Родительские инвестиции


Родительские инвестиции в эволюционной биологии и эволюционной психологии — это любые родительские затраты (например, время, энергия, ресурсы), приносящие пользу потомству . [1] [2] Родительские инвестиции могут осуществляться как мужчинами, так и женщинами (двойная родительская забота), только женщинами (исключительная материнская забота) или только мужчинами (исключительная отцовская забота ). Уход может быть предоставлен на любом этапе жизни потомства, от пренатального (например, охрана и инкубация яиц у птиц и плацентарное питание у млекопитающих) до послеродового периода (например, обеспечение питанием и защита потомства).
Теория родительских инвестиций, термин, придуманный Робертом Триверсом в 1972 году, предсказывает, что пол, который больше вкладывает в свое потомство, будет более избирательным при выборе партнера, а у пола, который меньше вкладывает средств, будет внутриполовая конкуренция за доступ к партнеру. Эта теория сыграла важную роль в объяснении половых различий в половом выборе и предпочтениях партнера во всем животном мире и у людей. [2]
История [ править ]
В 1859 году Чарльз Дарвин опубликовал книгу «Происхождение видов» . [3] концепцию естественного отбора Это представило миру , а также связанные с ней теории, такие как половой отбор . Впервые эволюционная теория была использована для объяснения того, почему самки «застенчивы», а самцы «пылки» и конкурируют друг с другом за внимание самок. В 1930 году Рональд Фишер написал «Генетическая теория естественного отбора» . [4] в котором он представил современную концепцию родительских инвестиций, представил гипотезу сексуального сына и представил принцип Фишера . В 1948 году Ангус Джон Бейтман опубликовал влиятельное исследование плодовых мух, в котором он пришел к выводу, что, поскольку производство женских гамет обходится дороже, чем мужские, репродуктивный успех самок ограничен способностью производить яйцеклетку, а репродуктивный успех самцов ограничен. был ограничен доступом к женщинам. [5] В 1972 году Триверс продолжил эту линию мышления, предложив теорию родительских инвестиций, которая описывает, как родительские инвестиции влияют на сексуальное поведение. Он заключает, что пол, который имеет более высокие родительские вложения, будет более избирательным при выборе партнера, а пол с более низкими вложениями будет конкурировать внутрисексуально за возможности спаривания. [2] В 1974 году Триверс расширил теорию родительских инвестиций, чтобы объяснить конфликт между родителями и потомками, конфликт между инвестициями, которые оптимальны с точки зрения родителей и детей. [6]
Родительская забота [ править ]
Теория родительских инвестиций — это раздел теории истории жизни . Самое раннее рассмотрение родительских инвестиций дано Рональдом Фишером в его книге 1930 года «Генетическая теория естественного отбора» . [7] при этом Фишер утверждал, что родительские расходы на потомство обоих полов должны быть равными. Клаттон-Брок расширил концепцию родительских инвестиций, включив в нее затраты на любой другой компонент родительской подготовленности. [ нужна ссылка ]
Самцы зайки , как правило, не делают различий между своим потомством и потомством другого самца в полиандрических или полигинандрических системах. Они увеличивают свой собственный репродуктивный успех за счет кормления потомства в зависимости от их собственного доступа к самке на протяжении всего периода спаривания, что обычно является хорошим показателем отцовства . [8] Такая неизбирательная родительская забота со стороны самцов наблюдается и у красногубых собачек . [9]

У некоторых насекомых родительские вложения самцов даются в форме брачного подарка. Например, самки декоративной моли во время совокупления получают от самца сперматофор, содержащий питательные вещества, сперму и защитные токсины. Этот дар, который может составлять до 10% массы тела самца, представляет собой общий родительский вклад, который вкладывает самец. [10]
У некоторых видов, таких как люди и многие птицы, потомство альтрициально и неспособно постоять за себя в течение длительного периода времени после рождения. У этих видов самцы вкладывают больше в свое потомство, чем родители-самцы скороспелых видов, поскольку в противном случае репродуктивный успех пострадает.
Выгоды от родительских инвестиций в потомство велики и связаны с влиянием на состояние, рост, выживание и, в конечном итоге, на репродуктивный успех потомства. Например, у цихлиды Tropheus moorii самка очень сильно вкладывает родительские силы в свое потомство, потому что она вынашивает детенышей во рту, а во время высиживания во рту все питание, которое она принимает, идет на кормление детенышей, и она фактически морит себя голодом. При этом ее детеныши становятся крупнее, тяжелее и быстрее, чем они были бы без него. Эти преимущества очень выгодны, поскольку снижают риск быть съеденными хищниками, а размер обычно является определяющим фактором в конфликтах за ресурсы. [11] Однако такие преимущества могут быть достигнуты за счет способности родителей воспроизводить потомство в будущем, например, из-за повышенного риска травм при защите потомства от хищников, потери возможностей спаривания при выращивании потомства и увеличения временного интервала до следующего воспроизводства.
Особый случай родительского вклада — это когда детеныши действительно нуждаются в питании и защите, но генетические родители фактически не участвуют в усилиях по воспитанию собственного потомства. Например, у Bombus terrestris бесплодные рабочие самки часто не размножаются самостоятельно, а вместо этого выращивают выводок своей матери. Это часто встречается у социальных перепончатокрылых из-за гаплодиплоидии , при которой самцы гаплоидны, а самки диплоидны. Это гарантирует, что сестры будут более связаны друг с другом, чем когда-либо со своими собственными потомками, что стимулирует их помогать воспитывать детенышей своей матери, а не своих собственных. [12]
В целом, родители отбираются таким образом, чтобы максимизировать разницу между выгодами и затратами, и родительская забота, вероятно, будет развиваться, когда выгоды превысят затраты.
Конфликт родителей и потомков [ править ]
Воспроизводство обходится дорого. Особи ограничены в степени, в которой они могут уделять время и ресурсы производству и воспитанию молодняка, и такие расходы также могут нанести ущерб их будущему состоянию, выживанию и дальнейшим репродуктивным результатам.Однако такие затраты обычно выгодны потомству, поскольку улучшают его состояние, выживаемость и репродуктивный успех. Эти различия могут привести к конфликту между родителями и потомками . Родители естественным образом отбираются таким образом, чтобы максимизировать разницу между выгодами и издержками, и родительская забота будет иметь тенденцию существовать тогда, когда выгоды существенно превышают затраты.
Родители в равной степени связаны со всем потомством, и поэтому, чтобы оптимизировать их приспособленность и шанс воспроизвести свои гены, они должны равномерно распределить свои инвестиции между нынешним и будущим потомством. Однако любой отдельный потомок больше связан с самим собой (они имеют 100% общей ДНК), чем со своими братьями и сестрами (братья и сестры обычно имеют 50% общей ДНК), поэтому для приспособленности потомства лучше всего, если родитель(и) вкладывают в них больше средств. Чтобы оптимизировать приспособленность, родитель хотел бы инвестировать в каждого потомка одинаково, но каждый потомок хотел бы получить большую долю родительских инвестиций. Родитель выбирается для инвестирования в потомство до тех пор, пока инвестиции в текущее потомство не станут дороже, чем инвестиции в будущее потомство. [13]
У итерородящих видов, у которых особи могут пережить несколько репродуктивных периодов в течение своей жизни, может существовать компромисс между инвестициями в нынешнее потомство и будущим воспроизводством. Родителям необходимо найти баланс между требованиями своих детей и собственными потребностями. Этот потенциальный негативный эффект родительской заботы был четко формализован Триверсом в 1972 году, который первоначально определил термин « родительские инвестиции» как означающий любые инвестиции родителя в отдельного потомка, которые увеличивают шансы потомства на выживание (и, следовательно, репродуктивный успех ) за счет способность родителя инвестировать в других потомков . [2]

Пингвины — яркий пример вида, который радикально жертвует своим здоровьем и благополучием в обмен на выживание своего потомства. Такое поведение, которое приносит пользу не только индивидууму, но и генетическому коду, из которого возникает индивидуум, можно увидеть у Королевского пингвина. Хотя некоторые животные действительно демонстрируют альтруистическое поведение по отношению к особям, не состоящим в прямом родстве, многие из этих видов поведения проявляются в основном в отношениях между родителями и потомками. Во время размножения самцы голодают в месте размножения в течение пяти недель, ожидая возвращения самки на свою инкубационную смену. Однако в этот период самцы могут решить отказаться от яйца, если самка задерживается с возвращением в места размножения. [14]
Это показывает, что эти пингвины изначально жертвуют собственным здоровьем в надежде увеличить выживаемость своего яйца. Но наступает момент, когда затраты самцов пингвинов становятся слишком высокими по сравнению с прибылью от успешного сезона размножения. Олоф Олссон исследовал корреляцию между количеством опыта размножения особи и продолжительностью ожидания, прежде чем особь откажется от яйца. Он предположил, что чем опытнее человек, тем лучше он сможет пополнить истощенные резервы тела, что позволит ему оставаться в яйце в течение более длительного периода времени. [14]
Принесение в жертву самцами своего веса и возможного выживания ради увеличения шансов на выживание их потомства является компромиссом между текущим репродуктивным успехом и будущим выживанием родителей. [14] Этот компромисс имеет смысл по сравнению с другими примерами альтруизма, основанного на родстве, и является ярким примером использования альтруизма в попытке повысить общую приспособленность генетического материала человека за счет будущего выживания человека.
матери и потомства инвестициях в Конфликт
Конфликт между матерью и потомством также изучался на животных и людях. Один такой случай был задокументирован в середине 1970-х годов этологом Вульфом Шифенхёвелем . Женщины эйпо на западе Новой Гвинеи участвуют в культурной практике, согласно которой они рожают недалеко от деревни. После рождения ребенка каждая женщина взвешивала, стоит ли ей оставить ребенка или оставить его в кустах поблизости, что неизбежно заканчивалось смертью ребенка. [15] Вероятность выживания и наличие ресурсов в деревне были факторами, которые повлияли на решение о том, оставлять ребенка или нет. Во время одного из показанных родов мать почувствовала, что ребенок слишком болен и не выживет, поэтому она завернула ребенка, готовясь оставить ребенка в кустах; однако, увидев, что ребенок движется, мать развернула ребенка и принесла его в деревню, продемонстрировав смену жизни и смерти. [15] Этот конфликт между матерью и ребенком привел к отстраненному поведению в Бразилии, о чем свидетельствует работа Шепера-Хьюза, поскольку «многие младенцы Альто остаются [редактированы] не только некрещеными, но и безымянными, пока не начнут ходить или говорить». [16] или если возник медицинский кризис и ребенку потребовалось срочное крещение . Этот конфликт между выживанием, как эмоциональным, так и физическим, вызвал сдвиг в культурных практиках, что привело к появлению новых форм вклада матери в ребенка.
забота Аллородительская
Аллородительская забота, также называемая «алломатерингом», представляет собой когда член сообщества, помимо биологических родителей младенца, участвует в уходе за потомством. [17] Под термин «аллородительский уход» подпадает ряд видов поведения, некоторые из которых: ношение, кормление, присмотр, защита и уход. Посредством аллородительской заботы можно уменьшить стресс для родителей, особенно матери, тем самым уменьшая негативное воздействие конфликта между родителями и детьми на мать. [18] Очевидный альтруистический характер поведения может показаться противоречащим теории естественного отбора Дарвина, поскольку забота о потомстве, которое не является собственным, не увеличит его прямую приспособленность, но отнимет время, энергию и ресурсы у воспитания собственного потомства. Однако такое поведение можно объяснить эволюционно как увеличение косвенной приспособленности, поскольку потомство, скорее всего, будет родственником, не являющимся потомком и, следовательно, несущим часть генетики аллородителя. [17]
и ситуации направление Потомство
Инвестиционное поведение родителей повышает шансы на выживание потомства и не требует, чтобы основные механизмы были совместимы с эмпатией, применимой к взрослым, или ситуациям, связанным с неродственным потомством, и не требует, чтобы потомство каким-либо образом отвечало взаимностью на альтруистическое поведение. [19] [20] Инвестирующие родители не более уязвимы перед эксплуатацией со стороны других взрослых.
Триверса Теория родительских инвестиций
Родительские инвестиции по определению Роберта Триверса в 1972 году. [21] это инвестиции родителя в потомство, которые увеличивают шансы потомства на выживание и, следовательно, репродуктивный успех за счет способности родителя инвестировать в другое потомство. Крупные родительские инвестиции в значительной степени снижают шансы родителей инвестировать в других потомков. Родительские инвестиции можно разделить на две основные категории: инвестиции в спаривание и инвестиции в воспитание. Инвестиции в спаривание состоят из полового акта и вложенных половых клеток. Инвестиции в выращивание — это время и энергия, затраченные на воспитание потомства после зачатия. У большинства видов родительский вклад самки в спаривание и воспитание значительно превосходит вклад самца. Что касается половых клеток (яйцеклеток и сперматозоидов), то вложением женщин обычно является большая часть как генетического материала, так и общей мужественности, в то время как мужчины обычно производят тысячи сперматозоидов ежедневно.
инвестиции Человеческие родительские
Человеческие женщины имеют фиксированный запас около 400 яйцеклеток , в то время как сперматозоиды у мужчин поставляются со скоростью двенадцать миллионов в час. [22] Кроме того, у женщин происходит оплодотворение и вынашивание беременности , и эти затраты перевешивают затраты мужчины на одну эффективную сперматозоидную клетку. Кроме того, для женщин один половой акт может привести к 38-недельному сроку беременности и последующим обязательствам, связанным с воспитанием, например, грудным вскармливанием . Из теории родительских инвестиций Трайверса следует несколько выводов. Первый вывод заключается в том, что женщины часто, но не всегда, больше инвестируют в секс. Тот факт, что они зачастую больше инвестируют в секс, приводит ко второму выводу: эволюция отдает предпочтение самкам, которые более избирательны в отношении своих партнеров, чтобы гарантировать, что половой акт не приведет к ненужным или расточительным затратам. Третий вывод заключается в том, что, поскольку женщины вкладывают больше средств и играют важную роль в репродуктивном успехе своего потомства, они являются ценным ресурсом для мужчин; в результате мужчины часто конкурируют за сексуальный доступ к женщинам.
как более инвестирующий секс Мужчины
Для многих видов единственным типом полученных мужских инвестиций являются половые клетки. В этом плане инвестиции женщин значительно превышают инвестиции мужчин, как упоминалось ранее. Однако есть и другие способы, которыми самцы вкладывают деньги в свое потомство. Например, самец может находить пищу, как на примере воздушных шаров. [23] Он может найти безопасную среду, в которой самка сможет кормиться или откладывать яйца, как это видно на примере многих птиц. [24] [25]
Он также может защищать детенышей и предоставлять им возможность учиться в юном возрасте, как это происходит со многими волками. В целом, основная роль, которую берут на себя самцы, — это защита самки и ее детенышей. Это часто может уменьшить несоответствие инвестиций, вызванное первоначальным инвестированием половых клеток. Есть некоторые виды, такие как мормонский сверчок , морской конек-игла и самцы панамской ядовитой стрелы-лягушки, которые вкладывают больше средств. Среди видов, в которые больше вкладывает самец, самец также является более разборчивым полом, предъявляя более высокие требования к избранной самке. Например, самка, которую они часто выбирают, обычно содержит на 60% больше яиц, чем отвергнутые самки. [26]
Это связывает теорию родительских инвестиций (PIT) с половым отбором : если родительские инвестиции больше для самца, чем для самки, обычно самка конкурирует за партнера, как показывают Phalaropidae и полиандрические виды птиц. Самки этих видов обычно более агрессивны, ярко окрашены и крупнее самцов. [27] предполагая, что более инвестирующий пол имеет больший выбор при выборе партнера по сравнению с полом, участвующим в внутриполовом отборе.
Женщины как ценный ресурс для мужчин [ править ]
Второе предсказание, вытекающее из теории Триверса, заключается в том, что тот факт, что самки вкладывают больше средств в потомство, делает их ценным ресурсом для самцов, поскольку обеспечивает выживание их потомства, что является движущей силой естественного отбора . Следовательно, пол, который вкладывает меньше средств в потомство, будет конкурировать между собой за размножение с полом, вкладывающим больше средств. Другими словами, самцы будут конкурировать за самок. Утверждалось, что ревность возникла для предотвращения риска потенциальной потери родительских инвестиций в потомство. [22]
Если самец перенаправляет свои ресурсы на другую самку, это означает дорогостоящую потерю времени, энергии и ресурсов для ее потомства. Однако риски для мужчин выше, поскольку, хотя женщины вкладывают больше средств в свое потомство, у них больше уверенности в материнстве, поскольку они сами вынашивают ребенка. Однако самцы никогда не могут иметь 100%-ную уверенность в отцовстве и, следовательно, рискуют вкладывать ресурсы и время в потомство, которое генетически не связано. Эволюционная психология рассматривает ревность как адаптивный ответ на эту проблему.
Применение теории Триверса в реальной жизни [ править ]
Теория Триверса оказала большое влияние, поскольку ее предсказания соответствуют различиям в сексуальном поведении мужчин и женщин, о чем свидетельствуют различные исследования. Межкультурное исследование Басса (1989). [28] показывает, что самцы настроены на физическую привлекательность, поскольку это сигнализирует о молодости и плодовитости и обеспечивает мужской репродуктивный успех, который увеличивается за счет совокупления с как можно большим количеством плодовитых самок. Женщины, с другой стороны, настроены на ресурсы, предоставляемые потенциальными партнерами, поскольку их репродуктивный успех увеличивается за счет обеспечения выживания их потомства, и один из способов сделать это — получить ресурсы для них. С другой стороны, другое исследование показывает, что мужчины более распущены, чем женщины, что еще раз подтверждает эту теорию. Кларк и Хэтфилд [29] обнаружили, что 75% мужчин были готовы заняться сексом с незнакомкой женщиной, когда ей было предложено, по сравнению с 0% женщин. С другой стороны, 50% женщин согласились на свидание с незнакомцем мужского пола. Это говорит о том, что мужчины стремятся к краткосрочным отношениям, в то время как женщины отдают предпочтение долгосрочным отношениям.
Однако эти предпочтения (мужская распущенность и женская разборчивость) можно объяснить и по-другому. В западных культурах мужская распущенность поощряется посредством доступности порнографических журналов и видеороликов, ориентированных на мужскую аудиторию. С другой стороны, как западная, так и восточная культуры препятствуют женской распущенности с помощью социальных ограничений, таких как позор шлюх . [30]
PIT (Теория родительских инвестиций) также объясняет закономерности сексуальной ревности . [22] Мужчины с большей вероятностью будут проявлять стрессовую реакцию, представляя, как их партнеры проявляют сексуальную неверность (вступают в сексуальные отношения с кем-то другим), а женщины демонстрируют больший стресс, когда представляют своего партнера эмоционально неверным (влюбленным в другую женщину). PIT объясняет это тем, что сексуальная неверность женщины снижает отцовскую уверенность мужчины, поэтому он будет проявлять больший стресс из-за страха перед рогоносцем . С другой стороны, женщина боится потерять ресурсы, которые предоставляет ей партнер. Если у ее партнера есть эмоциональная привязанность к другой самке, вполне вероятно, что он не будет так много вкладывать в свое потомство, поэтому в этих обстоятельствах проявляется более сильная реакция на стресс.
Резкая критика теории исходит от ее анализа Торнхилла и Палмера в книге «Естественная история изнасилования: биологические основы сексуального принуждения» . [31] поскольку это, кажется, оправдывает изнасилование и сексуальное принуждение женщин. Торнхилл и Палмер заявили, что изнасилование — это развитый метод поиска партнера в среде, где женщины выбирают партнера. Поскольку PIT утверждает, что самцы стремятся совокупиться с как можно большим количеством плодовитых самок, выбор, который есть у женщин, может отрицательно повлиять на репродуктивный успех самцов. Если бы женщины не выбирали себе партнеров, Торнхилл и Палмер утверждают, что изнасилований не было бы. При этом игнорируются различные социокультурные факторы, такие как тот факт, что насилуют не только плодовитых женщин – 34% несовершеннолетних жертв изнасилования моложе 12 лет, [32] а это означает, что они не достигли фертильного возраста, поэтому в их изнасиловании нет эволюционного преимущества. 14% изнасилований в Англии совершаются над мужчинами. [33] кто не может увеличить репродуктивный успех мужчины, так как не будет зачатия. Таким образом, то, что Торнхилл и Палмер назвали «развитым механизмом», может оказаться не очень выгодным.
стратегий сексуальных Против
Теория Триверс упускает из виду, что у женщин действительно есть краткосрочные отношения, например, на одну ночь, в то время как не все мужчины ведут себя беспорядочно. Альтернативным объяснением PIT (теории родительских инвестиций) и предпочтений партнера может быть теория сексуальных стратегий Басса и Шмитта. [34] SST утверждает, что оба пола стремятся к краткосрочным и долгосрочным отношениям, но ищут разные качества в своих краткосрочных и долгосрочных партнерах. Для краткосрочных отношений женщины предпочтут привлекательного партнера, но в долгосрочных отношениях они могут быть готовы пожертвовать этой привлекательностью ради ресурсов и обязательств. С другой стороны, мужчины могут принять сексуально заинтересованного партнера в краткосрочных отношениях, но, чтобы обеспечить свою отцовскую уверенность, вместо этого они будут искать верного партнера.
Международная политика [ править ]
Теория родительских инвестиций не только используется для объяснения эволюционных явлений и человеческого поведения, но также описывает повторяющиеся события в международной политике. В частности, родительские инвестиции упоминаются при описании конкурентного поведения между государствами и определении агрессивного характера внешней политики. Гипотеза родительских инвестиций утверждает, что размер коалиций и физическая сила ее членов-мужчин определяют, будут ли ее действия по отношению к иностранным соседям агрессивными или дружелюбными. [35] По словам Триверс, у мужчин были относительно низкие родительские инвестиции, и поэтому они были вынуждены оказаться в более жесткой конкурентной ситуации из-за ограниченных репродуктивных ресурсов. Половой отбор имел место естественным образом, и мужчины эволюционировали, чтобы решить свои уникальные репродуктивные проблемы. Помимо других адаптаций, мужская психология также развилась, чтобы напрямую помогать мужчинам в такой внутрисексуальной конкуренции. [35]
Одним из важных психологических событий стало принятие решения о том, бежать ли или активно вступить в войну с другой конкурирующей группой. Двумя основными факторами, на которые мужчины ссылались в таких ситуациях, были (1) является ли коалиция, частью которой они являются, больше, чем ее оппозиция, и (2) обладают ли мужчины в их коалиции большей физической силой, чем другие. Мужская психология, переданная в древнем прошлом, была передана в наше время, заставляя мужчин частично думать и вести себя так, как они это делали во время войн предков. Согласно этой теории, лидеры международной политики не были исключением. Например, Соединенные Штаты рассчитывали выиграть войну во Вьетнаме благодаря своей большей военной мощи по сравнению с врагами. Однако победа, согласно традиционному правилу увеличения размера коалиции, не произошла потому, что США не учли должным образом другие факторы, такие как настойчивость местного населения. [35]
Гипотеза родительских инвестиций утверждает, что мужская физическая сила коалиции по-прежнему определяет агрессивность современных конфликтов между государствами. Хотя эта идея может показаться необоснованной, учитывая, что мужская физическая сила является одним из наименее определяющих аспектов современной войны, человеческая психология, тем не менее, развилась, чтобы действовать на этой основе. Более того, хотя может показаться, что мотивация поиска партнера больше не является определяющим фактором, в современных войнах сексуальность, такая как изнасилование, несомненно очевидна в конфликтах даже по сей день. [35]

Половой отбор [ править ]
У многих видов самцы могут производить большее количество потомков в течение своей жизни, сводя к минимуму родительские вложения в пользу затрат времени на оплодотворение любой плодовитой самки репродуктивного возраста. Напротив, самка может иметь гораздо меньшее количество потомков в течение своей репродуктивной жизни, отчасти из-за более высоких обязательств родителей. Самки будут более избирательны («разборчивы») в отношении партнеров, чем самцы, выбирая самцов с хорошей приспособленностью (например, генами, высоким статусом, ресурсами и т. д.), чтобы компенсировать отсутствие прямых родительских инвестиций со стороны самцов. и, следовательно, повысить репродуктивный успех. Роберта Триверса Теория родительского вклада предсказывает, что пол, вносящий наибольшие вклады в лактацию , воспитание и защиту потомства, будет более разборчивым в спаривании ; и что пол, который меньше инвестирует в потомство, будет конкурировать посредством внутриполового отбора за доступ к полу, вкладывающему больше средств (см. принцип Бейтмана). [36] ).
Ожидается, что у видов, где оба пола уделяют большое внимание родительской заботе, возникает взаимный выбор. Примером этого являются хохлатые конюги , у которых родители несут равную ответственность за высиживание единственного яйца и воспитание птенца. У хохлатых конюг орнаментированы оба пола. [37]
в человека вклад Родительский
В ходе эволюции люди увеличили уровень родительского вклада, как биологического, так и поведенческого. Плод требует больших инвестиций со стороны матери, а новорожденный требует больших инвестиций со стороны общества. обладают виды, у которых новорожденные детеныши не способны самостоятельно передвигаться и требуют родительской опеки Высокой степенью альтрициальности . Человеческие дети рождаются неспособными позаботиться о себе и требуют дополнительных родительских усилий после рождения, чтобы выжить. [38]
инвестиции Материнские
Триверс (1972) [2] выдвинули гипотезу, что увеличение инвестиций, обусловленных биологическими обязательствами, предсказывает увеличение добровольных инвестиций. Матери вкладывают внушительные суммы в своих детей еще до их рождения. Время и питательные вещества, необходимые для развития плода, а также риски, связанные как с введением этих питательных веществ, так и с родами, представляют собой значительные инвестиции. Чтобы убедиться, что эти инвестиции не напрасны, матери, скорее всего, будут вкладывать средства в своих детей после их рождения, чтобы быть уверенными, что они выживут и добьются успеха. По сравнению с большинством других видов, человеческие матери отдают больше ресурсов своему потомству с более высоким риском для собственного здоровья, даже до рождения ребенка. Это связано с развитием более медленного жизненного цикла, при котором через более длительные интервалы рождается меньшее количество более крупных потомков, что требует увеличения родительских вложений. [39] [40]
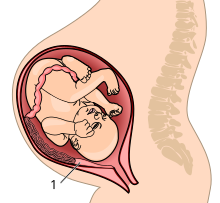
Развивающемуся человеческому плоду – и особенно мозгу – для роста необходимы питательные вещества. На более поздних неделях беременности плоду требуется все больше питательных веществ по мере увеличения роста мозга. [41] Грызуны и приматы имеют наиболее инвазивный фенотип плаценты — гемохориальную плаценту, при которой хорион разрушает эпителий матки и имеет прямой контакт с материнской кровью. Другие плацентарные фенотипы отделены от материнского кровотока по крайней мере одним слоем ткани. Более инвазивная плацента обеспечивает более эффективную передачу питательных веществ между матерью и плодом, но она также сопряжена с риском. Плод способен выделять гормоны непосредственно в кровоток матери, чтобы «потребовать» увеличения ресурсов. Это может привести к проблемам со здоровьем матери, например, к преэклампсии . Во время родов отслойка хориона плаценты может вызвать обильное кровотечение. [42]
Акушерская дилемма также усложняет роды и приводит к увеличению материнских инвестиций. Люди развили как прямохождение, так и большой размер мозга. Эволюция прямохождения изменила форму таза и уменьшила родовые пути, в то время как мозг стал больше. Уменьшение размера родовых путей означало, что дети рождаются на более ранних стадиях развития, когда их мозг меньше. Люди рожают детенышей с мозгом, развитым на 25%, тогда как другие приматы рожают потомство с мозгом, развитым на 45-50%. [43] Второе возможное объяснение ранних родов у людей — это энергия, необходимая для роста и поддержания большего мозга. Поддержка большего мозга во время беременности требует энергии, которую мать, возможно, не сможет вложить. [44]
Акушерская дилемма усложняет роды, а отличительной чертой человека является потребность в помощи во время родов. Измененная форма двуногого таза требует, чтобы детеныши покидали родовые пути лицом в сторону от матери, в отличие от всех других видов приматов. Из-за этого матери становится сложнее очистить дыхательные пути ребенка, убедиться, что пуповина не обмотана вокруг шеи, и вытащить ребенка, не сгибая его тело в неправильную сторону. [45]
Потребность человека в акушерке также требует социальности . Чтобы гарантировать присутствие акушерки, люди должны объединяться в группы. Спорно утверждалось, что люди обладают эусоциальностью . [46] как муравьи и пчелы, в которых родительский вклад относительно высок, совместная забота о потомстве и разделение труда. Неясно, что развилось первым; социальность, прямохождение или присутствие на родах. Бонобо , наши ближайшие ныне живущие родственники наряду с шимпанзе , отличаются высокой социальностью самок, и рождение детей среди бонобо также является социальным событием. [47] [48] Социальность могла быть предпосылкой для родовспоможения, а прямохождение и родовспоможение могли возникнуть еще пять миллионов лет назад. [38]
С возрастом самки приматов их способность к размножению снижается. Гипотеза бабушки описывает эволюцию менопаузы, которая может быть, а может и не быть уникальной для людей среди приматов. [49] По мере старения женщин затраты на инвестиции в дополнительное воспроизводство увеличиваются, а выгоды уменьшаются. В период менопаузы выгоднее остановить размножение и начать инвестировать во внуков. Бабушки уверены в своем генетическом родстве с внуками, особенно с детьми дочерей, поскольку материнская уверенность в собственных детях высока, а дочери уверены в своем материнстве и в отношении своих детей. Также было высказано предположение, что бабушки предпочитают инвестировать в дочерей своих дочерей, потому что Х-хромосомы несут больше ДНК, а их внучки наиболее тесно связаны с ними. [50]
инвестиции Отцовские
По мере того, как альтрициальность увеличивалась, инвестиции со стороны других лиц, помимо матери, становились все более необходимыми. Высокая социальность означала, что родственники женского пола присутствовали, чтобы помочь матери, но отцовский вклад также увеличился. Отцовские инвестиции увеличиваются по мере того, как становится все труднее иметь дополнительных детей, а также по мере того, как увеличивается влияние инвестиций на приспособленность потомства. [51]
Мужчины с большей вероятностью, чем женщины, не вкладывают родительских средств в своих детей, а дети отцов с низкими инвестициями с большей вероятностью будут вкладывать меньше родительских вложений в своих собственных детей. Отсутствие отца является фактором риска как для ранней сексуальной активности, так и для подростковой беременности. [52] [53] [54] [55] Отсутствие отца повышает уровень стресса у детей, что связано с более ранним началом половой жизни и усилением ориентации на краткосрочный спаривание. [56] [57] [58] [59] [60] Дочери отсутствующих отцов с большей вероятностью будут искать краткосрочных партнеров, и одна теория объясняет это предпочтением внешней (непартнерской) социальной поддержки из-за предполагаемого неопределенного будущего и неопределенного наличия заинтересованных партнеров в условиях высокого стресса. [61]
стратегий как предиктор спаривания Инвестиции

Скрытая овуляция [ править ]
Женщины могут забеременеть только во время овуляции. У человека овуляция скрыта или не сигнализируется извне. Скрытая овуляция снижает уверенность в отцовстве, поскольку мужчины не уверены, когда у женщины овуляция. [62] Предполагается, что эволюция скрытой овуляции является результатом альтрициальности и повышенной потребности в отцовских вложениях: если мужчины не уверены в времени овуляции, лучшим способом успешного воспроизводства будет неоднократное спаривание с женщиной на протяжении всего ее цикла, что требует парных связей, что, в свою очередь, увеличивает отцовские инвестиции. [63]
Ориентация спаривания [ править ]
Социосексуальность была впервые описана Альфредом Кинси как готовность вступать в случайные и ничем не обусловленные сексуальные отношения. [64] Социосексуальная ориентация описывает социосексуальность по шкале от неограниченной до ограниченной. Лица с неограниченной социосексуальной ориентацией имеют более высокую открытость к сексу в менее преданных отношениях, а лица с ограниченной социосексуальной ориентацией имеют меньшую открытость к случайным сексуальным связям. [65] [66] Однако сегодня признано, что социосексуальность в действительности не существует в одномерном масштабе. Люди, которые менее открыты для случайных связей, не всегда ищут преданных отношений, а люди, которые менее заинтересованы в серьезных отношениях, не всегда заинтересованы в случайных отношениях. [67] Краткосрочная и долгосрочная брачная ориентация являются современными дескрипторами открытости к свободным и преданным отношениям соответственно. [68]
Теория родительских инвестиций, предложенная Триверсом, утверждает, что пол с более высокими обязательными инвестициями будет более избирательным в выборе сексуальных партнеров, а пол с более низкими обязательными инвестициями будет менее избирательным и более заинтересованным в «случайных» возможностях спаривания. Пол с большим вкладом не может воспроизводиться так часто, в результате чего пол с меньшим вкладом конкурирует за возможности спаривания. [21] [69] У человека женщины имеют более высокие обязательные вложения ( беременность и роды), чем мужчины ( выработка спермы ). [34] Женщины с большей вероятностью будут иметь более высокую долгосрочную ориентацию к спариванию, а мужчины с большей вероятностью будут иметь более высокую краткосрочную ориентацию к спариванию. [67]
Краткосрочные и долгосрочные брачные ориентации влияют на предпочтения женщин в отношении мужчин. Исследования показали, что женщины уделяют большое внимание карьерной ориентации, амбициям и преданности только тогда, когда рассматривают долгосрочного партнера. [70] Когда речь не идет о браке, женщины уделяют больше внимания физической привлекательности. [71] Как правило, женщины предпочитают мужчин, которые, вероятно, будут выполнять большие родительские вложения и иметь хорошие гены. Женщины предпочитают мужчин с хорошим финансовым положением, более целеустремленных, более спортивных и здоровых. [72]
Некоторые неточные теории были основаны на теории родительских инвестиций. «Гипотеза структурного бессилия» [73] предлагает, чтобы женщины стремились найти партнеров с доступом к высоким уровням ресурсов, потому что, будучи женщинами, они напрямую исключены из этих ресурсов. Однако эта гипотеза была опровергнута исследованиями, которые показали, что финансово успешные женщины придают еще большее значение финансовому положению, социальному статусу и обладанию профессиональными степенями. [74]

Половой диморфизм [ править ]
Половой диморфизм - это разница в размерах тела между самцами и самками одного вида в результате как межполового, так и внутриполового отбора, то есть полового отбора , действующего как внутри полов, так и между ними. Высокий уровень полового диморфизма и больший размер тела у самцов обусловлен сочетанием конкуренции между самцами и селективного давления самок. [75] Виды приматов, у которых группы состоят из многих самок и одного самца, имеют более высокий половой диморфизм, чем виды, у которых есть одновременно несколько самок и самцов или одна самка и один самец. Наибольшим половым диморфизмом обладают полигинные приматы, меньшим – полигамные и моногамные. [76] [77] Снижение полигинии связано с увеличением отцовских инвестиций. [78] [79]
Демографический переход [ править ]
Демографический переход описывает современное снижение уровня рождаемости и смертности. С дарвиновской точки зрения не имеет смысла, что семьи с большими ресурсами рожают меньше детей. Одним из объяснений демографического перехода является увеличение родительских инвестиций, необходимых для воспитания детей, которые смогут поддерживать тот же уровень ресурсов, что и их родители. [80]
См. также [ править ]
- Эффект Золушки
- Стоимость воспитания ребенка
- Родственный отбор
- Дезертирство помощника
- Родительская забота у птиц
- Преждевременность и альтрициальность
- теория выбора r/K
Ссылки [ править ]
- ^ Клаттон-Брок, TH 1991. Эволюция родительской заботы . Принстон, Нью-Джерси: Princeton U. Press. стр. 9
- ^ Jump up to: Перейти обратно: а б с д и Триверс, Р.Л. (1972). Родительские инвестиции и половой отбор. В Б. Кэмпбелле (ред.), Половой отбор и происхождение человека , 1871–1971 (стр. 136–179). Чикаго, Иллинойс: Олдин. ISBN 0-435-62157-2 .
- ^ Дарвин, Чарльз (2009), «Публикация о «происхождении видов» - 3 октября 1859 г. – 31 декабря 1859 г.», в книге Дарвина Фрэнсиса (редактор), « Жизнь и письма Чарльза Дарвина» , Кембриджский университет. Press, стр. 205–255, doi : 10.1017/cbo9780511702891.007 , ISBN. 9780511702891
- ^ Фишер, Рональд Эйлмер (1930). Генетическая теория естественного отбора . Оксфорд: Кларендон Пресс. дои : 10.5962/bhl.title.27468 .
- ^ Бейтман, AJ (декабрь 1948 г.). «Внутриполовой отбор у дрозофилы» . Наследственность . 2 (3): 349–368. дои : 10.1038/hdy.1948.21 . ISSN 0018-067X . ПМИД 18103134 .
- ^ ТРИВЕРС, РОБЕРТ Л. (февраль 1974 г.). «Конфликт родителей и детей» . Американский зоолог . 14 (1): 249–264. дои : 10.1093/icb/14.1.249 . ISSN 0003-1569 .
- ^ Эдвардс, AWF (1 апреля 2000 г.). «Генетическая теория естественного отбора» . Генетика . 154 (4): 1419–1426. дои : 10.1093/генетика/154.4.1419 . ПМК 1461012 . PMID 10747041 – через www.genetics.org.
- ^ Берк, Т.; Дэвиест, Северная Каролина; Бруфорд, Миссури; Хэтчвелл, Би Джей (1989). «Родительская забота и брачное поведение полиандрических зайцев Prunellamodularis, связанных с отцовством с помощью дактилоскопии ДНК». Природа . 338 (6212): 249–51. Бибкод : 1989Natur.338..249B . дои : 10.1038/338249a0 . S2CID 4333938 .
- ^ Сантос, РС (1995). «Аллопатеринская забота о красногубой собачке». Журнал биологии рыб . 47 (2): 350–353. Бибкод : 1995JFBio..47..350S . дои : 10.1111/j.1095-8649.1995.tb01904.x .
- ^ Келли, Кейтлин А.; Норбутус, Аманда Дж.; Лагаланте, Энтони Ф.; Айенгар, Викрам К. (2012). «Мужские брачные феромоны как индикаторы генетического качества арктиидной бабочки ( Utetheisa ornatrix )» . Поведенческая экология . 23 (5): 1009–14. дои : 10.1093/beheco/ars064 .
- ^ Шюрх, Роджер, Роджер и Таборски, Барбара (2005). «Функциональное значение буккального питания у ротовой цихлиды Tropheus moorii ». Поведение . 142 (3): 265–281. дои : 10.1163/1568539053778274 . JSTOR 4536244 .
- ^ Дэвис, Николас Б.; Джон Р. Кребс и Стюарт А. Уэст (2012). Введение в поведенческую экологию . Уайли-Блэквелл. стр. 367–371.
- ^ Баретт Л., Данбар Р. и Лисетт Дж. (2002). Эволюционная психология человека. Пэлгрейв Пресс.
- ^ Jump up to: Перейти обратно: а б с Олссон, Олоф (1997). «Отказ от кладки: решение у королевских пингвинов, зависящее от штата». Журнал птичьей биологии . 28 (3): 264–267. дои : 10.2307/3676979 . JSTOR 3676979 .
- ^ Jump up to: Перейти обратно: а б Лосось, Кэтрин А.; Шекелфорд, Тодд К. (сентябрь 2007 г.). Семейные отношения: эволюционный взгляд . Оксфордская стипендия онлайн. ISBN 9780195320510 .
- ^ Шепер-Хьюз, Нэнси (декабрь 1985 г.). «Культура, дефицит и материнское мышление: материнская отстраненность и выживание младенцев в бразильских трущобах». Этос . 13 (4): 291–317. дои : 10.1525/eth.1985.13.4.02a00010 . ISSN 0091-2131 .
- ^ Jump up to: Перейти обратно: а б Соломон, Нэнси Г. и Лорен Д. Хейс. «Биологические основы аллородительского поведения млекопитающих». Замещающие родители: биологические и социальные перспективы алло-родительского воспитания в человеческих обществах , под редакцией Джиллиан Бентли и Рут Мейс, NED – новое издание, 1-е изд., Berghahn Books, 2009, стр. 13–49. JSTOR , www.jstor.org/stable/j.ctt9qch9m.7.
- ^ Хрди, Сара Блаффер. «ЗАЧЕМ НУЖНА ДЕРЕВНЯ». «Матери и другие» , издательство Гарвардского университета, Кембридж, Массачусетс; Лондон, Англия, 2009 г., стр. 65–110. JSTOR , www.jstor.org/stable/j.ctt1c84czb.5.
- ^ Морган, CL (1894). Введение в сравнительную психологию . Лондон: В. Скотт.
- ^ Эпштейн, Р. (1984). Принцип бережливости и некоторые приложения в психологии . Журнал разума и поведения, 5
- ^ Jump up to: Перейти обратно: а б Триверс, Р. (1972). Родительские инвестиции и половой отбор. Половой отбор и происхождение человека, Альдин де Грюйтер, Нью-Йорк, 136–179.
- ^ Jump up to: Перейти обратно: а б с Басс, ДМ; Ларсен, Р.Дж.; Вестен, Д.; Семмельрот, Дж. (1992). «Половые различия в ревности: эволюция, физиология и психология» (PDF) . Психологическая наука . 3 (4): 251–255. дои : 10.1111/j.1467-9280.1992.tb00038.x . S2CID 27388562 .
- ^ Кессель, Э.Л. (1955). «Спаривание воздушных шаров». Систематическая зоология . 4 (3): 97–104. дои : 10.2307/2411862 . JSTOR 2411862 .
- ^ Рояма, Т (1966). «Новая интерпретация ухаживающего кормления». Исследование птиц . 13 (2): 116–129. Бибкод : 1966BirdS..13..116R . дои : 10.1080/00063656609476115 .
- ^ Стоукс, AW, и Уильямс, HW (1971). Брачное питание куриных птиц. Аук, 543-559. Чикаго.
- ^ Триверс, Р. (1985). Социальная эволюция. Менло-Парк, Калифорния: Бенджамин/Каммингс.
- ^ Отсутствие, DL (1968). Экологические приспособления для размножения птиц.
- ^ Басс, DM (1989). «Половые различия в предпочтениях человека к партнеру: эволюционные гипотезы, проверенные в 37 культурах» . Поведенческие и мозговые науки . 12 (1): 1–14. дои : 10.1017/s0140525x00023992 . S2CID 3807679 .
- ^ Кларк, РД; Хэтфилд, Э. (1989). «Гендерные различия в восприимчивости к сексуальным предложениям» (PDF) . Журнал психологии и сексуальности человека . 2 (1): 39–55. дои : 10.1300/j056v02n01_04 .
- ^ Армстронг, Э.А.; Гамильтон, Лейтенант; Армстронг, EM; Сили, Дж.Л. (2014). « Пол, социальный класс и дискурс «хороших девочек» в кампусе» (PDF) . Социальная психология Ежеквартально . 77 (2): 100–122. дои : 10.1177/0190272514521220 . S2CID 12935534 .
- ^ Торнхилл, Р., и Палмер, Коннектикут (2001). Естественная история изнасилования: Биологические основы сексуального принуждения. Пресс-центр МТИ.
- ^ «Дети и подростки: Статистика – ДОЖДЬ» .
- ^ «Статистика изнасилований» .
- ^ Jump up to: Перейти обратно: а б Басс, ДМ; Шмитт, Д.П. (1993). «Теория сексуальных стратегий: эволюционный взгляд на спаривание людей» (PDF) . Психологический обзор . 100 (2): 204–32. дои : 10.1037/0033-295x.100.2.204 . ПМИД 8483982 .
- ^ Jump up to: Перейти обратно: а б с д Лопес, Энтони С.; Макдермотт, Роуз; Петерсен, Майкл Банг (2011). «Государства в уме: эволюция, коалиционная психология и международная политика». Международная безопасность . 36 (2): 48–83. дои : 10.1162/isec_a_00056 . S2CID 57562816 .
- ^ Бейтман Эй Джей (декабрь 1948 г.). «Внутриполовой отбор у дрозофилы» . Наследственность . 2 (3): 349–68. дои : 10.1038/hdy.1948.21 . ПМИД 18103134 .
- ^ Амундсен, Тронд (1 апреля 2000 г.). «Почему самок птиц украшают». Тенденции в экологии и эволюции . 15 (4): 149–55. дои : 10.1016/S0169-5347(99)01800-5 . ПМИД 10717684 .
- ^ Jump up to: Перейти обратно: а б Розенберг, Карен; Треватан, Венда (ноябрь 2002 г.). «Рождение, акушерство и эволюция человека». BJOG: Международный журнал акушерства и гинекологии . 109 (11): 1199–1206. дои : 10.1046/j.1471-0528.2002.00010.x . ISSN 1470-0328 . ПМИД 12452455 . S2CID 35070435 .
- ^ Гарратт, М.; Гайяр, Ж.-М.; Брукс, Колорадо; Леметр, Ж.-Ф. (22 апреля 2013 г.). «Диверсификация плаценты связана с изменением темпа жизни» . Труды Национальной академии наук . 110 (19): 7760–7765. Бибкод : 2013PNAS..110.7760G . дои : 10.1073/pnas.1305018110 . ISSN 0027-8424 . ПМЦ 3651450 . ПМИД 23610401 .
- ^ Билби, Дж.; Мейс, генеральный директор; Бининда-Эмондс, ORP; Кардилло, М.; Гиттлман, Дж.Л.; Джонс, Кентукки; Орм, CDL; Первис, А. (июнь 2007 г.). «Континуум быстрого и медленного развития в истории жизни млекопитающих: эмпирическая переоценка». Американский натуралист . 169 (6): 748–757. дои : 10.1086/516847 . ISSN 0003-0147 . ПМИД 17479461 . S2CID 18663841 .
- ^ Соареш, Майкл Дж; Варберг, Каела М; Икбал, Хуршид (22 февраля 2018 г.). «Гемохориальная плацентация: развитие, функции и адаптации †» . Биология размножения . 99 (1): 196–211. дои : 10.1093/biolre/ioy049 . ISSN 0006-3363 . ПМК 6044390 . ПМИД 29481584 .
- ^ Махмуд Ф. Фаталла (2013). «Как эволюция человеческого мозга повлияла на сексуальное и репродуктивное здоровье женщин». Понимание репродуктивной биологии . 6 : 11–18. дои : 10.4137/rbi.s12217 . ISSN 1178-6426 .
- ^ Венда., Треватан (2011). Человеческое рождение: эволюционная перспектива . Нью-Брансуик: Издатели транзакций. ISBN 9781412815024 . OCLC 669122326 .
- ^ Уэллс, Джонатан К.К.; ДеСильва, Джереми М.; Сток, Джей Т. (2012). «Акушерская дилемма: древняя игра в русскую рулетку или изменчивая дилемма, чувствительная к экологии?» . Американский журнал физической антропологии . 149 (С55): 40–71. дои : 10.1002/ajpa.22160 . ISSN 0002-9483 . ПМИД 23138755 .
- ^ Треватан, Венда Р. (июнь 1996 г.). «Эволюция прямохождения и вспомогательных родов». Медицинская антропология Ежеквартальный журнал . 10 (2): 287–290. дои : 10.1525/maq.1996.10.2.02a00100 . ISSN 0745-5194 . ПМИД 8744088 .
- ^ Гинтис, Герберт (ноябрь 2012 г.). «Битва Титанов» . Бионаука . 62 (11): 987–991. дои : 10.1525/bio.2012.62.11.8 . ISSN 1525-3244 .
- ^ Демуру, Элиза; Феррари, Пьер Франческо; Палаги, Элизабетта (сентябрь 2018 г.). «Является ли присутствие родов уникальной особенностью человека? Новые данные свидетельствуют о том, что самки бонобо защищают и поддерживают рожениц». Эволюция и поведение человека . 39 (5): 502–510. doi : 10.1016/j.evolhumbehav.2018.05.003 . ISSN 1090-5138 . S2CID 149489276 .
- ^ Дуглас, Памела Хайди (10 июля 2014 г.). «Общественность самки во время дневного рождения дикого бонобо в Луикотале, Демократическая Республика Конго». Приматы . 55 (4): 533–542. дои : 10.1007/s10329-014-0436-0 . ISSN 0032-8332 . ПМИД 25007717 . S2CID 18719002 .
- ^ Уокер, Маргарет Л.; Херндон, Джеймс Г. (1 сентября 2008 г.). «Менопауза у нечеловеческих приматов? 1» . Биология размножения . 79 (3): 398–406. дои : 10.1095/biolreprod.108.068536 . ISSN 0006-3363 . ПМК 2553520 . ПМИД 18495681 .
- ^ Фокс, Молли; Джохау, Йоханнес; Кнапп, Лесли А. (2011). «Эгоистичный ген бабушки: роль Х-хромосомы и неопределенности отцовства в эволюции бабушкиного поведения и долголетия» . Международный журнал эволюционной биологии . 2011 : 165919. дои : 10.4061/2011/165919 . ISSN 2090-052X . ПМК 3118636 . ПМИД 21716697 .
- ^ Таттерсолл, Ян (10 августа 2010 г.). «Палеоантропология: два новых предложения» . Эволюция: образование и информационно-пропагандистская деятельность . 3 (3): 464–465. дои : 10.1007/s12052-010-0263-8 . ISSN 1936-6426 .
- ^ Макланахан, СС (1999). Отсутствие отца и благополучие детей . Как справиться с разводом, воспитанием детей-одиночек и повторным браком: взгляд на риск и устойчивость, 117-145.
- ^ Кирнан, Кентукки; Хобкрафт, Дж. (1997). «Развод родителей в детстве: возраст первого полового акта, партнерство и родительство» (PDF) . Исследования народонаселения . 51 (1): 41–55. дои : 10.1080/0032472031000149716 .
- ^ Эллис, Би Джей; Бейтс, Дж. Э.; Додж, Калифорния; Фергюссон, DM; Джон Хорвуд, Л.; Петтит, Г.С.; Вудворд, Л. (2003). «Подвергает ли отсутствие отца дочери особому риску ранней сексуальной активности и подростковой беременности?» . Развитие ребенка . 74 (3): 801–821. дои : 10.1111/1467-8624.00569 . ПМЦ 2764264 . ПМИД 12795391 .
- ^ Скарамелла, Л.В.; Конгер, РД; Саймонс, РЛ; Уитбек, Л.Б. (1998). «Прогнозирование риска беременности в позднем подростковом возрасте: социальная контекстуальная перспектива» (PDF) . Психология развития . 34 (6): 1233–45. дои : 10.1037/0012-1649.34.6.1233 . ПМИД 9823508 .
- ^ Бельский, Дж.; Стейнберг, Л.; Дрейпер, П. (1991). «Детский опыт, межличностное развитие и репродуктивная стратегия: эволюционная теория социализации». Развитие ребенка . 62 (4): 647–670. дои : 10.2307/1131166 . JSTOR 1131166 . ПМИД 1935336 .
- ^ Коли, РЛ; Чейз-Лансдейл, Польша (1998). «Подростковая беременность и родительство: последние данные и будущие направления» (PDF) . Американский психолог . 53 (2): 152–66. дои : 10.1037/0003-066x.53.2.152 . ПМИД 9491745 . S2CID 1814668 . Архивировано из оригинала (PDF) 02 марта 2019 г.
- ^ Антфолк, Дж.; Сьёлунд, А. (2018). «Высокие родительские инвестиции в детство связаны с повышением ценности партнера во взрослом возрасте» (PDF) . Личность и индивидуальные различия . 127 (1): 144–150. дои : 10.1016/j.paid.2018.02.004 . S2CID 141063266 .
- ^ Линн, Д.Б.; Сори, WL (1959). «Влияние отсутствия отца на норвежских мальчиков и девочек». Журнал аномальной и социальной психологии . 59 (2): 258–62. дои : 10.1037/h0040784 . ПМИД 14419160 .
- ^ Маламут, Нью-Мексико (1981). «Склонность к изнасилованию среди мужчин» (PDF) . Журнал социальных проблем . 37 (4): 138–157. дои : 10.1111/j.1540-4560.1981.tb01075.x . Архивировано из оригинала (PDF) 13 декабря 2012 г.
- ^ Чисхолм, Дж.С.; Куинливан, Дж.А.; Петерсен, RW; Коул, Д.А. (2005). «Ранний стресс предсказывает возраст менархе и первых родов, привязанность взрослого человека и ожидаемую продолжительность жизни» (PDF) . Человеческая природа . 16 (3): 233–265. дои : 10.1007/s12110-005-1009-0 . ПМИД 26189749 . S2CID 207392022 .
- ^ Силлен-Туллберг, Биргитта; Моллер, Андерс П. (январь 1993 г.). «Взаимосвязь между скрытой овуляцией и системами спаривания у антропоидных приматов: филогенетический анализ». Американский натуралист . 141 (1): 1–25. дои : 10.1086/285458 . ISSN 0003-0147 . ПМИД 19426020 . S2CID 2120481 .
- ^ БЕНШУФ, Л; ТОРНХИЛЛ, Р. (апрель 1979 г.). «Эволюция моногамии и скрытой овуляции у человека». Журнал социальных и биологических систем . 2 (2): 95–106. дои : 10.1016/0140-1750(79)90001-0 . ISSN 0140-1750 .
- ^ Кинси, Альфред К.; Помрой, Уорделл Р.; Мартин, Клайд Э. (июнь 2003 г.). «Сексуальное поведение мужчины» . Американский журнал общественного здравоохранения . 93 (6): 894–898. дои : 10.2105/ajph.93.6.894 . ISSN 0090-0036 . ПМЦ 1643237 . ПМИД 12773346 .
- ^ Гангестад, Юго-Запад; Симпсон, Дж.А.; ДиДжеронимо, К.; Бик, М. (1992). «Дифференциальная точность восприятия человека по различным чертам: исследование функциональной гипотезы» (PDF) . Журнал личности и социальной психологии . 62 (4): 688–98. дои : 10.1037/0022-3514.62.4.688 . ПМИД 1583592 .
- ^ Симпсон, Дж.А.; Гангестад, Юго-Запад (1991). «Индивидуальные различия в социосексуальности: доказательства конвергентной и дискриминантной достоверности». Журнал личности и социальной психологии . 60 (6): 870–83. дои : 10.1037/0022-3514.60.6.870 . ПМИД 1865325 .
- ^ Jump up to: Перейти обратно: а б Хольцман, Николас С.; Струбе, Майкл Дж. (18 июля 2013 г.). «Помимо краткосрочного спаривания, долгосрочное спаривание однозначно связано с человеческой личностью» . Эволюционная психология . 11 (5): 1101–29. дои : 10.1177/147470491301100514 . ISSN 1474-7049 . ПМИД 24342881 .
- ^ Джексон, Джени Джеймс; Киркпатрик, Ли А. (ноябрь 2007 г.). «Структура и измерение человеческих стратегий спаривания: к многомерной модели социосексуальности». Эволюция и поведение человека . 28 (6): 382–391. doi : 10.1016/j.evolhumbehav.2007.04.005 . ISSN 1090-5138 .
- ^ Уэйд, MJ; Шустер, С.М. (2002). «Эволюция родительской заботы в контексте полового отбора: критическая переоценка теории родительских инвестиций». Американский натуралист . 160 (3): 285–292. дои : 10.1086/341520 . ПМИД 18707439 . S2CID 8929417 .
- ^ Басс, ДМ; Шмитт, Д.П. (1993). «Теория сексуальных стратегий: эволюционный взгляд на спаривание людей» (PDF) . Психологический обзор . 100 (2): 204–232. дои : 10.1037/0033-295x.100.2.204 . ПМИД 8483982 .
- ^ Шейб, JE (1997, июнь). Критерии выбора партнера, зависящие от контекста: женские компромиссы в контексте долгосрочных и внепарных отношений . Документ, представленный на ежегодном собрании Общества человеческого поведения и эволюции, Университет Аризоны, Тусон, Аризона.
- ^ Басс, доктор медицины (1999). Эволюционная психология: новая наука о разуме. (2-е изд.). США: Pearson Education, Inc.
- ^ Дэвид, МБ; Барнс, М. (1986). «Предпочтения в выборе партнера для человека» (PDF) . Журнал личности и социальной психологии . 50 (3): 559–570. дои : 10.1037/0022-3514.50.3.559 .
- ^ Басс, DM (1989). «Половые различия в предпочтениях человека к партнеру: проверка эволюционных гипотез в 37 культурах» . Поведенческие и мозговые науки . 12 :1–49. дои : 10.1017/s0140525x00023992 . S2CID 3807679 .
- ^ Ланде, Рассел (март 1980 г.). «Половой диморфизм, половой отбор и адаптация полигенных признаков». Эволюция . 34 (2): 292–305. дои : 10.2307/2407393 . ISSN 0014-3820 . JSTOR 2407393 . ПМИД 28563426 .
- ^ Чеверуд, Джеймс М.; Доу, Малкольм М.; Лейтенеггер, Уолтер (ноябрь 1985 г.). «Количественная оценка филогенетических ограничений в сравнительном анализе: половой диморфизм массы тела среди приматов». Эволюция . 39 (6): 1335–1351. дои : 10.1111/j.1558-5646.1985.tb05699.x . ISSN 0014-3820 . ПМИД 28564267 . S2CID 27531072 .
- ^ Лейтенеггер, Уолтер; Келли, Джеймс Т. (январь 1977 г.). «Связь полового диморфизма размеров собак и размеров тела с социальными, поведенческими и экологическими коррелятами у человекообразных приматов». Приматы . 18 (1): 117–136. дои : 10.1007/bf02382954 . ISSN 0032-8332 . S2CID 36131958 .
- ^ Лейтенеггер, Уолтер; Чеверуд, Джеймс (декабрь 1982 г.). «Корреляты полового диморфизма у приматов: экологические и размерные переменные». Международный журнал приматологии . 3 (4): 387–402. CiteSeerX 10.1.1.536.6939 . дои : 10.1007/bf02693740 . ISSN 0164-0291 . S2CID 38220186 .
- ^ Раллс, Кэтрин (сентябрь 1977 г.). «Половой диморфизм у млекопитающих: птичьи модели и вопросы без ответа». Американский натуралист . 111 (981): 917–938. дои : 10.1086/283223 . ISSN 0003-0147 . S2CID 38569769 .
- ^ Боргерхофф Малдер, Моник (июль 1998 г.). «Демографический переход: приблизились ли мы к эволюционному объяснению?». Тенденции в экологии и эволюции . 13 (7): 266–270. дои : 10.1016/s0169-5347(98)01357-3 . ISSN 0169-5347 . ПМИД 21238295 .
Дальнейшее чтение [ править ]
- Клаттон-Брок, TH и К. Годфрей. 1991. «Родительские инвестиции», в «Поведенческой экологии: эволюционный подход» . Под редакцией Дж. Р. Кребса и Н. Б. Дэвиса, стр. 234–262. Бостон: Блэквелл.
- Басс, Дэвид М. (март 1989 г.). «Половые различия в предпочтениях человека к партнеру: эволюционные гипотезы, проверенные в 37 культурах» . Поведенческие и мозговые науки . 12 (1): 1–14. дои : 10.1017/S0140525X00023992 .
- Бельский Дж., Стейнберг Л., Дрейпер П. (август 1991 г.). «Детский опыт, межличностное развитие и репродуктивная стратегия: и эволюционная теория социализации». Развитие ребенка . 62 (4): 647–70. дои : 10.1111/j.1467-8624.1991.tb01558.x . ПМИД 1935336 .
- Вудворд, Кевин; Ричардс, Мириам Х. (2005). «Модель родительских инвестиций и минимальные критерии выбора партнера у человека» . Поведенческая экология . 16 (1): 57–61. дои : 10.1093/beheco/arh121 .
- Гири, округ Колумбия (2005). Эволюция отцовских инвестиций. В Д. М. Бассе (ред.), Справочнике по эволюционной психологии (стр. 483–505). Хобокен, Нью-Джерси: John Wiley & Sons. Полный текст. Архивировано 20 декабря 2019 г. на Wayback Machine.