Микроядро
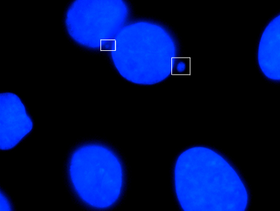
Микроядро ядро — небольшое , которое образуется всякий раз, когда хромосома или фрагмент хромосомы не включается в одно из дочерних ядер во время деления клетки . Обычно это признак генотоксических явлений и хромосомной нестабильности . Микроядра обычно встречаются в раковых клетках и могут указывать на события геномного повреждения, которые могут увеличить риск развития или дегенеративных заболеваний. [1]
Микроядра образуются во время анафазы из отстающих ацентрических хромосом или фрагментов хроматид, вызванных неправильно репарированными или нерепарированными разрывами ДНК или нерасхождением хромосом. Эта неправильная сегрегация хромосом может быть результатом гипометилирования повторяющихся последовательностей, присутствующих в прицентромерной ДНК, нарушений в белках кинетохор или их сборки, дисфункции веретенообразного аппарата или дефектных генов контрольной точки анафазы. [2] Микроядра могут способствовать нестабильности генома , способствуя катастрофическому мутационному событию, называемому хромотрипсис . [3] Многие микроядерные анализы были разработаны для проверки наличия этих структур и определения их частоты в клетках, подвергшихся воздействию определенных химических веществ или стрессовых условий.
Термин «микроядро» может также относиться к меньшему ядру у инфузорных простейших , таких как Paramecium . В митозе оно делится путем деления , а при конъюгации пара микроядер гамет подвергается взаимному слиянию с образованием ядра зиготы , дающего начало макронуклеусам и микроядрам особей следующего цикла деления. [4]
Открытие
[ редактировать ]Микроядра в новообразованных эритроцитах у людей известны как тельца Хауэлла-Джолли, поскольку эти структуры были впервые идентифицированы и описаны в эритроцитах гематологами Уильямом Хауэллом и Джастином Джолли. Позже было обнаружено, что эти структуры связаны с дефицитом витаминов, таких как фолиевая кислота и B12. Взаимосвязь между образованием микроядер и воздействием факторов окружающей среды была впервые обнаружена на клетках кончика корня, подвергшихся ионизирующему излучению. Индукция микроядер химическим веществом была впервые зарегистрирована в асцитных опухолевых клетках Эрлиха, обработанных колхицином. [2]
Формирование
[ редактировать ]Микроядра в основном возникают из ацентрических фрагментов хромосом или отстающих целых хромосом, которые не включаются в дочерние ядра, образующиеся в результате митоза, поскольку им не удается правильно прикрепиться к веретену во время сегрегации хромосом в анафазе. Эти полные хромосомы или фрагменты хроматид в конечном итоге покрываются ядерной мембраной и структурно похожи на обычные ядра, хотя и меньше по размеру. Это маленькое ядро называется микроядром. Образование микроядер можно наблюдать только в клетках, подвергающихся ядерному делению, и его можно четко увидеть с помощью цитохалазина B для блокирования цитокинеза с образованием двуядерных клеток . [2]
Фрагменты ацентрических хромосом могут возникать различными путями. Один из способов заключается в том, что нарушение восстановления двухцепочечных разрывов ДНК может привести к симметричным или асимметричным обменам хроматид и хромосом, а также фрагментов хроматид и хромосом. Если повреждение ДНК превышает восстановительную способность клетки, невосстановленные двухцепочечные разрывы ДНК также могут привести к образованию ацентрических фрагментов хромосомы. Другой способ возникновения эксцентрических фрагментов хромосом заключается в том, что дефекты в генах, связанных с гомологичной рекомбинационной репарацией (например, ATM, BRCA1, BRCA2 и RAD51), приводят к дисфункциональному безошибочному пути гомологичной рекомбинационной репарации ДНК и заставляют клетку прибегнуть к ошибке. -склонный к немумологичному соединению концов путь репарации (NHEJ), увеличивающий вероятность неправильного восстановления разрывов ДНК, образования дицентрических хромосом и фрагментов ацентрических хромосом. Если ферменты пути репарации NHEJ также повреждены, разрывы ДНК могут вообще не восстановиться. Кроме того, одновременная эксцизионная репарация поврежденных или неподходящих оснований, включенных в ДНК, которые находятся рядом и на противоположных комплементарных цепях ДНК, может привести к двухцепочечным разрывам ДНК и образованию микроядер, особенно если этап заполнения пробелов пути репарации не завершен. [2]
Микроядра также могут образовываться из фрагментированных хромосом, когда нуклеоплазматические мостики (NPB) образуются, растягиваются и разрываются во время телофазы. [2]
Образование микроядер также может быть результатом неправильной сегрегации хромосом во время анафазы. Гипометилирование цитозина в центромерных и прицентромерных областях и повторы сателлитной ДНК более высокого порядка в центромерной ДНК могут приводить к таким событиям хромосомной потери. Классическая сателлитная ДНК обычно сильно метилирована по остаткам цитозина, но может стать почти полностью неметилированной из-за синдрома ИКФ (иммунодефицит, нестабильность центромеры и синдром аномалий лица) или после лечения ингибиторами ДНК-метилтрансферазы. Поскольку на сборку белков кинетохор на центромерах влияет метилирование белков цитозина и гистонов, снижение целостности гетерохроматина в результате гипометилирования может мешать прикреплению микротрубочек к хромосомам и распознаванию напряжения от правильных соединений микротрубочек с кинетохорами. Другими возможными причинами потери хромосом, которые могут привести к образованию микроядер, являются дефекты взаимодействия кинетохор и микротрубочек, дефекты сборки митотического веретена, дефекты контрольных точек митоза, аномальная амплификация центросом и слияние концов теломер, которые приводят к образованию дицентрических хромосом, которые отсоединяются от веретена во время митоза. анафаза. Микроядра, возникшие в результате потери хромосом, и фрагменты ацентрических хромосом можно различить с помощью зондов панцентромерной ДНК. [2]
Идентификация
[ редактировать ]Количество микроядер в клетке можно предсказать по следующей формуле:
AF — число ацентрических фрагментов и F = 0,5 — 0,5P, где P — вероятность того, что фрагменты войдут в традиционное ядро и не образуют микроядро. [5]

В одном исследовании, в котором для окрашивания ядерного материала использовалось окрашивание по Гимзе, были установлены следующие критерии идентификации микроядер:
1) диаметр менее 1/3 первичного ядра,
2) невтягиваемость (исключает мелкие частицы пятен),
3) цвет такой же, как у основного ядра, или светлее его (исключая крупные частицы пятен),
4) расположение в пределах 3–4 ядерных диаметров главного ядра, не касаясь его, и
5) не более двух, связанных с одним первичным ядром (3 и более микроядер, вероятно, являются полиморфами или прорубицитами с ядерными фрагментами). [6]
Анализы
[ редактировать ]Микроядерные тесты предоставляют важную информацию о способности химического вещества влиять на структуру и функцию хромосом. Например, многие известные канцерогены человека дают положительный результат при микроядерных тестах на млекопитающих. В ходе этих тестов организмы обрабатывают химическим веществом и измеряют полученную частоту микроядерных клеток. Если наблюдается заметное увеличение количества клеток с микроядрами, можно сделать вывод, что химическое вещество вызывает структурные и/или численные хромосомные повреждения. Поскольку микроядерные тесты необходимо проводить на активно делящихся клетках, идеальными кандидатами являются стволовые клетки костного мозга и эритроциты, которые они производят в результате деления клеток. Эти клетки постоянно и быстро обновляются, а отсутствие настоящего ядра в эритроцитах делает микроядра легко видимыми под микроскопом. [1]
Системы микроядерного анализа очень экономичны, требуют гораздо меньше навыков для оценки, чем обычные метафазные тесты, и работают намного быстрее, чем эти традиционные тесты. Поскольку микроядерные анализы надежно и быстро отражают хромосомные аберрации, они чрезвычайно полезны для быстрой оценки хромосомных повреждений. В частности, анализ CBMNcyt (микроядерный цитом с блокировкой цитокинеза) чрезвычайно универсален и является одним из предпочтительных методов измерения уровня хромосомного повреждения и хромосомной нестабильности в клетках. Анализ микроядер с блокировкой цитокинеза (CBMN) был впервые разработан для оценки микроядер в клетках, завершивших деление ядра, путем блокирования их на двуядерной стадии перед цитокинезом. Позже он превратился в «цитомный» анализ CBMN для дальнейшего изучения гибели клеток, цитостаза и биомаркеров повреждения ДНК. Основным недостатком использования микроядерных тестов является то, что они не могут определить различные типы хромосомных аберраций и на них могут влиять скорость митоза и доля гибели клеток, что искажает результаты. [2]
Паттерны в формировании
[ редактировать ]Многочисленные исследования показали, что частота микроядер у женщин выше, чем у мужчин, и что количество микроядер увеличивается примерно до 70 лет. Уровень микроядер колебался от 0,5–1,4% у мужчин до 0,9–1,8% у женщин. Гендерные различия в основном наблюдались в более молодых возрастных группах (<= 50 лет) с почти двукратной разницей между мужчинами и женщинами. Закономерность количества микроядер после 70 лет противоречива. Некоторые исследования показали, что у лиц старше 70 лет частота микроядер увеличивается у представителей обоих полов. С другой стороны, другие исследования показали, что в самых старших возрастных группах частота микроядер выравнивается. Дефицит микроядер в некоторых старших возрастных группах можно объяснить тем, что микроядерные клетки преимущественно элиминируются путем апоптоза. Однако более высокая частота микроядер соответствует снижению эффективности репарации ДНК и повышению нестабильности генома, что характерно для пожилых людей. Возрастное увеличение частоты микроядер также хорошо коррелирует с возрастным увеличением гипоплоидии и возрастным увеличением потери половых хромосом. С другой стороны, выравнивание частоты микроядер у пожилых людей предполагает порог геномной нестабильности, который невозможно преодолеть, если человек хочет выжить. Если бы это было так, женщины, похоже, достигают этого порога быстрее, чем мужчины. [7]
Половые хромосомы способствуют большинству событий потери хромосом с возрастом. У женщин Х-хромосома может составлять до 72% наблюдаемых микроядер, из которых у 37%, по-видимому, отсутствует функциональная сборка кинетохор, возможно, из-за инактивации Х-хромосомы. Многочисленные исследования показали, что частоты аутосомно-позитивных микроядер у обоих полов и половых хромосом-позитивных МН у мужчин были сходны и оставались неизменными в старших группах, в то время как частота Х-позитивных МН у женщин превышала среднюю частоту аутосомных МН. -положительный МН и продолжал увеличиваться до самого старшего возраста. [2]
Частота хромосомных аберраций, повреждений клеток и микроядер у курильщиков значительно выше, чем у некурящих. [8]
У нормальных людей и многих других млекопитающих, у которых нет ядер в эритроцитах, микроядра быстро удаляются селезенкой . Следовательно, высокая частота микроядер в периферической крови человека указывает на разрыв или отсутствие селезенки. У мышей они не удаляются, что является основой микроядерного теста in vivo .
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Перейти обратно: а б «Микронуклеус» . ntp.niehs.nih.gov . Архивировано из оригинала 18 октября 2016 г. Проверено 14 октября 2016 г.
- ^ Перейти обратно: а б с д и ж г час Фенек, М.; Кирш-Волдерс, М.; Натараджан, АТ; Сюрральес, Дж.; Кротт, Дж.В.; Парри, Дж.; Норппа, Х.; Истмонд, Д.А.; Такер, доктор медицинских наук (1 января 2011 г.). «Молекулярные механизмы формирования микроядер, нуклеоплазматического мостика и ядерных зачатков в клетках млекопитающих и человека» . Мутагенез . 26 (1): 125–132. дои : 10.1093/mutage/geq052 . ISSN 0267-8357 . ПМИД 21164193 .
- ^ Умбрейт, Нил Т.; Чжан, Ченг-Чжун; Линч, Люк Д.; Блейн, Логан Дж.; Ченг, Анна М.; Турдо, Ришар; Солнце, Лили; Альмубарак, Ханна Ф.; Судья Ким; Митчелл, Томас Дж.; Спектор, Александр (17 апреля 2020 г.). «Механизмы, генерирующие сложность генома рака из-за ошибки деления одной клетки» . Наука . 368 (6488): eaba0712. дои : 10.1126/science.aba0712 . ISSN 0036-8075 . ПМЦ 7347108 . ПМИД 32299917 .
- ^ Одно или несколько предыдущих предложений включают текст из публикации, которая сейчас находится в свободном доступе : Чисхолм, Хью , изд. (1911). « Микронуклеус ». Британская энциклопедия . Том. 18 (11-е изд.). Издательство Кембриджского университета. п. 391.
- ^ Сэвидж, Джон РК (1 января 1988 г.). «Комментарий к количественной связи между микроядрами и хромосомными аберрациями». Письма об исследованиях мутаций . 207 (1): 33–36. дои : 10.1016/0165-7992(88)90008-5 . ПМИД 3336377 .
- ^ Земляк, Пол И.; Хеддл, Джон А. (1 декабря 1976 г.). «Производство микроядер из хромосомных аберраций в облученных культурах лимфоцитов человека». Мутационные исследования/Фундаментальные и молекулярные механизмы мутагенеза . 41 (2–3): 321–331. дои : 10.1016/0027-5107(76)90105-6 . ПМИД 796719 .
- ^ Войда, Алина; Зенткевич, Ева; Витт, Михал (1 мая 2007 г.). «Влияние возраста и пола на частоту нерасхождения микроядер и хромосом у долгожителей и более молодых людей» . Мутагенез . 22 (3): 195–200. дои : 10.1093/mutage/gem002 . ISSN 0267-8357 . ПМИД 17284771 .
- ^ Бандана Гангули, Бани (1 августа 1993 г.). «Деление клеток, хромосомное повреждение и образование микроядер в периферических лимфоцитах здоровых доноров: в зависимости от возраста донора». Исследования мутаций/старение ДНК . 295 (3): 135–148. дои : 10.1016/0921-8734(93)90015-У . ПМИД 7689700 .