Последний универсальный общий предок
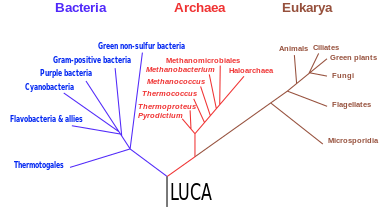
Последний универсальный общий предок ( LUCA ) — это предполагаемая общая предковая клетка , из которой произошли три домена жизни : бактерии , археи и эукарии . Предполагается, что это был «клеточный организм , который имел липидный бислой и использовал ДНК, РНК и белок». [2] LUCA также определяется как «гипотетический организм, предок всех трех доменов». [3] LUCA, вероятно, существовал не позднее 3,6 миллиарда лет назад, а возможно, еще 4,3 миллиарда лет назад или раньше. [4] Природа этой точки или стадии расхождения остается темой исследования.
Считается, что все более ранние формы жизни, предшествовавшие этому расхождению, и все существующие организмы имеют общее происхождение . На основе формального статистического теста эта теория универсального общего происхождения (UCA) подтверждается по сравнению с конкурирующими гипотезами множественного происхождения. Первый универсальный общий предок (FUCA) является гипотетическим неклеточным предком LUCA и других ныне вымерших сестринских линий.
Вопрос о том, происходит ли генезис вирусов до или после LUCA, а также разнообразие существующих вирусов и их хозяев, остается предметом исследования.
Хотя никаких ископаемых свидетельств существования LUCA не существует, детальное биохимическое сходство всей современной жизни (разделенной на три домена) делает ее существование широко признанным биохимиками. Его характеристики можно вывести из общих черт современных геномов . Эти гены описывают сложную форму жизни со многими коадаптированными функциями, включая механизмы транскрипции и трансляции для преобразования информации из ДНК в мРНК и в белки .
Историческая справка
[ редактировать ]
напрямую Филогенетическое древо отображает идею эволюции путем происхождения от одного предка . [5] Раннее древо жизни было набросано Жаном-Батистом Ламарком в его «Зоологической философии» в 1809 году. [6] [7] Чарльз Дарвин Более знаменитую теорию универсального общего происхождения в результате эволюционного процесса предложил в своей книге «Происхождение видов» в 1859 году: «Поэтому я должен сделать вывод по аналогии, что, вероятно, все органические существа, когда-либо жившие на этой Земле, произошли от некоего одна изначальная форма, в которую впервые была вдохнута жизнь». [8] Последнее предложение книги начинается с повторения гипотезы:
Есть величие в этом взгляде на жизнь, с ее несколькими силами, первоначально вдохнутыми в несколько форм или в одну...
— [8]
Термин «последний универсальный общий предок» или «LUCA» впервые был использован в 1990-х годах для обозначения такого первичного организма. [9] [10] [11]
Выводы об особенностях LUCA
[ редактировать ]Анаэробный термофил
[ редактировать ]

В 2016 году Мэдлин К. Вайс и ее коллеги генетически проанализировали 6,1 миллиона генов, кодирующих белки, и 286 514 белковых кластеров из секвенированных прокариот, геномов представляющих множество филогенетических деревьев , и идентифицировали 355 белковых кластеров, которые, вероятно, были общими для LUCA. Результаты их анализа весьма специфичны, хотя и дискуссионны. Они описывают LUCA как « анаэробный , CO 2 -фиксирующий, H 2 -зависимый с путем Вуда-Люнгдала (восстановительный путь ацетил-коэнзима А ), N 2 -фиксирующий и термофильный . Биохимия LUCA была изобилует кластерами FeS и механизмами радикальных реакций. ." [13] Кофакторы ферредоксина также обнаруживают «зависимость от переходных металлов , флавинов , S-аденозилметионина , кофермента А , метилирований , молибдоптерина , корринов и селена . Его генетический код требует нуклеозидных модификаций и S-аденозилметионин-зависимых » . [13] Они показывают, что метаногенные клостридии были базальными, около корня филогенетического дерева , в 355 исследованных белковых линиях и что, следовательно, LUCA, возможно, обитали в анаэробных гидротермальных источниках в геохимически активной среде, богатой H 2 , CO 2 и железо, где океанская вода взаимодействовала с горячей магмой под дном океана . [13] Предполагается даже, что LUCA также выросла из H 2 и CO 2 посредством обратного неполного цикла Кребса. [14] Другими метаболическими путями, предполагаемыми при LUCA, являются пентозофосфатный путь , гликолиз и глюконеогенез . [15] Даже если филогенетические данные могут указывать на среду гидротермальных жерл для термофильных LUCA, это не является доказательством того, что зарождение жизни произошло в гидротермальных жерлах, поскольку массовые вымирания могли уничтожить ранее существовавшие ветви жизни. [16]
Хотя общую анатомию LUCA можно реконструировать лишь с большой степенью неопределенности, ее биохимические механизмы можно описать довольно подробно, основываясь на «универсальных» свойствах, которые в настоящее время присущи всем независимо живущим организмам на Земле. [17]

У LUCA определенно были гены и генетический код . [12] Его генетическим материалом, скорее всего, была ДНК. [18] так что он жил после мира РНК . [а] [21] ДНК сохранялась двухцепочечной благодаря ферменту , ДНК-полимеразе который распознает структуру и направленность ДНК. [22] Целостность ДНК поддерживается группой ферментов репарации , включая ДНК-топоизомеразу . [23] Если генетический код был основан на двухцепочечной ДНК , он выражался путем копирования информации на одноцепочечную РНК. РНК вырабатывалась ДНК-зависимой РНК-полимеразой с использованием нуклеотидов, аналогичных нуклеотидам ДНК. [18] У него было несколько ДНК-связывающих белков , таких как белки гистоновой складки. [24] Генетический код был выражен в белках . Они были собраны из 20 свободных аминокислот путем трансляции информационной РНК посредством механизма рибосом , транспортных РНК и группы родственных белков. [18]
LUCA, вероятно, была способна к сексуальному взаимодействию в том смысле, что присутствовали адаптивные функции генов, которые способствовали передаче ДНК между особями популяции для облегчения генетической рекомбинации . Гомологичные генные продукты, способствующие генетической рекомбинации, присутствуют у бактерий, архей и эукариот, например, белок RecA у бактерий, белок RadA у архей и белки Rad51 и Dmc1 у эукариот. [25]
Функциональность LUCA, а также доказательства ранней эволюции мембранозависимых биологических систем позволяют предположить, что LUCA имела клеточность и клеточные мембраны. [26] Что касается общей структуры клетки, она содержала цитоплазму на водной основе , эффективно окруженную двухслойной липидной мембраной; он был способен размножаться путем деления клеток. [27] Он имел тенденцию исключать натрий и концентрировать калий с помощью специальных переносчиков ионов (или ионных насосов). Клетка размножалась путем дублирования всего своего содержимого с последующим клеточным делением . Клетка использовала хемиосмос для производства энергии. Он также восстанавливает CO 2 и окисляет 2 ( метаногенез или ацетогенез ) ацетилтиоэфиров посредством . H [28] [29]
Согласно филогенетическому брекетингу и анализу групп предполагаемых потомков LUCA, LUCA, по-видимому, представляла собой небольшой одноклеточный организм. Вероятно, у него был кольцеобразный клубок ДНК , свободно плавающий внутри клетки. Морфологически он, скорее всего, не выделялся бы среди смешанной популяции мелких современных бактерий. Создатель трехдоменной системы Карл Везе заявил, что по своему генетическому механизму LUCA была бы «более простой и рудиментарной сущностью, чем отдельные предки, породившие три [домена] (и их потомки)». [1]
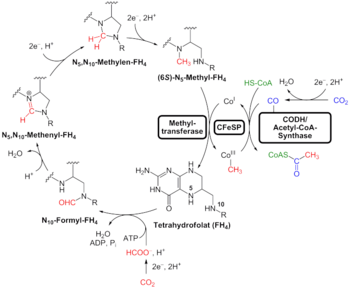
Альтернативой поиска «универсальных» признаков является использование анализа генома для выявления филогенетически древних генов. Это дает представление о LUCA, который может жить в суровых геохимически условиях и похож на современных прокариотов. Анализ биохимических путей подразумевает тот же тип химии, что и филогенетический анализ. Вайс и его коллеги пишут, что «Эксперименты... демонстрируют, что... путь ацетил-КоА [химические вещества, используемые в анаэробном дыхании] формиат , метанол , ацетильные фрагменты и даже пируват возникают спонтанно... из CO 2 , самородных металлов и воды. ", комбинация, присутствующая в гидротермальных источниках. [30]
Эксперимент показывает, что Zn 2+ , Кр 3+ , а Fe может стимулировать 6 из 11 реакций древнего анаболического пути, называемого обратным циклом Кребса, в кислых условиях, что означает, что LUCA могла обитать либо в гидротермальных источниках, либо в кислых, богатых металлами гидротермальных полях. [31]
Поскольку и бактерии, и археи имеют различия в структуре фосфолипидов и клеточной стенки, ионной перекачке, большинстве белков, участвующих в репликации ДНК и гликолизе, предполагается, что LUCA имела проницаемую мембрану без ионного насоса. Появление Na + /ЧАС + антипортеры, вероятно, приводят к эволюции непроницаемых мембран, присутствующих у эукариот, архей и бактерий. Утверждается, что «Поздняя и независимая эволюция гликолиза, но не глюконеогенеза, полностью согласуется с тем, что LUCA приводится в действие естественными градиентами протонов через дырявые мембраны. Некоторые противоречивые черты, вероятно, связаны с поздней эволюцией клеточных мембран, особенно клеточной стенки. , синтез которого зависит от мембраны и репликации ДНК». [32] Хотя у LUCA, вероятно, была ДНК, неизвестно, могла ли она реплицировать ДНК, и предполагается, что она «просто могла быть химически стабильным хранилищем для репликации на основе РНК». [33] Вполне вероятно, что проницаемая мембрана LUCA состояла из архейных липидов ( изопреноидов ) и бактериальных липидов ( жирных кислот ). Изопреноиды могли бы улучшить стабилизацию мембраны LUCA в окружающей экстремальной среде обитания. Ник Лейн и соавторы заявляют, что «преимущества и недостатки включения изопреноидов в клеточные мембраны в различных микроокружениях могли привести к дивергенции мембран, при этом более поздний биосинтез фосфолипидов привел к образованию уникальных головных групп G1P и G3P у архей и бактерий соответственно. Если это так, то свойства, придаваемые мембранными изопреноидами, относят к разделению липидов еще в зарождении жизни ». [34]
УФ-свет с длиной волны 200–280 нм (в то время не защищенный озоновым слоем) мог бы повредить нуклеотиды на поверхности, поскольку он может вызвать мутации или ошибки транскрипции и, в конечном итоге, разрушительные последствия для организмов на клеточном уровне. Однако вполне вероятно, что LUCA существовала в УФ-среде из-за преобладания фотолиазы на древе жизни. Фотолиаза использует УФ-излучение для запуска фотореактивации — механизма, который восстанавливает повреждения от радиации, вызванные УФ-излучением.
Альтернативные интерпретации
[ редактировать ]Некоторые другие исследователи оспорили выводы Вайса и др., сделанные в 2016 году. Сара Беркемер и Шон МакГлинн утверждают, что Weiss et al. выборка семейств белков была недостаточной, так что филогенетические деревья не были полными и не могли правильно описать эволюцию белков. Попытка объяснить окружающую среду LUCA почти универсальным распределением генов сопряжена с двумя рисками (как в Weiss et al., 2016). С одной стороны, это рискует ошибочно приписать события конвергенции или горизонтального переноса генов вертикальному спуску; с другой стороны, это рискует ошибочно отнести потенциальные семейства генов LUCA к событиям горизонтального переноса генов.
Филогеномный и геохимический анализ набора белков, которые, вероятно, восходят к LUCA, показывают, что он имел K + -зависимой ГТФазы, а ионный состав и концентрация ее внутриклеточной жидкости были, по-видимому, высокими К + / Что + соотношение, NH +
4 , Фе 2+ , Колорадо 2+ , Является 2+ , мг 2+ , Мн 2+ , Зн 2+ , пирофосфат и PO 3−
4 , что подразумевает наличие наземной среды обитания с горячими источниками . Возможно, у него был метаболизм, основанный на фосфатах. Кроме того, эти белки не были связаны с автотрофией (способностью организма создавать собственное органическое вещество ), что позволяет предположить, что LUCA вел гетеротрофный образ жизни (потребляя органическое вещество) и что его рост зависел от органического вещества, производимого физической средой. [35] Ник Лейн утверждает, что Na + /ЧАС + антипортеры могли бы легко объяснить низкую концентрацию Na + в LUCA и ее потомках.
Присутствие ферментов, управляющих энергией CODH / ацетил-коэнзим А- синтазы, в LUCA может быть совместимо не только с автотрофом , но и с жизнью в качестве миксотрофа или гетеротрофа . [36] Вайс и др. В 2018 году ответили, что ни один фермент не определяет трофический образ жизни и что гетеротрофы произошли от автотрофов. [37]
Доказательства того, что LUCA был мезофильным
[ редактировать ]Теперь несколько доказательств позволяют предположить, что LUCA не была термофильной.
Содержание пар нуклеотидов G+C (по сравнению с наличием пар A+T) может указывать на тепловой оптимум организма, поскольку они более термически стабильны за счет дополнительной водородной связи. В результате они чаще встречаются в рРНК термофилов; однако этого не наблюдается в реконструированной рРНК LUCA. [38] [39] [16]
Идентификация термофильных генов в LUCA подверглась критике. [40] поскольку вместо этого они могут представлять собой гены, которые позже развились у архей или бактерий, а затем мигрировали между ними посредством горизонтального переноса генов , как в гипотезе Вёзе 1998 года. [41] LUCA мог быть мезофилом, который фиксировал CO 2 и полагался на H 2 и жил недалеко от гидротермальных источников. [42]
Еще одним доказательством того, что LUCA был мезофильным, является аминокислотный состав его белков. Обилие аминокислот I, V, Y, W, R, E и L (обозначаемых IVYWREL) в белках организма коррелирует с оптимальной температурой его роста. [43] Согласно филогенетическому анализу, содержание белков LUCA в IVYWREL позволяет предположить, что идеальная температура для него была ниже 50°C. [16]
Наконец, данные о том, что бактерии и археи независимо претерпели фазы повышенной, а затем и пониженной термотолерантности, предполагают резкий климатический сдвиг после LUCA, который повлиял на обе популяции и может объяснить кажущееся генетическое распространение термотолерантной генетики. [44]
Возраст
[ редактировать ]Исследования, проведенные с 2000 по 2018 год, показали, что время существования LUCA становится все более древним. В 2000 году оценки возраста LUCA варьировались от 3,5 до 3,8 миллиардов лет назад в палеоархее . [45] за несколько сотен миллионов лет до самых ранних ископаемых свидетельств жизни , возраст кандидатов которых варьируется от 3,48 до 4,28 миллиардов лет назад. [46] [47] [48] [49] [50] Это определило возникновение первых форм жизни вскоре после поздней тяжелой бомбардировки , которая, как считалось, неоднократно стерилизовала поверхность Земли. Тем не менее, в исследовании 2018 года Холли Беттс и ее коллеги применили модель молекулярных часов к геномной и палеонтологической летописи (102 вида, 29 общих генов, кодирующих белки, в основном рибосомальные), и пришли к выводу, что LUCA предшествовала поздней тяжелой бомбардировке (что делает LUCA более 3,9). миллиардов лет назад). , которое они датировали 4,520 миллиарда лет назад, не было никакого события стерилизации Они предположили, что после события формирования Луны , и пришли к выводу, что наиболее вероятная дата LUCA находится в пределах 50 миллионов лет от этого события. [51] Исследование 2022 года показало, что возраст LUCA составляет около 3,6–4,2 миллиарда лет. [52] Исследование 2024 года показало, что LUCA жили около 4,2 миллиарда лет назад (с доверительным интервалом 4,09–4,33 миллиарда лет назад). [4]
Корень древа жизни
[ редактировать ]В 1990 году была представлена новая концепция древа жизни , разделившая живой мир на три ствола, классифицированных как домены Бактерии , Археи и Эукарии . [1] [54] [55] [2] Это первое дерево, основанное исключительно на молекулярной филогенетике и включающее эволюцию микроорганизмов. Его назвали «универсальным филогенетическим деревом в укорененной форме». [1] Это дерево и его корни стали предметом споров. [54] [б]
Тем временем были предложены многочисленные модификации этого дерева, в основном касающиеся роли и важности горизонтального переноса генов для его укоренения и ранних разветвлений (например, [57] [53] ). Поскольку наследственность происходит как вертикально, так и горизонтально, древо жизни могло быть более паутинным или сетчатым на ранней стадии и более древовидным, когда оно росло трехствольным. [53] Предположительно, горизонтальный перенос генов уменьшается с ростом стабильности клеток. [3]
Модифицированная версия дерева, основанная на нескольких молекулярных исследованиях, имеет корень между монофилетическим доменом Bacteria и кладой , образованной архей и эукариотами . [57] Небольшое меньшинство исследований помещает корень в домен бактерий типа Bacillota . [58] или заявить, что тип Chloroflexota (ранее Chloroflexi) является базальным по отношению к кладе с архей, эукариотами и остальными бактериями (как предложил Томас Кавальер-Смит ). [59] Метагеномный анализ выявил двухдоменную систему с доменами Archaea и Bacteria; с этой точки зрения на древо жизни эукариоты произошли от архей. [60] [61] [62] С более поздним генофондом потомков LUCA, разделяющим общую структуру правила AT/GC и стандартные двадцать аминокислот, горизонтальный перенос генов стал бы возможным и мог бы быть обычным явлением. [63]
Природа LUCA остается спорной. на основе первичного метаболизма (sensu Wächtershäuser ) В 1994 году Отто Кандлер предложил последовательное расхождение трёх областей жизни. [1] из мультифенотипической популяции предклеток , достигнутой путем постепенных эволюционных улучшений ( клеточности ). [64] [65] [66] Эти фенотипически разнообразные преклетки представляли собой метаболизирующиеся, самовоспроизводящиеся сущности, демонстрирующие частый взаимный обмен генетической информацией. Таким образом, в этом сценарии не было «первой ячейки». Это может объяснить единство и в то же время разделение на три линии (три области) жизни. Предклеточную теорию Кандлера поддерживает Вехтерсхойзер. [67] [68] В 1998 году Карл Везе , основываясь на концепции мира РНК, предположил, что ни один отдельный организм не может считаться LUCA, и что генетическое наследие всех современных организмов получено в результате горизонтального переноса генов между древним сообществом организмов. [69] Другие авторы сходятся во мнении, что существовал «сложный коллективный геном». [70] во времена LUCA, и что горизонтальный перенос генов был важен в эволюции более поздних групп; [70] Николас Глансдорфф утверждает, что LUCA «находилась в метаболически и морфологически гетерогенном сообществе, постоянно перемещаясь вокруг генетического материала» и «оставалась эволюционной сущностью, хотя и слабо определенной и постоянно меняющейся, пока продолжалась эта распущенность». [71]
Широко распространена теория универсального общего происхождения жизни. В 2010 году на основе «огромного множества молекулярных последовательностей, доступных сейчас из всех областей жизни», [72] Д. Л. Теобальд опубликовал « формальный тест » универсального общего происхождения (UCA). Речь идет об общем происхождении всех существующих наземных организмов, каждый из которых является генеалогическим потомком одного вида из далекого прошлого. Его формальный тест предпочитал существование универсального общего происхождения широкому классу альтернативных гипотез, включая горизонтальный перенос генов. Основные биохимические принципы предполагают, что все организмы имеют общее происхождение. [73]
Предполагаемый ранее неклеточный предок LUCA - это Первый универсальный общий предок (FUCA). [74] [75] Таким образом, FUCA будет предком каждой современной клетки, а также древних, ныне вымерших клеточных линий, не являющихся потомками LUCA. Предполагается, что у FUCA были и другие потомки, кроме LUCA, ни у кого из которых нет современных потомков. Считается, что некоторые гены этих древних, ныне вымерших клеточных линий были горизонтально перенесены в геном ранних потомков LUCA. [63]
LUCA и вирусы
[ редактировать ]Происхождение вирусов остается спорным. Поскольку для репликации вирусам необходимы клетки-хозяева, вполне вероятно, что они возникли после образования клеток . Вирусы могут даже иметь множественное происхождение, и разные типы вирусов могли развиваться независимо на протяжении истории жизни. [2] Существуют разные гипотезы происхождения вирусов, например, раннее вирусное происхождение из мира РНК или более позднее вирусное происхождение из эгоистичной ДНК . [2]
Судя по тому, как вирусы в настоящее время распределяются среди бактерий и архей , можно предположить, что LUCA стала жертвой нескольких вирусов, предков тех, которые теперь имеют эти два домена в качестве своих хозяев. [76] Более того, LUCA, по-видимому, предшествовала обширная эволюция вирусов, поскольку желеобразная структура белков капсида является общей для РНК- и ДНК-вирусов во всех трех сферах жизни. [77] [78] Вирусы LUCA, вероятно, представляли собой в основном вирусы с дцДНК из групп, называемых дуплоднавирией и вариднавирией . Две другие группы вирусов с одноцепочечной ДНК внутри Monodnaviria , Microviridae и Tubulavirales , вероятно, заразили последнего общего предка бактерий. Последний общий предок архей, вероятно, был носителем веретенообразных вирусов. Все это вполне могло повлиять на LUCA, и в этом случае каждый из них должен был быть потерян в хост-домене, где он больше не существует. Напротив, РНК-вирусы, по-видимому, не были важными паразитами LUCA, хотя при простом мышлении можно было бы предположить, что вирусы начинаются с РНК-вирусов, непосредственно происходящих из мира РНК. Вместо этого ко времени существования LUCA РНК-вирусы, вероятно, уже вытеснили ДНК-вирусы. [76]
LUCA могла быть предком некоторых вирусов, поскольку у нее могло быть как минимум два потомка: LUCELLA, последний универсальный клеточный предок, предок всех клеток, и архаичный предок вироклеток, предок крупных и средних клеток. ДНК-вирусы . [79] Вирусы могли появиться до LUCA, но после Первого универсального общего предка (FUCA), согласно гипотезе редукции, согласно которой гигантские вирусы развились из первичных клеток, которые стали паразитическими . [63]
См. также
[ редактировать ]- Абиогенез – жизнь, возникающая из неживой материи.
- Целлюляризация - научная теория, объясняющая происхождение и формирование клеток.
- Хемотон - Абстрактная модель фундаментальной единицы жизни.
- Дарвиновский порог - период эволюции первых клеток.
- Последний общий предок эукариот - процесс формирования первой эукариотической клетки.
- Митохондриальная Ева - самый недавний общий предок всех живых людей по материнской линии.
- Предклетка - Гипотетическая жизнь до появления полных клеток
- Протометаболизм
- Хронология эволюционной истории жизни
- Urmetazoan - Гипотетический последний общий предок всех животных.
- Y-хромосомный Адам - самый недавний общий предок всех живых людей по отцовской линии.
Примечания
[ редактировать ]- ^ Другие исследования предполагают, что LUCA могла быть полностью определена через РНК , [19] состоял из гибридного генома РНК-ДНК или обладал ретровирусоподобным генетическим циклом, в котором ДНК служила стабильным генетическим хранилищем. [20]
- ^ Одна дискуссия касалась бывшей кладистической гипотезы: дереву нельзя было приписать корень обычным алгоритмическим способом, потому что для этого потребовалась бы внешняя группа для ссылки. В случае универсального дерева никакой внешней группы не существовало бы. Кладистический метод использовался, «например, для того, чтобы укоренить пурпурные бактерии. Но установление корня вселенского древа жизни, порядка ветвления среди первичных королевств, было совсем другим делом». [56]
Ссылки
[ редактировать ]- ^ Перейти обратно: а б с д и Вёзе, ЧР ; Кандлер, О .; Уилис, ML (июнь 1990 г.). «На пути к естественной системе организмов: предложение по доменам архей, бактерий и эукариев» . ПНАС . 87 (12): 4576–4579. Бибкод : 1990PNAS...87.4576W . дои : 10.1073/pnas.87.12.4576 . ПМК 54159 . ПМИД 2112744 .
- ^ Перейти обратно: а б с д Мэдиган, Майкл Т.; Айер, Дженнифер; Бакли, Дэниел Х.; Сэттли, Мэтью; Шталь, Дэвид А. (2022). Брок Биология микроорганизмов (16-е изд.). Харлоу: Образование Пирсона. стр. Раздел 3, глава 13: 431 (LUCA), 435 (дерево жизни), 428, 438, 439 (вирусы). ISBN 978-1-292-40479-0 .
- ^ Перейти обратно: а б Гарольд, Франклин М. (2014). В поисках клеточной истории: эволюция строительных блоков жизни . Чикаго, Лондон: Издательство Чикагского университета. ISBN 978-0-226-17428-0 . Архивировано из оригинала 31 октября 2023 года . Проверено 12 октября 2023 г.
- ^ Перейти обратно: а б Муди, Эдмунд Р.Р.; Альварес-Карретеро, Сандра; Махендрараджа, Тара А.; Кларк, Джеймс В.; Беттс, Холли С.; Домбровский, Нина; Санто, Ленард Л.; Бойл, Ричард А.; Дэйнс, Стюарт; Чен, Си; Лейн, Ник; Ян, Цзыхэн; Шилдс, Грэм А.; Сёллёси, Гергеи Дж.; Спанг, Аня (12 июля 2024 г.). «Природа последнего универсального общего предка и его влияние на раннюю систему Земли» . Экология природы и эволюция : 1–13. дои : 10.1038/s41559-024-02461-1 . ISSN 2397-334X .
- ^ Грегори, Т. Райан (2008). «Понимание эволюционных деревьев» . Эволюция: образование и информационно-пропагандистская деятельность . 1 (2): 121–137. дои : 10.1007/s12052-008-0035-x . S2CID 15488906 .
- ^ Ламарк, Жан Батист Пьер Антуан де Моне де (1994) [1809]. Зоологическая философия (PDF) . Париж. п. 737.
{{cite book}}: CS1 maint: отсутствует местоположение издателя ( ссылка ) - ^ Ноубл, Денис (1 июля 2020 г.). «Редакционная статья: Чарльз Дарвин, Жан-Батист Ламарк и аргументы XXI века об основах биологии» . Прогресс биофизики и молекулярной биологии . 153 : 1–4. doi : 10.1016/j.pbiomolbio.2020.02.005 . ПМИД 32092299 . S2CID 211475380 . Архивировано из оригинала 1 марта 2022 года . Проверено 23 декабря 2022 г.
- ^ Перейти обратно: а б Дарвин, Чарльз (1859). Происхождение видов путем естественного отбора . Джон Мюррей . стр. 484, 490. Архивировано из оригинала 8 октября 2022 года . Проверено 8 октября 2022 г.
- ^ Уикхэм, Джин Стивен (март 1995 г.). Молекулярно-филогенетический анализ природных гипертермофильных микробных сообществ (кандидатская диссертация). Университет Индианы . п. 4. ПроКвест 304192982
- ^ Фортерре, Патрик (1997). «Археи: чему мы можем научиться из их последовательностей?». Текущее мнение в области генетики и развития . 7 (6): 764–770. дои : 10.1016/s0959-437x(97)80038-x . ПМИД 9468785 .
- ^ Кунин Евгений Владимирович ; Гальперин, Михаил Ю. (2003). Последовательность – Эволюция – Функция: Вычислительные подходы в сравнительной геномике . Бостон, Массачусетс: Клувер. п. 252. ИСБН 978-1-4757-3783-7 . OCLC 55642057 .
- ^ Перейти обратно: а б с Вайс, Мэдлин С.; Прейнер, Мартина; Ксавье, Джоана С.; Зиморски, Верена; Мартин, Уильям Ф. (16 августа 2018 г.). «Последний универсальный предок между древней земной химией и зарождением генетики» . ПЛОС Генетика . 14 (8): e1007518. дои : 10.1371/journal.pgen.1007518 . ПМК 6095482 . ПМИД 30114187 . S2CID 52019935 .
- ^ Перейти обратно: а б с Вайс, Мэдлин С.; Соуза, Флорида; Мрнявац, Н.; и др. (2016). «Физиология и среда обитания последнего всеобщего общего предка» (PDF) . Природная микробиология . 1 (9): 16116. doi : 10.1038/nmicrobiol.2016.116 . ПМИД 27562259 . S2CID 2997255 . Архивировано (PDF) из оригинала 18 апреля 2022 года . Проверено 10 октября 2022 г.
- ^ Харрисон, Стюарт А.; Палмейра, Ракель Нуньес; Халперн, Аарон; Лейн, Ник (1 ноября 2022 г.). «Биофизическая основа возникновения генетического кода в протоклетках» . Biochimica et Biophysica Acta (BBA) — Биоэнергетика . 1863 (8): 148597. doi : 10.1016/j.bbabio.2022.148597 . ISSN 0005-2728 . PMID 35868450 .
- ^ Харрисон, Стюарт А.; Лейн, Ник (12 декабря 2018 г.). «Жизнь как руководство к синтезу пребиотических нуклеотидов» . Природные коммуникации . 9 (1): 5176. Бибкод : 2018NatCo...9.5176H . дои : 10.1038/s41467-018-07220-y . ISSN 2041-1723 . ПМК 6289992 . ПМИД 30538225 .
- ^ Перейти обратно: а б с Кантин, Марджори Д.; Фурнье, Грегори П. (6 июля 2017 г.). «Адаптация окружающей среды от зарождения жизни до последнего вселенского общего предка» . Происхождение жизни и эволюция биосфер . 48 (1): 35–54. дои : 10.1007/s11084-017-9542-5 . hdl : 1721.1/114219 . ISSN 0169-6149 . ПМИД 28685374 . S2CID 254888920 . Архивировано из оригинала 23 февраля 2024 года . Проверено 4 декабря 2023 г.
- ^ Вехтерсхойзер, Гюнтер (1998). «На пути к реконструкции геномов предков путем выравнивания кластеров генов». Систематическая и прикладная микробиология . 21 (4): 473–474, IN1, 475–477. Бибкод : 1998SyApM..21N1475W . дои : 10.1016/S0723-2020(98)80058-1 .
- ^ Перейти обратно: а б с Вехтерсхойзер, Гюнтер (1998). «На пути к реконструкции геномов предков путем выравнивания кластеров генов». Систематическая и прикладная микробиология . 21 (4): 473–474, IN1, 475–477. Бибкод : 1998SyApM..21N1475W . дои : 10.1016/S0723-2020(98)80058-1 .
- ^ Маршалл, Майкл. «Жизнь началась с планетарного мегаорганизма» . Новый учёный . Архивировано из оригинала 25 июля 2016 года . Проверено 31 июля 2016 г.
- ^ Кунин Евгений Владимирович; Мартин, Уильям (1 декабря 2005 г.). «О происхождении геномов и клеток в неорганических компартментах» . Тенденции в генетике . 21 (12): 647–654. дои : 10.1016/j.tig.2005.09.006 . ПМЦ 7172762 . ПМИД 16223546 .
- ^ Гарвуд, Рассел Дж. (2012). «Закономерности в палеонтологии: первые 3 миллиарда лет эволюции» . Палеонтология онлайн . 2 (11): 1–14. Архивировано из оригинала 26 июня 2015 года . Проверено 25 июня 2015 г.
- ^ Кунин Евгений Владимирович ; Крупович, М.; Ишино, С.; Ишино, Ю. (2020). «Механизм репликации LUCA: общий источник репликации и транскрипции ДНК» . БМК Биология . 18 (1): 61. дои : 10.1186/s12915-020-00800-9 . ПМЦ 7281927 . ПМИД 32517760 .
- ^ Ахмад, Музаммил; Сюй, Дунъи; Ван, Вэйдун (23 мая 2017 г.). «Топоизомеразы типа IA могут быть «волшебниками» как для ДНК, так и для РНК во всех сферах жизни» . Биология РНК . 14 (7): 854–864. дои : 10.1080/15476286.2017.1330741 . ПМЦ 5546716 . ПМИД 28534707 .
- ^ Лупас, Андрей Н.; Альва, Викрам (2018). «Гистоны появились еще до раскола между бактериями и архей». Биоинформатика . 35 (14): 2349–2353. doi : 10.1093/биоинформатика/bty1000 . ПМИД 30520969 .
- ^ Бернштейн, Х., Бернштейн, К. (2017). Сексуальное общение у архей, предшественников эукариотического мейоза. В: Вицани, Г. (ред.) Биокоммуникация архей. Спрингер, Чам. https://doi.org/10.1007/978-3-319-65536-9_7 Архивировано 23 февраля 2024 г. в Wayback Machine.
- ^ Гогартен, Иоганн Петер; Таиз, Линкольн (1992). «Эволюция АТФаз, перекачивающих протоны: укоренение древа жизни» . Исследования фотосинтеза . 33 (2): 137–146. Бибкод : 1992PhoRe..33..137G . дои : 10.1007/bf00039176 . ISSN 0166-8595 . ПМИД 24408574 . S2CID 20013957 . Архивировано из оригинала 23 февраля 2024 года . Проверено 4 декабря 2023 г.
- ^ Вехтерсхойзер, Гюнтер (1998). «На пути к реконструкции геномов предков путем выравнивания кластеров генов». Систематическая и прикладная микробиология . 21 (4): 473–474, IN1, 475–477. Бибкод : 1998SyApM..21N1475W . дои : 10.1016/S0723-2020(98)80058-1 .
- ^ Мартин, В.; Рассел, MJ (октябрь 2007 г.). «О происхождении биохимии на щелочно-гидротермальном источнике» . Философские труды Лондонского королевского общества. Серия Б, Биологические науки . 362 (1486): 1887–1925. дои : 10.1098/rstb.2006.1881 . ПМЦ 2442388 . ПМИД 17255002 .
- ^ Лейн, Ник ; Аллен, Дж. Ф.; Мартин, Уильям (апрель 2010 г.). «Как LUCA зарабатывала на жизнь? Хемиосмос в зарождении жизни». Биоэссе . 32 (4): 271–280. дои : 10.1002/bies.200900131 . ПМИД 20108228 .
- ^ Вайс, Мэдлин С.; Прейнер, Мартина; Ксавье, Джоана С.; Зиморски, Верена; Мартин, Уильям Ф. (16 августа 2018 г.). «Последний универсальный предок между древней земной химией и зарождением генетики» . ПЛОС Генетика . 14 (8): e1007518. дои : 10.1371/journal.pgen.1007518 . ПМК 6095482 . ПМИД 30114187 . S2CID 52019935 .
- ^ Муховска, Камила Б.; Варма, Шриджит Дж.; Шевалло-Беру, Элоди; Летюилье-Карл, Лукас; Ли, Гуан; Моран, Джозеф (2 октября 2017 г.). «Металлы способствуют последовательности обратного цикла Кребса» . Экология и эволюция природы . 1 (11): 1716–1721. Бибкод : 2017NatEE...1.1716M . дои : 10.1038/s41559-017-0311-7 . ISSN 2397-334X . ПМЦ 5659384 . ПМИД 28970480 .
- ^ Соджо, Виктор; Помянковский, Эндрю; Лейн, Ник (12 августа 2014 г.). «Биоэнергетическая основа дивергенции мембран архей и бактерий» . ПЛОС Биология . 12 (8): e1001926. дои : 10.1371/journal.pbio.1001926 . ISSN 1545-7885 . ПМК 4130499 . ПМИД 25116890 .
- ^ Вайс, Мэдлин С.; Прейнер, Мартина; Ксавье, Джоана С.; Зиморски, Верена; Мартин, Уильям Ф. (16 августа 2018 г.). «Последний универсальный предок между древней земной химией и зарождением генетики» . ПЛОС Генетика . 14 (8): e1007518. дои : 10.1371/journal.pgen.1007518 . ПМК 6095482 . ПМИД 30114187 . S2CID 52019935 .
- ^ Джордан, Сан-Франциско; Ни, Э.; Лейн, Ник (18 октября 2019 г.). «Изопреноиды повышают стабильность мембран жирных кислот при зарождении жизни, что потенциально может привести к раннему разделению липидов» . Фокус на интерфейсе . 9 (6). дои : 10.1098/rsfs.2019.0067 . ISSN 2042-8901 . ПМК 6802135 . ПМИД 31641436 .
 В эту статью включен текст из этого источника, доступного по лицензии CC BY 4.0 .
В эту статью включен текст из этого источника, доступного по лицензии CC BY 4.0 . - ^ Мулкиджанян Армен Ю.; Бычков Андрей Ю; Диброва Дарья Викторовна; Гальперин, Михаил Юрьевич; Кунин, Евгений В. (2012). «Происхождение первых клеток на земных бескислородных геотермальных полях» . Труды Национальной академии наук . 109 (14): Е821-30. Бибкод : 2012PNAS..109E.821M . дои : 10.1073/pnas.1117774109 . ПМЦ 3325685 . ПМИД 22331915 .
- ^ Адам, Панайотис С.; Боррель, Гийом; Грибальдо, Симонетта (6 февраля 2018 г.). «Эволюционная история дегидрогеназы монооксида углерода / ацетил-КоА-синтазы, одного из старейших ферментативных комплексов» . ПНАС . 115 (6): E1166–E1173. Бибкод : 2018PNAS..115E1166A . дои : 10.1073/pnas.1716667115 . ПМК 5819426 . ПМИД 29358391 .
- ^ Вайс, Мэдлин С.; Прейнер, Мартина; Ксавье, Джоана С.; Зиморски, Верена; Мартин, Уильям Ф. (16 августа 2018 г.). «Последний универсальный предок между древней земной химией и зарождением генетики» . ПЛОС Генетика . 14 (8): e1007518. дои : 10.1371/journal.pgen.1007518 . ПМК 6095482 . ПМИД 30114187 . S2CID 52019935 .
- ^ Галтье, Николя; Турасс, Николя; Гуи, Маноло (8 января 1999 г.). «Негипертермофильный общий предок существующих форм жизни» . Наука . 283 (5399): 220–221. дои : 10.1126/science.283.5399.220 . ISSN 0036-8075 . ПМИД 9880254 . Архивировано из оригинала 23 февраля 2024 года . Проверено 4 декабря 2023 г.
- ^ Груссен, Матье; Буссау, Бастьен; Чарльз, Сандрин; Бланкар, Самуэль; Гуи, Маноло (23 октября 2013 г.). «Молекулярный сигнал адаптации к холодной температуре на ранних этапах жизни на Земле» . Письма по биологии . 9 (5): 20130608. doi : 10.1098/rsbl.2013.0608 . ISSN 1744-9561 . ПМЦ 3971708 . ПМИД 24046876 .
- ^ Гогартен, Иоганн Петер; Димер, Дэвид (2016). «Является ли LUCA термофильным предшественником?» . Природная микробиология . 1 (12): 16229. doi : 10.1038/nmicrobiol.2016.229 . ПМИД 27886195 . S2CID 205428194 . Архивировано из оригинала 3 апреля 2020 года . Проверено 25 июня 2019 г.
- ^ Вёзе, Карл (июнь 1998 г.). «Всемирный предок» . ПНАС . 95 (12): 6854–6859. Бибкод : 1998PNAS...95.6854W . дои : 10.1073/pnas.95.12.6854 . ПМК 22660 . ПМИД 9618502 .
- ^ Кампруби, Э.; де Леу, JW; Дом, Швейцария; Раулин, Ф.; Рассел, MJ; Спанг, А.; Тирумалай, MR; Уэстолл, Ф. (12 декабря 2019 г.). «Появление жизни» . Обзоры космической науки . 215 (8): 56. Бибкод : 2019ССРв..215...56С . дои : 10.1007/s11214-019-0624-8 . ISSN 1572-9672 .
- ^ Зельдович Константин Б; Березовский, Игорь Н; Шахнович, Евгений I (2007). «Детерминанты термофильной адаптации белков и ДНК» . PLOS Вычислительная биология . 3 (1): e5. arXiv : q-bio/0607004 . Бибкод : 2007PLSCB...3....5Z . дои : 10.1371/journal.pcbi.0030005 . ISSN 1553-7358 . ПМК 1769408 . ПМИД 17222055 .
- ^ Буссау, Бастьен; Бланкар, Самуэль; Нексулея, Анамария; Лартильо, Николя; Гуи, Маноло (26 ноября 2008 г.). «Параллельные адаптации к высоким температурам в архейском эоне» . Природа . 456 (7224): 942–945. Бибкод : 2008Natur.456..942B . дои : 10.1038/nature07393 . ISSN 0028-0836 . ПМИД 19037246 . S2CID 4348746 . Архивировано из оригинала 23 февраля 2024 года . Проверено 4 декабря 2023 г.
- ^ Дулитл, WF (февраль 2000 г.). «Искоренение древа жизни». Научный американец . 282 (2): 90–95. Бибкод : 2000SciAm.282b..90D . doi : 10.1038/scientificamerican0200-90 . JSTOR 26058605 . ПМИД 10710791 .
- ^ Ноффке, Н. ; Кристиан, Д.; Уэйси, Д.; Хазен, Р.М. (декабрь 2013 г.). «Микробно-индуцированные осадочные структуры, фиксирующие древнюю экосистему формации Дрессер возрастом около 3,48 миллиардов лет, Пилбара, Западная Австралия» . Астробиология . 13 (12): 1103–1124. Бибкод : 2013AsBio..13.1103N . дои : 10.1089/ast.2013.1030 . ПМК 3870916 . ПМИД 24205812 .
- ^ Отомо, Йоко; Какегава, Такеши; Исида, Акизуми; Нагасе, Тоширо; Розинг, Миник Т. (2013). «Свидетельства наличия биогенного графита в метаосадочных породах раннего архея Исуа». Природа Геонауки . 7 (1): 25–28. Бибкод : 2014NatGe...7...25O . дои : 10.1038/ngeo2025 .
- ^ Хасенкам, Т.; Андерссон, член парламента; Далби, КНЦ; и др. (2017). «Элементы эоархейской жизни, заключенные в минеральных включениях». Природа . 548 (7665): 78–81. Бибкод : 2017Natur.548...78H . дои : 10.1038/nature23261 . ПМИД 28738409 . S2CID 205257931 .
- ^ Белл, Элизабет А.; Бенке, Патрик; Харрисон, Т. Марк; Мао, Венди Л. (24 ноября 2015 г.). «Потенциально биогенный углерод сохранился в цирконе возрастом 4,1 миллиарда лет» . ПНАС . 112 (47): 14518–14521. Бибкод : 2015PNAS..11214518B . дои : 10.1073/pnas.1517557112 . ПМЦ 4664351 . ПМИД 26483481 .
- ^ Додд, Мэтью С.; Папино, Доминик; Гренн, Тор; и др. (2 марта 2017 г.). «Доказательства ранней жизни в осадках старейших гидротермальных источников Земли» (PDF) . Природа . 543 (7643): 60–64. Бибкод : 2017Natur.543...60D . дои : 10.1038/nature21377 . ПМИД 28252057 . S2CID 2420384 . Архивировано (PDF) из оригинала 23 июля 2018 года . Проверено 25 июня 2019 г.
- ^ Беттс, Холли С.; Путтик, Марк Н.; Кларк, Джеймс В.; Уильямс, Том А.; Донохью, Филип CJ; Пизани, Давиде (2018). «Комплексные геномные и ископаемые данные проливают свет на раннюю эволюцию жизни и происхождение эукариот» . Экология и эволюция природы . 2 (10): 1556–1562. Бибкод : 2018NatEE...2.1556B . дои : 10.1038/s41559-018-0644-x . ПМК 6152910 . ПМИД 30127539 .
- ^ Муди, Эдмунд Р.Р.; Махендрараджа, Тара А; Домбровский, Нина; Кларк, Джеймс В.; Петижан, Селин; Оффре, Пьер; Сёллёси, Гергеи Дж; Спанг, Аня; Уильямс, Том А. (22 февраля 2022 г.). «Оценка самых глубоких ветвей древа жизни на основе древних вертикально развивающихся генов» . электронная жизнь . 11 . doi : 10.7554/eLife.66695 . ISSN 2050-084X . ПМЦ 8890751 . ПМИД 35190025 .
- ^ Перейти обратно: а б с Сметс, Барт Ф.; Баркай, Тамар (сентябрь 2005 г.). «Горизонтальный перенос генов: перспективы на стыке научных дисциплин» . Обзоры природы Микробиология . 3 (9): 675–678. дои : 10.1038/nrmicro1253 . ПМИД 16145755 . S2CID 2265315 .
- ^ Перейти обратно: а б Сапп, Ян А. (2009). Новые основы эволюции: на древе жизни . Нью-Йорк: Издательство Оксфордского университета. стр. Глава 19: 257 и далее (новая концепция древа жизни), главы 17–21 плюс заключительные замечания: 226–318 (обсуждение дерева и его укоренения), 286 и далее (LUCA). ISBN 978-0-199-73438-2 . Архивировано из оригинала 6 ноября 2023 года . Проверено 21 ноября 2023 г.
- ^ Мэдиган, Майкл Т.; Мартинко, Джон М.; Бендер, Келли С.; Бакли, Дэниел Х.; Шталь, Дэвид А. (2015). Брок Биология микроорганизмов (14-е изд.). Бостон: Pearson Education Limited. стр. 29, 374, 381. ISBN. 978-1-292-01831-7 .
- ^ Сапп 2009 , с. 255.
- ^ Перейти обратно: а б Браун, младший; Дулиттл, WF (1995). «Корень вселенского древа жизни, основанный на дупликации генов древней аминоацил-тРНК-синтетазы» . ПНАС . 92 (7): 2441–2445. Бибкод : 1995PNAS...92.2441B . дои : 10.1073/pnas.92.7.2441 . ПМК 42233 . ПМИД 7708661 .
- ^ Валас, Р.Э.; Борн, ЧП (2011). «Происхождение производного суперцарства: как грамположительная бактерия пересекла пустыню и стала археем» . Биология Директ . 6:16 . дои : 10.1186/1745-6150-6-16 . ПМК 3056875 . ПМИД 21356104 .
- ^ Кавальер-Смит, Том (2006). «Укоренение древа жизни посредством анализа переходов» . Биология Директ . 1:19 . дои : 10.1186/1745-6150-1-19 . ПМК 1586193 . ПМИД 16834776 .
- ^ Рэйманн, Кэйси; Брошье-Армане, Селин; Грибальдо, Симонетта (26 мая 2015 г.). «Двудоменное древо жизни связано с новым корнем архей» . Труды Национальной академии наук . 112 (21): 6670–6675. Бибкод : 2015PNAS..112.6670R . дои : 10.1073/pnas.1420858112 . ISSN 0027-8424 . ПМК 4450401 . ПМИД 25964353 .
- ^ Обнимаю, Лаура А.; Бейкер, Бретт Дж.; Анантараман, Картик; и др. (11 апреля 2016 г.). «Новый взгляд на древо жизни» . Природная микробиология . 1 (5): 16048. doi : 10.1038/nmicrobiol.2016.48 . ISSN 2058-5276 . ПМИД 27572647 . S2CID 3833474 .
- ^ Уильямс, Том А.; Фостер, Питер Г.; Кокс, Саймон Дж.; Эмбли, Т. Мартин (11 декабря 2013 г.). «Архейное происхождение эукариот поддерживает только две основные области жизни» . Природа . 504 (7479): 231–236. Бибкод : 2013Natur.504..231W . дои : 10.1038/nature12779 . ISSN 1476-4687 . ПМИД 24336283 . S2CID 4461775 . Архивировано из оригинала 1 октября 2022 года . Проверено 23 сентября 2022 г.
- ^ Перейти обратно: а б с Харрис, Хью МБ; Хилл, Колин (2021). «Место вирусам на древе жизни» . Границы микробиологии . 11 . дои : 10.3389/fmicb.2020.604048 . ISSN 1664-302X . ПМЦ 7840587 . ПМИД 33519747 .
- ^ Кандлер, Отто (1994). «Раннее разнообразие жизни». В Стефане Бенгтсоне (ред.). Ранняя жизнь на Земле. Нобелевский симпозиум 84 . Нью-Йорк: Издательство Колумбийского университета. стр. 152–160.
- ^ Кандлер, Отто (1995). «Биохимия клеточной стенки архей и ее филогенетические последствия». Журнал биологической физики . 20 (1–4): 165–169. дои : 10.1007/BF00700433 . S2CID 83906865 .
- ^ Кандлер, Отто (1998). «Раннее разнообразие жизни и происхождение трех областей: предложение» . У Юргена Вигеля; Майкл В.В. Адамс (ред.). Термофилы: ключи к молекулярной эволюции и происхождению жизни? . Лондон: Тейлор и Фрэнсис Лтд., стр. 19–31. ISBN 978-0-203-48420-3 . Архивировано из оригинала 25 февраля 2023 года . Проверено 21 июня 2023 г.
- ^ Вехтерсхойзер, Гюнтер (2003). «От предклеток к эукариям – история двух липидов» . Молекулярная микробиология . 47 (1): 13–22. дои : 10.1046/j.1365-2958.2003.03267.x . ПМИД 12492850 . S2CID 37944519 .
- ^ Вехтерсхойзер, Гюнтер (октябрь 2006 г.). «От вулканического происхождения хемоавтотрофной жизни до бактерий, архей и эукариев» . Философские труды Лондонского королевского общества. Серия Б, Биологические науки . 361 (1474): 1787–1808. дои : 10.1098/rstb.2006.1904 . ПМЦ 1664677 . ПМИД 17008219 .
- ^ Вёзе, Карл (июнь 1998 г.). «Всемирный предок» . ПНАС . 95 (12): 6854–6859. Бибкод : 1998PNAS...95.6854W . дои : 10.1073/pnas.95.12.6854 . ПМК 22660 . ПМИД 9618502 .
- ^ Перейти обратно: а б Эгель, Ричард (март 2012 г.). «Первичный эукариогенез: об общей природе доклеточных состояний, предков современной жизни» . Жизнь . 2 (1): 170–212. Бибкод : 2012Life....2..170E . дои : 10.3390/life2010170 . ПМК 4187143 . ПМИД 25382122 .
- ^ Глансдорф, Николас; Сюй, Ин; Лабедан, Бернар (9 июля 2008 г.). «Последний универсальный общий предок: появление, конституция и генетическое наследие неуловимого предшественника» . Биология Директ . 3 (1): 29. дои : 10.1186/1745-6150-3-29 . ПМЦ 2478661 . ПМИД 18613974 . S2CID 18250196 .
- ^ Сталь, М.; Пенни, Д. (май 2010 г.). «Происхождение жизни: общее происхождение подвергнуто испытанию» . Природа . 465 (7295): 168–169. Бибкод : 2010Natur.465..168S . дои : 10.1038/465168a . ПМИД 20463725 . S2CID 205055573 .
- ^ Теобальд, Д.Л. (май 2010 г.). «Формальная проверка теории универсального общего происхождения». Природа . 465 (7295): 219–222. Бибкод : 2010Natur.465..219T . дои : 10.1038/nature09014 . ПМИД 20463738 . S2CID 4422345 .
- ^ Просдочими, Франциско; Хосе, Марко В.; де Фариас, Савио Торрес (2019). «Первый универсальный общий предок (FUCA) как самый ранний предок линии LUCA (последнего UCA)» . В Понтаротти, Пьер (ред.). Эволюция, происхождение жизни, концепции и методы . Чам: Спрингер. стр. 43–54. дои : 10.1007/978-3-030-30363-1_3 . ISBN 978-3-030-30363-1 . S2CID 199534387 . Проверено 2 ноября 2023 г.
- ^ Просдочими, Франциско; Хосе, Марко В.; де Фариас, Савио Торрес (2019), «Первый универсальный общий предок (FUCA) как самый ранний предок линии LUCA (последнего UCA)», в Понтаротти, Пьер (редактор), « Эволюция, происхождение жизни, концепции и методы» , Чам: Springer, стр. 43–54, номер документа : 10.1007/978-3-030-30363-1_3 , ISBN. 978-3-030-30363-1 , S2CID 199534387 , заархивировано из оригинала 23 февраля 2024 г. , получено 2 ноября 2023 г.
- ^ Перейти обратно: а б Крупович, М.; Доля, В.В.; Кунин, Евгений В. (2020). «LUCA и его комплекс виром» (PDF) . Обзоры природы Микробиология . 18 (11): 661–670. дои : 10.1038/s41579-020-0408-x . PMID 32665595 . S2CID 220516514 . Архивировано (PDF) из оригинала 21 октября 2022 года . Проверено 15 августа 2021 г.
- ^ Фортерре, Патрик; Прангишвили, Давид (2009). «Происхождение вирусов». Исследования в области микробиологии . 160 (7): 466–472. дои : 10.1016/j.resmic.2009.07.008 . ПМИД 19647075 . S2CID 2767388 .
- ^ Дуржиньска, Юлия; Годзицка-Юзефиак, Анна (16 октября 2015 г.). «Вирусы и клетки переплетались с самого начала эволюции» . Вирусологический журнал . 12 (1): 169. дои : 10.1186/s12985-015-0400-7 . ПМК 4609113 . ПМИД 26475454 .
- ^ Насир, Аршан; Ким, Кён Мо; Каэтано-Аноллес, Густаво (1 сентября 2012 г.). «Вирусная эволюция» . Мобильные генетические элементы . 2 (5): 247–252. дои : 10.4161/mge.22797 . ISSN 2159-2543 . ПМЦ 3575434 . ПМИД 23550145 .
Дальнейшее чтение
[ редактировать ]- Лейн, Ник (2016) [2015]. Жизненно важный вопрос . Лондон: Профильные книги . ISBN 978-1781250372 .