Общее происхождение
| Часть серии о |
| Эволюционная биология |
|---|
 |
Общее происхождение — это концепция эволюционной биологии, применимая, когда один вид является предком двух или более видов позже во времени. Согласно современной эволюционной биологии, все живые существа могут быть потомками уникального предка, которого обычно называют последним универсальным общим предком (LUCA) всей жизни на Земле . [1] [2] [3] [4]
Общее происхождение — это эффект видообразования , при котором несколько видов происходят от одной предковой популяции. Чем более поздней была общая предковая популяция двух видов, тем теснее они связаны. Самый недавний общий предок всех ныне живущих организмов — это последний универсальный предок, [3] которые жили около 3,9 миллиардов лет назад . [5] [6] Двумя самыми ранними свидетельствами существования жизни на Земле являются графит, который оказался биогенным возрастом 3,7 миллиарда лет, в метаосадочных породах обнаруженных в западной Гренландии. [7] и микробного мата окаменелости возрастом 3,48 миллиарда лет, , обнаруженные в песчанике обнаруженном в Западной Австралии . [8] [9] Все ныне живущие на Земле организмы имеют общее генетическое наследие, хотя предположение о значительном горизонтальном переносе генов в ходе ранней эволюции привело к вопросам монофилии (единого происхождения) жизни. [3] 6331 группа генов Идентифицирована , общих для всех ныне живущих животных; они, возможно, произошли от одного общего предка , жившего 650 миллионов лет назад в докембрии . [10] [11]
Всеобщее общее происхождение в результате эволюционного процесса было впервые предложено британским натуралистом Чарльзом Дарвином в заключительном предложении его книги «Происхождение видов» 1859 года :
Есть величие в этом взгляде на жизнь, с ее различными силами, первоначально вдохнутыми в несколько форм или в одну; и что, пока эта планета продолжает вращаться в соответствии с неизменным законом гравитации, из столь простого начала развились и продолжают развиваться бесчисленные формы, самые красивые и удивительные. [12]
История
[ редактировать ]Идея о том, что все живые существа (включая то, что наука считает неживыми) взаимосвязаны, является повторяющейся темой во многих мировоззрениях коренных народов по всему миру. [13] Позже, в 1740-х годах, французский математик Пьер Луи Мопертюи пришел к идее, что все организмы имели общего предка и разошлись в результате случайных изменений и естественного отбора . [14] [15]
В 1790 году философ Иммануил Кант написал в «Критике , что суждения» сходство [а] животных форм подразумевает общий первоначальный тип и, следовательно, общего родителя. [16]
В 1794 году дед Чарльза Дарвина, Эразм Дарвин, спросил:
[Было бы] слишком смело вообразить, что через большой промежуток времени, с тех пор как Земля начала существовать, возможно, за миллионы веков до начала истории человечества, было бы слишком смело вообразить, что все теплые -кровные животные произошли из одной живой нити, которая великая Первопричина, наделенная животностью, способностью приобретать новые части, сопровождаемые новыми склонностями, направляемыми раздражениями, ощущениями, волениями и ассоциациями; и, таким образом, обладать способностью продолжать совершенствоваться посредством своей собственной внутренней активности и передавать эти улучшения через поколения своим потомкам, миру без конца? [17]
Взгляды Чарльза Дарвина на общее происхождение, выраженные в книге «Происхождение видов» , заключались в том, что вполне вероятно, что существовал только один прародитель всех форм жизни:
Поэтому я должен заключить по аналогии, что, вероятно, все органические существа, когда-либо жившие на этой земле, произошли от какой-то одной первичной формы, в которую впервые была вдохнута жизнь. [18]
Но он предваряет это замечание словами: «Аналогия привела бы меня еще на шаг дальше, а именно к убеждению, что все животные и растения произошли от какого-то одного прототипа. Но аналогия может оказаться обманчивым руководством». И в последующем издании [19] скорее он утверждает,
«Мы не знаем всех возможных переходных градаций между простейшими и наиболее совершенными органами; нельзя притворяться, что мы знаем все разнообразные способы распространения в течение долгих лет или что мы знаем, насколько несовершенны геологические летописи. Какими бы серьезными ни были эти трудности, по моему мнению, они не опровергают теорию происхождения нескольких созданных форм с последующей модификацией».
Общее происхождение получило широкое признание в научном сообществе после публикации Дарвина. [20] В 1907 году Вернон Келлог заметил, что «практически ни один натуралист, обладающий положением и признанными достижениями, не сомневается в теории происхождения». [21]
В 2008 году биолог Т. Райан Грегори отметил, что:
Никогда не было обнаружено ни одного надежного наблюдения, противоречащего общему представлению об общем происхождении. Поэтому неудивительно, что научное сообщество в целом приняло эволюционное происхождение как историческую реальность со времен Дарвина и считает его одним из наиболее достоверно установленных и фундаментально важных фактов во всей науке. [22]
Доказательство
[ редактировать ]Общая биохимия
[ редактировать ]Все известные формы жизни основаны на одной и той же фундаментальной биохимической организации: генетическая информация, закодированная в ДНК , транскрибируется в РНК под действием белков- и РНК- ферментов , затем транслируется в белки (очень похожими) рибосомами с помощью АТФ , НАДФН. и другие в качестве источников энергии. Анализ небольших различий в последовательностях широко распространенных веществ, таких как цитохром c, еще раз подтверждает универсальное общее происхождение. [23] Во всех организмах обнаружено около 23 белков, служащих ферментами , выполняющими основные функции, такие как репликация ДНК. Тот факт, что существует только один такой набор ферментов, является убедительным доказательством единого происхождения. [3] [24] 6331 ген Идентифицирован , общий для всех ныне живущих животных; они, возможно, произошли от одного общего предка , жившего 650 миллионов лет назад в докембрии . [10] [11]
Общий генетический код
[ редактировать ]| Аминокислоты | неполярный | полярный | базовый | кислый | Стоп-кодон |
| 1-й база | 2-я база | |||||||
|---|---|---|---|---|---|---|---|---|
| Т | С | А | Г | |||||
| Т | ТТТ | Фенил- аланин | ТСТ | Серин | ТАТ | Тирозин | ТГТ | Цистеин |
| ТТС | ТСС | ТАС | ТГК | |||||
| ТТА | Лейцин | ТСА | СЕГОДНЯ | Останавливаться | ТГА | Останавливаться | ||
| ТТГ | ТКГ | ЯРЛЫК | Останавливаться | ТГГ | Триптофан | |||
| С | СТТ | ЦКТ | Пролин | КОТ | Гистидин | ВКТ | Аргинин | |
| СТС | CCC | САС | КГК | |||||
| призыв к действию | Приблизительно | САА | Глютамин | CGA | ||||
| КТГ | CCG | ЦАГ | КГГ | |||||
| А | К | изолейцин | ДЕЙСТВОВАТЬ | Треонин | ААТ | Аспарагин | ВОСЕМЬ | Серин |
| УВД | АСС | ААС | АРУ | |||||
| МИНУТЫ | АСА | ААА | Лизин | НО | Аргинин | |||
| АТГ | Метионин | АКГ | ААГ | АГГ | ||||
| Г | ГТТ | Валин | GCT | Аланин | ДЫРА | Аспарагиновая кислота кислота | ГГТ | Глицин |
| ГТК | GCC | ПКК | ГГК | |||||
| ГТА | ГКА | ИДТИ | Глутаминовый кислота | ГГА | ||||
| ГТГ | ГКГ | кляп | ГГГ | |||||
Генетический код («таблица перевода», согласно которой информация ДНК транслируется в аминокислоты , а значит, и в белки) практически идентичен для всех известных форм жизни, от бактерий и архей до животных и растений . Универсальность этого кода обычно рассматривается биологами как окончательное свидетельство в пользу универсального общего происхождения. [23]
Способ, которым кодоны (триплеты ДНК) сопоставляются с аминокислотами, по-видимому, сильно оптимизирован. Ричард Эгель утверждает, что, в частности, гидрофобные (неполярные) боковые цепи хорошо организованы, что позволяет предположить, что они позволили самым ранним организмам создавать пептиды с водоотталкивающими областями, способными поддерживать важные реакции электронного обмена ( окислительно-восстановительные реакции ) для передачи энергии. [25]
Избирательно нейтральное сходство
[ редактировать ]Сходства, не имеющие адаптивного значения, не могут быть объяснены конвергентной эволюцией , и поэтому они обеспечивают убедительную поддержку универсального общего происхождения. Такие доказательства были получены из двух областей: аминокислотных последовательностей и последовательностей ДНК. Белки с одинаковой трехмерной структурой не обязательно должны иметь идентичные аминокислотные последовательности; любое несущественное сходство между последовательностями является свидетельством общего происхождения. В некоторых случаях существует несколько кодонов (триплетов ДНК), которые избыточно кодируют одну и ту же аминокислоту. Поскольку многие виды используют один и тот же кодон в одном и том же месте для обозначения аминокислоты, которая может быть представлена более чем одним кодоном, это свидетельствует о том, что у них был недавний общий предок. Если бы аминокислотные последовательности произошли от разных предков, они бы кодировались любым из избыточных кодонов, а поскольку правильные аминокислоты уже были бы на месте, естественный отбор не вызвал бы никаких изменений в кодонах, какими бы значительными они ни были. время было в наличии. Генетический дрейф может изменить кодоны, но крайне маловероятно, чтобы все избыточные кодоны во всей последовательности точно совпадали в нескольких линиях. Точно так же общие нуклеотидные последовательности, особенно если они явно нейтральны, например расположение интронов и псевдогенов , служат убедительным доказательством общего происхождения. [26]
Другие сходства
[ редактировать ]Биологи часто [ количественно ] указывают на универсальность многих аспектов клеточной жизни в качестве подтверждающего доказательства более убедительных доказательств, перечисленных выше. Эти сходства включают в себя переносчик энергии аденозинтрифосфат (АТФ) и тот факт, что все аминокислоты, обнаруженные в белках, являются левосторонними . Однако возможно, что эти сходства возникли из-за законов физики и химии , а не из-за универсального общего происхождения, и, следовательно, привели к конвергентной эволюции. Напротив, есть доказательства гомологии центральных субъединиц трансмембранных АТФаз во всех живых организмах, особенно того, как вращающиеся элементы связаны с мембраной. Это подтверждает предположение о LUCA как о клеточном организме, хотя первичные мембраны могли быть полупроницаемыми и позже эволюционировали в мембраны современных бактерий, а также на втором пути - в мембраны современных архей. [27]
Филогенетические деревья
[ редактировать ]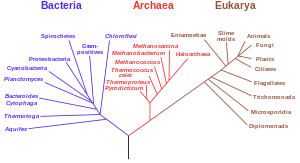
Еще одним важным доказательством являются подробные филогенетические деревья (т.е. «генеалогические деревья» видов), отображающие предполагаемые подразделения и общих предков всех ныне живущих видов. В 2010 году Дуглас Л. Теобальд опубликовал статистический анализ доступных генетических данных. [3] сопоставление их с филогенетическими деревьями, что дало «сильное количественное подтверждение единства жизни с помощью формального теста». [4]
Традиционно эти деревья строились с использованием морфологических методов, таких как внешний вид, эмбриология и т. д. Недавно стало возможным строить эти деревья, используя молекулярные данные, основанные на сходствах и различиях между генетическими и белковыми последовательностями. Все эти методы дают по существу схожие результаты, хотя большая часть генетических вариаций не влияет на внешнюю морфологию. То, что филогенетические деревья, основанные на различных типах информации, согласуются друг с другом, является убедительным доказательством реального общего происхождения. [28]
Возражения
[ редактировать ]Филогенетический анализ облаков генного обмена
[ редактировать ]Теобальд отметил, что существенный горизонтальный перенос генов мог произойти на ранней стадии эволюции. Бактерии сегодня сохраняют способность к обмену генами между отдаленно родственными линиями. Это ослабляет основное предположение филогенетического анализа о том, что сходство геномов подразумевает общее происхождение, поскольку достаточный обмен генами позволил бы линиям иметь большую часть своего генома независимо от того, был ли у них общий предок (монофилия) . Это привело к вопросам о едином происхождении жизни. [3] Однако биологи считают маловероятным, что совершенно неродственные протоорганизмы могли обмениваться генами, поскольку их разные механизмы кодирования привели бы лишь к искажению, а не к функционированию систем. Однако позже многие организмы, произошедшие от одного предка, могли легко иметь общие гены, которые работали одинаково, и, похоже, так оно и было. [3]
Конвергентная эволюция
[ редактировать ]Если бы ранние организмы под влиянием одних и тех же условий окружающей среды конвергентно развивали схожую биохимию , они могли бы независимо друг от друга приобрести сходные генетические последовательности. Соответственно, «формальный тест» Теобальда подвергся критике со стороны Такахиро Ёнезавы и его коллег. [29] за невключение рассмотрения конвергенции. Они утверждали, что теста Теобальда было недостаточно, чтобы отличить конкурирующие гипотезы. Теобальд защищал свой метод от этого утверждения, утверждая, что его тесты различают филогенетическую структуру и простое сходство последовательностей. Таким образом, утверждал Теобальд, его результаты показывают, что «настоящие универсально консервативные белки гомологичны ». [30] [31]
мир РНК
[ редактировать ]Выше упоминалась возможность того, что все живые организмы могут произойти от первоначального одноклеточного организма с ДНК геномом , и что это предполагает единое происхождение жизни. Хотя такой универсальный общий предок мог существовать, такая сложная сущность вряд ли возникла спонтанно из нежизни, и поэтому клетку с ДНК-геномом нельзя разумно рассматривать как источник жизни. Чтобы понять происхождение жизни, было высказано предположение, что клеточная жизнь, основанная на ДНК, произошла от относительно простых доклеточных самореплицирующихся молекул РНК, способных подвергаться естественному отбору . В ходе эволюции этот мир РНК сменился эволюционным возникновением мира ДНК. Мира независимо самореплицирующихся РНК-геномов, очевидно, больше не существует (РНК-вирусы зависят от клеток-хозяев с ДНК-геномами). Поскольку мир РНК, очевидно, исчез, неясно, как научные доказательства могут быть использованы для ответа на вопрос о том, существовало ли единое событие происхождения жизни, от которого произошла вся жизнь.
См. также
[ редактировать ]Библиография
[ редактировать ]- Кромби, AC ; Хоскин, Майкл [на словацком языке] (1970). «Научное движение и распространение научных идей, 1688–1751». В Бромли, Дж. С. (ред.). Возвышение Великобритании и России, 1688–1715/25 . Новая Кембриджская современная история . Том. 6. Лондон: Издательство Кембриджского университета . ISBN 978-0-521-07524-4 . LCCN 57014935 . OCLC 7588392 .
- Дарвин, Чарльз (1859). О происхождении видов путем естественного отбора, или о сохранении избранных рас в борьбе за жизнь (1-е изд.). Лондон: Джон Мюррей . LCCN 06017473 . OCLC 741260650 . Книга доступна в Полном собрании сочинений Чарльза Дарвина в Интернете . Проверено 23 ноября 2015 г.
- Дарвин, Эразм (1818 г.) [первоначально опубликовано в 1794 г.]. Зоономия; или Законы органической жизни . Том. 1 (4-е американское изд.). Филадельфия, Пенсильвания: Эдвард Эрл. Зоономия; или Законы органической жизни: в трех частях (Том 1) (1818 г.) в Интернет-архиве, дата обращения 23 ноября 2015 г.
- Харрис, К. Леон (1981). Эволюция: Бытие и откровения: с чтениями от Эмпедокла до Вильсона . Олбани, Нью-Йорк: Издательство Государственного университета Нью-Йорка . ISBN 978-0-87395-487-7 . LCCN 81002555 . OCLC 7278190 .
- Кант, Иммануил (1987) [Первоначально опубликовано в 1790 году в Пруссии под названием Kritik der Urteilskraft ]. Критика суждения . Перевод с введением Вернера С. Плухара; предисловие Мэри Дж. Грегор . Индианаполис, Индиана: Издательская компания Hackett . ISBN 978-0-87220-025-8 . LCCN 86014852 . OCLC 13796153 .
- Сокровище, Джеффри (1985). Становление современной Европы, 1648-1780 гг . Нью-Йорк: Метуэн . ISBN 978-0-416-72370-0 . LCCN 85000255 . OCLC 11623262 .
- Циглер, Генрих Эрнст (1904). Понятие инстинкта тогда и сейчас . Йена.
{{cite book}}: CS1 maint: отсутствует местоположение издателя ( ссылка )
Примечания
[ редактировать ]- ^ Теперь называется гомологией .
Ссылки
[ редактировать ]- ^ Вайс, Мэдлин С.; Соуза, Филиппа Л.; Мрнявац, Наталья; Нойкирхен, Синье; Реттгер, Мэйо; Нельсон-Сати, Шиджулал; Мартин, Уильям Ф. (25 июля 2016 г.). «Физиология и среда обитания последнего всеобщего общего предка» . Природная микробиология . 1 (9): 16116. doi : 10.1038/nmicrobiol.2016.116 . ISSN 2058-5276 . ПМИД 27562259 . S2CID 2997255 .
- ^ Фортерре, Патрик; Грибальдо, Симонетта; Брошье, Селин (октябрь 2005 г.). «[Лука: последний универсальный предок]» . Медицина/Науки . 21 (10): 860–865. дои : 10.1051/medsci/20052110860 . ISSN 0767-0974 . ПМИД 16197904 .
- ^ Перейти обратно: а б с д и ж г Теобальд, Дуглас Л. (13 мая 2010 г.). «Формальная проверка теории универсального общего происхождения». Природа . 465 (7295): 219–222. Бибкод : 2010Natur.465..219T . дои : 10.1038/nature09014 . ПМИД 20463738 . S2CID 4422345 .
- ^ Перейти обратно: а б Стил, Майк ; Пенни, Дэвид (13 мая 2010 г.). «Происхождение жизни: общее происхождение подвергнуто испытанию» . Природа . 465 (7295): 168–169. Бибкод : 2010Natur.465..168S . дои : 10.1038/465168a . ПМИД 20463725 . S2CID 205055573 .
- ^ Дулитл, В. Форд (февраль 2000 г.). «Искоренение Древа Жизни» (PDF) . Научный американец . 282 (2): 90–95. Бибкод : 2000SciAm.282b..90D . doi : 10.1038/scientificamerican0200-90 . ПМИД 10710791 . Архивировано из оригинала (PDF) 7 сентября 2006 г. Проверено 22 ноября 2015 г.
- ^ Глансдорф, Николас; Ин Сюй; Лабедан, Бернар (9 июля 2008 г.). «Последний универсальный общий предок: появление, конституция и генетическое наследие неуловимого предшественника» . Биология Директ . 3:29 . дои : 10.1186/1745-6150-3-29 . ПМЦ 2478661 . ПМИД 18613974 .
- ^ Отомо, Йоко; Какегава, Такеши; Исида, Акизуми; и др. (январь 2014 г.). «Свидетельства наличия биогенного графита в метаосадочных породах раннего архея Исуа». Природа Геонауки . 7 (1): 25–28. Бибкод : 2014NatGe...7...25O . дои : 10.1038/ngeo2025 .
- ^ Боренштейн, Сет (13 ноября 2013 г.). «Найдена самая старая окаменелость: познакомьтесь со своей микробной мамой» . Yahoo Новости . Ассошиэйтед Пресс . Архивировано из оригинала 10 ноября 2019 г. Проверено 22 ноября 2015 г.
- ^ Ноффке, Нора ; Кристиан, Дэниел; Уэйси, Дэвид; Хейзен, Роберт М. (16 декабря 2013 г.). «Микробно-индуцированные осадочные структуры, фиксирующие древнюю экосистему в формации Дрессер возрастом около 3,48 миллиардов лет, Пилбара, Западная Австралия» . Астробиология . 13 (12): 1103–1124. Бибкод : 2013AsBio..13.1103N . дои : 10.1089/ast.2013.1030 . ПМК 3870916 . ПМИД 24205812 .
- ^ Перейти обратно: а б Циммер, Карл (4 мая 2018 г.). «Самое первое животное появилось на фоне взрыва ДНК» . Нью-Йорк Таймс . Архивировано из оригинала 1 января 2022 г. Проверено 4 мая 2018 г.
- ^ Перейти обратно: а б Папс, Хорди; Холланд, Питер WH (30 апреля 2018 г.). «Реконструкция генома предков многоклеточных животных показывает увеличение новизны генома» . Природные коммуникации . 9 (1730 (2018)): 1730. Бибкод : 2018NatCo...9.1730P . дои : 10.1038/s41467-018-04136-5 . ПМЦ 5928047 . ПМИД 29712911 .
- ^ Дарвин 1859 , с. 490
- ^ Персонал, ИКТ (13 сентября 2018 г.). «Мы все связаны между собой: знания коренных народов, подтвержденные оцифрованным древом жизни» . Новости ИКТ . Проверено 5 мая 2021 г.
- ^ Кромби и Хоскин 1970 , стр. 62–63.
- ^ Сокровище 1985 , с. 142
- ^ Кант 1987 , с. 304 : «Несмотря на все разнообразие этих форм, они, кажется, были созданы в соответствии с общим архетипом, и эта аналогия между ними усиливает наше подозрение, что на самом деле они родственны, произведены общей первоначальной матерью».
- ^ Дарвин 1818 , с. 397 [§ 39.4.8]
- ^ Дарвин 1859 , с. 484
- ^ Дарвин, CR 1860. О происхождении видов посредством естественного отбора или о сохранении избранных рас в борьбе за жизнь. Лондон: Джон Мюррей. 2-е издание, второй выпуск, стр. 466.
- ^ Крог, Дэвид. (2005). Биология: Путеводитель по миру природы . Пирсон/Прентис Холл. п. 323. ISBN 978-0321946768 «Происхождение с модификациями было признано большинством ученых вскоре после публикации книги Дарвина «Происхождение видов посредством естественного отбора» в 1859 году. Ученые приняли это, потому что оно объяснило множество аспектов живого мира».
- ^ Келлог, Вернон Л. (1907). Дарвинизм сегодня . Генри Холт и компания. п. 3
- ^ Грегори, Т. Райан (2008). «Эволюция как факт, теория и путь» . Эволюция: образование и информационно-пропагандистская деятельность . 1 : 46–52. дои : 10.1007/s12052-007-0001-z .
- ^ Перейти обратно: а б Найт, Робин; Фриланд, Стивен Дж.; Ландвебер, Лаура Ф. (январь 2001 г.). «Перемонтирование клавиатуры: возможность развития генетического кода». Обзоры природы Генетика . 2 (1): 49–58. дои : 10.1038/35047500 . ПМИД 11253070 . S2CID 12267003 .
- ^ Тан, Кер (14 мая 2010 г.). «Все виды произошли из одной клетки, результаты исследования» . Нэшнл Географик . Архивировано из оригинала 15 мая 2010 года . Проверено 22 ноября 2017 г. .
- ^ Эгель, Ричард (март 2012 г.). «Первичный эукариогенез: об общей природе доклеточных состояний, предков современной жизни» . Жизнь . 2 (1): 170–212. Бибкод : 2012Life....2..170E . дои : 10.3390/life2010170 . ПМК 4187143 . ПМИД 25382122 .
- ^ Шарма, Н.С. (2005). Преемственность и эволюция животных . Публикации Миттала. стр. 32–. ISBN 978-81-8293-018-6 .
- ^ Лейн, Ник (2015). Жизненно важный вопрос: почему жизнь такая, какая она есть? . Профильные книги. ISBN 978-1781250365 .
- ^ Теобальд, Дуглас Л. «Предсказание 1.3: Согласованность независимых филогений» . 29+ доказательств макроэволюции: научное обоснование общего происхождения . Версия 2.89. Фонд TalkOrigins . Проверено 20 ноября 2009 г.
- ^ Ёнедзава, Такахиро; Хасегава, Масами (16 декабря 2010 г.). «Было ли доказано всеобщее общее происхождение?» . Природа . 468 (7326): Е9. Бибкод : 2010Natur.468E...9Y . дои : 10.1038/nature09482 . ПМИД 21164432 . S2CID 4318346 .
- ^ Теобальд, Дуглас Л. (16 декабря 2010 г.). «Ответ Теобальда» . Природа . 468 (7326): Е10. Бибкод : 2010Natur.468E..10T . дои : 10.1038/nature09483 . S2CID 4317014 .
- ^ Теобальд, Дуглас Л. (24 ноября 2011 г.). «Об универсальном общем происхождении, сходстве последовательностей и филогенетической структуре: грехи P-значений и достоинства байесовских доказательств» . Биология Директ . 6 (1): 60. дои : 10.1186/1745-6150-6-60 . ПМЦ 3314578 . ПМИД 22114984 .