202000295)» . Биоэссе . 44 (2): e2100070. doi : 10.1002/bies.202100070 . ПМИД 34993976 . S2CID 245824320 .
- ^ Хигучи Р., Боуман Б., Фрейбергер М., Райдер О.А., Уилсон А.С. (15 ноября 1984 г.). «Последовательности ДНК квагги, вымершего представителя семейства лошадей». Природа . 312 (5991): 282–284. Бибкод : 1984Natur.312..282H . дои : 10.1038/312282a0 . ПМИД 6504142 . S2CID 4313241 .
- ^ Jump up to: а б Райх Д., Грин Р.Э., Кирхер М., Краузе Дж., Паттерсон Н., Дюран Э.Ю. и др. (декабрь 2010 г.). «Генетическая история архаической группы гомининов из Денисовой пещеры в Сибири» . Природа . 468 (7327): 1053–1060. Бибкод : 2010Natur.468.1053R . дои : 10.1038/nature09710 . ПМК 4306417 . ПМИД 21179161 .
- ^ Пенниси Э (май 2013 г.). «Эволюция человека. Дополнительные геномы из Денисовой пещеры показывают смешение ранних групп человека». Наука . 340 (6134): 799. Бибкод : 2013Sci...340..799P . дои : 10.1126/science.340.6134.799 . ПМИД 23687020 .
- ^ Мунье А., Маршаль Ф., Кондеми С. (март 2009 г.). «Является ли Homo heidelbergensis отдельным видом? Новый взгляд на нижнюю челюсть Мауэра». Журнал эволюции человека . 56 (3): 219–246. Бибкод : 2009JHumE..56..219M . дои : 10.1016/j.jhevol.2008.12.006 . ПМИД 19249816 .
- ^ Картмилл М., Смит Ф.Х. (2009). Человеческая родословная . Джон Уайли и сыновья . ISBN 978-0471214915 . Проверено 21 апреля 2013 г.
- ^ Jump up to: а б Мейер М., Фу К., Аксиму-Петри А., Глок И., Никель Б., Арсуага Дж.Л. и др. (январь 2014 г.). «Последовательность митохондриального генома гоминина из Сима-де-лос-Уэсос» (PDF) . Природа . 505 (7483): 403–406. Бибкод : 2014Natur.505..403M . дои : 10.1038/nature12788 . ПМИД 24305051 . S2CID 4456221 . Архивировано из оригинала (PDF) 12 февраля 2014 года.
- ^ Циммер К. «Возвращение к жизни вымерших видов» . Нэшнл Географик. Архивировано из оригинала 12 декабря 2016 года . Проверено 26 марта 2014 г.
- ^ Грей Р. (31 января 2009 г.). «Вымерший козерог воскрешен путем клонирования» . Телеграф . Архивировано из оригинала 2 августа 2009 года . Проверено 9 апреля 2014 г.
- ^ Чёрч Г. «Мультиплексная автоматизированная геномная инженерия (MAGE): машина, ускоряющая эволюцию, производит революцию в дизайне генома» . Институт Висса. Архивировано из оригинала 12 октября 2016 года . Проверено 26 марта 2014 г.
- ^ Jump up to: а б Редакция (14 мая 2013 г.). «Почему попытки вернуть вымершие виды из мертвых упускают суть» . Научный американец . Проверено 9 апреля 2014 г.
{{cite news}}:|last=имеет общее имя ( справка ) - ^ Пимм С.Л., Рассел Г.Дж., Гиттлман Дж.Л., Брукс Т.М. (июль 1995 г.). «Будущее биоразнообразия». Наука . 269 (5222): 347–350. Бибкод : 1995Sci...269..347P . дои : 10.1126/science.269.5222.347 . ПМИД 17841251 . S2CID 35154695 .
- ^ Хоган М. «Причины вымирания» . Энциклопедия Земли . Проверено 9 апреля 2014 г.
- ^ Церковь G (20 августа 2013 г.). «Восстановление вымирания — хорошая идея» . Научный американец . Проверено 9 апреля 2014 г.
- ^ «Метод: Принципы» . Лаборатория аминокислотной геохронологии . Университет Северной Аризоны. Архивировано из оригинала 14 марта 2012 года . Проверено 26 марта 2014 г.
- ^ Jump up to: а б с Пойнар Х.Н., Хёсс М., Бада Дж.Л., Паабо С. (май 1996 г.). «Рацемизация аминокислот и сохранение древней ДНК». Наука . 272 (5263): 864–866. Бибкод : 1996Sci...272..864P . дои : 10.1126/science.272.5263.864 . hdl : 2060/19980202342 . ПМИД 8629020 . S2CID 20626782 .
- ^ Хенн Б.М., Жиньу Ч.Р., Фельдман М.В., Маунтин Дж.Л. (январь 2009 г.). «Характеристика временной зависимости оценок скорости мутаций митохондриальной ДНК человека» . Молекулярная биология и эволюция . 26 (1): 217–230. дои : 10.1093/molbev/msn244 . ПМИД 18984905 .
- ^ «Митохондриальная ДНК» . Обучение ДНК-аналитика . NFSTC Наука на службе справедливости. Архивировано из оригинала 1 февраля 2014 года . Проверено 23 апреля 2014 г.
- ^ Ланг Б.Ф., Грей М.В., Бургер Дж. (1999). «Эволюция митохондриального генома и происхождение эукариот». Ежегодный обзор генетики . 33 : 351–397. дои : 10.1146/annurev.genet.33.1.351 . ПМИД 10690412 .
- ^ Jump up to: а б Кано Р.Дж., Пойнар Х.Н. (сентябрь 1993 г.). «Быстрое выделение ДНК из ископаемых и музейных образцов, пригодных для ПЦР». БиоТехники . 15 (3): 432–4, 436. PMID 8217155 .
- ^ Jump up to: а б Тальявиа М, Масса Б, Альбанезе И, Ла Фарина М (29 марта 2011 г.). «Извлечение ДНК из музейных образцов прямокрылых» (PDF) . Аналитические письма . 44 (6): 1058–1062. дои : 10.1080/00032719.2010.506939 . hdl : 10447/57711 . S2CID 95380144 .
- ^ Jump up to: а б с д и Ходжес С.Б., Швейцер М.Х. Молекулярная палеонтология (PDF) . Энциклопедия палеонтологии. стр. 752–754.
- ^ Jump up to: а б Туросс Н. (июнь 1994 г.). «Биохимия древней ДНК в кости». Эксперименты . 50 (6): 530–535. дои : 10.1007/bf01921721 . ПМИД 7517371 . S2CID 27896535 .
- ^ Туросс Н., Статоплос Л. (1993). «[9] Древние белки в ископаемых костях». Древние белки в ископаемых костях . Методы энзимологии. Том. 224. стр. 121–129. дои : 10.1016/0076-6879(93)24010-р . ISBN 9780121821258 . ПМИД 8264383 .