Кокколит

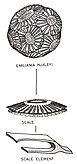
Кокколиты — отдельные пластинки или чешуйки карбоната кальция, образованные кокколитофорами (одноклеточным фитопланктоном типа Emiliania huxleyi ) и покрывающие поверхность клеток, расположенные в виде сферической оболочки, называемой коккосферой .
Обзор
[ редактировать ]Кокколитофоры представляют собой сферические клетки диаметром около 5–100 микрометров, окруженные известковыми пластинками, называемыми кокколитами , диаметром около 2–25 микрометров. [1] Кокколитофоры — важная группа, насчитывающая около 200 видов морского фитопланктона. [2] которые покрывают себя оболочкой из карбоната кальция, называемой «коккосферой». Они важны с экологической и биогеохимической точки зрения, но причина их кальцинирования остается неясной. Одной из ключевых функций может быть то, что коккосфера обеспечивает защиту от хищничества микрозоопланктона , который является одной из основных причин гибели фитопланктона в океане. [3]
![Частичный разрез кокколитофорора со слоем кокколита [4]](//upload.wikimedia.org/wikipedia/commons/thumb/b/b3/Cross_section_of_a_coccolithophore_with_coccolith_layer.png/355px-Cross_section_of_a_coccolithophore_with_coccolith_layer.png) Частичный разрез кокколитофора со слоем кокколита [4]
Частичный разрез кокколитофора со слоем кокколита [4]![Клетка кокколитофора окружена щитом из кокколитов. Клетка, несущая кокколит, называется коккосферой.[5][6]](//upload.wikimedia.org/wikipedia/commons/thumb/2/23/Diagram_of_a_coccolithophore_cell_and_its_shield_of_coccoliths.png/484px-Diagram_of_a_coccolithophore_cell_and_its_shield_of_coccoliths.png) Клетка кокколитофора окружена щитом из кокколитов. Клетка, несущая кокколит, называется коккосферой . [5] [6]
Клетка кокколитофора окружена щитом из кокколитов. Клетка, несущая кокколит, называется коккосферой . [5] [6]
![Частичный разрез кокколитофорора со слоем кокколита [4]](http://upload.wikimedia.org/wikipedia/commons/thumb/b/b3/Cross_section_of_a_coccolithophore_with_coccolith_layer.png/355px-Cross_section_of_a_coccolithophore_with_coccolith_layer.png)
![Клетка кокколитофора окружена щитом из кокколитов. Клетка, несущая кокколит, называется коккосферой.[5][6]](http://upload.wikimedia.org/wikipedia/commons/thumb/2/23/Diagram_of_a_coccolithophore_cell_and_its_shield_of_coccoliths.png/484px-Diagram_of_a_coccolithophore_cell_and_its_shield_of_coccoliths.png)
Кокколитофоры были неотъемлемой частью морских планктонных сообществ начиная с юрского периода . [7] [8] Сегодня кокколитофоры вносят ~ 1–10% в первичную продукцию на поверхности океана. [9] и ~50% – пелагическим осадкам CaCO 3 . [10] Их известковая оболочка увеличивает скорость погружения фотосинтетически связанного CO 2 в глубины океана за счет балластирования органического вещества. [11] [12] В то же время биогенное осаждение карбоната кальция при формировании кокколита снижает общую щелочность морской воды и выделяет CO 2 . [13] [14] Таким образом, кокколитофоры играют важную роль в морском углеродном цикле , влияя на эффективность биологического углеродного насоса и поглощение океаном атмосферного CO 2 . [3]
По состоянию на 2021 год неизвестно, почему кокколитофоры кальцифицируются и как их способность производить кокколиты связана с их экологическим успехом. [15] [16] [17] [18] [19] Наиболее вероятным преимуществом наличия коккосферы является защита от хищников и вирусов. [20] [18] Вирусная инфекция является важной причиной гибели фитопланктона в океанах. [21] недавно было показано, что кальцификация может влиять на взаимодействие между кокколитофором и его вирусом. [22] [23] Основными хищниками морского фитопланктона являются микрозоопланктон, такой как инфузории и динофлагелляты . По оценкам, они потребляют около двух третей первичной продукции океана. [24] а микрозоопланктон может оказывать сильное пастбищное давление на популяции кокколитофоров. [25] Хотя кальцификация не предотвращает хищничество, утверждается, что коккосфера снижает эффективность выпаса, затрудняя хищнику использование органического содержимого кокколитофоров. [26] Гетеротрофные протисты способны выборочно выбирать добычу на основе ее размера или формы, а также с помощью химических сигналов. [27] [28] и, таким образом, может отдавать предпочтение другой добыче, которая доступна и не защищена кокколитами. [3]
Формирование и состав
[ редактировать ]| Часть серии, посвященной |
| Биоминерализация |
|---|
 |
Кокколиты образуются внутри клетки в везикулах, происходящих из тела Гольджи . Когда кокколит сформирован, эти везикулы сливаются с клеточной стенкой, и кокколит экзоцитозируется и включается в коккосферу. Кокколиты либо рассеиваются после гибели и распада коккосферы, либо постоянно сбрасываются некоторыми видами. Они погружаются в толщу воды, образуя важную часть глубоководных отложений (в зависимости от глубины воды). Томас Хаксли был первым человеком, наблюдавшим эти формы в современных морских отложениях, и дал им название «кокколиты» в отчете, опубликованном в 1858 году. [29] [30] Кокколиты состоят из карбоната кальция в виде минерала кальцита и являются основным компонентом меловых отложений, таких как белые скалы Дувра (отложившиеся в меловой период), в которых они были впервые описаны Генри Клифтоном Сорби в 1861 году. [31]
 Разрушенная коккосфера Pleurochrysis carterae
Разрушенная коккосфера Pleurochrysis carterae

Типы
[ редактировать ]Существует два основных типа кокколитов: гетерококколиты и голококколиты . Гетерококколиты состоят из радиального массива кристаллических единиц сложной формы. Голококколиты состоят из мельчайших (~0,1 микрометра) кальцитовых ромбоэдров , расположенных непрерывными рядами. Первоначально считалось, что эти два типа кокколитов производятся разными семействами кокколитофоров. Однако теперь благодаря совокупности наблюдений за полевыми образцами и лабораторными культурами известно, что два типа кокколитов производятся одним и тем же видом, но на разных стадиях жизненного цикла. Гетерококколиты образуются в диплоидной фазе жизненного цикла, а голококколиты - в гаплоидной фазе. Как в полевых пробах, так и в лабораторных культурах существует возможность наблюдения клетки, покрытой комбинацией гетерококколитов и голококколитов. Это указывает на переход от диплоидной к гаплоидной фазе вида. Такое сочетание кокколитов наблюдалось в полевых образцах, многие из которых были доставлены из Средиземноморья. [32] [33]

Форма
[ редактировать ]Кокколиты также классифицируются в зависимости от формы. Общие формы включают в себя: [34] [35]
- Калиптролит - корзинчатая форма с отверстиями у основания.
- Канеолит – диско- или чашеобразный.
- Цератолит – имеет форму подковы или поперечного рычага.
- Крибрилит – дискообразный, с многочисленными перфорациями в центральной части.
- Циртолит - выпуклый диск, может с выступающим центральным отростком.
- Дисколит - эллипсоидной формы с приподнятым венчиком, в некоторых случаях высокий венчик образует вазообразную или чашеобразную структуру.
- Геликолит - плаколит со спиральным краем.
- Лопадолит – корзинчатая или чашеобразная форма с высоким венчиком, открывающаяся дистально.
- Пенталит – пятиугольная форма, состоящая из пяти четырехгранных кристаллов.
- Плаколит - ободок, состоящий из двух пластин, сложенных друг на друга.
- Призматолит - многоугольный, может иметь перфорацию.
- Рабдолит – одиночная пластинка с булавовидным центральным отростком.
- Скафолит – ромбоэдрический, с параллельными линиями в центре.

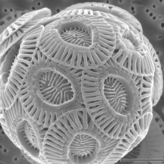
![Структуры кокколита представителей Noelaerhabdaceae.[36] Каждый морфовид связан со СЭМ-изображением на следующей диаграмме.](//upload.wikimedia.org/wikipedia/commons/2/28/Coccolith_structures_of_representative_Noelaerhabdaceae.jpg) Кокколитовые структуры представителей Noelaerhabdaceae . [36]
Кокколитовые структуры представителей Noelaerhabdaceae . [36]
Каждый морфовид связан со СЭМ-изображением на следующей диаграмме.
![Структуры кокколита представителей Noelaerhabdaceae.[36] Каждый морфовид связан со СЭМ-изображением на следующей диаграмме.](http://upload.wikimedia.org/wikipedia/commons/2/28/Coccolith_structures_of_representative_Noelaerhabdaceae.jpg)
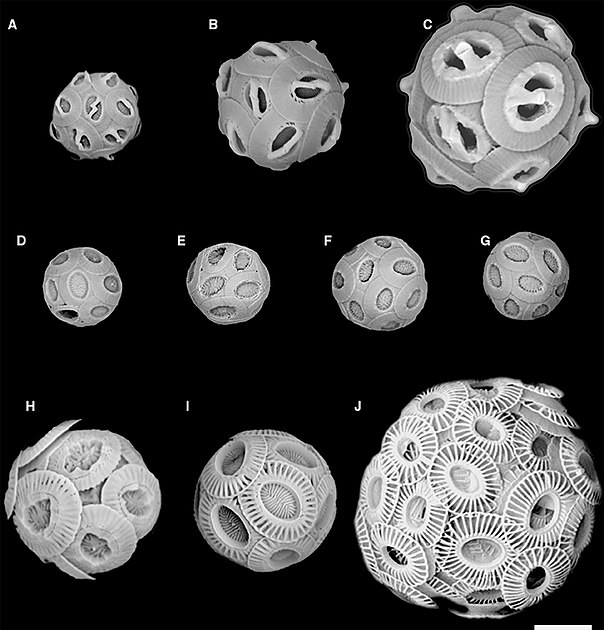 СЭМ-изображения соответствуют рисункам кокколита на предыдущей диаграмме.
СЭМ-изображения соответствуют рисункам кокколита на предыдущей диаграмме.
(A) Gephyrocapsa ericsonii RCC4032 (B) Gephyrocapsa muellerae (C) Gephyrocapsa Oceanica (D) Reticulofenestra parvular RCC4033; (E) ретикулофенестра парвулярная RCC4034; (F) парвулярный ретикулофенестра RCC4035; (G) ретикулофенестра парвулярная RCC4036; (H) Emiliania huxleyi морфотип R ; (I) Emiliania huxleyi морфотип А; (J) Emiliania huxleyi морфотип B.

Функция
[ редактировать ]Хотя кокколиты представляют собой чрезвычайно сложные структуры, формирование которых является сложным продуктом клеточных процессов, их функция неясна. Гипотезы включают защиту от выпаса зоопланктона или заражения бактериями или вирусами ; поддержание плавучести ; выделение углекислого газа для фотосинтеза ; фильтровать вредное ультрафиолетовое излучение; или у глубоко обитающих видов - для концентрации света для фотосинтеза.
Ископаемая запись
[ редактировать ]Поскольку кокколиты состоят из кальцита с низким содержанием магния, наиболее стабильной формы карбоната кальция, они легко окаменевают. Они встречаются в отложениях вместе с аналогичными микрофоссилиями неопределенного родства (нанолитами) от верхнего триаса до современности. Они широко используются в качестве биостратиграфических маркеров и палеоклиматических индикаторов. Кокколиты и родственные им окаменелости называются известковыми нанофоссилиями или известковым наннопланктоном (нанопланктоном) .
Ссылки
[ редактировать ]- ^ Мохеймани, Северная Каролина; Уэбб, JP; Боровицка, Массачусетс (2012), «Биоремедиация и другие потенциальные применения кокколитофоридных водорослей: обзор. Биоремедиация и другие потенциальные применения кокколитофоридных водорослей: обзор», Algal Research , 1 (2): 120–133, doi : 10.1016/ j.algal.2012.06.002
- ^ Янг, младший; Гейзен, М.; Проберт, И. (2005). «Обзор избранных аспектов биологии кокколитофоров, имеющих значение для оценки палеобиоразнообразия» (PDF) . Микропалеонтология . 51 (4): 267–288. дои : 10.2113/gsmicropal.51.4.267 .
- ^ Перейти обратно: а б с Хауност, Матиас; Рибезель, Ульф; Д'Амор, Франческо; Келтинг, Оле; Бах, Леннарт Т. (30 июня 2021 г.). «Влияние карбонатного кальция панциря кокколитофоров на питание и рост динофлагеллятного хищника» . Границы морской науки . 8 . Фронтирс Медиа С.А. дои : 10.3389/fmars.2021.664269 . ISSN 2296-7745 .
 Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 . - ^ Ирие, Такахиро; Бесшо, Кадзухиро; Финдли, Хелен С.; Калози, Пьеро (15 октября 2010 г.). «Увеличение затрат из-за закисления океана приводит к более сильной кальцификации фитопланктона: оптимальная стратегия роста кокколитофоров» . ПЛОС ОДИН . 5 (10). Публичная научная библиотека (PLoS): e13436. дои : 10.1371/journal.pone.0013436 . ISSN 1932-6203 . Измененный материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Алоизи, Г. (6 августа 2015 г.). «Ковариация скорости метаболизма и размера клеток у кокколитофоров» . Биогеонауки . 12 (15). Коперник ГмбХ: 4665–4692. Бибкод : 2015BGeo...12.4665A . дои : 10.5194/bg-12-4665-2015 . ISSN 1726-4189 . S2CID 6227548 .
- ^ Хендерикс, Йоринтье (2008). «Правила размера кокколитофоров — реконструкция геометрии древних клеток и доли клеточного кальцита на основе ископаемых кокколитов». Морская микропалеонтология . 67 (1–2). Эльзевир Б.В.: 143–154. Бибкод : 2008МарМП..67..143H . doi : 10.1016/j.marmicro.2008.01.005 . ISSN 0377-8398 .
- ^ Баун, Пол Р.; Лиз, Джеки А.; Янг, Джереми Р. (2004). «Эволюция и разнообразие известкового наннопланктона с течением времени». Кокколитофоры . стр. 481–508. дои : 10.1007/978-3-662-06278-4_18 . ISBN 978-3-642-06016-8 .
- ^ Хэй, Уильям В. (2004). «Карбонатные потоки и известковый наннопланктон». Кокколитофоры . стр. 509–528. дои : 10.1007/978-3-662-06278-4_19 . ISBN 978-3-642-06016-8 .
- ^ Поултон, Алекс Дж.; Ади, Тим Р.; Балч, Уильям М.; Холлиган, Патрик М. (2007). «Связь скорости кальцификации кокколитофоров с динамикой сообщества фитопланктона: региональные различия и последствия для экспорта углерода». Глубоководные исследования, часть II: Актуальные исследования в океанографии . 54 (5–7): 538–557. Бибкод : 2007DSRII..54..538P . дои : 10.1016/j.dsr2.2006.12.003 .
- ^ Брокер, Уоллес; Кларк, Элизабет (2009). «Соотношение кокколита CaCO3 и фораминифер CaCO3 в глубоководных отложениях позднеголоцена» . Палеоокеанография . 24 (3). Бибкод : 2009PalOc..24.3205B . дои : 10.1029/2009PA001731 .
- ^ Клаас, Кристина; Арчер, Дэвид Э. (2002). «Связь тонущего органического вещества с различными типами минерального балласта в глубоководных водах: влияние на соотношение осадков» . Глобальные биогеохимические циклы . 16 (4): 1116. Бибкод : 2002GBioC..16.1116K . дои : 10.1029/2001GB001765 . S2CID 34159028 .
- ^ Хондзё, Сусуму; Манганини, Стивен Дж.; Кришфилд, Ричард А.; Франсуа, Роджер (2008). «Потоки твердых частиц органического углерода в недра океана и факторы, контролирующие биологический насос: синтез глобальных программ по улавливанию отложений с 1983 года». Прогресс в океанографии . 76 (3): 217–285. Бибкод : 2008Proce..76..217H . дои : 10.1016/j.pocean.2007.11.003 .
- ^ Франкиньюль, Мишель; Кэнон, Кристина; Гаттузо, Жан-Пьер (1994). «Морская кальцификация как источник углекислого газа: положительная обратная связь с увеличением содержания CO2 в атмосфере» . Лимнология и океанография . 39 (2): 458–462. Бибкод : 1994LimOc..39..458F . дои : 10.4319/lo.1994.39.2.0458 . hdl : 2268/246251 .
- ^ Рост, Бьёрн; Рибезель, Ульф (2004). «Кокколитофоры и биологический насос: реакция на изменения окружающей среды». Кокколитофоры . стр. 99–125. дои : 10.1007/978-3-662-06278-4_5 . ISBN 978-3-642-06016-8 .
- ^ Янг, младший (1987). Возможные функциональные интерпретации морфологии кокколита. Нью-Йорк: Springer-Verlag, 305–313.
- ^ Янг, младший (1994). «Функции кокколитов», в книге «Кокколитофоры», под редакцией А. Винтер и У. Г. Сиссера (Кембридж: издательство Кембриджского университета), 63–82.
- ^ Рэйвен, Дж.А.; Кроуферд, К. (2012). «Экологический контроль кальцификации кокколитофоров» . Серия «Прогресс в области морской экологии» . 470 : 137–166. Бибкод : 2012MEPS..470..137R . дои : 10.3354/meps09993 . hdl : 10453/114799 .
- ^ Перейти обратно: а б Монтейро, Фанни М.; Бах, Леннарт Т.; Браунли, Колин; Баун, Пол; Рикаби, Розалинда EM; Поултон, Алекс Дж.; Тиррелл, Тоби; Бофорт, Люк; Дуткевич, Стефани; Гиббс, Саманта; Гутовска, Магдалена А.; Ли, Рене; Рибезель, Ульф; Янг, Джереми; Риджвелл, Энди (2016). «Почему кальцинируется морской фитопланктон» . Достижения науки . 2 (7): e1501822. Бибкод : 2016SciA....2E1822M . дои : 10.1126/sciadv.1501822 . ПМЦ 4956192 . ПМИД 27453937 .
- ^ Мюллер, Мариус Н. (2019). «О происхождении и функции кальцификации кокколитофоров» . Границы морской науки . 6 . дои : 10.3389/fmars.2019.00049 .
- ^ Хамм, Кристиан; Сметачек, Виктор (2007). «Доспехи: почему, когда и как». Эволюция первичных продуцентов в море . стр. 311–332. дои : 10.1016/B978-012370518-1/50015-1 . ISBN 9780123705181 .
- ^ Брюссаард, Корина П.Д. (2004). «Вирусный контроль популяций фитопланктона - обзор1». Журнал эукариотической микробиологии . 51 (2): 125–138. дои : 10.1111/j.1550-7408.2004.tb00537.x . ПМИД 15134247 . S2CID 21017882 .
- ^ Джонс, Кристофер Т.; Грабб, Остин Р.; Нисимов, Юзеф И.; Натале, Фрэнк; Кнапп, Вики; Муи, Элвин; Фредрикс, Хелен Ф.; Ван Мой, Бенджамин А.С.; Бидл, Кей Д. (2019). «Взаимосвязь между кальцификацией и кокколитовирусной инфекцией» . Экологическая микробиология . 21 (6): 1896–1915. дои : 10.1111/1462-2920.14362 . ПМЦ 7379532 . ПМИД 30043404 .
- ^ Хауност, Матиас; Рибезель, Ульф; Бах, Леннарт Т. (2020). «Оболочка карбоната кальция Emiliania huxleyi обеспечивает ограниченную защиту от вирусной инфекции» . Границы морской науки . 7 . дои : 10.3389/fmars.2020.530757 .
- ^ Кальбет, Альберт; Лэндри, Майкл Р. (2004). «Рост фитопланктона, выпас микрозоопланктона и круговорот углерода в морских системах». Лимнология и океанография . 49 (1): 51–57. Бибкод : 2004LimOc..49...51C . дои : 10.4319/lo.2004.49.1.0051 . hdl : 10261/134985 . S2CID 22995996 .
- ^ Майерс, KMJ; Поултон, Эй Джей; Дэниелс, CJ; Уэллс, СР; Вудворд, служба скорой помощи; Тарран, Джорджия; Виддикомб, CE; Мэр, диджей; Аткинсон, А.; Гиринг, SLC (2019). «Рост и смертность кокколитофоров весной в шельфовом море с умеренным климатом (Кельтское море, апрель 2015 г.)» . Прогресс в океанографии . 177 : 101928. Бибкод : 2019Proce.17701928M . дои : 10.1016/j.pocean.2018.02.024 . S2CID 135347218 .
- ^ Янг, младший (1994) «Функции кокколитов». В: Кокколитофоры , редакторы А. Винтер и У. Г. Сиссер (Кембридж: издательство Кембриджского университета), 63–82.
- ^ Тиллманн, Урбан (2004). «Взаимодействие между планктонными микроводорослями и простейшими травоядными1». Журнал эукариотической микробиологии . 51 (2): 156–168. дои : 10.1111/j.1550-7408.2004.tb00540.x . ПМИД 15134250 . S2CID 36526359 .
- ^ Брекельс, Миннесота; Робертс, ЕС; Арчер, SD; Малин, Г.; Стейнке, М. (2011). «Роль растворенных инфохимических веществ в обеспечении взаимодействия хищник-жертва у гетеротрофной динофлагелляты Oxyrris marina» . Журнал исследований планктона . 33 (4): 629–639. дои : 10.1093/plankt/fbq114 .
- ^ Хаксли, Томас Генри (1858). «Приложение А» . Глубоководные зондирования в северной части Атлантического океана между Ирландией и Ньюфаундлендом, произведенные на корабле HMS Cyclops под командованием лейтенанта Джозефа Дэймана в июне и июле 1857 года . Лондон: Британское Адмиралтейство. С. 63–68 [64].
- ^ Хаксли, Томас Генри (1868). «О некоторых организмах, обитающих на большой глубине в Северной Атлантическом океане» . Ежеквартальный журнал микроскопической науки . Новая серия. 8 : 203–212.
- ^ Сорби, Генри Клифтон (1861). «Об органическом происхождении так называемых «кристаллоидов» мела» . Анналы и журнал естественной истории . Сер. 3. 8 (45): 193–200. дои : 10.1080/00222936108697404 .
- ^ Фортуньо, Хосе Мануэль; Крос, Луиса (30 марта 2002 г.). «Атлас кокколитофоров северо-западного Средиземноморья» . Сциентия Марина . 66 (С1): 1–182. дои : 10.3989/scimar.2002.66s11 . hdl : 10261/5441 . ISSN 1886-8134 .
- ^ Малинверно, Э; Димиза, доктор медицины; Триантафиллу, М.В.; Дермицакис, доктор медицинских наук; Корселли, К. (2008). Кокколитофоры Восточного Средиземноморья: Взгляд в морской микромир . Афины: Издательская группа «ИОН». ISBN 978-960-411-660-7 .
- ^ Амос Винтер; Уильям Г. Сиссер (2006). Кокколитофоры . Издательство Кембриджского университета. стр. 54–58. ISBN 978-0-521-03169-1 .
- ^ Кармело Р. Томас (2012). Морской фитопланктон: Путеводитель по голым жгутиконосцам и кокколитофоридам . Академическая пресса. стр. 161–165. ISBN 978-0-323-13827-7 .
- ^ Бендиф, Эль-Махди; Проберт, Ян; Диас-Росас, Франциско; Томас, Даниэла; ван ден Энг, Гер; Янг, Джереми Р.; фон Дассов, Питер (24 мая 2016 г.). «Недавняя сетчатая эволюция экологически доминирующей линии кокколитофоров» . Границы микробиологии . 7 . Фронтирс Медиа С.А. дои : 10.3389/fmicb.2016.00784 . ISSN 1664-302X . Измененный материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
Внешние ссылки
[ редактировать ]- Веб-сайт EHUX — сайт, посвященный Emiliania huxleyi , содержащий очерки о цветении, функции кокколита и т. д.
- Сайт Международной ассоциации наннопланктона - включает иллюстрированное руководство по терминологии кокколитов и несколько галерей изображений.
- Наннотакс - иллюстрированное руководство по систематике кокколитофоров и других нанноокаменелостей.
- Cocco Express - Теги экспрессированных последовательностей кокколитофоридов (EST) и база данных микрочипов
- Возможные функции кокколитов
| Базы данных органов управления : Национальные |
|---|