Введение в эволюцию
| Часть серии о |
| Эволюционная биология |
|---|
 |
В биологии происходит эволюция — это процесс изменения всех форм жизни на протяжении поколений, а эволюционная биология — это изучение того, как эволюция . Биологические популяции развиваются посредством генетических изменений, которые соответствуют изменениям в организмов характеристиках наблюдаемых . К генетическим изменениям относятся мутации , вызванные повреждением или ошибками репликации ДНК организмов . Поскольку генетическая изменчивость популяции беспорядочно меняется из поколения в поколение, естественный отбор постепенно приводит к тому, что признаки становятся более или менее распространенными в зависимости от относительного репродуктивного успеха организмов с этими признаками.
Возраст Земли составляет около 4,5 миллиардов лет. [1] [2] [3] Самые ранние неоспоримые свидетельства существования жизни на Земле датируются как минимум 3,5 миллиарда лет назад. [4] [5] [6] Эволюция не пытается объяснить происхождение жизни (вместо этого она охватывает абиогенез ), но она объясняет, как ранние формы жизни превратились в сложную экосистему, которую мы видим сегодня. [7] На основании сходства между всеми современными организмами предполагается, что вся жизнь на Земле возникла в результате общего происхождения от последнего универсального предка , от которого все известные виды разошлись в процессе эволюции. [8]
Все особи имеют наследственный материал в виде генов, полученных от родителей, который они передают любому потомству. Среди потомства существуют вариации генов из-за введения новых генов посредством случайных изменений, называемых мутациями, или перетасовки существующих генов во время полового размножения . [9] [10] Потомство отличается от родителя незначительными случайными способами. Если эти различия полезны, потомство с большей вероятностью выживет и размножится. Это означает, что большее количество потомков в следующем поколении будет иметь такую полезную разницу, и у особей не будет равных шансов на репродуктивный успех. Таким образом, черты, которые приводят к лучшей адаптации организмов к условиям жизни, становятся более распространенными в потомственных популяциях. [9] [10] Эти различия накапливаются, что приводит к изменениям в популяции. Этот процесс ответственен за множество разнообразных форм жизни в мире.
Современное понимание эволюции началось с публикации в 1859 году Чарльза Дарвина книги «Происхождение видов» . Кроме того, работа Грегора Менделя с растениями помогла объяснить наследственные закономерности генетики . [11] Открытия окаменелостей в палеонтологии , достижения в области популяционной генетики и глобальная сеть научных исследований предоставили дополнительную информацию о механизмах эволюции. Теперь ученые хорошо понимают происхождение новых видов ( видообразование ) и наблюдали процесс видообразования в лаборатории и в дикой природе. Эволюция — это основная научная теория , которую биологи используют для понимания жизни и которая используется во многих дисциплинах, включая медицину , психологию , биологию сохранения , антропологию , судебно-медицинскую экспертизу , сельское хозяйство и другие социально-культурные приложения.
Простой обзор [ править ]
Основные идеи эволюции можно резюмировать следующим образом:
- жизни Формы размножаются и поэтому имеют тенденцию становиться более многочисленными.
- Такие факторы, как хищничество и конкуренция, препятствуют выживанию особей.
- Каждый потомок отличается от своего родителя (родителей) незначительными случайными способами.
- Если эти различия полезны, потомство с большей вероятностью выживет и размножится.
- Это делает вероятным, что большее количество потомков в следующем поколении будет иметь полезные различия и меньшее количество будет иметь вредные различия.
- Эти различия накапливаются на протяжении поколений, что приводит к изменениям внутри популяции.
- Со временем популяции могут разделиться или разветвиться на новые виды.
- Эти процессы, известные под общим названием «эволюция», ответственны за множество разнообразных форм жизни, наблюдаемых в мире.
Естественный отбор [ править ]

В XIX веке естествознания были популярны коллекции и музеи . Европейская экспансия и военно-морские экспедиции привлекали натуралистов , а кураторы великих музеев демонстрировали сохранившиеся и живые образцы разновидностей жизни. Чарльз Дарвин был английским выпускником, получившим образование и обученным дисциплинам естествознания. Такие естествоиспытатели будут собирать, каталогизировать, описывать и изучать огромные коллекции образцов, хранящиеся и управляемые кураторами этих музеев. Дарвин служил корабельным натуралистом на борту HMS Beagle , отправленного в пятилетнюю исследовательскую экспедицию по всему миру. Во время своего путешествия он наблюдал и собирал множество организмов, очень интересуясь разнообразными формами жизни вдоль побережий Южной Америки и соседних Галапагосских островов . [12] [13]

Дарвин приобрел обширный опыт, собирая и изучая естественную историю форм жизни из отдаленных мест. Благодаря своим исследованиям он сформулировал идею о том, что каждый вид развился от предков со схожими характеристиками. В 1838 году он описал, как это произойдет благодаря процессу, который он назвал естественным отбором. [14]
Размер популяции зависит от того, сколько и сколько ресурсов способны ее поддержать. Чтобы численность населения оставалась неизменной из года в год, должно существовать равновесие или баланс между численностью населения и доступными ресурсами. Поскольку организмы производят больше потомства, чем может поддержать их окружающая среда, не все особи могут выжить в каждом поколении. Должна быть конкурентная борьба за ресурсы, способствующие выживанию. В результате Дарвин понял, что выживание определяется не только случайностью. Вместо этого выживание организма зависит от различий каждого отдельного организма или «особенностей», которые помогают или препятствуют выживанию и воспроизводству. Хорошо адаптированные особи, вероятно, оставят больше потомства, чем их менее адаптированные конкуренты. Черты, препятствующие выживанию и воспроизводству, исчезнут с течением поколений. Черты, которые помогают организму выживать и размножаться, накапливаются в течение поколений. Дарвин понял, что неравная способность особей к выживанию и размножению может вызвать постепенные изменения в популяции, и использовал термин естественный отбор для описания этого процесса. [15] [16]
Наблюдения за изменчивостью животных и растений легли в основу теории естественного отбора. Например, Дарвин заметил, что орхидеи и насекомые имеют тесную связь, позволяющую опылять растения. что орхидеи имеют разнообразные структуры, привлекающие насекомых, поэтому пыльца цветов Он отметил , прилипает к телам насекомых. Таким способом насекомые переносят пыльцу от мужской орхидеи к женской. Несмотря на сложный внешний вид орхидей, эти специализированные части состоят из тех же основных структур, что и другие цветы. В своей книге «Оплодотворение орхидей» (1862 г.) Дарвин предположил, что цветы орхидей были адаптированы из ранее существовавших частей посредством естественного отбора. [17]
Дарвин все еще исследовал и экспериментировал со своими идеями о естественном отборе, когда получил письмо от Альфреда Рассела Уоллеса, в котором описывалась теория, очень похожая на его собственную. Это привело к немедленной совместной публикации обеих теорий. И Уоллес, и Дарвин рассматривали историю жизни как генеалогическое древо , где каждая развилка ветвей дерева являлась общим предком. Кончики конечностей представляли современные виды, а ветви — общих предков, которые были общими для многих различных видов. Чтобы объяснить эти отношения, Дарвин сказал, что все живые существа связаны между собой, а это означало, что вся жизнь должна произойти от нескольких форм или даже от одного общего предка. Он назвал этот процесс нисхождением с модификацией . [16]
Дарвин опубликовал свою теорию эволюции путем естественного отбора в книге «Происхождение видов» в 1859 году. [18] Его теория означает, что вся жизнь, включая человечество , является продуктом продолжающихся природных процессов. Утверждение о том, что вся жизнь на Земле имеет общего предка, встретило возражения со стороны некоторых религиозных групп . Их возражения контрастируют с уровнем поддержки теории более чем 99 процентами представителей научного сообщества сегодня. [19]
Естественный отбор обычно приравнивают к выживанию наиболее приспособленных , но это выражение возникло в Герберта Спенсера в «Принципах биологии» 1864 году, через пять лет после того, как Чарльз Дарвин опубликовал свои оригинальные работы. Выживание наиболее приспособленных неверно описывает процесс естественного отбора, поскольку естественный отбор касается не только выживания, и не всегда выживает наиболее приспособленный. [20]
Источник вариаций [ править ]
Теория естественного отбора Дарвина заложила основу современной эволюционной теории, а его эксперименты и наблюдения показали, что организмы в популяциях различаются друг от друга, что некоторые из этих вариаций передаются по наследству и что на эти различия можно воздействовать путем естественного отбора. Однако он не смог объяснить источник этих изменений. Как и многие его предшественники, Дарвин ошибочно полагал, что наследуемые черты являются продуктом использования и неиспользования и что особенности, приобретенные в течение жизни организма, могут передаваться его потомству. Он искал примеры, такие как крупные наземные птицы, у которых ноги становятся сильнее благодаря упражнениям, а крылья становятся слабее из-за того, что они не летают до тех пор, пока, как страус , они не перестают летать вообще. [21] Это недоразумение называлось наследованием приобретенных признаков и было частью теории трансмутации видов, выдвинутой в 1809 году Жаном-Батистом Ламарком . В конце 19 века эта теория стала известна как ламаркизм . Дарвин выдвинул неудачную теорию, которую он назвал пангенезисом, чтобы попытаться объяснить, как приобретенные характеристики могут быть унаследованы. В 1880-х годах эксперименты Августа Вейсмана показали, что изменения, вызванные использованием и неиспользованием, не могут передаваться по наследству, и ламаркизм постепенно потерял популярность. [22]
Недостающую информацию, необходимую для объяснения того, как новые признаки могут передаваться от родителя к потомству, предоставила новаторская генетическая работа Грегора Менделя . Эксперименты Менделя с несколькими поколениями растений гороха продемонстрировали, что наследование работает путем разделения и перетасовки наследственной информации во время формирования половых клеток и рекомбинации этой информации во время оплодотворения. Это похоже на смешивание игральных карт в разных комбинациях , когда организм получает случайную смесь половины карт от одного родителя и половины карт от другого. Мендель назвал информационными факторами ; однако позже они стали известны как гены. Гены являются основными единицами наследственности в живых организмах. Они содержат информацию, управляющую физическим развитием и поведением организмов.
Гены состоят из ДНК . ДНК — это длинная молекула, состоящая из отдельных молекул, называемых нуклеотидами . Генетическая информация закодирована в последовательности нуклеотидов, составляющих ДНК, точно так же, как последовательность букв в словах несет информацию на странице. Гены подобны коротким инструкциям, составленным из «букв» алфавита ДНК. В совокупности весь набор этих генов дает достаточно информации, чтобы служить «инструкцией» по построению и управлению организмом. Однако инструкции, записанные в этом алфавите ДНК, могут быть изменены в результате мутаций, и это может изменить инструкции, содержащиеся в генах. Внутри клетки гены передаются в хромосомах , которые представляют собой пакеты для переноса ДНК. Именно перестановка хромосом приводит к появлению уникальных комбинаций генов у потомства. Поскольку гены взаимодействуют друг с другом в ходе развития организма, новые комбинации генов, образующиеся в результате полового размножения, могут увеличить генетическую изменчивость популяции даже без новых мутаций. [23] Генетическая изменчивость популяции также может увеличиваться, когда члены этой популяции скрещиваются с особями из другой популяции, вызывая поток генов между популяциями. Это может привести к появлению в популяции генов, которых раньше не было. [24]
Эволюция – это не случайный процесс. Хотя мутации в ДНК случайны, естественный отбор — не случайный процесс: вероятность репродуктивного успеха определяется окружающей средой. Эволюция — неизбежный результат несовершенного копирования самовоспроизводящихся организмов, размножающихся на протяжении миллиардов лет под избирательным давлением окружающей среды. Результатом эволюции не является идеально сконструированный организм. Конечными продуктами естественного отбора являются организмы, адаптированные к нынешней среде обитания. Естественный отбор не предполагает продвижения к конечной цели . Эволюция не стремится к созданию более продвинутых , более разумных или более сложных форм жизни. [25] Например, блохи (бескрылые паразиты) произошли от крылатого предка скорпиона , а змеи — это ящерицы , которым больше не нужны конечности, хотя у питонов все еще растут крошечные структуры, которые являются остатками задних ног их предков. [26] [27] Организмы — это всего лишь результат вариаций, которые достигают успеха или терпят неудачу в зависимости от условий окружающей среды в данный момент.
Быстрые изменения окружающей среды обычно приводят к вымиранию видов. [28] Из всех видов, существовавших на Земле, 99,9 процентов сейчас вымерли. [29] С момента зарождения жизни на Земле пять крупных массовых вымираний привели к значительному и внезапному сокращению разнообразия видов. Самое последнее, мел-палеогеновое вымирание , произошло 66 миллионов лет назад. [30]
Генетический дрейф [ править ]
Генетический дрейф является причиной изменения частоты аллелей в популяциях вида. Аллели — это различные варианты определенных генов. Они определяют такие вещи, как цвет волос , оттенок кожи , цвет глаз и группу крови ; другими словами, все генетические черты, которые различаются у разных людей. Генетический дрейф не приводит к появлению новых аллелей в популяции, но может уменьшить изменчивость внутри популяции за счет удаления аллеля из генофонда. Генетический дрейф вызван случайной выборкой аллелей. По-настоящему случайная выборка — это выборка, в которой никакие внешние силы не влияют на то, что выбрано. Это все равно что вытаскивать из коричневого бумажного пакета шарики одинакового размера и веса, но разного цвета. В любом потомстве присутствующие аллели являются образцами аллелей предыдущих поколений, и случайность играет роль в том, выживет ли особь для воспроизводства и передачи образца своего поколения следующему. Частота аллелей в популяции — это соотношение копий одной конкретной аллели, имеющих одну и ту же форму, к числу всех форм аллели, присутствующих в популяции. [31]
Генетический дрейф затрагивает меньшие популяции больше, чем большие популяции. [32]
- Принцип Вайнберга Харди
Принцип Харди-Вайнберга гласит, что при определенных идеализированных условиях, включая отсутствие давления отбора, в большой популяции частота аллелей не изменится по мере смены поколений. [33] Говорят, что популяция, удовлетворяющая этим условиям, находится в равновесии Харди – Вайнберга. В частности, Харди и Вайнберг показали, что доминантные и рецессивные аллели не имеют тенденции автоматически становиться более или менее частыми соответственно, как считалось ранее.
Условия равновесия Харди-Вайнберга включают отсутствие мутаций, иммиграции или эмиграции , которые могут напрямую изменять частоты аллелей. Кроме того, спаривание должно быть совершенно случайным, при этом все самцы (или в некоторых случаях самки) являются одинаково желательными партнерами. Это обеспечивает истинно случайное смешивание аллелей. [34] Популяция, находящаяся в равновесии Харди – Вайнберга, аналогична колоде карт ; сколько бы раз не перетасовывалась колода, новые карты не добавляются и старые не забираются. Карты в колоде представляют аллели генофонда популяции.
На практике ни одна популяция не может находиться в идеальном равновесии Харди-Вайнберга. Конечный размер популяции в сочетании с естественным отбором и многими другими эффектами приводят к изменению частот аллелей с течением времени.
в численности населения Узкое место

возникает Узкое место в популяции , когда популяция вида резко сокращается за короткий период времени из-за внешних сил. [35] В условиях настоящего узкого места в популяции сокращение не благоприятствует какой-либо комбинации аллелей; то, что люди выживают, является абсолютно случайным шансом. Узкое место может уменьшить или устранить генетическую изменчивость в популяции. популяции Дальнейшие события дрейфа после события «узкого места» также могут уменьшить генетическое разнообразие . Отсутствие созданного разнообразия может подвергнуть население риску других видов селективного давления. [36]
Типичным примером «узкого места» популяции является северный морской слон . Из-за чрезмерной охоты на протяжении XIX века популяция северного морского слона сократилась до 30 особей или меньше. Они полностью выздоровели, общее число особей достигло около 100 000 и продолжает расти. Однако последствия узкого места очевидны. У тюленей чаще возникают серьезные проблемы с болезнями или генетическими нарушениями, поскольку разнообразие в популяции практически отсутствует. [37]
Эффект Основателя [ править ]
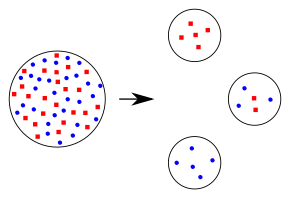
Эффект основателя возникает, когда небольшая группа из одной популяции отделяется и образует новую популяцию, часто из-за географической изоляции. Частота аллелей этой новой популяции, вероятно, отличается от частоты исходной популяции и изменит степень распространенности определенных аллелей в популяциях. Основатели популяции будут определять генетический состав и, возможно, выживание новой популяции на протяжении нескольких поколений. [34]
Одним из примеров эффекта основателя является миграция амишей в Пенсильванию в 1744 году. Двое основателей колонии в Пенсильвании несли рецессивный аллель синдрома Эллиса-ван Кревельда . Поскольку амиши, как правило, являются религиозными изолированными людьми, они скрещиваются между собой, и на протяжении поколений этой практики частота синдрома Эллиса-ван Кревельда у амишей намного выше, чем частота синдрома Эллиса-ван Кревельда среди населения в целом. [38]
Современный синтез [ править ]
Современный эволюционный синтез основан на представлении о том, что популяции организмов обладают значительной генетической изменчивостью, вызванной мутациями и рекомбинацией генов при половом размножении. Он определяет эволюцию как изменение частот аллелей внутри популяции, вызванное генетическим дрейфом, потоком генов между субпопуляциями и естественным отбором. Естественный отбор подчеркивается как важнейший механизм эволюции; большие изменения являются результатом постепенного накопления мелких изменений в течение длительных периодов времени. [39] [40]
Современный эволюционный синтез является результатом слияния нескольких различных научных областей с целью создания более связного понимания эволюционной теории. В 1920-х годах Рональд Фишер , Дж.Б.С. Холдейн и Сьюэлл Райт объединили теорию естественного отбора Дарвина со статистическими моделями менделевской генетики , основав дисциплину популяционной генетики. В 1930-х и 1940-х годах были предприняты попытки объединить популяционную генетику, наблюдения полевых натуралистов за распространением видов и подвидов, а также анализ летописи окаменелостей в единую объяснительную модель. [41] Применение принципов генетики к естественным популяциям такими учеными, как Феодосий Добжанский и Эрнст Майр , продвинуло понимание процессов эволюции. Работа Добжанского 1937 года «Генетика и происхождение видов» помогла преодолеть разрыв между генетикой и полевой биологией , представив математические работы популяционных генетиков в форме, более полезной для полевых биологов, и показав, что дикие популяции обладают гораздо большей генетической изменчивостью при географически изолированных территориях. подвиды и резервуары генетического разнообразия в рецессивных генах, чем предполагали модели ранних популяционных генетиков. Майр на основе понимания генов и прямых наблюдений за эволюционными процессами в ходе полевых исследований ввел концепцию биологического вида, которая определяла вид как группу скрещивающихся или потенциально скрещивающихся популяций, репродуктивно изолированных от всех других популяций. И Добжанский, и Майр подчеркивали важность подвидов, репродуктивно изолированных географическими барьерами, для появления новых видов. Палеонтолог Джордж Гейлорд Симпсон помог объединить палеонтологию со статистическим анализом летописи окаменелостей, который показал закономерность, соответствующую ветвящемуся и ненаправленному пути эволюции организмов, предсказанному современным синтезом. [39]
эволюции Доказательства

Научные доказательства эволюции основаны на многих аспектах биологии и включают окаменелости , гомологичные структуры и молекулярное сходство между ДНК видов.
Ископаемая запись [ править ]
Исследования в области палеонтологии , изучение окаменелостей, подтверждают идею о том, что все живые организмы связаны между собой. Окаменелости свидетельствуют о том, что накопленные изменения в организмах в течение длительных периодов времени привели к появлению разнообразных форм жизни, которые мы видим сегодня. Окаменелость сама по себе раскрывает структуру организма и взаимоотношения между нынешними и вымершими видами, позволяя палеонтологам построить генеалогическое древо всех форм жизни на Земле. [42]
Современная палеонтология началась с работ Жоржа Кювье . Кювье отметил, что в осадочных породах каждый слой содержит определенную группу окаменелостей. Более глубокие слои, которые он считал более древними, содержали более простые формы жизни. Он отметил, что многие формы жизни из прошлого сегодня уже не существуют. Одним из успешных вкладов Кювье в понимание летописи окаменелостей было установление факта вымирания. Пытаясь объяснить вымирание, Кювье предложил идею «революций» или катастрофизма , в которой он предположил, что геологические катастрофы происходили на протяжении всей истории Земли, уничтожая большое количество видов. [43] Теория революций Кювье позже была заменена униформистскими теориями, особенно теориями Джеймса Хаттона и Чарльза Лайелла , которые предположили, что геологические изменения Земли были постепенными и последовательными. [44] Однако нынешние данные в летописи окаменелостей подтверждают концепцию массового вымирания. В результате общая идея катастрофизма вновь стала обоснованной гипотезой, по крайней мере, для некоторых быстрых изменений форм жизни, которые появляются в летописях окаменелостей.
В настоящее время обнаружено и идентифицировано очень большое количество окаменелостей. Эти окаменелости служат хронологической записью эволюции. Летопись окаменелостей предоставляет примеры переходных видов , которые демонстрируют наследственные связи между прошлыми и настоящими формами жизни. [45] Одним из таких переходных ископаемых является археоптерикс , древний организм, который имел отличительные характеристики рептилии (например, длинный костистый хвост и конические зубы ), но также имел характеристики птиц (например, перья и поперечные рычаги ). Смысл такой находки заключается в том, что современные рептилии и птицы произошли от общего предка. [46]
Сравнительная анатомия [ править ]
Сравнение сходства между организмами по форме или внешнему виду частей, называемое их морфологией , издавна было способом классификации жизни на близкородственные группы. Это можно сделать, сравнивая структуру взрослых организмов разных видов или сравнивая закономерности роста, деления и даже миграции клеток во время развития организма.
Таксономия [ править ]
Таксономия – это раздел биологии, который называет и классифицирует все живые существа. Ученые используют морфологическое и генетическое сходство, чтобы классифицировать формы жизни на основе наследственных связей. Например, орангутанги , гориллы , шимпанзе и люди принадлежат к одной и той же таксономической группе, называемой семейством — в данном случае семейству Hominidae . Эти животные сгруппированы вместе из-за сходства морфологии, обусловленного общим происхождением (так называемая гомология ). [47]

Убедительные доказательства эволюции получены благодаря анализу гомологичных структур: структур у разных видов, которые больше не выполняют одну и ту же задачу, но имеют схожую структуру. [48] Так обстоит дело с передними конечностями млекопитающих . Передние конечности человека, кошки , кита и летучей мыши имеют поразительно схожую костную структуру. Однако передние конечности каждого из этих четырех видов выполняют разные задачи. Те же кости, из которых построены крылья летучей мыши, используемые для полета, также образуют ласты кита, которые используются для плавания. Такая «конструкция» не имеет смысла, если они не связаны друг с другом и созданы специально для своих конкретных задач. Теория эволюции объясняет эти гомологичные структуры: все четыре животных имели общего предка, и каждое из них претерпело изменения на протяжении многих поколений. Эти изменения в строении привели к тому, что передние конечности стали приспособлены для выполнения различных задач. [49]

Однако анатомические сравнения могут ввести в заблуждение, поскольку не все анатомические сходства указывают на близкое родство. Организмы, живущие в схожей среде обитания, часто развивают сходные физические особенности — процесс, известный как конвергентная эволюция . И акулы , и дельфины имеют схожие формы тела, но имеют лишь отдаленное родство: акулы — это рыбы , а дельфины — млекопитающие. Такое сходство является результатом того, что обе группы населения подвергаются одинаковому давлению отбора . В обеих группах предпочтение отдавалось изменениям, способствующим плаванию. Таким образом, со временем они приобрели схожий внешний вид (морфологию), хотя и не были тесно связаны. [50]
Эмбриология [ править ]
В некоторых случаях анатомическое сравнение структур эмбрионов двух или более видов дает доказательства наличия общего предка, что может быть неочевидно у взрослых форм. По мере развития эмбриона эти гомологии могут быть потеряны из виду, и структуры могут брать на себя разные функции. Частью классификации группы позвоночных (в которую входят люди) является наличие хвоста (выходящего за пределы ануса ) и глоточных щелей . Обе структуры появляются на определенной стадии эмбрионального развития, но не всегда очевидны во взрослой форме. [51]
Из-за морфологического сходства, присутствующего у эмбрионов разных видов во время развития, когда-то предполагалось, что организмы воспроизводят свою эволюционную историю в эмбрионе. Считалось, что человеческие эмбрионы прошли сначала стадию амфибии , затем стадию рептилии, прежде чем завершить свое развитие как млекопитающих. Такая реконструкция, часто называемая теорией перепросмотра , не подтверждается научными данными. Однако происходит то, что первые стадии развития у широких групп организмов схожи. [52] на очень ранних Например, стадиях все позвоночные кажутся чрезвычайно похожими, но не совсем похожими на какие-либо предковые виды. По мере продолжения разработки из этой базовой модели выявляются специфические особенности.
Рудиментарные структуры [ править ]
Гомология включает уникальную группу общих структур, называемых рудиментарными структурами . Рудиментарными называют анатомические части, которые имеют минимальную ценность для организма, который ими обладает. Эти, казалось бы, нелогичные структуры являются остатками органов, игравших важную роль у предковых форм. Так обстоит дело с китами, у которых есть небольшие рудиментарные кости, которые кажутся остатками костей ног их предков , ходивших по суше. [53] У людей также есть рудиментарные структуры, в том числе ушные мышцы , зубы мудрости , аппендикс , копчик , волосы на теле (в том числе гусиная кожа ) и полулунная складка в углу глаза . [54]
Биогеография [ править ]

Биогеография – это наука о географическом распространении видов. Данные биогеографии, особенно биогеографии океанических островов , сыграли ключевую роль в убеждении Дарвина и Альфреда Рассела Уоллеса в том, что виды эволюционировали по схеме разветвления общего происхождения. [55] На островах часто обитают эндемичные виды, виды, которые больше нигде не встречаются, но эти виды часто связаны с видами, обитающими на ближайшем континенте. Кроме того, на островах часто встречаются скопления близкородственных видов, которые занимают совершенно разные экологические ниши , то есть имеют разные способы существования в окружающей среде. Такие кластеры формируются в процессе адаптивной радиации , когда один предковый вид колонизирует остров, имеющий множество открытых экологических ниш, а затем диверсифицируется, эволюционируя в различные виды, приспособленные для заполнения этих пустых ниш. Хорошо изученные примеры включают вьюрков Дарвина , группу из 13 видов вьюрков, эндемичных для Галапагосских островов, и гавайских медоносов , группу птиц, которая когда-то, до вымирания, вызванного человеком, насчитывала 60 видов, выполняющих разнообразные экологические роли, и все они произошли от предок, похожий на одиночного вьюрка, прибывший на Гавайские острова около 4 миллионов лет назад. [56] Другим примером является альянс Silversword , группа многолетних видов растений, также эндемичных для Гавайских островов, которые обитают в различных средах обитания и имеют различные формы и размеры, включая деревья, кустарники и коврики, прилегающие к земле, но которые могут быть гибридизированы друг с другом и с некоторыми видами тарвида, встречающимися на западном побережье Северной Америки ; Похоже, что один из этих тарвидов в прошлом колонизировал Гавайи и дал начало всему альянсу Серебряного Меча. [57]
биология Молекулярная

Каждый живой организм (за исключением, пожалуй, РНК- вирусов ) содержит молекулы ДНК, несущие генетическую информацию. Гены — это участки ДНК, которые несут эту информацию и влияют на свойства организма. Гены определяют внешний вид человека и в некоторой степени его поведение. Если два организма тесно связаны, их ДНК будет очень похожа. [58] С другой стороны, чем более отдаленно родственны два организма, тем больше у них различий. Например, братья являются близкими родственниками и имеют очень схожую ДНК, в то время как двоюродные братья имеют более отдаленное родство и имеют гораздо больше различий в ДНК. Сходства в ДНК используются для определения отношений между видами почти так же, как они используются для демонстрации отношений между особями. Например, сравнение шимпанзе с гориллами и людьми показывает, что сходство между ДНК человека и шимпанзе составляет целых 96 процентов. Сравнение ДНК показывает, что люди и шимпанзе более тесно связаны друг с другом, чем любой из видов с гориллами. [59] [60] [61]
Область молекулярной систематики фокусируется на измерении сходства этих молекул и использовании этой информации для выяснения того, как различные типы организмов связаны в ходе эволюции. Эти сравнения позволили биологам построить дерево взаимоотношений эволюции жизни на Земле. [62] Они даже позволили ученым разгадать взаимоотношения между организмами, чьи общие предки жили так давно, что во внешнем виде организмов не осталось никакого реального сходства.
Искусственный отбор [ править ]

Искусственный отбор – это контролируемое разведение домашних растений и животных. Люди определяют, какое животное или растение будет воспроизводиться и кто из потомков выживет; таким образом, они определяют, какие гены будут переданы будущим поколениям. Процесс искусственного отбора оказал существенное влияние на эволюцию домашних животных. Например, люди вывели разные типы собак путем контролируемого разведения. Различия в размерах между чихуахуа и немецким догом являются результатом искусственного отбора. Несмотря на совершенно разный внешний вид, они и все другие собаки произошли от нескольких волков , одомашненных людьми на территории современного Китая менее 15 000 лет назад. [63]
Искусственный отбор позволил получить большое разнообразие растений. Что касается кукурузы , недавние генетические данные свидетельствуют о том, что одомашнивание произошло 10 000 лет назад в центральной Мексике. [64] [ ненадежный источник? ] До одомашнивания съедобная часть дикой формы была небольшой, и ее было трудно собрать. Сегодня Центр сотрудничества в области генетики кукурузы • Фондовый центр хранит коллекцию из более чем 10 000 генетических вариаций кукурузы, возникших в результате случайных мутаций и хромосомных отклонений от исходного дикого типа. [65]
При искусственном отборе появляется новая порода или разновидность со случайными мутациями, привлекательными для человека, тогда как при естественном отборе выживающим видом является вид со случайными мутациями, полезными для него в нечеловеческой среде обитания. И при естественном, и при искусственном отборе вариации являются результатом случайных мутаций, а лежащие в их основе генетические процессы по существу одинаковы. [66] Дарвин внимательно наблюдал за результатами искусственного отбора у животных и растений, чтобы сформировать многие из своих аргументов в пользу естественного отбора. [67] Большая часть его книги «Происхождение видов» была основана на этих наблюдениях за многими разновидностями домашних голубей, возникшими в результате искусственного отбора. Дарвин предположил, что если люди смогут добиться драматических изменений в домашних животных за короткие периоды времени, то естественный отбор, продлившийся миллионы лет, мог бы привести к различиям, наблюдаемым в живых существах сегодня.
Коэволюция [ править ]
Коэволюция – это процесс, в котором два или более видов влияют на эволюцию друг друга. На все организмы влияет жизнь вокруг них; однако в ходе коэволюции есть свидетельства того, что генетически детерминированные черты каждого вида являются прямым результатом взаимодействия между двумя организмами. [58]
Широко задокументированный случай коэволюции — это отношения между Pseudomyrmex , разновидностью муравьев , и акацией , растением, которое муравьи используют для еды и убежища. Отношения между ними настолько близки, что привели к эволюции особых структур и поведения у обоих организмов. Муравей защищает акацию от травоядных и очищает лесную подстилку от семян конкурирующих растений. В ответ на это у растения появились опухшие шипы, которые муравьи используют в качестве убежища, и особые части цветка, которые муравьи едят. [68] Такая коэволюция не означает, что муравьи и дерево решили вести себя альтруистично . Скорее, небольшие генетические изменения в популяции как у муравьев, так и у деревьев принесли пользу каждому. Это преимущество давало немного более высокий шанс передачи характеристики следующему поколению. Со временем последовательные мутации создали отношения, которые мы наблюдаем сегодня.
Видообразование [ править ]

При подходящих обстоятельствах и достаточном количестве времени эволюция приводит к появлению новых видов. Ученые изо всех сил пытались найти точное и всеобъемлющее определение вида . Эрнст Майр определил вид как популяцию или группу популяций, члены которых могут естественным образом скрещиваться друг с другом для производства жизнеспособного и плодовитого потомства. (Представители одного вида не могут производить жизнеспособное, плодовитое потомство от представителей других видов). [69] Определение Майра получило широкое признание среди биологов, но не применимо к таким организмам, как бактерии , которые размножаются бесполым путем .
Видообразование — это событие разделения линии, в результате которого из одной общей предковой популяции образуются два отдельных вида. [15] Широко распространенный метод видообразования называется аллопатрическим видообразованием . Аллопатрическое видообразование начинается, когда популяция становится географически разделенной. [48] Геологические процессы, такие как появление горных хребтов, образование каньонов или затопление сухопутных мостов из-за изменений уровня моря, могут привести к образованию отдельных популяций. Для того чтобы произошло видообразование, разделение должно быть существенным, чтобы генетический обмен между двумя популяциями был полностью нарушен. В своей отдельной среде обитания генетически изолированные группы следуют своим уникальным эволюционным путям. Каждая группа будет накапливать разные мутации, а также подвергаться разному давлению отбора. Накопленные генетические изменения могут привести к разделению популяций, которые больше не смогут скрещиваться, если воссоединятся. [15] Барьеры, препятствующие скрещиванию, бывают презиготными (предотвращают спаривание или оплодотворение) или постзиготическими (барьеры, возникающие после оплодотворения). Если скрещивание уже невозможно, то их будут считать разными видами. [70] Результатом четырех миллиардов лет эволюции стало разнообразие жизни вокруг нас: сегодня существует около 1,75 миллиона различных видов. [71] [72]
Обычно процесс видообразования протекает медленно и занимает очень длительные промежутки времени; таким образом, прямые наблюдения на протяжении всей человеческой жизни редки. Однако видообразование наблюдается у современных организмов, а прошлые события видообразования зафиксированы в окаменелостях. [73] [74] [75] Ученые задокументировали образование пяти новых видов рыб-цихлид от одного общего предка, который был изолирован менее 5000 лет назад от родительского стада в озере Нагубаго. [76] Доказательством видообразования в данном случае была морфология (внешний вид) и отсутствие естественного скрещивания. У этих рыб сложные брачные ритуалы и разнообразная окраска; Небольшие модификации, внесенные в новый вид, изменили процесс выбора партнера, и возникшие пять форм не удалось убедить скрещиваться. [77]
Механизм [ править ]
Теория эволюции получила широкое признание в научном сообществе и служит связующим звеном между различными областями биологии. [19] Эволюция обеспечивает биологию прочной научной базой. Значимость эволюционной теории резюмируется Феодосием Добжанским следующим образом: « Ничто в биологии не имеет смысла, кроме как в свете эволюции ». [78] [79] Тем не менее теория эволюции не статична. В научном сообществе ведется много дискуссий относительно механизмов эволюционного процесса. Например, скорость, с которой происходит эволюция, все еще обсуждается. Кроме того, существуют противоречивые мнения относительно того, что является первичной единицей эволюционных изменений — организм или ген.
Скорость изменения [ править ]
Дарвин и его современники рассматривали эволюцию как медленный и постепенный процесс. Эволюционные деревья основаны на идее, что глубокие различия между видами являются результатом множества небольших изменений, которые накапливаются в течение длительных периодов времени.
Основой градуализма послужили работы геологов Джеймса Хаттона и Чарльза Лайеля. Точка зрения Хаттона предполагает, что глубокие геологические изменения были совокупным продуктом относительно медленного продолжающегося действия процессов, которые все еще можно увидеть в действии сегодня, в отличие от катастрофизма, который продвигал идею о том, что внезапные изменения имели причины, действие которых больше нельзя увидеть. Для биологических изменений был принят униформистский подход. Такая точка зрения может показаться противоречащей летописи окаменелостей, которая часто свидетельствует о внезапном появлении новых видов, а затем сохранении в этой форме в течение длительного периода времени. В 1970-х годах палеонтологи Найлс Элдридж и Стивен Джей Гулд разработали теоретическую модель, которая предполагает, что эволюция, хотя и медленный процесс с человеческой точки зрения, претерпевает периоды относительно быстрых изменений (от 50 000 до 100 000 лет). [80] чередующиеся с длительными периодами относительной стабильности. Их теория называется прерывистым равновесием и объясняет летопись окаменелостей, не противореча идеям Дарвина. [81]
Единица изменения [ править ]

Общей единицей отбора в эволюции является организм. Естественный отбор происходит, когда репродуктивный успех особи улучшается или снижается из-за унаследованных характеристик, а репродуктивный успех измеряется количеством выживших потомков особи. Взгляд на организм подвергался сомнению со стороны различных биологов, а также философов. Эволюционный биолог Ричард Докинз предполагает, что многое можно понять, если посмотреть на эволюцию с точки зрения генов; то есть естественный отбор действует как эволюционный механизм как на гены, так и на организмы. [82] В своей книге 1976 года «Эгоистичный ген » он объясняет:
Индивиды не являются стабильными вещами, они мимолетны. Хромосомы тоже тасуются до небытия, как карты в руках вскоре после раздачи. Но сами карты перетасовываются. Карты — это гены. Гены не уничтожаются при кроссинговере, они просто меняют партнеров и продолжают свой путь. Конечно, они идут дальше. Это их дело. Они — репликаторы, а мы — их машины выживания. Когда мы выполнили свою задачу, нас отбрасывают в сторону. Но гены — обитатели геологического времени: гены вечны. [83]
Другие считают, что отбор действует на многих уровнях, а не только на одном уровне организма или гена; например, Стивен Джей Гулд призвал к иерархическому подходу к отбору. [84]
См. также [ править ]
Ссылки [ править ]
- ^ «Возраст Земли» . Геологическая служба США . 9 июля 2007 года . Проверено 29 мая 2015 г.
- ^ Далримпл 2001 , стр. 205–221.
- ^ Манхеса, Жерар; Аллегре, Клод Ж .; Дюпреа, Бернар; Хамельн, Бруно (май 1980 г.). «Изотопное исследование свинца основных-ультраосновных слоистых комплексов: предположения о возрасте Земли и характеристиках примитивной мантии». Письма о Земле и планетологии . 47 (3): 370–382. Бибкод : 1980E&PSL..47..370M . дои : 10.1016/0012-821X(80)90024-2 .
- ^ Шопф, Дж. Уильям ; Кудрявцев Анатолий Б.; Чая, Эндрю Д.; Трипати, Абхишек Б. (5 октября 2007 г.). «Свидетельства архейской жизни: строматолиты и микроокаменелости». Докембрийские исследования . 158 (3–4): 141–155. Бибкод : 2007PreR..158..141S . doi : 10.1016/j.precamres.2007.04.009 .
- ^ Шопф, Дж. Уильям (29 июня 2006 г.). «Ископаемые свидетельства архейской жизни» . Философские труды Королевского общества Б. 361 (1470): 869–885. дои : 10.1098/rstb.2006.1834 . ПМЦ 1578735 . ПМИД 16754604 .
- ^ Рэйвен и Джонсон 2002 , с. 68
- ^ «Заблуждения об эволюции» . Понимание эволюции . Калифорнийский университет в Беркли . Проверено 26 сентября 2015 г.
- ^ Футуйма 2005а
- ↑ Перейти обратно: Перейти обратно: а б Гулд 2002 г.
- ↑ Перейти обратно: Перейти обратно: а б Грегори, Т. Райан (июнь 2009 г.). «Понимание естественного отбора: основные понятия и распространенные заблуждения» . Эволюция: образование и информационно-пропагандистская деятельность . 2 (2): 156–175. дои : 10.1007/s12052-009-0128-1 .
- ^ Ри, Сын Ён. «Грегор Мендель (1822–1884)» . Доступ к совершенству . Атланта, Джорджия: Национальный музей здравоохранения. Архивировано из оригинала 27 декабря 2014 г. Проверено 7 января 2015 г.
- ^ Фарбер 2000 , с. 136
- ^ Дарвин 2005 г.
- ↑ Перейти обратно: Перейти обратно: а б Элдридж, Найлз (весна 2006 г.). «Исповедь дарвиниста» . Ежеквартальный обзор Вирджинии . 82 (2). Шарлоттсвилл, Вирджиния: Университет Вирджинии : 32–53 . Проверено 7 января 2015 г.
- ↑ Перейти обратно: Перейти обратно: а б с Кваммен, Дэвид (ноябрь 2004 г.). «Был ли Дарвин не прав?» . National Geographic (дополнительно онлайн). Вашингтон, округ Колумбия: Национальное географическое общество . Архивировано из оригинала 15 декабря 2007 года . Проверено 23 декабря 2007 г.
- ↑ Перейти обратно: Перейти обратно: а б ван Вай, Джон (2002). «Чарльз Дарвин: джентльмен-натуралист» . Полное собрание сочинений Чарльза Дарвина в Интернете . OCLC 74272908 . Проверено 16 января 2008 г.
- ^ ван Вай, Джон (2002). «Оплодотворение орхидей» . Полное собрание сочинений Чарльза Дарвина в Интернете . OCLC 74272908 . Проверено 7 января 2008 г.
- ^ Дарвин 1859 .
- ↑ Перейти обратно: Перейти обратно: а б Дельгадо, Синтия (28 июля 2006 г.). «В поисках эволюции в медицине» . Запись НИЗ . 58 (15): 1, 8–9.
- ^ Футуйма 2005b , стр. 93–98.
- ^ Дарвин 1872 , с. 108 , «Последствия увеличения использования и неиспользования частей, контролируемые естественным отбором».
- ^ Гизелин, Майкл Т. (сентябрь – октябрь 1994 г.). «Воображаемый Ламарк: взгляд на фальшивую «историю» в школьных учебниках» . Учебное письмо . Саусалито, Калифорния: Лига учебников. OCLC 23228649 . Архивировано из оригинала 12 февраля 2008 г. Проверено 23 января 2008 г.
- ^ «Секс и генетическая перетасовка» . Понимание эволюции . Калифорнийский университет в Беркли . Проверено 8 января 2015 г.
- ^ «Поток генов» . Понимание эволюции . Калифорнийский университет в Беркли . Проверено 8 января 2015 г.
- ^ Гулд 1980 , с. 24
- ^ Бейдер, Ларс; Холл, Брайан К. (ноябрь 2002 г.). «Конечности у китов и отсутствие конечностей у других позвоночных: механизмы эволюционной трансформации и потери». Эволюция и развитие . 4 (6): 445–458. дои : 10.1046/j.1525-142X.2002.02033.x . ПМИД 12492145 . S2CID 8448387 .
- ^ Буфнер, Джулия К.; Бухтова, Марсела; Фу, Кэтрин; и др. (25 июня 2007 г.). «Эмбриональное развитие Python sebae – I: критерии стадирования и макроскопический морфогенез скелета головы и конечностей». Зоология . 110 (3): 212–230. дои : 10.1016/j.zool.2007.01.005 . ПМИД 17499493 .
- ^ «Часто задаваемые вопросы об эволюции» . Библиотека эволюции (веб-ресурс). Эволюция . Бостон, Массачусетс: Образовательный фонд WGBH ; Clear Blue Sky Productions, Inc. 2001. OCLC 48165595 . Проверено 23 января 2008 г.
- ^ «Современное массовое вымирание?» . Библиотека эволюции (веб-ресурс). Эволюция. Бостон, Массачусетс: Образовательный фонд WGBH; Clear Blue Sky Productions, Inc. 2001. OCLC 48165595 . Проверено 23 января 2008 г.
- ^ Бамбах, Ричард К.; Нолл, Эндрю Х .; Ван, Стив К. (декабрь 2004 г.). «Происхождение, исчезновение и массовое истощение морского разнообразия» (PDF) . Палеобиология . 30 (4): 522–542. Бибкод : 2004Pbio...30..522B . doi : 10.1666/0094-8373(2004)030<0522:OEAMDO>2.0.CO;2 . S2CID 17279135 .
- ^ Футуйма 1998 , с. Глоссарий
- ^ Элстранд, Норман К.; Элам, Дайан Р. (ноябрь 1993 г.). «Популяционно-генетические последствия небольшой численности популяции: значение для сохранения растений». Ежегодный обзор экологии и систематики . 24 : 217–242. doi : 10.1146/annurev.es.24.110193.001245 .
- ^ Юэнс 2004
- ↑ Перейти обратно: Перейти обратно: а б Кэмпбелл и Рис, 2002 , стр. 445–463.
- ^ Робинсон, Ричард, изд. (2003). «Узкое место народонаселения» . Генетика . Нью-Йорк: Справочник Macmillan, США . ISBN 978-0-02-865606-9 . LCCN 2002003560 . OCLC 49278650 . Проверено 7 апреля 2011 г.
- ^ Футуйма 1998 , стр. 303–304.
- ^ Хельцель, А. Рус; Флейшер, Роберт С.; Кампанья, Клаудио; и др. (июль 2002 г.). «Влияние узкого места в популяции на симметрию и генетическое разнообразие северного морского слона» . Журнал эволюционной биологии . 15 (4): 567–575. дои : 10.1046/j.1420-9101.2002.00419.x . S2CID 85821330 .
- ^ «Генетический дрейф и эффект основателя» . Библиотека эволюции (веб-ресурс). Эволюция. Бостон, Массачусетс: Образовательный фонд WGBH ; Clear Blue Sky Productions, Inc. 2001. OCLC 48165595 . Проверено 7 апреля 2009 г.
- ↑ Перейти обратно: Перейти обратно: а б Ларсон 2004 , стр. 231–237.
- ^ Моран, Лоуренс (22 января 1993 г.). «Современный синтез генетики и эволюции» . Архив TalkOrigins . Хьюстон, Техас: Фонд TalkOrigins . Проверено 9 января 2015 г.
- ^ Боулер 2003 , стр. 325–339.
- ^ «Ископаемая пластинка – эпопея жизни» . Виртуальный музей окаменелостей . Роджер Перкинс . Проверено 31 августа 2007 г.
- ^ Таттерсолл 1995 , стр. 5–6.
- ^ Лайель 1830 , с. 76
- ^ НАН 2008 , с. 22
- ^ Гулд 1995 , с. 360
- ^ Даймонд 1992 , с. 16
- ↑ Перейти обратно: Перейти обратно: а б «Глоссарий» . Библиотека эволюции (веб-ресурс). Эволюция. Бостон, Массачусетс: Образовательный фонд WGBH; Clear Blue Sky Productions, Inc. 2001. OCLC 48165595 . Проверено 23 января 2008 г.
- ^ Майр 2001 , стр. 25–27.
- ^ Джонсон, Джордж Б. «Конвергентная и дивергентная эволюция» . Справочники доктора Джорджа Джонсона . Сент-Луис, Миссури: Txtwriter, Inc. Архивировано из оригинала 10 марта 2008 г. Проверено 6 января 2015 г.
- ^ Вейхерт и Преш 1975 , с. 8
- ^ Миллер, Кеннет Р .; Левин, Джозеф (1997). «Геккель и его эмбрионы: заметка об учебниках» . Ресурсы эволюции . Рехобот, Массачусетс: Биология Миллера и Левина . Проверено 2 сентября 2007 г.
- ^ Теобальд, Дуглас (2004). «29+ доказательств макроэволюции» (PDF) . Архив TalkOrigins . Хьюстон, Техас: The TalkOrigins Foundation, Inc. Часть 2: «Киты с задними конечностями» . Проверено 27 января 2008 г.
- ^ Джонсон, Джордж Б. «Рудиментарные структуры» . Справочники доктора Джорджа Джонсона . Сент-Луис, Миссури: Txtwriter, Inc. Архивировано из оригинала 10 марта 2008 г. Проверено 6 января 2015 г.
- ^ Боулер 2003 , стр. 150–151, 174.
- ^ Койн 2009 , стр. 102–103.
- ^ Карр, Джеральд Д. «Адаптивная радиация и гибридизация в Гавайском альянсе Серебряного меча» . Гонолулу, Гавайи: Гавайского университета Факультет ботаники . Проверено 10 января 2015 г.
- ↑ Перейти обратно: Перейти обратно: а б НАН 1998 г. , «Эволюция и природа науки».
- ^ Ловгрен, Стефан (31 августа 2005 г.). «Шимпанзе и люди на 96 процентов одинаковы, результаты исследования генов» . Национальные географические новости . Вашингтон, округ Колумбия: Национальное географическое общество. Архивировано из оригинала 5 сентября 2005 года . Проверено 23 декабря 2007 г.
- ^ Кэрролл, Гренье и Уэзерби, 2005 г.
- ^ «Минусы: настройки 46-полосного трека» . UCSC Геномная биоинформатика . Санта-Крус, Калифорния: Калифорнийский университет, Санта-Крус . Проверено 10 января 2015 г.
- ^ Сиссарелли, Франческа Д.; Доркс, Тобиас; фон Меринг, Кристиан; и др. (3 марта 2006 г.). «На пути к автоматической реконструкции древа жизни с высоким разрешением». Наука . 311 (5765): 1283–1287. Бибкод : 2006Sci...311.1283C . CiteSeerX 10.1.1.381.9514 . дои : 10.1126/science.1123061 . ПМИД 16513982 . S2CID 1615592 .
- ^ МакГурти, Кристина (22 ноября 2002 г.). «Происхождение собак прослежено» . Новости Би-би-си . Лондон: Би-би-си . Проверено 14 декабря 2007 г.
- ^ Холл, Харди (7 ноября 2005 г.). «Бегство трансгенов: находятся ли традиционные сорта кукурузы в Мексике под угрозой со стороны трансгенных культур кукурузы» . Научно-творческий ежеквартальный журнал . Ванкувер и Келоуна, Британская Колумбия: Университет Британской Колумбии . Проверено 14 декабря 2007 г.
- ^ «Информация о кукурузном кооперативе» . Национальная система зародышевой плазмы растений . Министерство сельского хозяйства США ; Служба сельскохозяйственных исследований . 23 ноября 2010 года . Проверено 10 января 2015 г.
- ^ Сильверман, Дэвид (2002). «Лучшие книги методом проб и ошибок» . Большая инструкция жизни . Архивировано из оригинала 17 января 2015 г. Проверено 4 апреля 2008 г.
- ^ Вилнер, Эдуардо (март 2006 г.). «Искусственный отбор Дарвина как эксперимент». Исследования по истории и философии науки. Часть C: Исследования по истории и философии биологических и биомедицинских наук . 37 (1): 26–40. дои : 10.1016/j.shpsc.2005.12.002 . ПМИД 16473266 .
- ^ Янзен, Дэниел Х. (1974). «Акации с опухшими шипами Центральной Америки» (PDF) . Смитсоновский вклад в ботанику . 13 (13). Вашингтон, округ Колумбия: Издательство Смитсоновского института : 1–131. дои : 10.5479/si.0081024X.13 . OCLC 248582653 . Проверено 31 августа 2007 г.
- ^ Майр 2001 , стр. 165–169.
- ^ Саллоуэй, Фрэнк Дж. (декабрь 2005 г.). «Эволюция Чарльза Дарвина» . Смитсоновский институт . Вашингтон, округ Колумбия: Смитсоновский институт . Проверено 11 января 2015 г.
- ^ Кавальер-Смит, Томас (29 июня 2006 г.). «Эволюция клеток и история Земли: застой и революция» . Философские труды Королевского общества B: Биологические науки . 361 (1470): 969–1006. дои : 10.1098/rstb.2006.1842 . ПМЦ 1578732 . ПМИД 16754610 .
- ^ «Сколько существует видов?» . Enviroliteracy.org . Вашингтон, округ Колумбия: Совет экологической грамотности. 17 июня 2008 года . Проверено 11 января 2015 г.
- ^ Джиггинс, Крис Д.; Уздечка, Джон Р. (март 2004 г.). «Видообразование яблочной личинки: смесь винтажей?». Тенденции в экологии и эволюции . 19 (3): 111–114. дои : 10.1016/j.tree.2003.12.008 . ПМИД 16701238 .
- ^ Боксхорн, Джозеф (1 сентября 1995 г.). «Наблюдаемые случаи видообразования» . Архив TalkOrigins . Хьюстон, Техас: Фонд TalkOrigins, Inc. Проверено 10 мая 2007 г.
- ^ Вайнберг, Джеймс Р.; Старчак, Виктория Р.; Йорг, Даниэле (август 1992 г.). «Доказательства быстрого видообразования после события основателя в лаборатории». Эволюция . 46 (4): 1214–1220. дои : 10.2307/2409766 . JSTOR 2409766 . ПМИД 28564398 .
- ^ Майр 1970 , с. 348
- ^ Майр 1970 г.
- ^ Добжанский, Феодосий (март 1973 г.). «Ничто в биологии не имеет смысла, кроме как в свете эволюции». Американский учитель биологии . 35 (3): 125–129. CiteSeerX 10.1.1.525.3586 . дои : 10.2307/4444260 . JSTOR 4444260 . S2CID 207358177 .
- ^ Скотт, Юджини К. (17 октября 2008 г.) [первоначально опубликовано на PBS Online, май 2000 г.]. «Можно и нельзя преподавать эволюцию» . Национальный центр научного образования (блог). Беркли, Калифорния: Национальный центр научного образования . Проверено 1 января 2008 г.
- ^ Аяла, Франсиско Дж. (2005). « Структура эволюционной теории : о монументальном шедевре Стивена Джея Гулда». Теология и наука . 3 (1): 104. дои : 10.1080/14746700500039800 . S2CID 4293004 .
- ^ Гулд, Стивен Джей (август 1991 г.). «Опус 200» (PDF) . Естественная история . 100 (8). Research Triangle Park, Северная Каролина: Журнал Natural History, Inc.: 12–18.
- ^ Райт, Сьюэлл (сентябрь 1980 г.). «Геническая и организменная селекция». Эволюция . 34 (5): 825–843. дои : 10.2307/2407990 . JSTOR 2407990 . ПМИД 28581131 .
- ^ Докинз 1976 , с. 35
- ^ Гулд, Стивен Джей; Ллойд, Элизабет А. (12 октября 1999 г.). «Индивидуальность и адаптация на разных уровнях отбора: как нам назвать и обобщить единицу дарвинизма?» . Учеб. Натл. акад. наук. США 96 (21): 11904–11909. Бибкод : 1999PNAS...9611904G . дои : 10.1073/pnas.96.21.11904 . ЧВК 18385 . ПМИД 10518549 .
Библиография [ править ]
- Боулер, Питер Дж. (2003). Эволюция: История идеи (3-е полностью переработанное и расширенное изд.). Беркли, Калифорния: Издательство Калифорнийского университета . ISBN 978-0-520-23693-6 . LCCN 2002007569 . OCLC 49824702 .
- Кэмпбелл, Нил А .; Рис, Джейн Б. (2002). «Эволюция популяций» . Биология . Том. 6-й. Сан-Франциско, Калифорния: Бенджамин Каммингс . ISBN 978-0-8053-6624-2 . LCCN 2001047033 . OCLC 47521441 .
- Кэрролл, Шон Б .; Гренье, Дженнифер К.; Уэзерби, Скотт Д. (2005). От ДНК к разнообразию: молекулярная генетика и эволюция дизайна животных (2-е изд.). Молден, Массачусетс: Blackwell Publishing . ISBN 978-1-4051-1950-4 . LCCN 2003027991 . OCLC 53972564 .
- Койн, Джерри А. (2009). Почему эволюция верна . Нью-Йорк: Викинг . ISBN 978-0-670-02053-9 . LCCN 2008033973 . OCLC 233549529 .
- Далримпл, Дж. Брент (2001). «Возраст Земли в двадцатом веке: проблема (в основном) решена». В Льюисе, CLE; Нелл, С.Дж. (ред.). Возраст Земли: с 4004 г. до н.э. по 2002 г. н.э. Специальное издание Геологического общества. Том. 190. Лондон: Лондонское геологическое общество . стр. 205–221. Бибкод : 2001GSLSP.190..205D . дои : 10.1144/gsl.sp.2001.190.01.14 . ISBN 978-1-86239-093-5 . LCCN 2003464816 . ОСЛК 48570033 . S2CID 130092094 .
{{cite book}}:|journal=игнорируется ( помогите ) - Дарвин, Чарльз (1859). О происхождении видов путем естественного отбора, или о сохранении избранных рас в борьбе за жизнь (1-е изд.). Лондон: Джон Мюррей . LCCN 06017473 . OCLC 741260650 .
- Дарвин, Чарльз (1872). Происхождение видов путем естественного отбора, или сохранение избранных рас в борьбе за жизнь (6-е изд.). Лондон: Джон Мюррей. LCCN 13003393 . OCLC 228738610 .
- Дарвин, Чарльз (2005). Дарвин: Неизгладимая печать: эволюция идеи . Отредактировано, с предисловием и комментариями Джеймса Д. Уотсона . Филадельфия, Пенсильвания: Бегущая пресса . ISBN 978-0-7624-2136-7 . OCLC 62172858 .
- Докинз, Ричард (1976). Эгоистичный ген (1-е изд.). Нью-Йорк: Издательство Оксфордского университета . ISBN 978-0-19-857519-1 . LCCN 76029168 . ОСЛК 2681149 .
- Даймонд, Джаред (1992). Третий шимпанзе: эволюция и будущее человеческого животного (1-е изд.). Нью-Йорк: ХарперКоллинз . ISBN 978-0-06-018307-3 . LCCN 91050455 . OCLC 24246928 .
- Юэнс, Уоррен Дж. (2004). Математическая популяционная генетика . Междисциплинарная прикладная математика. Том. Я. Теоретическое введение (2-е изд.). Нью-Йорк: Springer-Verlag Нью-Йорк . ISBN 978-0-387-20191-7 . LCCN 2003065728 . OCLC 53231891 .
- Фарбер, Пол Лоуренс (2000). В поисках порядка в природе: традиция натуралистов от Линнея до Э.О. Вильсона . Вводные исследования Джона Хопкинса по истории науки. Балтимор, Мэриленд: Издательство Университета Джонса Хопкинса . ISBN 978-0-8018-6390-5 . LCCN 99089621 . OCLC 43115207 .
- Футуйма, Дуглас Дж. (1998). Эволюционная биология (3-е изд.). Сандерленд, Массачусетс: Sinauer Associates . ISBN 978-0-87893-189-7 . LCCN 97037947 . ОСЛК 37560100 .
- Футуйма, Дуглас Дж. (2005a). Эволюция . Сандерленд, Массачусетс: Sinauer Associates. ISBN 978-0-87893-187-3 . LCCN 2004029808 . OCLC 57311264 .
- Футуйма, Дуглас Дж. (2005b). «Природа естественного отбора» (PDF) . В Cracraft, Джоэл; Байби, Роджер В. (ред.). Эволюционная наука и общество: воспитание нового поколения . Колорадо-Спрингс, Колорадо: Учебная программа по биологическим наукам . ISBN 978-1-929614-23-3 . OCLC 64228003 . Архивировано из оригинала (PDF) 18 июля 2015 г. Проверено 6 декабря 2014 г. «Пересмотренные материалы BSCS, симпозиума AIBS, ноябрь 2004 г., Чикаго, Иллинойс»
- Гулд, Стивен Джей (1980). Большой палец панды: больше размышлений о естествознании (1-е изд.). Нью-Йорк: WW Norton & Company . ISBN 978-0-393-01380-1 . LCCN 80015952 . OCLC 6331415 .
- Гулд, Стивен Джей (1995). Динозавр в стоге сена: размышления в естествознании . Нью-Йорк: Книги Гармонии. ISBN 978-0-517-70393-9 . LCCN 95051333 . OCLC 33892123 .
- Гулд, Стивен Джей (2002). Структура эволюционной теории . Кембридж, Массачусетс: Belknap Press издательства Гарвардского университета . ISBN 978-0-674-00613-3 . LCCN 2001043556 . OCLC 47869352 .
- Ларсон, Эдвард Дж. (2004). Эволюция: замечательная история научной теории . Нью-Йорк: Современная библиотека . ISBN 978-0-679-64288-6 . LCCN 2003064888 . OCLC 53483597 .
- Лайель, Чарльз (1830). Принципы геологии как попытка объяснить прежние изменения земной поверхности с помощью причин, действующих сейчас . Лондон: Джон Мюррей. LCCN 45042424 . OCLC 670356586 .
- Майр, Эрнст (1970). Популяции, виды и эволюция: краткое изложение видов животных и эволюции . Кембридж, Массачусетс: Belknap Press издательства Гарвардского университета. ISBN 978-0-674-69010-3 . LCCN 79111486 . OCLC 114063 .
- Майр, Эрнст (2001). Что такое эволюция . Нью-Йорк: Основные книги. ISBN 978-0-465-04426-9 . LCCN 2001036562 . OCLC 47443814 .
- МакКинни, Майкл Л. (1997). «Как редким видам удается избежать вымирания? Палеонтологический взгляд». В Кунине, Уильям Э.; Гастон, Кевин Дж. (ред.). Биология редкости: причины и последствия редких и общих различий (1-е изд.). Лондон; Нью-Йорк: Чепмен и Холл . ISBN 978-0-412-63380-5 . LCCN 96071014 . OCLC 36442106 .
- Миллер, Дж. Тайлер; Спулман, Скотт Э. (2012). Наука об окружающей среде (14-е изд.). Бельмонт, Калифорния: Брукс/Коул . ISBN 978-1-111-98893-7 . LCCN 2011934330 . OCLC 741539226 .
- Национальная академия наук (1998). Учение об эволюции и природе науки . Вашингтон, округ Колумбия: Издательство национальных академий . ISBN 978-0-309-06364-7 . LCCN 98016100 . OCLC 245727856 . Проверено 10 января 2015 г.
- Национальная академия наук; Медицинский институт (2008). Наука, эволюция и креационизм . Вашингтон, округ Колумбия: Издательство национальных академий. ISBN 978-0-309-10586-6 . LCCN 2007015904 . OCLC 123539346 . Проверено 31 июля 2014 г.
- Рауп, Дэвид М. (1992) [первоначально опубликовано в 1991 году]. Вымирание: плохие гены или невезение? . Том. 131. Введение Стивена Джея Гулда. Нью-Йорк: WW Norton & Company. стр. 46–49. ISBN 978-0-393-30927-0 . LCCN 90027192 . OCLC 28725488 . ПМИД 11540932 .
{{cite book}}:|journal=игнорируется ( помогите ) - Рэйвен, Питер Х .; Джонсон, Джордж Б. (2002). Биология (6-е изд.). Бостон, Массачусетс: МакГроу-Хилл . ISBN 978-0-07-112261-0 . LCCN 2001030052 . OCLC 45806501 .
- Таттерсолл, Ян (1995). Ископаемый след: откуда мы знаем то, что, по нашему мнению, мы знаем об эволюции человека . Нью-Йорк: Издательство Оксфордского университета. ISBN 978-0-19-506101-7 . LCCN 94031633 . ОСЛК 30972979 .
- Вейхерт, Чарльз К.; Преш, Уильям (1975). Элементы анатомии хордовых . Нью-Йорк: МакГроу-Хилл. ISBN 978-0-07-069008-0 . LCCN 74017151 . ОСЛК 1032188 .
Дальнейшее чтение [ править ]
- Чарльзуорт, Брайан ; Чарльзуорт, Дебора (2003). Эволюция: очень краткое введение . Очень короткие вступления. Оксфорд; Нью-Йорк: Издательство Оксфордского университета. ISBN 978-0-19-280251-4 . LCCN 2003272247 . OCLC 51668497 .
- Эллис, Р. Джон (2010). Как работает наука: Эволюция: учебник для студентов . Дордрехт; Нью-Йорк: Спрингер . дои : 10.1007/978-90-481-3183-9 . ISBN 978-90-481-3182-2 . LCCN 2009941981 . OCLC 465370643 .
- Хорвиц, Лесли Алан (2002). Полное руководство идиота по эволюции . Индианаполис, Индиана: Alpha Books . ISBN 978-0-02-864226-0 . LCCN 2001094735 . OCLC 48402612 .
- Круконис, Грег; Барр, Трейси Л. (2008). Эволюция для чайников . Хобокен, Нью-Джерси: John Wiley & Sons . ISBN 978-0-470-11773-6 . LCCN 2008922285 . OCLC 183916075 .
- Паллен, Марк Дж. (2009). Примерное руководство по эволюции . Грубые руководства Справочные руководства. Лондон; Нью-Йорк: Грубые гиды . ISBN 978-1-85828-946-5 . LCCN 2009288090 . OCLC 233547316 .
- Сис, Питер (2003). Древо жизни: книга, изображающая жизнь Чарльза Дарвина, натуралиста, геолога и мыслителя (1-е изд.). Нью-Йорк: Фаррар, Штраус и Жиру . ISBN 978-0-374-45628-3 . LCCN 2002040706 . OCLC 50960680 .
- Томсон, Кейт (2005). Окаменелости: очень краткое введение . Очень короткие вступления. Оксфорд; Нью-Йорк: Издательство Оксфордского университета. ISBN 978-0-19-280504-1 . LCCN 2005022027 . OCLC 61129133 .
- Вильярреал, Луис (2005). Вирусы и эволюция жизни . АСМ Пресс.
- Циммер, Карл (2010). Запутанный банк: введение в эволюцию . Гринвуд-Виллидж, Колорадо: Издательство Roberts & Company. ISBN 978-0-9815194-7-0 . LCCN 2009021802 . OCLC 403851918 .
Внешние ссылки [ править ]
- «Введение в эволюцию и естественный отбор» . Ханская академия . Проверено 6 января 2015 г.
- Брэйн, Маршалл (25 июля 2001 г.). «Как работает эволюция» . Как все работает . Проверено 6 января 2015 г.
- «Архив Talk.Origins: часто задаваемые вопросы по эволюции» . Архив TalkOrigins . Хьюстон, Техас: Фонд TalkOrigins, Inc. Проверено 12 января 2015 г.
- «Понимание эволюции: ваш универсальный ресурс информации об эволюции» . Калифорнийский университет в Беркли . Проверено 8 января 2015 г.