Целлюляризация
В эволюционной биологии термин «клеточность» ( клеточность ) использовался в теориях, объясняющих эволюцию клеток , например, в доклеточной теории . [1] [2] [3] дело об эволюции первых клеток на этой планете и в синцитиальной теории [4] пытаясь объяснить происхождение Metazoa от одноклеточных организмов.
Процессы клеточного развития в многоядерных клетках ( синцитии , множественные синцитии) животных и растений также называют целлюляризацией, часто называемой синцитиальной целлюляризацией .
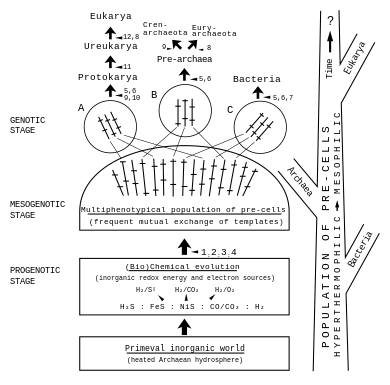
Ключ:
1 Восстановительное образование органических соединений из CO или CO2 с помощью координационной химии Me-серы.
2 использование различных источников окислительно-восстановительной энергии и образование примитивных ферментов и шаблонов.
3 элемента аппарата транскрипции и трансляции и свободные ассоциации
4 образование предклеток
5 стабилизированных кольцевых или линейных геномов
6 цитоплазматических мембран
7 жестких муреиновых клеточных стенок
8 различных немуреиновых жестких клеточных стенок
9 гликопротеиновая клеточная оболочка или гликокаликс
10 цитоскелет
11 сложных хромосом и ядерная мембрана
12 клеточных органелл посредством эндосимбиоза».
Доклеточная теория
[ редактировать ]Согласно Отто Кандлера доклеточной теории , [1] [2] [3] ранняя эволюция жизни и первичный метаболизм (см. Гипотезу мира железа и серы - первый сценарий метаболизма, согласно Вехтерсхойзеру). [5] [6] ) привели к ранней диверсификации жизни посредством эволюции мультифенотипической популяции предклеток , [1] [2] [3] из которых произошли три группы-основатели A, B, C, а затем из них клетки-предшественники (здесь называемые протоклетками) трех областей жизни. [7] возникло последовательно. [1] [2] [3] [8]
В этом сценарии три области жизни произошли не от наследственной почти полной «первой клетки» и не от клеточного организма, который часто называют последним универсальным общим предком ( LUCA) . [9] [10] [11] ), а из популяции развивающихся предклеток. Кандлер ввел термин «клеточность» для своей концепции последовательной эволюции клеток посредством процесса эволюционных улучшений. [1] [2] [3]
Его концепция может объяснить квазислучайное распределение эволюционно важных особенностей между тремя доменами и в то же время существование наиболее основных биохимических особенностей (генетический код, набор белковых аминокислот и т. д.) во всех трех доменах (единство жизни), а также о тесном родстве архей и эукариев. Предклеточную теорию Кандлера поддерживает Вехтерсхойзер. [8]
По мнению Кандлера, защита хрупких первичных форм жизни от окружающей среды посредством изобретения оболочек (т.е. мембран, стенок) была существенным улучшением. Например, появление жестких клеточных стенок благодаря изобретению и разработке пептидогликана. [12] у бактерий (домен Бактерии ), возможно, было предпосылкой их успешного выживания, радиации и колонизации практически всех местообитаний геосферы и гидросферы. [3]
Предполагается совместная эволюция биосферы и геосферы: «Эволюционирующая жизнь может проникнуть в более разнообразные среды обитания, даже в микроаэробные среды обитания на мелководье, освещенные поверхностные воды. Непрерывные изменения физической среды на стареющей и остывающей Земле привели к дальнейшей диверсификации местообитаний и способствовали оппортунистическому распространению примитивной жизни на многочисленные фенотипы на основе каждой из различных хемолитоавтотрофий. Одновременно с накоплением органического вещества, происходящего от хемолитоавтотрофной жизни, могла развиться также условно-патогенная и облигатная гетеротрофная жизнь». [1] : 155 ф.
Детали предложения Кандлера о раннем разнообразии жизни представлены на схеме, где цифры указывают на эволюционные улучшения. [3]
Синцитиальная теория или теория реснитчато-акоэль
[ редактировать ]Эта теория также известна как теория клеточности . Это теория, объясняющая происхождение Metazoa . Идею предложил Хаджи (1953). [4] и Хэнсон (1977). [13]
Эта теория целлюляризации (синцитиальная) утверждает, что многоклеточные животные произошли от одноклеточных инфузорий с множеством ядер, прошедших клеточную обработку. Во-первых, у инфузории появился вентральный рот для питания, и все ядра переместились на одну сторону клетки. Во-вторых, эпителий был создан мембранами, образующими барьеры между ядрами. Таким способом из одной многоядерной клетки ( синцития ) был создан многоклеточный организм. [14]
Пример и критика
[ редактировать ]Плоские черви турбеллярии
[ редактировать ]Согласно синцитиальной теории, предок инфузорий путем нескольких процессов клеточной формации эволюционировал в известных в настоящее время турбеллярийных плоских червей , которые, следовательно, являются наиболее примитивными многоклеточными животными. Теория клеточности основана на большом сходстве инфузорий и плоских червей. И инфузории, и плоские черви имеют реснички , двусторонне симметричны и синцитиальны . Поэтому теория предполагает, что двусторонняя симметрия более примитивна, чем радиальная симметрия . Однако современные биологические данные показывают, что наиболее примитивные формы многоклеточных животных обладают радиальной симметрией, и, таким образом, радиально-симметричные животные, такие как книдарии, не могут происходить от двусторонних плоских червей. [15]
Делая вывод о том, что первыми многоклеточными животными были плоские черви, можно также предположить, что более простые организмы, такие как губки , гребневики и книдарии, произошли от более сложных животных. [16] Однако большинство современных молекулярных исследований показали, что губки — самые примитивные многоклеточные животные. [17] [18]
Зародышевые листки образуются одновременно
[ редактировать ]Синцитиальная теория отвергает теорию зародышевых листков . В ходе развития турбеллярии ( Acoela ) образуются три участка без образования зародышевых листков. Из этого был сделан вывод, что зародышевые листки формируются одновременно в процессе целлюляризации. Это противоречит теории зародышевого слоя, согласно которой эктодерма , энтодерма и мезодерма (у более сложных животных) формируют эмбрион. [19]
Макро- и микроядро инфузорий.
[ редактировать ]Существует множество доказательств того, что инфузории являются предками многоклеточных животных. Инфузории имеют два типа ядер: микронуклеус , который используется в качестве зародышевого ядра, и макронуклеус , который регулирует вегетативный рост. [20] Такое деление ядер является уникальной особенностью инфузорий и не встречается ни у каких других представителей животного мира. [21] Поэтому маловероятно, что инфузории действительно являются предками многоклеточных животных. Это подтверждается молекулярно- филогенетическим исследованием. Инфузории никогда не встречались рядом с животными ни в одной молекулярной филогении . [22]
Жгутиковые сперматозоиды
[ редактировать ]Более того, синцитиальная теория не может объяснить жгутиковые сперматозоиды многоклеточных животных. Поскольку предок ресничек не имеет жгутиков и маловероятно, что жгутики возникли как признак de novo у многоклеточных животных, синцитиальная теория делает практически невозможным объяснение происхождения жгутиковых сперматозоидов. [19]
Из-за отсутствия молекулярных и морфологических доказательств этой теории альтернативная колониальная теория Геккеля в настоящее время получает широкое признание.
Дополнительные теории см. в основной статье « Многоклеточные организмы» .
Целлюляризация в синцитии (целлюляризация синцития)
[ редактировать ]Развитие клеток синцития (многоядерных клеток) называется целлюляризацией синцития. Синцитии довольно часто встречаются у животных и растений. Целлюляризация синцития происходит, например, при эмбриональном развитии животных и при развитии эндосперма растений. Вот два примера:
дрозофилы melanogaster Развитие
[ редактировать ]В эмбриональном развитии Drosophila melanogaster происходят первые 13 ядерных делений, образующих синцитиальную бластодерму, состоящую примерно из 6000 ядер. На более поздней стадии гаструляции между ядрами образуются мембраны и клеточная обработка завершается. [23]
Клеточность синцития у растений
[ редактировать ]Термин «синцитиальная клеточность» используется, например, для обозначения процесса развития клеток в эндосперме Poaceae , например , ячменя ( Hordeum vulgare ), [24] рис ( Oryza sativa ). [25]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Перейти обратно: а б с д и ж Кандлер, Отто (1994). «Раннее разнообразие жизни». В Стефане Бенгтсоне (ред.). Ранняя жизнь на Земле. Нобелевский симпозиум 84 . Нью-Йорк: Columbia UP, стр. 152–160.
- ^ Перейти обратно: а б с д и Кандлер, Отто (1995). «Биохимия клеточной стенки архей и ее филогенетические последствия». Журнал биологической физики . 20 (1–4): 165–169. дои : 10.1007/BF00700433 . S2CID 83906865 .
- ^ Перейти обратно: а б с д и ж г час Кандлер, Отто (1998). «Раннее разнообразие жизни и происхождение трех областей: предложение» . У Юргена Вигеля; Майкл В.В. Адамс (ред.). Термофилы: ключи к молекулярной эволюции и происхождению жизни? . Лондон: Тейлор и Фрэнсис Лтд., стр. 19–31. ISBN 978-0-203-48420-3 .
- ^ Перейти обратно: а б Хаджи, Дж. (1 декабря 1953 г.). «Попытка реконструировать систему классификации животных» . Систематическая биология . 2 (4): 145–154. дои : 10.2307/sysbio/2.4.145 . ISSN 1063-5157 .
- ^ Вехтерсхойзер, Гюнтер (декабрь 1988 г.). «До ферментов и шаблонов: теория поверхностного метаболизма» (PDF) . Обзоры микробиологии и молекулярной биологии . 52 (4): 452–484. дои : 10.1128/мр.52.4.452-484.1988 . ПМЦ 373159 . ПМИД 3070320 .
- ^ Вехтерсхойзер, Гюнтер (1998). «Доводы в пользу гипертермофильного, хемолитоавтотрофного происхождения жизни в железо-серном мире» . У Юргена Вигеля; Майкл В.В. Адамс (ред.). Термофилы: ключи к молекулярной эволюции и происхождению жизни? . Лондон: Тейлор и Фрэнсис Лтд., стр. 19–31. ISBN 978-0-203-48420-3 .
- ^ Вёзе, Карл Р .; Кандлер, Отто ; Уилис, ML (июнь 1990 г.). «На пути к естественной системе организмов: предложение по доменам архей, бактерий и эукариев» . Труды Национальной академии наук Соединенных Штатов Америки . 87 (12): 4576–4579. Бибкод : 1990PNAS...87.4576W . дои : 10.1073/pnas.87.12.4576 . ПМК 54159 . ПМИД 2112744 .
- ^ Перейти обратно: а б Вехтерсхойзер, Гюнтер (2003). «От предклеток к эукариям – история двух липидов» . Молекулярная микробиология . 47 (1): 13–22. дои : 10.1046/j.1365-2958.2003.03267.x . ПМИД 12492850 . S2CID 37944519 .
- ^ Гарольд, Франклин М. (2014). В поисках клеточной истории: эволюция строительных блоков жизни . Чикаго, Лондон: Издательство Чикагского университета. ISBN 978-0-226-17428-0 .
- ^ Мэдиган, Майкл Т.; Мартинко, Джон М.; Бендер, Келли С.; Бакли, Дэниел Х.; Шталь, Дэвид А. (2015). Брок Биология микроорганизмов (14-е изд.). Бостон: Pearson Education Limited. стр. 29, 374, 381. ISBN. 978-1-292-01831-7 .
- ^ Мэдиган, Майкл Т.; Айер, Дженнифер; Бакли, Дэниел Х.; Сэттли, Мэтью; Шталь, Дэвид А. (2022). Брок Биология микроорганизмов (16-е изд.). Харлоу: Образование Пирсона. стр. 431, 435. ISBN. 978-1-292-40479-0 .
- ^ Шлейфер, Карл-Хайнц; Кандлер, Отто (1972). «Типы пептидогликана клеточных стенок бактерий и их таксономическое значение» . Бактериологические обзоры . 36 (4): 407–775. дои : 10.1128/бр.36.4.407-477.1972 . ПМК 408328 . ПМИД 4568761 .
- ^ Хэнсон, Эрл Д. (1977). Происхождение и ранняя эволюция животных (1-е изд.). Миддлтаун, Коннектикут: Издательство Уэслианского университета. ISBN 0819550086 . ОСЛК 2597099 .
- ^ Клаутау, М.; Руссо, CAM (2016), «Многоклеточные животные, происхождение», Энциклопедия эволюционной биологии , Elsevier, стр. 1–6, doi : 10.1016/b978-0-12-800049-6.00270-5 , ISBN 9780128004265
- ^ Пилато, Джованни (2007). Происхождение и филогения многоклеточных животных и теория эндодермы как вторичного слоя . Фоксвелл и Дэвис. ISBN 978-1905868063 . OCLC 488084010 .
- ^ Ваггонер, Бен (25 апреля 2001 г.), «Эукариоты и мультиклетки: происхождение», eLS , John Wiley & Sons, Ltd, doi : 10.1038/npg.els.0001640 , ISBN 0470016175
- ^ Шютце, Иоахим; Краско, Анатолий; Кастодио, Марсио Рейс; Ефремова, Софла М.; Мюллер, Изабель М.; Мюллер, Вернер Э.Г. (7 января 1999 г.). «Эволюционные взаимоотношения Metazoa внутри эукариот на основе молекулярных данных Porifera» . Труды Лондонского королевского общества. Серия Б: Биологические науки . 266 (1414): 63–73. дои : 10.1098/rspb.1999.0605 . ISSN 0962-8452 . ПМК 1689648 . ПМИД 10081159 .
- ^ Мануэль, Микаэль; Верхайде, Герт; Моргенштерн, Буркхард; Эрпенбек, Дирк; Шрайбер, Фабиан; Джексон, Дэниел Дж.; Лейс, Салли; Гюядер, Эрве Ле; Винкер, Патрик (28 апреля 2009 г.). «Филогеномика возрождает традиционные взгляды на глубокие взаимоотношения животных» . Современная биология . 19 (8): 706–712. Бибкод : 2009CBio...19..706P . дои : 10.1016/j.cub.2009.02.052 . ISSN 0960-9822 . ПМИД 19345102 .
- ^ Перейти обратно: а б Р.Л.Котпал, профессор (2012). Современный учебник зоологии: Беспозвоночные . Публикации Растоги. ISBN 9788171339037 .
- ^ Прескотт, DM (июнь 1994 г.). «ДНК мерцательных простейших» . Микробиологические обзоры . 58 (2): 233–267. дои : 10.1128/MMBR.58.2.233-267.1994 . ISSN 0146-0749 . ПМК 372963 . ПМИД 8078435 .
- ^ Липскомб, Диана (март 1991 г.). «Протоктисты под рукой. Справочник протоктиста Л. Маргулиса Дж. О. Корлисса М. Мелконяна, ди-джея Чепмена» . Бионаука . 41 (3): 169–170. дои : 10.2307/1311459 . ISSN 0006-3568 . JSTOR 1311459 .
- ^ Шлегель, Мартин (сентябрь 1994 г.). «Молекулярная филогения эукариот». Тенденции в экологии и эволюции . 9 (9): 330–335. дои : 10.1016/0169-5347(94)90153-8 . ISSN 0169-5347 . ПМИД 21236876 .
- ^ Кампос-Ортега, Хосе А.; Хартенштейн, Волкер (11 ноября 2013 г.). Эмбриональное развитие Drosophila melanogaster . Springer Science & Business Media. ISBN 9783662224892 .
- ^ Журнальная статья «Динамика количества транскриптов во время целлюляризации развивающегося эндосперма ячменя» Рунсюань Чжан, Мэтью Р. Такер, Рэйчел А. Бертон, Нил Дж. Ширли, Алан Литтл, Дженни Моррис, Линда Милн, Келли Хьюстон, Пит Э. Хедли, Робби Во, Джеффри Б. Финчер (2016). «Динамика количества транскриптов во время целлюляризации развивающегося эндосперма ячменя» . Физиология растений . 170 (3): 1549–1565. дои : 10.1104/стр.15.01690 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Масанори Мизутани, Такума Наганума, Кен-ичи Цуцуми, Ясуси Сайто (2010). «Синцитий-специфическая экспрессия ингибитора Orysa; KRP3 CDK: значение его участия в контроле клеточного цикла в синцитиальном эндосперме риса (Oryza sativa L.)» . Журнал экспериментальной ботаники . 61 (3): 791–798. дои : 10.1093/jxb/erp343 . ПМЦ 2814109 . ПМИД 19933315 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка )