Филогенетическое дерево
| Часть серии о |
| Эволюционная биология |
|---|
 |
Филогенетическое дерево , филогения или эволюционное дерево — это графическое представление, которое показывает историю эволюции между набором видов или таксонов в течение определенного времени. [ 1 ] [ 2 ] Другими словами, это диаграмма ветвления или дерево, показывающее эволюционные отношения между различными биологическими видами или другими объектами, основанные на сходстве и различиях в их физических или генетических характеристиках. В эволюционной биологии вся жизнь на Земле теоретически является частью единого филогенетического дерева, что указывает на общее происхождение . Филогенетика - это изучение филогенетических деревьев. Основная задача — найти филогенетическое дерево, представляющее оптимальное эволюционное происхождение между набором видов или таксонов. Вычислительная филогенетика (также филогенетический вывод) фокусируется на алгоритмах, участвующих в поиске оптимального филогенетического дерева в филогенетическом ландшафте. [ 1 ] [ 2 ]
Филогенетические деревья могут быть укорененными и неукорененными. В корневом филогенетическом дереве каждый узел с потомками представляет предполагаемого самого последнего общего предка этих потомков, [ 3 ] а длины ребер в некоторых деревьях можно интерпретировать как оценки времени. Каждый узел называется таксономической единицей. Внутренние узлы обычно называют гипотетическими таксономическими единицами, поскольку их нельзя наблюдать напрямую. Деревья полезны в таких областях биологии, как биоинформатика , систематика и филогенетика . Некорневые деревья иллюстрируют только родство листовых узлов и не требуют знания или предположения о родовом корне.
История
[ редактировать ]Идея древа жизни возникла из древних представлений о лестничном переходе от низших к высшим формам жизни (например, в Великой Цепи Бытия ). Ранние представления «ветвящихся» филогенетических деревьев включают «палеонтологическую карту», показывающую геологические взаимоотношения между растениями и животными в книге «Элементарная геология» Эдварда Хичкока (первое издание: 1840 г.).
Чарльз Дарвин представил схематическое эволюционное «дерево» в своей книге 1859 года «Происхождение видов» . Спустя столетие биологи-эволюционисты все еще используют древовидные диаграммы для изображения эволюции , поскольку такие диаграммы эффективно передают концепцию, согласно которой видообразование происходит посредством адаптивного и полуслучайного разделения линий.
Термин филогенетический , или филогения , происходит от двух древнегреческих слов φῦλον ( phûlon ), означающих «раса, родословная», и γένεσις ( genesis ), означающих «происхождение, источник». [ 4 ] [ 5 ]
Характеристики
[ редактировать ]Укорененное дерево
[ редактировать ]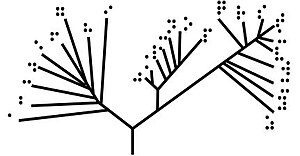
Корневое филогенетическое дерево (см. два рисунка вверху) — это ориентированное дерево с уникальным узлом — корнем — соответствующим (обычно вменяемому ) самому последнему общему предку всех объектов на листьях дерева. Корневой узел не имеет родительского узла, но является родительским для всех остальных узлов дерева. Таким образом, корень является узлом степени 2, в то время как другие внутренние узлы имеют минимальную степень 3 (где «степень» здесь относится к общему количеству входящих и исходящих ребер). [ нужна ссылка ]
Наиболее распространенным методом укоренения деревьев является использование непротиворечивой внешней группы — достаточно близкой, чтобы можно было сделать выводы на основе данных о признаках или молекулярного секвенирования, но достаточно далекой, чтобы быть явной внешней группой. Другой метод — укоренение средней точки. Кроме того, дерево можно укоренить с помощью модели нестационарной подстановки . [ 6 ]
Неукорененное дерево
[ редактировать ]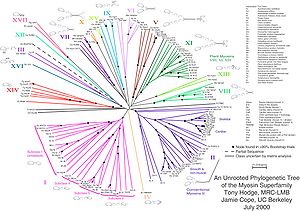
Некорневые деревья иллюстрируют родство листовых узлов, не делая предположений о происхождении. Они не требуют знания или предположения о родовом корне. [ 8 ] Некорневые деревья всегда можно создать из корневых, просто опуская корень. Напротив, определение корня неукорененного дерева требует некоторых средств идентификации происхождения. Обычно это делается путем включения внешней группы во входные данные, чтобы корень обязательно находился между внешней группой и остальными таксонами в дереве, или путем введения дополнительных предположений об относительных скоростях эволюции в каждой ветви, например приложения гипотезы часов . молекулярных [ 9 ]
Раздвоение против раздвоения
[ редактировать ]Как корневые, так и некорневые деревья могут быть как раздвоенными , так и многоветвистыми. Корневое разветвляющееся дерево имеет ровно два потомка, возникающих из каждого внутреннего узла (то есть оно образует бинарное дерево ), а некорневое разветвляющееся дерево принимает форму некорневого бифуркационного дерева , свободного дерева, имеющего ровно три соседа в каждом внутреннем узле. Напротив, корневое разветвленное дерево может иметь более двух дочерних элементов в некоторых узлах, а некорневое разветвленное дерево может иметь более трех соседей в некоторых узлах. [ нужна ссылка ]
Маркированные и немаркированные
[ редактировать ]Деревья с корнями и без корней могут быть как помеченными, так и не помеченными. Помеченное дерево имеет определенные значения, присвоенные его листьям, тогда как немаркированное дерево, иногда называемое формой дерева, определяет только топологию. Некоторые деревья на основе последовательностей, построенные на основе небольшого геномного локуса, такие как Phylotree, [ 10 ] имеют внутренние узлы, помеченные предполагаемыми предковыми гаплотипами.
Перечисление деревьев
[ редактировать ]
Число возможных деревьев для заданного количества листовых узлов зависит от конкретного типа дерева, но помеченных деревьев всегда больше, чем немаркированных, разветвленных деревьев больше, чем раздвоенных, и больше корневых деревьев, чем некорневых. Последнее различие наиболее значимо с биологической точки зрения; оно возникает потому, что на неукорененном дереве много мест, куда можно пустить корень. Для раздвоенных помеченных деревьев общее количество корневых деревьев равно:
- для , представляет количество конечных узлов. [ 11 ]



Для раздвоенных помеченных деревьев общее количество некорневых деревьев равно: [ 11 ]
- для .


Среди помеченных раздвоенных деревьев количество некорневых деревьев с листьев равно числу укорененных деревьев с листья. [ 2 ]

Число укоренившихся деревьев быстро растет в зависимости от количества кончиков. На 10 советов приходится более возможны раздвоенные деревья, причем число разветвляющихся деревьев растет быстрее, примерно с . Последних в 7 раз больше, чем первых.

| Маркированный листья |
Двоичный некорневые деревья |
Двоичный деревья с корнями |
Многофуркация деревья с корнями |
Все возможное деревья с корнями |
|---|---|---|---|---|
| 1 | 1 | 1 | 0 | 1 |
| 2 | 1 | 1 | 0 | 1 |
| 3 | 1 | 3 | 1 | 4 |
| 4 | 3 | 15 | 11 | 26 |
| 5 | 15 | 105 | 131 | 236 |
| 6 | 105 | 945 | 1,807 | 2,752 |
| 7 | 945 | 10,395 | 28,813 | 39,208 |
| 8 | 10,395 | 135,135 | 524,897 | 660,032 |
| 9 | 135,135 | 2,027,025 | 10,791,887 | 12,818,912 |
| 10 | 2,027,025 | 34,459,425 | 247,678,399 | 282,137,824 |
Особые виды деревьев
[ редактировать ]
Дендрограмма
[ редактировать ]Дендрограмма — это общее название дерева, филогенетического или нет, а, следовательно, и схематического представления филогенетического дерева. [ 12 ]
Кладограмма
[ редактировать ]Кладограмма представляет собой только образец ветвления; т. е. длина его ветвей не отражает время или относительную величину изменения символов, а его внутренние узлы не представляют предков. [ 13 ]
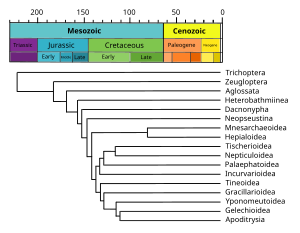
Филограмма
[ редактировать ]Филограмма — это филогенетическое дерево, длина ветвей которого пропорциональна степени изменения признаков. [ 15 ]
Хронограмма
[ редактировать ]Хронограмма — это филогенетическое дерево, которое явно представляет время через длину его ветвей. [ 16 ]
Дальгренограмма
[ редактировать ]Дальгренограмма — это диаграмма , представляющая собой поперечное сечение филогенетического дерева. [ нужна ссылка ]
Филогенетическая сеть
[ редактировать ]Филогенетическая сеть — это, строго говоря, не дерево, а скорее более общий граф или ориентированный ациклический граф в случае корневых сетей. Они используются для преодоления некоторых ограничений, присущих деревьям.
Схема шпинделя
[ редактировать ]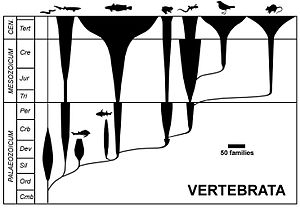
Диаграмму веретена, или пузырьковую диаграмму, часто называют ромерограммой, в честь ее популяризации американским палеонтологом Альфредом Ромером . [ 17 ] Он представляет таксономическое разнообразие (ширина по горизонтали) в зависимости от геологического времени (вертикальная ось), чтобы отразить изменение численности различных таксонов во времени. Диаграмма веретена не является эволюционным деревом: [ 18 ] таксономические веретена скрывают фактические отношения родительского таксона с дочерним таксоном [ 17 ] и имеют недостаток, связанный с парафилией родительской группы. [ 19 ] Этот тип диаграммы больше не используется в первоначально предложенной форме. [ 19 ]
Коралл жизни
[ редактировать ]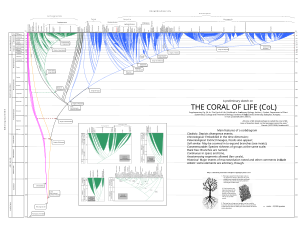
Дарвин [ 20 ] также упомянул, что коралл может быть более подходящей метафорой, чем дерево . Действительно, филогенетические кораллы полезны для изображения прошлой и настоящей жизни и имеют некоторые преимущества перед деревьями ( допускаются анастомозы и т. д.). [ 19 ]
Строительство
[ редактировать ]Филогенетические деревья, состоящие из нетривиального числа входных последовательностей, строятся с использованием вычислительной филогенетики методов . Методы матрицы расстояний, такие как соединение соседей или UPGMA , которые вычисляют генетическое расстояние на основе множественных выравниваний последовательностей , являются наиболее простыми в реализации, но не задействуют эволюционную модель. Многие методы выравнивания последовательностей, такие как ClustalW, также создают деревья, используя более простые алгоритмы (т. е. основанные на расстоянии) построения деревьев. Максимальная экономия — еще один простой метод оценки филогенетических деревьев, но он подразумевает неявную модель эволюции (т. е. экономию). Более продвинутые методы используют критерий оптимальности максимального правдоподобия , часто в рамках байесовской структуры , и применяют явную модель эволюции для оценки филогенетического дерева. [ 2 ] Определить оптимальное дерево с помощью многих из этих методов NP-сложно . [ 2 ] поэтому методы эвристического поиска и оптимизации используются в сочетании с функциями оценки дерева для определения достаточно хорошего дерева, соответствующего данным.
Методы построения деревьев можно оценить на основе нескольких критериев: [ 21 ]
- эффективность (сколько времени занимает вычисление ответа, сколько памяти требуется?)
- мощность (эффективно ли используются данные или информация тратится впустую?)
- последовательность (сойдется ли он к одному и тому же ответу неоднократно, если каждый раз давать разные данные для одной и той же модельной задачи?)
- надежность (хорошо ли он справляется с нарушениями предположений базовой модели?)
- фальсифицируемость (предупреждает ли нас, что его использовать нецелесообразно, т. е. когда предположения нарушаются?)
Методы построения деревьев также привлекли внимание математиков. Деревья также можно строить с помощью Т-теории . [ 22 ]
Форматы файлов
[ редактировать ]Деревья могут быть закодированы в различных форматах, каждый из которых должен представлять вложенную структуру дерева. Они могут кодировать или не кодировать длину ветвей и другие функции. Стандартизированные форматы имеют решающее значение для распространения и совместного использования деревьев, не полагаясь на графический вывод, который трудно импортировать в существующее программное обеспечение. Обычно используемые форматы:
Ограничения филогенетического анализа
[ редактировать ]Этот раздел нуждается в дополнительных цитатах для проверки . ( Октябрь 2012 г. ) |
Хотя филогенетические деревья, созданные на основе секвенированных генов или геномных данных у разных видов, могут дать представление об эволюции, этот анализ имеет важные ограничения. Самое главное, что деревья, которые они создают, не обязательно являются правильными – они не обязательно точно отражают историю эволюции включенных в них таксонов. Как и любой научный результат, они подлежат фальсификации путем дальнейшего изучения (например, сбора дополнительных данных, анализа существующих данных улучшенными методами). Данные, на которых они основаны, могут быть зашумлены ; [ 23 ] анализ может быть испорчен генетической рекомбинацией , [ 24 ] горизонтальный перенос генов , [ 25 ] гибридизация между видами, которые не были ближайшими соседями на дереве до того, как произошла гибридизация, и консервативные последовательности .
Кроме того, существуют проблемы с базированием анализа на одном типе признаков, например, на одном гене или белке , или только на морфологическом анализе, поскольку такие деревья, построенные на основе другого несвязанного источника данных, часто отличаются от первого, и поэтому требуется большая осторожность. в установлении филогенетических связей между видами. Это наиболее справедливо в отношении генетического материала, который подвергается латеральному переносу генов и рекомбинации , где разные блоки гаплотипов могут иметь разную историю. В этих типах анализа выходное дерево филогенетического анализа отдельного гена представляет собой оценку филогении гена (т. е. генного дерева), а не филогении таксонов ( т. е. дерева видов), из которых были выбраны эти признаки. в идеале оба должны быть очень близки. По этой причине в серьезных филогенетических исследованиях обычно используется комбинация генов, происходящих из разных геномных источников (например, из митохондриального или пластидного или ядерного генома). [ 26 ] или гены, которые, как ожидается, будут развиваться в различных режимах отбора, так что гомоплазия (ложная гомология ) вряд ли станет результатом естественного отбора.
Когда вымершие виды включаются в качестве конечных узлов в анализ (а не, например, для ограничения внутренних узлов), считается, что они не представляют прямых предков каких-либо существующих видов. Вымершие виды обычно не содержат высококачественной ДНК .
Диапазон полезных материалов ДНК расширился с развитием технологий экстракции и секвенирования. Разработка технологий, способных выводить последовательности из более мелких фрагментов или пространственных структур продуктов деградации ДНК, еще больше расширит диапазон ДНК, считающейся полезной.
Филогенетические деревья также могут быть выведены на основе ряда других типов данных, включая морфологию, наличие или отсутствие определенных типов генов, события вставки и делеции, а также любые другие наблюдения, которые, как считается, содержат эволюционный сигнал.
Филогенетические сети используются, когда раздвоенные деревья не подходят из-за этих осложнений, которые предполагают более сетчатую эволюционную историю отобранных организмов.
См. также
[ редактировать ]- Клад
- кладистика
- Вычислительная филогенетика
- Эволюционная биология
- Эволюционная таксономия
- Обобщенное выравнивание дерева
- Список программного обеспечения для филогенетики
- Список программного обеспечения для визуализации филогенетического дерева
- PANDIT , биологическая база данных, охватывающая белковые домены.
- Филогенетические сравнительные методы
- Филогенетическое примирение
- Таксономический ранг
- Tokogeny
Ссылки
[ редактировать ]- ^ Перейти обратно: а б Халафванд, Тайлер (2015). Поиск структуры в пространстве поиска филогении . Университет Далхаузи.
- ^ Перейти обратно: а б с д и Фельзенштейн Дж. (2004). Вывод о филогении Sinauer Associates: Сандерленд, Массачусетс.
- ^ Кинен, Т.; Вайнайна, Дж.; Майна, С.; Бойкин, Л. (21 апреля 2016 г.). «Укоренение деревьев, способы» . Энциклопедия эволюционной биологии : 489–493. дои : 10.1016/B978-0-12-800049-6.00215-8 . ISBN 9780128004265 . ПМК 7149615 .
- ^ Байи, Анатоль (1 января 1981 г.). Сокращено из Французско-греческого словаря . Париж: Хашетт. ISBN 978-2010035289 . OCLC 461974285 .
- ^ Байи, Анатоль. «Греко-французский словарь онлайн» . www.tabularium.be . Архивировано из оригинала 21 апреля 2014 года . Проверено 2 марта 2018 г.
- ^ Данг, Куонг Цао; Минь, Буй Куанг; Макши, Ханон; Мазель, Джоанна; Джеймс, Дженнифер Элеонора; Винь, Ле Си; Ланфир, Роберт (9 февраля 2022 г.). «nQMaker: оценка времени моделей необратимой замены аминокислот» . Систематическая биология . 71 (5): 1110–1123. дои : 10.1093/sysbio/syac007 . ПМЦ 9366462 . ПМИД 35139203 .
- ^ Ходж Т., Коуп М. (1 октября 2000 г.). «Генеалогическое древо миозина» . J Cell Sci . 113 (19): 3353–4. дои : 10.1242/jcs.113.19.3353 . ПМИД 10984423 . Архивировано из оригинала 30 сентября 2007 года.
- ^ « Факты о «дереве»: деревья с корнями и без корней» . Архивировано из оригинала 14 апреля 2014 г. Проверено 26 мая 2014 г.
- ^ У. Форд Дулитл (2002). «Искоренение Древа Жизни». Научный американец . 282 (2): 90–95. Бибкод : 2000SciAm.282b..90D . doi : 10.1038/scientificamerican0200-90 . ПМИД 10710791 .
Нет доступных тезисов
- ^ ван Овен, Маннис; Кайзер, Манфред (2009). «Обновленное комплексное филогенетическое древо глобальных вариаций митохондриальной ДНК человека» . Человеческая мутация . 30 (2): E386–E394. дои : 10.1002/humu.20921 . ПМИД 18853457 . S2CID 27566749 .
- ^ Перейти обратно: а б с Фельзенштейн, Джозеф (1 марта 1978 г.). «Число эволюционных деревьев» . Систематическая биология . 27 (1): 27–33. дои : 10.2307/2412810 . ISSN 1063-5157 . JSTOR 2412810 .
- ^ Фокс, Эмили. «Дендрограмма» . конечно . Архивировано из оригинала 28 сентября 2017 года . Проверено 28 сентября 2017 г.
- ^ Майр, Эрнст (1974) «Кладистический анализ или кладистическая классификация?». Журнал зоологической систематики и эволюционных исследований. 12: 94–128. doi:10.1111/j.1439-0469.1974.tb00160.x.
- ^ Лабандейра, CC; Дилчер, Д.Л.; Дэвис, доктор медицинских наук; Вагнер, Д.Л. (6 декабря 1994 г.). «Девяносто семь миллионов лет ассоциации покрытосеменных растений и насекомых: палеобиологическое понимание значения коэволюции» . Труды Национальной академии наук . 91 (25): 12278–12282. Бибкод : 1994PNAS...9112278L . дои : 10.1073/pnas.91.25.12278 . ISSN 0027-8424 . ПМК 45420 . ПМИД 11607501 .
- ^ Соарес, Антонио; Рабело, Рикардо; Дельбем, Александр (2017). «Оптимизация на основе анализа филограмм». Экспертные системы с приложениями . 78 : 32–50. дои : 10.1016/j.eswa.2017.02.012 . ISSN 0957-4174 .
- ^ Сантамария, Р.; Терон, Р. (26 мая 2009 г.). «Древовидная эволюция: визуальный анализ филогенетических деревьев» . Биоинформатика . 25 (15): 1970–1971. doi : 10.1093/биоинформатика/btp333 . ПМИД 19470585 .
- ^ Перейти обратно: а б «Эволюционная систематика: Веретенообразные диаграммы» . Палеос.com . 10.11.2014 . Проверено 07.11.2019 .
- ^ «Деревья, пузыри и копыта» . Мозг трехфунтовой обезьяны — Биология, программирование, лингвистика, филогения, систематика … 21 ноября 2007 г. Проверено 07.11.2019 .
- ^ Перейти обратно: а б с Подани, Янош (01.06.2019). «Коралл жизни» . Эволюционная биология . 46 (2): 123–144. Бибкод : 2019EvBio..46..123P . дои : 10.1007/s11692-019-09474-w . hdl : 10831/46308 . ISSN 1934-2845 .
- ^ Дарвин, Чарльз (1837). Тетрадь Б. п. 25.
- ^ Пенни, Д.; Хенди, доктор медицины; Сталь, Массачусетс (1992). «Прогресс в методах построения эволюционных деревьев». Тенденции экологии и эволюции . 7 (3): 73–79. дои : 10.1016/0169-5347(92)90244-6 . ПМИД 21235960 .
- ^ А. Дресс, К. Т. Хубер и В. Моултон. 2001. Метрические пространства в чистой и прикладной математике. Documenta Mathematica LSU 2001 : 121-139.
- ^ Таунсенд Дж. П., Су З, Текле Ю (2012). «Филогенетический сигнал и шум: прогнозирование силы набора данных для решения филогении». Генетика . 61 (5): 835–849. дои : 10.1093/sysbio/sys036 . ПМИД 22389443 .
- ^ Аренас М, Посада Д (2010). «Влияние рекомбинации на реконструкцию наследственных последовательностей» . Генетика . 184 (4): 1133–1139. дои : 10.1534/genetics.109.113423 . ПМЦ 2865913 . ПМИД 20124027 .
- ^ Вёзе С (2002). «Об эволюции клеток» . Proc Natl Acad Sci США . 99 (13): 8742–7. Бибкод : 2002PNAS...99.8742W . дои : 10.1073/pnas.132266999 . ПМЦ 124369 . ПМИД 12077305 .
- ^ Пархи, Дж.; Трипатия, ПС; Приядарши, Х.; Мандал, Южная Каролина; Панди, ПК (2019). «Диагностика митогенома для надежной филогении: случай группы рыб Cypriniformes». Джин . 713 : 143967. doi : 10.1016/j.gene.2019.143967 . ПМИД 31279710 . S2CID 195828782 .
Дальнейшее чтение
[ редактировать ]- Шух, RT и АВЗ Брауэр. 2009. Биологическая систематика: принципы и приложения (2-е изд.). ISBN 978-0-8014-4799-0
- Мануэль Лима , Книга деревьев: визуализация отраслей знаний , 2014, Princeton Architectural Press, Нью-Йорк.
- MEGA — бесплатная программа для рисования филогенетических деревьев.
- Гонтье, Н. 2011. «Изображение Древа Жизни: философские и исторические корни эволюционных древовидных диаграмм». Эволюция, Образование, Пропаганда 4: 515–538.
- Ян Сапп , Новые основы эволюции: на Древе жизни , 2009, Oxford University Press, Нью-Йорк.
Внешние ссылки
[ редактировать ]Изображения
[ редактировать ]- Y-хромосома человека 2002 Филогенетическое древо
- iTOL: интерактивное древо жизни
- Филогенетическое древо искусственных организмов, эволюционировавших на компьютерах. Архивировано 22 февраля 2016 г. на Wayback Machine.
- Филограмма эвтерийных млекопитающих Миямото и Гудмана.
Общий
[ редактировать ]- Обзор различных методов визуализации дерева доступен по адресу Пейдж, RDM (2011). «Пространство, время, форма: просмотр Древа Жизни». Тенденции в экологии и эволюции . 27 (2): 113–120. дои : 10.1016/j.tree.2011.12.002 . ПМИД 22209094 .
- OneZoom: Tree of Life – все живые виды в виде интуитивно понятного и масштабируемого исследователя фракталов (адаптивный дизайн)
- Откройте для себя жизнь Интерактивное дерево, основанное на проекте Национального научного фонда США «Сборка древа жизни».
- Филокод
- Множественное выравнивание 139 последовательностей миозина и филогенетическое древо
- Веб-проект «Древо жизни»
- Филогенетический вывод на сервере T-REX
- База данных таксономии NCBI [1]
- ETE: среда Python для исследования деревьев. Это программная библиотека для анализа, управления и визуализации филогенетических деревьев. Ссылка.
- Ежедневно обновляемое древо (последовательной) жизни Фанг, Х.; Оутс, Мэн; Петика, РБ; Гринвуд, Дж. М.; Сардар, Эй Джей; Рэкхэм, OJL; Донохью, PCJ; Стаматакис, А.; Де Лима Мораис, окружной прокурор; Гоф, Дж. (2013). «Ежедневно обновляемое древо (секвенированной) жизни как справочник для исследований генома» . Научные отчеты . 3 : 2015. Бибкод : 2013NatSR...3E2015F . дои : 10.1038/srep02015 . ПМК 6504836 . ПМИД 23778980 .