Гидра (род)
| Гидра | |
|---|---|
 | |
| Гидра почковающаяся | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Книдарийцы |
| Сорт: | Гидрозоа |
| Заказ: | Антоатеката |
| Семья: | гидриды День, 1846 г. |
| Род: | Гидра Линней , 1758 г. [1] |
| Разновидность [1] | |
Список | |
Гидра ( / ˈh aɪ d rə / . HY - ) род мелких пресноводных гидрозоев типа Cnidaria drə — Они родом из умеренных и тропических регионов. [2] [3] Род был назван Линнеем в 1758 году в честь Гидры , многоголового мифического зверя, побежденного Гераклом , поскольку, когда у животного отрубается часть, оно восстанавливается так же, как головы мифической гидры. Биологи особенно интересуются гидрой из-за ее регенеративной способности ; они, кажется, не умирают от старости или не стареют вообще .
Морфология
[ редактировать ]
Гидра имеет трубчатое радиально-симметричное тело длиной до 10 мм (0,39 дюйма) в развернутом состоянии, закрепленное простой клейкой ножкой, известной как базальный диск. Железистые клетки базального диска выделяют липкую жидкость, которая обеспечивает его адгезивные свойства.
На свободном конце тела находится ротовое отверстие, окруженное от одного до двенадцати тонкими подвижными щупальцами . Каждое щупальце, или книда (множественное число: книды), покрыто высокоспециализированными стрекательными клетками, называемыми книдоцитами . Книдоциты содержат специализированные структуры, называемые нематоцистами , которые выглядят как миниатюрные лампочки со спиральной нитью внутри. На узком внешнем крае книдоцита находится короткий пусковой волосок, называемый книдоцилом. При контакте с добычей содержимое нематоцисты взрывается, выпуская дротообразную нить, содержащую нейротоксины , в то, что вызвало выброс. Это может парализовать добычу, особенно если будет выпущено много сотен нематоцист.
Гидра имеет два основных слоя тела, что делает ее « диплобластической ». Слои разделены мезоглеей — гелеобразным веществом. Наружный слой — это эпидермис , а внутренний слой называется гастродермой , поскольку он выстилает желудок. Клетки, составляющие эти два слоя тела, относительно просты. Гидрамацин [4] — бактерицид , недавно обнаруженный на Гидре ; он защищает внешний слой от инфекции. Одна гидра состоит из 50 000–100 000 клеток, которые состоят из трех определенных популяций стволовых клеток , которые создают множество различных типов клеток. Эти стволовые клетки постоянно обновляются в организме . [5] На теле гидр есть две важные структуры: «голова» и «нога». Когда гидру разрезают пополам, каждая половина регенерирует и превращается в маленькую гидру ; «голова» регенерирует «ногу», а «нога» регенерирует «голову». Если гидру разрезать на множество сегментов, то средние ломтики образуют и «голову», и «ногу». [6]
Дыхание и выделение происходят путем диффузии по поверхности эпидермиса , при этом более крупные экскременты выводятся через рот. [7] [8]
Нервная система
[ редактировать ]Нервная система гидры представляет собой нервную сеть , структурно простую по сравнению с более развитыми нервными системами животных. У гидры нет узнаваемого мозга или настоящих мышц . Нервные сети соединяют сенсорные фоторецепторы и сенсорные нервные клетки, расположенные в стенках тела и щупальцах.
В строении нервной сети имеется два уровня:
- уровень 1 – сенсорные клетки или внутренние клетки; и
- уровень 2 – взаимосвязанные ганглиозные клетки, синапсированные с эпителиальными или двигательными клетками.
Некоторые имеют только два слоя нейронов . [9]
Движение и передвижение
[ редактировать ]
Если гидра встревожена или атакована, щупальца могут втянуться в маленькие бутоны, а сам столб тела может втянуться в небольшую студенистую сферу. Гидры обычно реагируют одинаково независимо от направления раздражителя, и это может быть связано с простотой нервных сетей.
Гидры, как правило, ведут сидячий или сидячий образ жизни , но иногда довольно легко передвигаются, особенно во время охоты. У них есть два различных метода перемещения: «петля» и «кувырок». Они делают это, наклоняясь и прикрепляясь к субстрату ртом и щупальцами, а затем перемещая ногу, что обеспечивает обычное прикрепление, этот процесс называется петлеобразованием. При сальто тело затем наклоняется и образует новое место крепления стопы. Благодаря этому процессу «петли» или «кувырка» гидра может перемещаться на несколько дюймов (около 100 мм) за день. Гидра может также передвигаться амебоидным движением основания или отрываться от субстрата и уплывать по течению.
Размножение и жизненный цикл
[ редактировать ]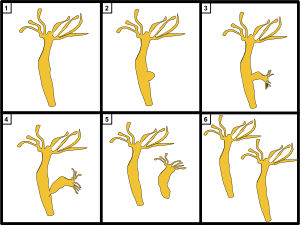
- Невоспроизводящийся
- Создание бутона
- Дочь растет
- Начало расщепления
- Дочь сломалась
- Дочерний клон родителя
У большинства видов гидр нет гендерной системы. Вместо этого, когда еды много, многие гидры размножаются бесполым путем почкованием . Почки формируются из стенки тела, вырастают в миниатюрных взрослых особей и отделяются при созревании.
Когда гидра хорошо питается, новый бутон может образовываться каждые два дня. [10] В суровых условиях, часто перед зимой или при плохих кормовых условиях, происходит половое размножение у некоторых гидр . Отеки стенок тела перерастают либо в яичники, либо в яички. Семенники выпускают в воду свободно плавающие гаметы , которые могут оплодотворить яйцеклетку в яичнике другой особи. Оплодотворенные яйца образуют прочную внешнюю оболочку, и когда взрослая особь умирает (из-за голода или холода), эти покоящиеся яйца падают на дно озера или пруда в ожидании лучших условий, после чего из них вылупляется нимфа Гидра . Некоторые гидр виды , такие как Hydracircumcincta и Hydra viridissima , являются гермафродитами. [11] и может производить как семенники, так и яичники одновременно.
Многие представители Hydrozoa претерпевают изменение тела от полипа до взрослой формы, называемой медузой , что обычно является той стадией жизни, на которой происходит половое размножение, но гидры не продвигаются дальше фазы полипа. [12]
Кормление
[ редактировать ]Гидра в основном питается водными беспозвоночными, такими как дафния и циклоп .
Во время кормления гидра вытягивает свое тело на максимальную длину, а затем медленно вытягивает щупальца. Несмотря на простоту конструкции, щупальца гидры необычайно растяжимы и могут быть в четыре-пять раз длиннее тела. После полного выпрямления щупальца медленно перемещаются в ожидании контакта с подходящей добычей. При контакте нематоцисты на щупальце стреляют в добычу, а само щупальце обвивается вокруг добычи. Большинство щупалец присоединяются к атаке в течение 30 секунд, чтобы усмирить борющуюся добычу. В течение двух минут щупальца окружают добычу и перемещают ее в открытое ротовое отверстие. В течение десяти минут добыча поглощается полостью тела, и начинается пищеварение. Гидра может значительно растягивать стенки своего тела. [ нужна ссылка ]
Пищевое поведение гидры демонстрирует сложность, казалось бы, простой нервной системы.
Некоторые виды гидры существуют во взаимоотношениях с различными видами одноклеточных водорослей . Водоросли защищены от хищников Гидрой ; в свою очередь, продукты фотосинтеза водорослей полезны в качестве источника пищи для гидры. [13] [14] и даже помогают поддерживать микробиом гидры . [15]
Измерение реакции на кормление
[ редактировать ]
Реакция на питание у гидры индуцируется глутатионом (особенно в восстановленном состоянии в виде GSH), высвобождаемым из поврежденных тканей раненой добычи. [16] Существует несколько методов, традиционно используемых для количественной оценки реакции на кормление. У некоторых измеряют продолжительность, в течение которой рот остается открытым. [17] Другие методы основаны на подсчете количества гидр среди небольшой популяции, показывающей реакцию на кормление после добавления глутатиона. [18] Недавно был разработан метод измерения реакции гидры на питание. [19] Было показано, что в этом методе линейное двумерное расстояние между кончиком щупальца и ртом гидры является прямой мерой степени пищевой реакции. Этот метод был проверен с использованием модели голодания, поскольку известно, что голодание вызывает усиление реакции гидры на кормление. [19]
Хищники
[ редактировать ]На вид Hydra oligactis охотится плоский червь Microstomum Lineare . [20] [21]
Регенерация тканей
[ редактировать ]Гидры подвергаются морфаллаксису (регенерации тканей) при ранении или разрыве. Обычно гидры размножаются путем отпрыскивания совершенно новой особи; зародыш располагается примерно на двух третях вниз по оси тела. Когда гидру разрезают пополам, каждая половина регенерирует и превращается в маленькую гидру ; «голова» регенерирует «ногу», а «нога» регенерирует «голову». Эта регенерация происходит без деления клеток. Если гидру разрезать на несколько сегментов, средние ломтики образуют и «голову», и «ногу». [6] Полярность регенерации объясняется двумя парами градиентов позиционных значений. Существует градиент активации и торможения как головы, так и ног. Активация и торможение головы работают в направлении, противоположном паре градиентов ног. [22] Доказательства этих градиентов были продемонстрированы в начале 1900-х годов в ходе экспериментов по прививке. Было показано, что ингибиторы обоих градиентов важны для блокирования образования почек. Место, где формируется бутон, находится там, где градиенты низкие как для головы, так и для ног. [6]
Гидры способны регенерировать из кусочков тканей тела и дополнительно после диссоциации тканей из агрегатов. [22] Этот процесс происходит не только в кусочках ткани, иссеченных из столба тела, но и в реагрегатах диссоциированных одиночных клеток. Установлено, что в этих агрегатах клетки, первоначально распределенные хаотично, подвергаются сортировке и образуют два слоя эпителиальных клеток, в которых более активную роль в этом процессе играют энтодермальные эпителиальные клетки. Активная подвижность этих энтодермальных эпителиальных клеток образует два слоя как в реагрегате, так и в регенерирующем кончике иссеченной ткани. Когда эти два слоя установлены, происходит процесс формирования рисунка для формирования головы и ног. [23]
Не старение
[ редактировать ]Дэниел Мартинес заявил в статье 1998 года в журнале «Экспериментальная геронтология» , что гидры бессмертны биологически . [24] Эта публикация широко цитировалась как доказательство того, что гидры не стареют (не стареют) и что они являются доказательством существования нестареющих организмов в целом. В 2010 году Престон Эстеп опубликовал (также в «Экспериментальной геронтологии ») письмо в редакцию, в котором утверждал, что данные Мартинеса опровергают гипотезу о том, что гидры не стареют. [25]
Спорная неограниченная продолжительность жизни гидры привлекла большое внимание ученых. Сегодняшние исследования, похоже, подтверждают исследование Мартинеса. [26] Стволовые клетки гидры обладают способностью к неограниченному самообновлению. Транскрипционный фактор « forkhead box O » (FoxO) был идентифицирован как критический фактор непрерывного самообновления Hydra . [26] В ходе экспериментов резкое снижение прироста популяции произошло в результате подавления FoxO . [26]
У билатерально-симметричных организмов ( Bilateria ) фактор транскрипции FoxO влияет на реакцию на стресс, продолжительность жизни и увеличение количества стволовых клеток. Если этот транскрипционный фактор сбит в двухсторонних модельных организмах, таких как плодовые мухи и нематоды , продолжительность их жизни значительно снижается. В экспериментах на H. vulgaris (радиально-симметричный представитель типа Cnidaria ), когда уровни FoxO были снижены, наблюдалось негативное влияние на многие ключевые особенности гидры , но гибели не наблюдалось, поэтому считается, что этому могут способствовать и другие факторы. очевидное отсутствие старения у этих существ. [5]
восстановление ДНК
[ редактировать ]Гидры способны к двум типам репарации ДНК : эксцизионной репарации нуклеотидов и эксцизионной репарации оснований . [27] Пути репарации облегчают репликацию ДНК, устраняя повреждения ДНК. Их идентификация у гидры была частично основана на присутствии в ее геноме генов, гомологичных генам, присутствующим у других генетически хорошо изученных видов, играющих ключевую роль в этих путях репарации ДНК. [27]
Геномика
[ редактировать ]Сравнительный анализ ортологов , проведенный за последнее десятилетие, показал, что гидра имеет как минимум 6071 общий ген с человеком. Гидра становится все более лучшей модельной системой по мере того, как становится доступным все больше генетических подходов. [5] Трансгенные гидры стали привлекательными модельными организмами для эволюции иммунитета изучения . [28] Проект генома Hydra magnipapillata был опубликован в 2010 году . [29]
геномов книдарий обычно меньше 500 МБ ( мегабаз Размер ), как у Hydra viridissima , размер генома которой составляет около 300 МБ. Напротив, геномы бурых гидр имеют размер примерно 1 Гб. Это связано с тем, что геном бурой гидры является результатом события расширения с участием LINE , типа мобильных элементов , в частности, одного семейства класса CR1. Это расширение уникально для этой подгруппы рода Hydra и отсутствует у зеленой гидры, у которой повторяющийся ландшафт, как и у других книдарий. Эти характеристики генома делают гидру привлекательной для изучения видоизменений, управляемых транспозонами, и расширения генома. [30]
Из-за простоты своего жизненного цикла по сравнению с другими гидрозойными гидры потеряли множество генов, соответствующих типам клеток или метаболическим путям, предковая функция которых до сих пор неизвестна.
Геном гидры отдает предпочтение проксимальным промоторам . Благодаря этой особенности было создано множество линий репортерных клеток с областями, расположенными примерно от 500 до 2000 оснований выше интересующего гена. Его цис-регуляторные элементы ( CRE ) в основном расположены менее чем на 2000 пар оснований выше ближайшего сайта инициации транскрипции, но есть CRE, расположенные дальше.
Его хроматин имеет конфигурацию Рабля. Существуют взаимодействия между центромерами разных хромосом и центромерами и теломерами одной и той же хромосомы. По сравнению с другими книдариями у него наблюдается большое количество межцентромерных взаимодействий, вероятно, из-за потери нескольких субъединиц конденсина II . Он организован в домены, охватывающие от десятков до сотен мегабаз, содержащие эпигенетически совместно регулируемые гены и окруженные границами, расположенными внутри гетерохроматина . [31]
Транскриптомика
[ редактировать ]Различные типы клеток гидры экспрессируют семейства генов разного эволюционного возраста. Клетки-предшественники (стволовые клетки, предшественники нейронов и нематоцист, а также половые клетки) экспрессируют гены из семейств, которые предшествовали многоклеточным животным . Среди дифференцированных клеток некоторые экспрессируют гены из семейств, происходящих от основания многоклеточных животных, например, железистые и нейрональные клетки, а другие экспрессируют гены из более новых семейств, происходящих от основания книдарий или медузозоа , например нематоцисты. Интерстициальные клетки содержат факторы трансляции, функция которых сохраняется по крайней мере 400 миллионов лет. [31]
См. также
[ редактировать ]- Лернейская гидра — греческое мифологическое водное существо, в честь которого назван род.
- Turritopsis dohrnii , еще одна книдария (медуза), которую ученые считают бессмертной.
Ссылки
[ редактировать ]- ^ Перейти обратно: а б Шухерт П. (2011). Шухерт П. (ред.). « Гидра Линней, 1758 год» . Мировая база данных Hydrozoa . Всемирный реестр морских видов . Проверено 20 декабря 2011 г.
- ^ Гилберсон Л. (1999). Руководство по зоологической лаборатории (4-е изд.). Primis Custom Publishing.
- ^ Соломон Э., Берг Л., Мартин Д. (2002). Биология (6-е изд.). Брукс/Коул .
- ^ Юнг С., Дингли А.Дж., Огюстен Р., Антон-Эркслебен Ф., Станисак М., Гельхаус С., Гутсманн Т., Хаммер М.Ю., Подщун Р., Бонвин А.М., Лейпп М., Бош Т.К., Гретцингер Дж. (январь 2009 г.). «Гидрамацин-1, структура и антибактериальная активность белка базальных многоклеточных гидр» . Журнал биологической химии . 284 (3): 1896–905. дои : 10.1074/jbc.M804713200 . ПМИД 19019828 . S2CID 3887876 .
- ^ Перейти обратно: а б с Томчик С., Фишер К., Остад С., Галлиот Б. (январь 2015 г.). «Гидра — мощная модель для изучения старения» . Размножение и развитие беспозвоночных . 59 (суп1): 11–16. Бибкод : 2015ИнвРД..59С..11Т . дои : 10.1080/07924259.2014.927805 . ПМЦ 4463768 . ПМИД 26120246 .
- ^ Перейти обратно: а б с Гилберт С.Ф. (2000). «Регенерация» . Биология развития (6-е изд.).
- ^ «Ресурсный центр Olympus по микроскопии | Видеогалерея прудовой жизни – Гидра (Coelenterata)» . olympus.magnet.fsu.edu . Проверено 21 сентября 2019 г.
- ^ Кантор, Марвин Х.; Рахат, Менахем (1982). «Регуляция дыхания и фотосинтеза у Hydra viridis и ее отдельных косимбионтов: влияние питательных веществ». Физиологическая зоология . 55 (3). Издательство Чикагского университета: 281–288. дои : 10.1086/physzool.55.3.30157891 . ISSN 0031-935X . JSTOR 30157891 . S2CID 86961916 .
- ^ Джи Н, Флавелл С.В. (апрель 2017 г.). «Гидра: визуализация нервных сетей в действии» . Современная биология . 27 (8): Р294–Р295. дои : 10.1016/j.cub.2017.03.040 . hdl : 1721.1/114954 . ПМИД 28441559 .
- ^ Паттон В.К. (август 2014 г.). «Гидра (кишечнополостная)». Мультимедийная энциклопедия Гролье . Гролье Онлайн.
- ^ Гольштейн Т., Эмшерманн П. (1995). Cnidaria: Пресноводная фауна Hydrozoa Центральной Европы. Том 1/2+ 3 . Штутгарт: Академическое издательство «Спектр». ISBN 978-3-8274-0836-5 .
- ^ Хикман, Кливленд П. младший (2019). Комплексные принципы зоологии (Восемнадцатое изд.). Нью-Йорк, штат Нью-Йорк. ISBN 978-1-260-20519-0 . OCLC 1097367369 .
{{cite book}}: CS1 maint: отсутствует местоположение издателя ( ссылка ) - ^ Торингтон, Глин; Маргулис, Линн (1981). «Hydra viridis; перенос метаболитов между гидрой и симбиотическими водорослями» . Биологический вестник . 160 (1): 175–188. дои : 10.2307/1540911 . ISSN 0006-3185 . JSTOR 1540911 . ПМИД 6164406 . S2CID 21008864 .
- ^ Маскатин, Леонард; Ленхофф, Ховард М. (15 ноября 1963 г.). «Симбиоз: о роли водорослей, симбиотических с гидрой» . Наука . 142 (3594): 956–958. Бибкод : 1963Sci...142..956M . дои : 10.1126/science.142.3594.956 . ISSN 0036-8075 . ПМИД 17753799 . S2CID 28578967 .
- ^ Батия, Джей; Шредер, Катя; Фрауне, Себастьян; Лахнит, Тим; Розенстиль, Филип; Бош, Томас К.Г. (6 июня 2022 г.). «Симбиотические водоросли Hydra viridissima играют ключевую роль в поддержании гомеостатической бактериальной колонизации» . Передний. Микробиол . 13 : 869666. дои : 10.3389/fmicb.2022.869666 . ПМЦ 9207534 . ПМИД 35733963 .
- ^ Лумис В.Ф. (октябрь 1955 г.). «Глутатионовый контроль специфических реакций питания гидры». Анналы Нью-Йоркской академии наук . 62 (9): 211–27. Бибкод : 1955NYASA..62..211L . дои : 10.1111/j.1749-6632.1955.tb35372.x . S2CID 85570550 .
- ^ Беллис С.Л., Ло, округ Колумбия, Роудс, DE (ноябрь 1994 г.). «Аффинная очистка белков, связывающих глутатион Hydra» . Письма ФЭБС . 354 (3): 320–4. дои : 10.1016/0014-5793(94)01154-0 . ПМИД 7957948 . S2CID 29262166 .
- ^ Вентурини Дж. (1987). «Рецептор гидры GSH. Фармакологические исследования и исследования связывания радиолигандов». Сравнительная биохимия и физиология. C. Сравнительная фармакология и токсикология . 87 (2): 321–4. дои : 10.1016/0742-8413(87)90015-6 . ПМИД 2888575 .
- ^ Перейти обратно: а б Кулкарни Р., Галанде С. (ноябрь 2014 г.). «Измерение реакции гидры на питание, вызванной глутатионом» . Журнал визуализированных экспериментов (93): e52178. дои : 10.3791/52178 . ПМЦ 4354099 . ПМИД 25490534 .
- ^ Кроне, Георг (2018). «Выживание органелл в чужеродном организме: нематоцисты гидры у плоского червя Microstomum Lineare» . Европейский журнал клеточной биологии . 97 (4): 289–299. дои : 10.1016/j.ejcb.2018.04.002 . ISSN 1618-1298 . ПМИД 29661512 .
- ^ Кроне, Георг (2020). «Нематоцисты гидры у плоского червя Microstomum Lineare: в поисках изменений, предшествующих их исчезновению от нового хозяина» . Исследования клеток и тканей . 379 (1): 63–71. дои : 10.1007/s00441-019-03149-w . ISSN 1432-0878 . ПМИД 31848750 . S2CID 209380951 .
- ^ Перейти обратно: а б Фудзисава Т. (февраль 2003 г.). «Регенерация гидры и эпителиопептиды» . Динамика развития . 226 (2): 182–9. дои : 10.1002/dvdy.10221 . ПМИД 12557197 . S2CID 26953455 .
- ^ Фудзисава, Тоситака (29 января 2003 г.). «Регенерация гидры и эпителиопептиды» . Динамика развития . 226 (2). Американская ассоциация анатомии ( Уайли ): 182–189. дои : 10.1002/dvdy.10221 . ISSN 1058-8388 . ПМИД 12557197 . S2CID 26953455 .
- ^ Мартинес Д.Е. (май 1998 г.). «Модель смертности предполагает отсутствие старения у гидры». Экспериментальная геронтология . 33 (3): 217–25. дои : 10.1016/S0531-5565(97)00113-7 . ПМИД 9615920 . S2CID 2009972 .
- ^ Эстеп П.В. (сентябрь 2010 г.). «Снижение бесполого размножения указывает на старение гидры: комментарий Мартинеса Д.: «Модель смертности предполагает отсутствие старения у гидры». Exp Gerontol 33, 217–25». Экспериментальная геронтология . 45 (9): 645–6. дои : 10.1016/j.exger.2010.03.017 . ПМИД 20398746 . S2CID 35408542 .
- ^ Перейти обратно: а б с Бём А.М., Халтурин К., Антон-Эркслебен Ф., Хеммрих Г., Клостермайер Ю.К., Лопес-Кинтеро Х.А., Оберг Х.Х., Пучерт М., Розенштиль П., Виттлиб Дж., Бош Т.С. (ноябрь 2012 г.). «FoxO является важнейшим регулятором поддержания стволовых клеток бессмертной гидры» . Труды Национальной академии наук Соединенных Штатов Америки . 109 (48): 19697–702. Бибкод : 2012PNAS..10919697B . дои : 10.1073/pnas.1209714109 . ПМЦ 3511741 . ПМИД 23150562 .
- ^ Перейти обратно: а б Барве, Апурва; Галанде, Алиша А.; Гаскадби, Сарой С.; Гаскадби, Сурендра (2021). «Репертуар восстановления ДНК загадочной гидры» . Границы генетики . 12 : 670695. doi : 10.3389/fgene.2021.670695 . ПМЦ 8117345 . ПМИД 33995496 .
- ^ «Центр трансгенной гидры, Кильский университет (Германия)» .
- ^ Чепмен Дж.А., Киркнесс Э.Ф., Симаков О., Хэмпсон С.Е., Митрос Т., Вайнмайер Т. и др. (март 2010 г.). «Динамический геном гидры» . Природа . 464 (7288): 592–6. Бибкод : 2010Natur.464..592C . дои : 10.1038/nature08830 . ПМЦ 4479502 . ПМИД 20228792 .
- ^ Вонг, Вайоминг; Симаков О; Мост, ДМ; Картрайт, П; Беллантуоно, Эй Джей; Кун, А; Гольштейн, ТВ; Дэвид, Китай; Стил, RE; Мартинес, Делавэр (2019). «Расширение одного семейства мобильных элементов связано с увеличением размера генома и радиацией у рода Hydra» . Proc Natl Acad Sci США . 116 (46): 22915–22917. Бибкод : 2019PNAS..11622915W . дои : 10.1073/pnas.1910106116 . ПМЦ 6859323 . ПМИД 31659034 .
- ^ Перейти обратно: а б Ж. Ф., Казе; С., Зиберт; ХМ, Литтл; П., Бертемес; А.С., Примак; П., Ладурнер; М., Ахрейнер; МТ, Фредриксен; RT, Морленд; С., Сингх; С., Чжан; ТГ, Вольфсберг; Т.Г., Шницлер; А.Д., Баксеванис; О., Симаков; Б., Хобмайер; CE, Джулиано (2023). «Эпигенетическая карта генома гидры в масштабе хромосом обнаруживает консервативные регуляторы состояния клеток» . Геномные исследования . 33 (2): 283–298. дои : 10.1101/гр.277040.122 . ПМЦ 10069465 . ПМИД 36639202 .
| Базы данных органов управления : Национальные |
|---|