Бесполое размножение

Бесполое размножение — тип размножения , не предполагающий слияния гамет или изменения числа хромосом . Потомство, возникающее в результате бесполого размножения от одноклеточных или многоклеточных организмов, наследует полный набор генов своего единственного родителя, и, таким образом, вновь созданная особь генетически и физически подобна родителю или является точным клоном родителя. Бесполое размножение является основной формой размножения одноклеточных организмов, таких как археи и бактерии . Многие эукариотические организмы, включая растения , животные и грибы , также могут размножаться бесполым путем. [1] У позвоночных наиболее распространенной формой бесполого размножения является партеногенез , который обычно используется в качестве альтернативы половому размножению во времена, когда репродуктивные возможности ограничены. Драконы Комодо и некоторые вараны могут размножаться бесполым путем. [2]
Хотя все прокариоты размножаются без образования и слияния гамет, механизмы латерального переноса генов , такие как конъюгация , трансформация и трансдукция, можно сравнить с половым размножением в смысле генетической рекомбинации в мейозе . [3] [4]
Типы бесполого размножения [ править ]
Деление [ править ]
Прокариоты ( археи и бактерии ) размножаются бесполым путем путем бинарного деления , при котором родительский организм делится надвое, образуя два генетически идентичных дочерних организма. Эукариоты (такие как простейшие и одноклеточные грибы ) могут размножаться функционально сходным образом путем митоза ; большинство из них также способны к половому размножению.
Множественное деление на клеточном уровне происходит у многих протистов , например у споровиков и водорослей . Ядро . родительской клетки несколько раз делится митозом , образуя несколько ядер Затем цитоплазма разделяется, образуя множество дочерних клеток . [5] [6] [7]
У apicomplexans множественное деление, или шизогония, проявляется либо как мерогония , спорогония или гаметогония . В результате мерогонии образуются мерозоиты , представляющие собой несколько дочерних клеток, возникающих внутри одной и той же клеточной мембраны. [8] [9] В результате спорогонии образуются спорозоиты , а в результате гаметогонии образуются микрогаметы .
Расцвет [ править ]

Некоторые клетки делятся путем почкования (например, пекарские дрожжи ), в результате чего образуются «материнская» и «дочерняя» клетки, которые изначально меньше родительской. Почкование известно также на многоклеточном уровне; животным примером является гидра , [10] который размножается почкованием. Из почек вырастают полностью созревшие особи, которые в конечном итоге отделяются от родительского организма.
Внутреннее почкование — это процесс бесполого размножения, которому благоприятствуют такие паразиты, как Toxoplasma gondii . Он включает в себя необычный процесс, при котором внутри материнской клетки образуются две ( эндодиогения ) или более ( эндополигения ) дочерние клетки, которые затем потребляются потомством до их разделения. [11]
Кроме того, почкование (внешнее или внутреннее) происходит у некоторых червей, таких как Taenia или Echinococcus ; эти черви производят цисты, а затем производят (инвагинированные или эвагинированные) протосколексы с почкованием .
Вегетативное размножение [ править ]

Вегетативное размножение — это тип бесполого размножения, обнаруженный у растений, при котором новые особи образуются без образования семян или спор и, следовательно, без сингамии или мейоза . [12] Примеры вегетативного размножения включают образование миниатюрных растений, называемых проростками, на специализированных листьях, например, у каланхоэ ( Bryophyllum daigremontianum ), и многие из них дают новые растения из корневищ или столонов (например, у клубники ). Некоторые растения размножаются путем образования луковиц или клубней , например луковицы тюльпанов и клубни георгин . В этих примерах все особи являются клонами, а клональная популяция может охватывать большую территорию. [13]
Образование спор [ править ]
Многие многоклеточные организмы производят споры в течение своего биологического жизненного цикла в процессе, называемом спорогенезом . Исключение составляют животные и некоторые протисты, у которых сразу же происходит мейоз с последующим оплодотворением. С другой стороны, растения и многие водоросли подвергаются споровому мейозу , при котором мейоз приводит к образованию гаплоидных спор, а не гамет. Эти споры превращаются в многоклеточные особи, называемые гаметофитами , без оплодотворения. Эти гаплоидные особи производят гаметы посредством митоза . Таким образом, мейоз и образование гамет происходят в отдельных многоклеточных поколениях или «фазах» жизненного цикла, называемых чередованием поколений . Поскольку половое размножение часто более узко определяется как слияние гамет ( оплодотворение ), образование спор в спорофитах растений и водорослях можно рассматривать как форму бесполого размножения (агамогенез), несмотря на то, что оно является результатом мейоза и подвергается снижению плоидности . Однако оба события (образование спор и оплодотворение) необходимы для завершения полового размножения в жизненном цикле растения.
Грибы и некоторые водоросли также могут использовать истинное бесполое образование спор , которое включает митоз, дающий начало репродуктивным клеткам, называемым митоспорами, которые после распространения развиваются в новый организм. Этот метод размножения встречается, например, у конидиальных грибов и красных водорослей Polysiphonia и включает спорогенез без мейоза. Таким образом, число хромосом споровой клетки такое же, как у родительской клетки, производящей споры. Однако митотический спорогенез является исключением, и большинство спор, например, у растений и многих водорослей, образуются путем мейоза . [14] [15] [16]
Фрагментация [ править ]

Фрагментация — это форма бесполого размножения, при которой новый организм вырастает из фрагмента родительского. Каждый фрагмент развивается в зрелую, взрослую особь. Фрагментация наблюдается у многих организмов. К животным, размножающимся бесполым путем, относятся планарии , многие кольчатые черви, включая полихет. [17] и некоторые олигохеты , [18] турбеллярии и морские звезды . Многие грибы и растения размножаются бесполым путем. Некоторые растения имеют специализированные структуры для размножения путем фрагментации, например почки мхов , и печеночников . Большинство лишайников , которые представляют собой симбиотический союз гриба и фотосинтезирующих водорослей или цианобактерий , размножаются путем фрагментации, чтобы гарантировать, что новые особи содержат обоих симбионтов. Эти фрагменты могут иметь форму соредий — пылевидных частиц, состоящих из грибных гиф, обернутых вокруг фотобионтных клеток.
Клональная фрагментация в многоклеточных или колониальных организмах — это форма бесполого размножения или клонирования, при которой организм разделяется на фрагменты. Каждый из этих фрагментов развивается в зрелых, полностью выросших особей, которые являются клонами исходного организма. У иглокожих этот метод размножения обычно известен как расщепление . [19] Из-за множества экологических и эпигенетических различий клоны, происходящие от одного и того же предка, на самом деле могут быть генетически и эпигенетически разными. [20]
Агамогенез [ править ]
Агамогенез – это любая форма размножения, не связанная с участием мужской гаметы. Примерами являются партеногенез и апомиксис .
Партеногенез [ править ]
Партеногенез — это форма агамогенеза , при которой из неоплодотворенной яйцеклетки развивается новая особь. Оно зарегистрировано у более чем 2000 видов. [21] Партеногенез происходит в дикой природе у многих беспозвоночных (например, водяных блох, коловраток , тлей, палочников , некоторых муравьев, пчел и ос-паразитов) и позвоночных животных (в основном рептилий, амфибий и рыб). Это также было зарегистрировано у домашних птиц и у генетически измененных лабораторных мышей. [22] [23] Растения также могут участвовать в партеногенезе посредством процесса, называемого апомиксисом . Однако многие считают этот процесс не самостоятельным методом размножения, а нарушением механизмов полового размножения. [24] Партеногенетические организмы можно разделить на две основные категории: факультативные и облигатные.
Факультативный партеногенез [ править ]

При факультативном партеногенезе самки могут размножаться как половым, так и бесполым путем. [21] Из-за множества преимуществ полового размножения большинство факультативных партенот размножаются бесполым путем только в случае необходимости. Обычно это происходит в тех случаях, когда найти пару становится сложно. Например, самки акул-зебр размножаются бесполым путем, если они не могут найти себе пару в своей океанской среде обитания. [2]
Ранее считалось, что партеногенез редко встречается у позвоночных и возможен только у очень мелких животных. Однако в последние годы он был обнаружен у многих других видов. Сегодня самый крупный вид, размножающийся партеногенным путем, — это варан Комодо длиной 10 футов и весом более 300 фунтов. [25] [26]

Гетерогония — это форма факультативного партеногенеза, при которой самки через определенные промежутки времени чередуют половое и бесполое размножение (см. Чередование полового и бесполого размножения ). Тля — это одна из групп организмов, которая занимается этим типом размножения. Они используют бесполое размножение для быстрого размножения и создания крылатого потомства, которое может колонизировать новые растения и размножаться половым путем осенью, чтобы отложить яйца на следующий сезон. [27] Однако некоторые виды тли являются облигатными партенотами. [28]
Облигатный партеногенез [ править ]

При облигатном партеногенезе самки размножаются только бесполым путем. [21] Одним из примеров этого является пустынная ящерица-хлыстохвост , гибрид двух других видов. Обычно гибриды бесплодны, но благодаря партеногенезу этот вид смог создать стабильные популяции. [29]
Гиногенез — это форма облигатного партеногенеза, при которой сперматозоид используется для инициации размножения. Однако гены сперматозоида никогда не внедряются в яйцеклетку. Самым известным примером этого является амазонская молли . Поскольку они являются облигатными партенотами, в их виде нет самцов, поэтому они зависят от самцов близкородственного вида ( моллинезии Sailfin ). в получении спермы [30]
и нуцеллярный Апомиксис эмбрионий
Апомиксис у растений – это образование нового спорофита без оплодотворения. Он важен для папоротников и цветковых растений, но очень редко встречается у других семенных растений. У цветковых растений термин «апомиксис» сейчас чаще всего используется для обозначения агамоспермии , образования семян без оплодотворения, но когда-то он включал и вегетативное размножение . Примером апомиктического растения может служить триплоидный европейский одуванчик . Апомиксис в основном встречается в двух формах: при гаметофитном апомиксисе зародыш возникает из неоплодотворенной яйцеклетки внутри диплоидного зародышевого мешка, образовавшегося без завершения мейоза. При нуцеллярной эмбрионологии зародыш формируется из диплоидной ткани нуцеллуса, окружающей зародышевый мешок. Нуцеллярный зародыш встречается в некоторых семенах цитрусовых . Мужской апомиксис может возникать в редких случаях, например, у сахарского кипариса Cupressus dupreziana , где генетический материал эмбриона полностью получен из пыльцы . [31] [32] [33]
Чередование полового и бесполого размножения [ править ]
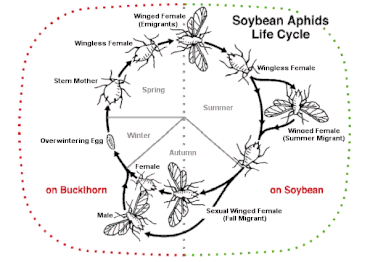
Некоторые виды могут чередовать сексуальные и бесполые стратегии (способность, известная как гетерогамия) , в зависимости от многих условий. Чередование наблюдается у некоторых видов коловраток (циклический партеногенез, например, у видов Brachionus ) и у некоторых видов насекомых.
Одним из примеров этого является тля , которая может проявлять гетерогонию. В этой системе самки рождаются беременными и производят потомство только женского пола. Этот цикл позволяет им размножаться очень быстро. Однако большинство видов размножаются половым путем один раз в год. Это переключение вызвано изменениями окружающей среды осенью и заставляет самок развивать яйца вместо эмбрионов. Этот динамичный репродуктивный цикл позволяет им производить специализированное потомство с полифенизмом — типом полиморфизма , при котором разные фенотипы развивались для выполнения определенных задач. [27]
Капская пчела Apis mellifera subsp. capensis может размножаться бесполым путем посредством процесса, называемого телитоки . Пресноводное ракообразное дафния весной размножается партеногенезом, быстро заселяя водоемы, а затем переключается на половое размножение по мере увеличения интенсивности конкуренции и хищничества. Могононтные коловратки рода Brachionus размножаются посредством циклического партеногенеза: при низкой плотности популяции самки размножаются бесполым путем, а при более высокой плотности накапливается химический сигнал, вызывающий переход к половому размножению. Многие протисты и грибы чередуют половое и бесполое размножение. Аналогичной способностью обладают несколько видов амфибий, рептилий и птиц. [ который? ] [ который? ]
Слизевик Dictyostelium при благоприятных условиях подвергается бинарному делению (митозу) в виде одноклеточных амеб. Однако, когда условия становятся неблагоприятными, клетки объединяются и следуют одному из двух разных путей развития, в зависимости от условий. На социальном пути они образуют многоклеточную слизню, которая затем образует плодовое тело со спорами, образующимися бесполым путем. В половом пути две клетки сливаются, образуя гигантскую клетку, которая развивается в большую кисту. Когда эта макроциста прорастает, она высвобождает сотни амебных клеток, которые являются продуктом мейотической рекомбинации между двумя исходными клетками. [34]
Гифы плесени обыкновенной ( Rizopus ) способны образовывать как митотические, так и мейотические споры. Многие водоросли аналогичным образом переключаются между половым и бесполым размножением. [35] Ряд растений используют как половые, так и бесполые способы для образования новых растений, некоторые виды меняют свои основные способы размножения с полового на бесполое в различных условиях окружающей среды. [36]
у видов Наследование бесполых
У коловратки Brachionus calyciflorus бесполое размножение (облигатный партеногенез ) может наследоваться по рецессивному аллелю, что приводит к утрате полового размножения у гомозиготного потомства. [37] [38]
Наследование бесполого размножения по единственному рецессивному локусу обнаружено также у паразитоидной осы Lysiphlebus fabarum . [39]
Примеры у животных [ править ]
Бесполое размножение встречается почти у половины типов животных. [40] Партеногенез происходит у акулы-молота. [41] и черноперая акула . [42] В обоих случаях акулы достигли половой зрелости в неволе в отсутствие самцов, и в обоих случаях было показано, что потомство генетически идентично матерям. хлыстохвост из Нью-Мексико Еще один пример – .
Некоторые рептилии используют систему определения пола ZW , в результате которой рождаются либо самцы (с половыми хромосомами ZZ), либо самки (с половыми хромосомами ZW или WW). До 2010 года считалось, что хромосомная система ZW, используемая рептилиями, неспособна производить жизнеспособное потомство WW, но было обнаружено, что самка удава (ZW) произвела жизнеспособное потомство женского пола с хромосомами WW. [43] Самка удава могла выбрать любое количество партнеров-самцов (и успешно это делала в прошлом), но в данном случае она размножалась бесполым путем, создав 22 детеныша женского пола с половыми хромосомами WW.
Полиэмбриония — широко распространенная форма бесполого размножения у животных, при которой оплодотворенная яйцеклетка или более поздняя стадия эмбрионального развития расщепляется с образованием генетически идентичных клонов. У животных это явление лучше всего изучено у паразитических перепончатокрылых . У девятипоясных броненосцев этот процесс обязателен и обычно приводит к образованию генетически идентичных четверок. У других млекопитающих монозиготные близнецы не имеют очевидной генетической основы, хотя встречаются часто. Сегодня в мире насчитывается по меньшей мере 10 миллионов идентичных человеческих близнецов и тройняшек.
Бделлоидные коловратки размножаются исключительно бесполым путем, причем все особи класса Bdelloidea — самки. Асексуальность развилась у этих животных миллионы лет назад и сохраняется с тех пор. Есть данные, позволяющие предположить, что бесполое размножение позволило животным выработать новые белки благодаря эффекту Мезельсона , которые позволили им лучше выживать в периоды обезвоживания. [44] Бделлоидные коловратки чрезвычайно устойчивы к повреждениям от ионизирующей радиации благодаря тем же адаптациям, сохраняющим ДНК, которые используются для выживания в состоянии покоя. [45] Эти адаптации включают чрезвычайно эффективный механизм восстановления двухцепочечных разрывов ДНК. [46] Этот механизм репарации был изучен у двух видов Bdelloidea: Adineta vaga , [46] и Филодина розеола . [47] и, по-видимому, включает митотическую рекомбинацию между гомологичными участками ДНК внутри каждого вида.
Молекулярные данные убедительно свидетельствуют о том, что некоторые виды палочников рода Timema использовали только бесполое (партеногенетическое) размножение в течение миллионов лет - самый длительный период, известный для любого насекомого. [48] Подобные результаты позволяют предположить, что клещей вид Oppiella nova мог размножаться полностью бесполым путем в течение миллионов лет. [49]
У травяных трипсов рода Aptinothrips наблюдался несколько переходов к асексуальности, вероятно, по разным причинам. [50]
значение бесполого Адаптивное размножения
Полное отсутствие полового размножения сравнительно редко встречается у многоклеточных организмов , особенно у животных . Не совсем понятно, почему среди них так распространена способность к размножению половым путем. Текущие гипотезы [51] предполагают, что бесполое размножение может иметь краткосрочные выгоды, когда важен быстрый рост населения или в стабильной среде, в то время как половое размножение дает чистое преимущество, позволяя более быстро создавать генетическое разнообразие, позволяя адаптироваться к изменяющейся окружающей среде. Ограничения развития [52] может лежать в основе того, почему немногие животные полностью отказались от полового размножения в своем жизненном цикле. Почти все бесполые способы размножения поддерживают мейоз либо в модифицированной форме, либо в качестве альтернативного пути. [53] Факультативно апомиктические растения повышают частоту сексуальности по сравнению с апомиксисом после абиотического стресса. [53] Еще одним препятствием на пути перехода от полового к бесполому размножению может быть сопутствующая потеря мейоза и защитная рекомбинационная репарация повреждений ДНК, являющаяся одной из функций мейоза. [54] [55]
См. также [ править ]
Ссылки [ править ]
- ^ Энгельштедтер, Ян (июнь 2017 г.). «Бесполый, но не клональный: эволюционные процессы в аутомиктических популяциях | Генетика» . Генетика . 206 (2): 993–1009. doi : 10.1534/genetics.116.196873 . ПМК 5499200 . ПМИД 28381586 . Проверено 21 августа 2018 г.
- ^ Jump up to: Перейти обратно: а б Даджен, Кристин Л.; Коултон, Лора; Кость, Рен; Овенден, Дженнифер Р.; Томас, Северин (16 января 2017 г.). «Переход от полового размножения к партеногенетическому у зебровой акулы» . Научные отчеты . 7 (1): 40537. Бибкод : 2017NatSR...740537D . дои : 10.1038/srep40537 . ISSN 2045-2322 . ПМЦ 5238396 . ПМИД 28091617 .
- ^ Нарра, HP; Охман, Х. (2006). «Какая польза от секса бактериям?» . Современная биология . 16 (17): 705–710 рэндов. Бибкод : 2006CBio...16.R705N . дои : 10.1016/j.cub.2006.08.024 . ПМИД 16950097 .
- ^ Джинна Брайнер (20 декабря 2006 г.). «Самка дракона Комодо рожает девственно» . www.livscience.com . Проверено 18 декабря 2022 г.
- ^ «Размножение клеток» . Британская энциклопедия .
- ^ Образовательное издательство Британника (2011). Грибы, водоросли и протисты . Издательская группа Розен. ISBN 978-1-61530-463-9 .
- ^ П. Пураник; Аша Бхат (2007). Формы и функции животных: Беспозвоночные . Саруп и сыновья. ISBN 978-81-7625-791-6 .
- ^ Маргулис, Линн ; МакХанн, Хизер И.; Олендзенски, Лотарингия (2001). Иллюстрированный словарь протоктистов: словарь водорослей, апикомплексов, инфузорий, фораминифер, микроспор, водяных плесеней, слизевиков и других протоктистов . Джонс и Бартлетт учатся. ISBN 978-0-86720-081-2 .
- ^ Ёсинори Танада; Гарри К. Кайя (1993). Патология насекомых . Профессиональное издательство Персидского залива. ISBN 978-0-12-683255-6 .
- ^ Левенгук, Антони Ван (31 декабря 1703 г.). «IV. Часть письма г-на Антони ван Левенгука, ФРС, касающаяся зеленых сорняков, растущих в воде, и некоторых животных, обнаруженных возле них». Философские труды Лондонского королевского общества . 23 (283): 1304–1311. дои : 10.1098/rstl.1702.0042 . ISSN 0261-0523 . S2CID 186209549 .
- ^ Смит, Джеймс Десмонд; Уэйклин, Дерек (1994). Введение в паразитологию животных (3-е изд.). Издательство Кембриджского университета. стр. 101–102. ISBN 978-0-521-42811-8 .
- ^ «Бесполое размножение» . Ucmp.berkeley.edu . Проверено 13 августа 2010 г.
- ^ «Празднование полевых цветов – Увядание золота – Как растут осины» . Fs.fed.us. 11 мая 2010 г. Архивировано из оригинала 23 сентября 2010 г.
- ^ «Завод». Britannica Academic , Британская энциклопедия, 15 июня 2021 г. По состоянию на 20 января 2022 г.
- ^ Кард, В. (2016). Водоросли. В MS Hill (ред.), Biology (2-е изд., Том 1, стр. 21–23). Справочник Macmillan США.
- ^ «Грибок». Britannica Academic , Британская энциклопедия, 4 октября 2018 г. По состоянию на 20 января 2022 г.
- ^ Руперт, Э.Э.; Фокс, РС; Барнс, Р.Д. (2004). «Аннелида». Зоология беспозвоночных (7-е изд.). Брукс / Коул. стр. 434–441 . ISBN 978-0-03-025982-1 .
- ^ Руперт, Э.Э.; Фокс, Р.С. и Барнс, Р.Д. (2004). «Аннелида». Зоология беспозвоночных (7-е изд.). Брукс / Коул. стр. 466–469 . ISBN 978-0-03-025982-1 .
- ^ Шилд, Хелен Нильссон; Обст, Матиас; Щит, Маттиас; Окессон, Бертил (2009). «Стволовые клетки в бесполом размножении морских беспозвоночных» . У Баруха Ринкевича; Валерия Матранга (ред.). Стволовые клетки в морских организмах . Бег. стр. 125. ISBN 978-90-481-2766-5 .
- ^ Нойхоф, Моран; Левин, Майкл; Рехави, Одед (26 августа 2016 г.). «Вертикально и горизонтально передаваемые воспоминания – стирание границ между регенерацией и наследованием у планарий» . Биология Открытая . 5 (9): 1177–1188. дои : 10.1242/bio.020149 . ПМК 5051648 . ПМИД 27565761 .
- ^ Jump up to: Перейти обратно: а б с «Партеногенез | Определение, типы и факты» . Британская энциклопедия . Проверено 3 декабря 2020 г.
- ^ Коно, Томохиро; Обата, Яёи; Ву, Кьонг; Нива, Кацутоши; Оно, Юкико; Ямамото, Юджи; Пак Ын Сон; Со, Чон Сон; Огава, Хидехико (22 апреля 2004 г.). «Рождение партеногенетических мышей, способных развиваться до взрослого возраста» . Природа . 428 (6985): 860–864. Бибкод : 2004Natur.428..860K . дои : 10.1038/nature02402 . ISSN 1476-4687 . ПМИД 15103378 . S2CID 4353479 .
- ^ Рамачандран, Р.; МакДэниел, компакт-диск (2018). «Партеногенез у птиц: обзор» . Размножение . 155 (6): Р245–Р257. дои : 10.1530/REP-17-0728 . ISSN 1741-7899 . ПМИД 29559496 .
- ^ Озиас-Акинс, Пегги; Коннер, Джоан А. (1 января 2012 г.), Альтман, Ари; Хасегава, Пол Майкл (ред.), «16 – Регулирование апомиксиса» , Биотехнология растений и сельское хозяйство , Сан-Диего: Academic Press, стр. 243–254, ISBN 978-0-12-381466-1 , получено 12 декабря 2020 г.
- ^ Ям, Филип. «Странно, но факт: вараны Комодо показывают, что «непорочное зачатие» возможно» . Научный американец . Проверено 13 декабря 2020 г.
- ^ «Комодский дракон» . Животные . 10 сентября 2010 года. Архивировано из оригинала 17 октября 2016 года . Проверено 13 декабря 2020 г.
- ^ Jump up to: Перейти обратно: а б Стерн, Дэвид Л. (24 июня 2008 г.). «Тля» . Современная биология . 18 (12): Р504–Р505. Бибкод : 2008CBio...18.R504S . дои : 10.1016/j.cub.2008.03.034 . ISSN 0960-9822 . ПМЦ 2974440 . ПМИД 18579086 .
- ^ Дедривер, Калифорния; Ле Галлик, JF; Маэо, Ф; Саймон, Джей Си; Дедривер, Ф (2013). «Генетика облигатного партеногенеза у видов тлей и ее последствия для поддержания альтернативных способов размножения» . Наследственность . 110 (1): 39–45. дои : 10.1038/hdy.2012.57 . ISSN 0018-067X . ПМЦ 3522239 . ПМИД 22990313 .
- ^ Крюс, Д.; Фицджеральд, Коннектикут (1980). « «Половое» поведение партеногенетических ящериц (Cnemidophorus)» . Труды Национальной академии наук Соединенных Штатов Америки . 77 (1): 499–502. Бибкод : 1980PNAS...77..499C . дои : 10.1073/pnas.77.1.499 . ISSN 0027-8424 . ПМЦ 348299 . ПМИД 16592761 .
- ^ Тоблер, Майкл; Шлупп, Инго (22 июня 2005 г.). «Паразиты половых и бесполых моллинезий (Poecilia, Poeciliidae, Teleostei): аргументы в пользу Красной Королевы?» . Письма по биологии . 1 (2): 166–168. дои : 10.1098/rsbl.2005.0305 . ISSN 1744-9561 . ПМК 1626213 . ПМИД 17148156 .
- ^ «Апомиксис | размножение | Британика» . www.britanica.com . Проверено 23 марта 2023 г.
- ^ Чжан, Сыци; Лян, Мэй; Ван, Нан; Сюй, Цян; Дэн, Сюсинь; Чай, Лицзюнь (1 марта 2018 г.). «Размножение древесных многолетних цитрусовых: обновленная информация о нуцеллярном эмбрионе и самонесовместимости» . Размножение растений . 31 (1): 43–57. дои : 10.1007/s00497-018-0327-4 . ISSN 2194-7961 . ПМИД 29457194 . S2CID 254022638 .
- ^ Лотси, Йоханнес Паулюс; ботаников Международной ассоциации (1907). Progressus rei botanicae = Fortschritte der Botanik = Прогресс ботаники . Полет. 2. Йена: Г. Фишер.
- ^ Мехротра, РС; Анежа, КР (декабрь 1990 г.). Введение в микологию . Нью Эйдж Интернэшнл. стр. 83 и далее. ISBN 978-81-224-0089-2 .
- ^ Коул, Кэтлин М.; Шит, Роберт Г. (1990). Биология красных водорослей . Издательство Кембриджского университета. п. 469. ИСБН 978-0-521-34301-5 .
- ^ Эдвард Г. Рики; Фахри А. Баззаз (2005). Репродуктивное распределение у растений . Академическая пресса. п. 99. ИСБН 978-0-12-088386-8 .
- ^ Штельцер, К.-П.; Шмидт, Дж.; Видлеройтер, А.; Рисс, С. (2010). «Потеря полового размножения и карликовость у мелких многоклеточных животных» . ПЛОС ОДИН . 5 (9): e12854. Бибкод : 2010PLoSO...512854S . дои : 10.1371/journal.pone.0012854 . ПМЦ 2942836 . ПМИД 20862222 .
- ^ Шойерль, Т.; Рисс, С.; Стельцер, КП (2011). «Фенотипические эффекты аллели, вызывающей облигатный партеногенез у коловратки» . Журнал наследственности . 102 (4): 409–415. doi : 10.1093/jhered/esr036 . ПМК 3113615 . ПМИД 21576287 .
- ^ Сандрок, К.; Ворбургер, К. (2011). «Однолокусное рецессивное наследование бесполого размножения у паразитоидной осы» . Курс. Биол . 21 (5): 433–7. Бибкод : 2011CBio...21..433S . дои : 10.1016/j.cub.2011.01.070 . ПМИД 21353557 . S2CID 1438839 .
- ^ Минелли, Алессандро (2009). Бесполое размножение и регенерация . Издательство Оксфордского университета. стр. 123–127. ISBN 978-0198566205 .
{{cite book}}:|work=игнорируется ( помогите ) - ^ Сэвидж, Джульетта Эйлперин (23 мая 2007 г.). «Женщины акул могут размножаться в одиночку, обнаружили исследователи» . Вашингтон Пост .
- ^ Чепмен, Д.Д.; Фирчау, Б.; Шивджи, М.С. (11 октября 2008 г.). « Непорочное зачатие» от акулы подтверждено: второй случай за всю историю» . Журнал биологии рыб . 73 (6). Sciencedaily.com: 1473–1477. дои : 10.1111/j.1095-8649.2008.02018.x .
- ^ «Удав рождает детей без отца» . Новости CBC – Технологии и наука. 3 ноября 2010 г. Проверено 20 октября 2014 г.
- ^ Пушкина-Станчева, Н.Н.; МакГи, Б.М.; Бошетти, К.; Толлетер, Д.; Чакраборти, С.; Попова А.В.; Меерсман, Ф.; МакХерел, Д.; Хинча, ДК (2007). «Функциональное расхождение бывших аллелей у древних бесполых беспозвоночных» . Наука . 318 (5848): 268–71. Бибкод : 2007Sci...318..268P . дои : 10.1126/science.1144363 . ПМИД 17932297 .
- ^ Гладышев, Евгений и Мезельсон, Мэтью (1 апреля 2008 г.). «Чрезвычайная устойчивость бделлоидных коловраток к ионизирующей радиации» . Труды Национальной академии наук . 105 (13): 5139–5144. Бибкод : 2008PNAS..105.5139G . дои : 10.1073/pnas.0800966105 . ПМК 2278216 . ПМИД 18362355 .
- ^ Jump up to: Перейти обратно: а б Хеспелс Б., Кнапен М., Хано-Мамбрес Д., Хойскин А.С., Пино Ф., ЛУКАС С., Кошул Р., Ван Донинк К. (июль 2014 г.). «Ворота к генетическому обмену? Двухцепочечные разрывы ДНК у бделлоидной коловратки Adineta vaga, подвергшейся высыханию» (PDF) . Дж. Эвол. Биол . 27 (7): 1334–45. дои : 10.1111/jeb.12326 . ПМИД 25105197 . Архивировано (PDF) из оригинала 9 октября 2022 года.
- ^ Уэлч, Дэвид Б. Марк; Уэлч, Джессика Л. Марк и Мезельсон, Мэтью (1 апреля 2008 г.). «Доказательства вырожденной тетраплоидии у бделлоидных коловраток» . Труды Национальной академии наук . 105 (13): 5145–9. Бибкод : 2008PNAS..105.5145M . дои : 10.1073/pnas.0800972105 . ПМЦ 2278229 . ПМИД 18362354 .
- ^ Швандер, Т.; и др. (2011). «Молекулярные доказательства древней асексуальности палочников Timema» (PDF) . Современная биология . 21 (13): 1129–34. Бибкод : 2011CBio...21.1129S . дои : 10.1016/j.cub.2011.05.026 . hdl : 11370/8c189a5e-f36b-4199-934c-53347c0e2131 . ПМИД 21683598 . S2CID 2053974 . Архивировано (PDF) из оригинала 9 октября 2022 года.
- ^ Йенс, Брандт, Александр Ван, Патрик Тран Блюм, Кристиан Ансельметти, Йоан Дюма, Зои Фиге, Эмерик Франсуа, Клементина М. Галтье, Николя Хеймбургер, Бастиан Джарон, Камил С. Лабедан, Марджори Мараун, Марк Паркер, Даррен Дж. Робинсон- Рехави, Марк Шефер, Ина Симион, Пауль Шой, Стефан Швандер, Таня Баст (2021). Дивергенция гаплотипов поддерживает долговременную асексуальность панцирного клеща Oppiella nova . НАТЛ АКАД НАУК. OCLC 1312207506 .
{{cite book}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ ван дер Коой, CJ; Швандер, Т. (2014). «Эволюция асексуальности с помощью различных механизмов у травяного трипса (Thysanoptera: Aptinothrips )». Эволюция . 86 (7): 1883–1893. дои : 10.1111/evo.12402 . ПМИД 24627993 . S2CID 14853526 .
- ^ Доусон, К.Дж. (октябрь 1995 г.). «Преимущество бесполого размножения: когда оно двойное?». Журнал теоретической биологии . 176 (3): 341–347. Бибкод : 1995JThBi.176..341D . дои : 10.1006/jtbi.1995.0203 .
- ^ Энгельштедтер, Дж. (ноябрь 2008 г.). «Ограничения на развитие бесполого размножения». Биоэссе . 30 (11–12): 1138–1150. doi : 10.1002/bies.20833 . ПМИД 18937362 . S2CID 5357709 .
- ^ Jump up to: Перейти обратно: а б Хёрандль, Эльвира; Хадачек, Франц (2013). «Гипотеза инициации окислительного повреждения мейоза» . Размножение растений . 26 (4): 351–367. дои : 10.1007/s00497-013-0234-7 . ПМЦ 3825497 . ПМИД 23995700 .
- ^ Бернштейн, Х.; Хопф, ФА; Мишод, RE (1987). «Молекулярная основа эволюции пола». Молекулярная генетика развития . Достижения генетики. Том. 24. С. 323–70. дои : 10.1016/s0065-2660(08)60012-7 . ISBN 9780120176243 . ПМИД 3324702 .
- ^ Avise, J. (2008) Клональность: генетика, экология и эволюция сексуального воздержания у позвоночных животных. См. стр. 22–25. Издательство Оксфордского университета. ISBN 019536967X ISBN 978-0195369670
Дальнейшее чтение [ править ]
- Авис, Дж. (2008). Клональность: генетика, экология и эволюция сексуального воздержания у позвоночных животных . Издательство Оксфордского университета. ISBN 978-0-19-536967-0 .
- Грэм, Л.; Грэм, Дж.; Уилкокс, Л. (2003). Биология растений . Река Аппер-Седл, Нью-Джерси: Pearson Education. стр. 258–259. ISBN 978-0-13-030371-4 .
- Рэйвен, штат Пенсильвания; Эверт, РФ; Эйххорн, SE (2005). Биология растений (7-е изд.). Нью-Йорк: WH Freeman and Company. ISBN 978-0-7167-6284-3 .
Внешние ссылки [ править ]
| Национальный | |
|---|---|
| Другой | |