Раковина протиста

| Часть серии, посвященной |
| Биоминерализация |
|---|
 |
Многие протисты имеют защитные панцири или панцири . [2] обычно изготавливается из кремнезема (стекла) или карбоната кальция (мела). Протисты — это разнообразная группа эукариотных организмов, которые не являются растениями, животными или грибами. Обычно это микроскопические одноклеточные организмы , живущие в воде или во влажной среде.
Раковины протистов часто представляют собой твердые минерализованные формы, которые устойчивы к разложению и могут пережить смерть простейшего в виде микроископаемого . Хотя протисты обычно очень малы, они повсеместно распространены. Их численность такова, что их раковины играют огромную роль в формировании океанских отложений и в глобальном круговороте элементов и питательных веществ.
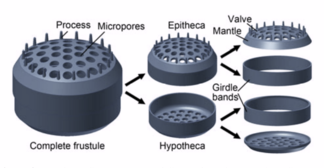
Роль раковин протистов зависит от типа протистов. Простейшие, такие как диатомеи и радиолярии, имеют сложные, похожие на стекло оболочки из кремнезема , которые являются твердыми и защитными и служат барьером, предотвращающим потерю воды. Раковины имеют небольшие поры, которые обеспечивают газообмен и поглощение питательных веществ. Кокколитофоры и фораминиферы также имеют твердые защитные панцири, но раковины состоят из карбоната кальция . Эти раковины повышают плавучесть, позволяя организмам плавать в толще воды и легче передвигаться.
Помимо защиты и поддержки, панцири протистов также служат учёным средством идентификации. Изучая характеристики раковин, можно идентифицировать различные виды протистов и изучить их экологию и эволюцию.
Протисты
[ редактировать ]Клеточная жизнь, вероятно, зародилась как одноклеточные прокариоты (включая современные бактерии и археи), а затем развилась в более сложные эукариоты . Эукариоты включают такие организмы, как растения, животные, грибы и «протисты». Протисты обычно одноклеточные и микроскопические. Они могут быть гетеротрофными , то есть получают питательные вещества, потребляя другие организмы, или автотрофными , то есть они производят собственную пищу посредством фотосинтеза или хемосинтеза , или миксотрофными , то есть они производят свою собственную пищу, используя сочетание этих методов.
Термин «протист» исторически использовался для обозначения группы биологически сходных организмов; однако современные исследования показали, что это парафилетическая группа, которая не содержит всех потомков общего предка. Как таковой он не представляет собой кладу и в настоящее время официально не используется в научных целях. Тем не менее, этот термин продолжает неофициально использоваться для обозначения тех эукариот , которых нельзя классифицировать как растения, грибы или животные.
Большинство протистов слишком малы, чтобы их можно было увидеть невооруженным глазом. Это очень разнообразные организмы, которые в настоящее время разделены на 18 типов, но их нелегко классифицировать. [3] [4] Исследования показали, что большое разнообразие протистов существует в океанах, глубоких морских жерлах и речных отложениях, что позволяет предположить, что большое количество эукариотических микробных сообществ еще не обнаружено. [5] [6] Как эукариоты, протисты обладают внутри своей клетки по крайней мере одним ядром , а также такими органеллами, как митохондрии и тельца Гольджи . Многие протисты бесполы , но могут быстро размножаться посредством митоза или фрагментации ; другие (включая фораминиферы ) могут размножаться половым или бесполым путем. [7]
В отличие от клеток бактерий и архей клетки протистов и других эукариот высокоорганизованы. Растения, животные и грибы обычно многоклеточные и обычно макроскопические . Большинство протистов одноклеточные и микроскопические, но есть исключения: некоторые морские протисты не являются ни одноклеточными, ни микроскопическими, например морские водоросли .
Оболочки на основе кремния
[ редактировать ]
Хотя кремний легко доступен в форме силикатов , очень немногие организмы используют его напрямую. Диатомовые водоросли , радиолярии и кремнистые губки используют биогенный кремнезем в качестве структурного материала для своих скелетов. кремнезема У более развитых растений фитолиты (фитолиты опала) представляют собой твердые микроскопические тельца, встречающиеся в клетке; некоторым растениям, включая рис , для роста необходим кремнезем. [8] [9] [10] Было показано, что кремнезем улучшает прочность стенок растительных клеток и структурную целостность некоторых растений. [11]
Диатомовые водоросли
[ редактировать ]Диатомовые водоросли образуют (спорный) тип, содержащий около 100 000 признанных видов, в основном одноклеточных водорослей. Диатомовые водоросли производят около 20 процентов кислорода, производимого на планете каждый год. [12] поглощают более 6,7 миллиардов тонн кремния из вод, в которых они обитают, ежегодно [13] и составляют почти половину органического материала, обнаруженного в океанах.
Диатомовые водоросли заключены в защитные кремнеземные (стеклянные) оболочки, называемые панцирями . Красиво спроектированная и сложная структура многих из этих панцирей такова, что их часто называют «морскими драгоценностями». [14] Каждая панцирь состоит из двух взаимосвязанных частей, покрытых крошечными отверстиями, через которые диатомовые водоросли обмениваются питательными веществами и отходами. [15] Панцири мертвых диатомей дрейфуют на дно океана, где за миллионы лет они могут накапливаться на глубине до полумили . [16]
Диатомовые водоросли используют кремний в форме биогенного кремнезема (BSiO 2 ), [17] который поглощается белком-транспортером кремния и преимущественно используется при построении этих защитных структур клеточной стенки. [18] Кремний попадает в океан в растворенной форме, например, в виде кремниевой кислоты или силиката . [19] Поскольку диатомеи являются одними из основных потребителей этих форм кремния, они вносят большой вклад в концентрацию кремния во всем океане. Кремний образует профиль питательных веществ в океане из-за продуктивности диатомей на небольших глубинах, что означает меньшую концентрацию кремния в верхних слоях океана и большую концентрацию кремния в глубоком океане. [19]
Продуктивность диатомовых водорослей в верхних слоях океана способствует увеличению количества кремния, экспортируемого в нижние слои океана. [20] Когда клетки диатомей лизируются в верхних слоях океана, их питательные вещества, такие как железо, цинк и кремний, переносятся в нижние слои океана посредством процесса, называемого морским снегом . Морской снег предполагает перенос твердых частиц органического вещества вниз за счет вертикального смешивания растворенного органического вещества . [21] Доступность кремния, по-видимому, имеет решающее значение для продуктивности диатомовых водорослей, и пока диатомовые водоросли могут использовать кремниевую кислоту, диатомовые водоросли обеспечивают концентрацию других важных питательных веществ в глубоком океане. [22]
В прибрежных зонах диатомеи служат основными фитопланктонными организмами и вносят большой вклад в производство биогенного кремнезема. Однако в открытом океане роль диатомовых водорослей в ежегодном мировом производстве кремнезема снижается. Диатомовые водоросли субтропических круговоротов Северной Атлантики и Северной Тихого океана обеспечивают лишь около 6% мирового годового производства морского кремнезема, в то время как Южный океан производит около одной трети глобального морского биогенного кремнезема. [23] Южный океан называют «биогеохимическим водоразделом», поскольку из этого региона выносится лишь незначительное количество кремния. [24]


 Диатомеи – один из наиболее распространенных видов фитопланктона.
Диатомеи – один из наиболее распространенных видов фитопланктона. Их защитные оболочки (фрустлы) изготовлены из кремния.
Их защитные оболочки (фрустлы) изготовлены из кремния.
 Они бывают разных форм и размеров
Они бывают разных форм и размеров






 Кварцифицированная панцирь пеннатной диатомеи с двумя перекрывающимися половинками.
Кварцифицированная панцирь пеннатной диатомеи с двумя перекрывающимися половинками. Guinardia delicatula , диатомовая водоросль, ответственная за цветение водорослей в Северном море и Ла-Манше. [25]
Guinardia delicatula , диатомовая водоросль, ответственная за цветение водорослей в Северном море и Ла-Манше. [25] Ископаемая диатомовая водоросль
Ископаемая диатомовая водоросль Существует более 100 000 видов диатомовых водорослей , на долю которых приходится 50% первичной продукции океана.
Существует более 100 000 видов диатомовых водорослей , на долю которых приходится 50% первичной продукции океана.







Панцири диатомовых водорослей накапливались более 100 миллионов лет, оставляя по всему земному шару богатые залежи нано- и микроструктурированного оксида кремния в виде диатомовой земли . Эволюционные причины образования нано- и микроструктурированного кремнезема фотосинтезирующими водорослями еще не ясны. Однако в 2018 году было показано, что поглощение ультрафиолетового света наноструктурированным кремнеземом защищает ДНК в клетках водорослей, и это может быть эволюционной причиной образования стеклянных клеток. [28] [29]


| Внешние видео | |
|---|---|
Радиолярии
[ редактировать ]

Радиолярии — это одноклеточные хищные протисты , заключенные в сложные шаровидные раковины, обычно сделанные из кремнезема и пронизанные отверстиями. Их название происходит от латинского слова «радиус». Они ловят добычу, вытягивая части своего тела через отверстия. Как и в случае с кремнеземными панцирями диатомей, раковины радиолярий могут опускаться на дно океана, когда радиолярии умирают, и сохраняются как часть океанских отложений. Эти останки в виде микрокаменелостей предоставляют ценную информацию о прошлых условиях океана. [32]
 Как и диатомеи, радиолярии бывают разных форм.
Как и диатомеи, радиолярии бывают разных форм. Раковины радиолярий, как и диатомеи, обычно состоят из силиката.
Раковины радиолярий, как и диатомеи, обычно состоят из силиката. Однако акантарианские радиолярии имеют раковины из сульфата стронция. кристаллов
Однако акантарианские радиолярии имеют раковины из сульфата стронция. кристаллов




 Схематическая диаграмма сферической раковины радиолярий в разрезе
Схематическая диаграмма сферической раковины радиолярий в разрезе Кладококк абиетинус
Кладококк абиетинус




| Внешние видео | |
|---|---|
Скорлупы на основе кальция
[ редактировать ]Кокколитофоры
[ редактировать ]Кокколитофоры — это мелкие одноклеточные фотосинтезирующие протисты с двумя жгутиками для передвижения. Большинство из них защищено оболочкой, называемой коккосферой . Коккосферы покрыты декоративными круглыми пластинками или чешуйками, называемыми кокколитами . Кокколиты состоят из карбоната кальция. Термин кокколитофор происходит от греческого слова « камень, несущий семя» , что связано с их небольшим размером и камнями кокколита, которые они несут. При правильных условиях они цветут, как и другой фитопланктон, и могут сделать океан молочно-белым . [36]

 Кокколитофоры названы в честь документального сериала BBC «Голубая планета».
Кокколитофоры названы в честь документального сериала BBC «Голубая планета».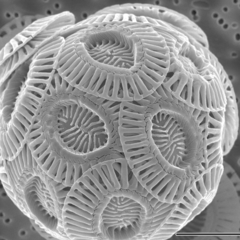 Кокколитофор . Emiliania huxleyi
Кокколитофор . Emiliania huxleyi





Есть преимущества для простейших, имеющих защитный панцирь. На диаграмме слева ниже показаны некоторые преимущества, которые кокколитофоры получают от ношения кокколитов. На диаграмме (A) представлен ускоренный фотосинтез, включая механизмы концентрации углерода (CCM) и повышенное поглощение света за счет рассеяния редких фотонов для глубоко обитающих видов. (B) представляет собой защиту от фотоповреждений, включая защиту солнцезащитного козырька от ультрафиолетового света (УФ) и фотосинтетического активного излучения (ФАР), а также рассеивание энергии в условиях яркого освещения. (C) представляет собой броневую защиту, включающую защиту от вирусных/бактериальных инфекций и выпаса селективными и неселективными травоядными. [38]

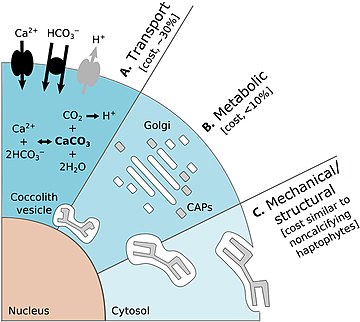
Есть также затраты на простейших, имеющих защитные панцири. Диаграмма справа выше показывает некоторые энергетические затраты, которые несет кокколитофор при переноске кокколитов. На диаграмме энергетические затраты указаны в процентах от общего фотосинтетического бюджета. (А) представляет собой транспортные процессы, включающие транспорт в клетку из окружающей морской воды субстратов первичной кальцификации Ca 2 + и HCO 3 - (черные стрелки) и удаление конечного продукта H+ из клетки (серая стрелка). Транспорт Ca 2 + через цитоплазму к везикуле кокколита (CV) является доминирующей затратой, связанной с кальцификацией. (B) представляет собой метаболические процессы, включающие синтез связанных с кокколитом полисахаридов (CAP – серые прямоугольники) комплексом Гольджи (белые прямоугольники), которые регулируют зарождение и геометрию кристаллов CaCO 3 . Завершенный кокколит (серая пластинка) представляет собой сложную структуру из сложно расположенных CAP и CaCO 3 кристаллов . (C) Механические и структурные процессы ответственны за секрецию завершенных кокколитов, которые транспортируются из своего исходного положения, прилегающего к ядру, на периферию клетки, где они переносятся на поверхность клетки. [38]
Фораминиферы
[ редактировать ]

Как и радиолярии, фораминиферы ( сокращенно форамы ) представляют собой одноклеточные хищные протисты, также защищенные раковинами с отверстиями. Их название происходит от латинского слова «носители дыр». Их оболочки, часто называемые раковинами , могут быть однокамерными или многокамерными; многокамерные отверстия по мере роста добавляют больше камер. Самые известные из них сделаны из кальцита, но пробы могут быть также изготовлены из арагонита , агглютинированных частиц осадка, хитона или (редко) кремнезема. Большинство форамов донные , но около 40 современных видов — планктонные . [39] Они широко исследуются с использованием хорошо известных летописей окаменелостей, которые позволяют ученым делать много выводов о окружающей среде и климате прошлого. [32] У некоторых фораминифер раковины вообще отсутствуют. [40]


| Внешние видео | |
|---|---|
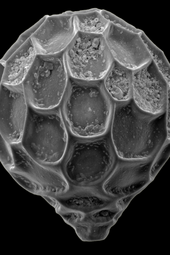
 разрез, показывающий камеры спирального отверстия
разрез, показывающий камеры спирального отверстия Живая аммиачная тепида, струящая гранулированную эктоплазму для улавливания пищи
Живая аммиачная тепида, струящая гранулированную эктоплазму для улавливания пищи Группа планктонных форм.
Группа планктонных форм. Ископаемые образования нуммулитид разных размеров из эоцена.
Ископаемые образования нуммулитид разных размеров из эоцена.![Египетские пирамиды были построены из известняка, содержащего нуммулиты.[42]](//upload.wikimedia.org/wikipedia/commons/thumb/a/af/All_Gizah_Pyramids.jpg/325px-All_Gizah_Pyramids.jpg)




![Египетские пирамиды были построены из известняка, содержащего нуммулиты.[42]](http://upload.wikimedia.org/wikipedia/commons/thumb/a/af/All_Gizah_Pyramids.jpg/325px-All_Gizah_Pyramids.jpg)
Другие снаряды
[ редактировать ]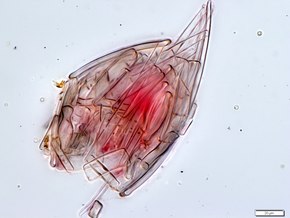
Тело клетки многих хоанофлагеллят окружено отличительным внеклеточным матриксом или перипластом . Эти клеточные покрытия сильно различаются по структуре и составу и используются систематиками для целей классификации. Многие хоанофлагелляты строят сложные корзинчатые «дома», называемые лориками , из нескольких склеенных вместе полосок кремнезема. [43] Функциональное значение перипласта неизвестно, но считается, что у сидячих организмов он способствует прикреплению к субстрату. Есть предположение, что у планктонных организмов перипласт увеличивает сопротивление, тем самым противодействуя силе, создаваемой жгутиком, и повышая эффективность питания. [43] [44]
| Внешние видео | |
|---|---|

Микрофоссилии и отложения
[ редактировать ]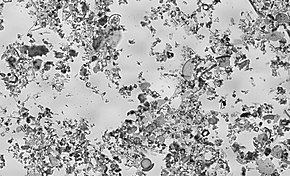
Панцири или скелеты многих протистов сохранились в геологических масштабах времени в виде микроокаменелостей. Микрофоссилии — это окаменелости , размер которых обычно составляет от 0,001 мм до 1 мм. [45] исследование которых требует применения световой или электронной микроскопии . Окаменелости, которые можно изучить невооруженным глазом или при малом увеличении, например, с помощью ручной линзы, называются макроокаменами .
Микрофоссилии являются обычным явлением в геологической летописи от докембрия до голоцена . Они наиболее распространены в морских отложениях , но также встречаются в солоноватой воде, пресной воде и наземных осадочных отложениях. все царства жизни и Хотя в летописях микроископаемых представлены протистов скелеты или цисты Chrysophyta , а , Pyrrophyta , Sarcodina , акритарх , наиболее распространенными формами являются хитинозоев также пыльца и споры сосудистых растений .
окаменелые микроорганизмы были обнаружены гидротермальных жерл В 2017 году в осадках в поясе Нуввуагиттук , или микрофоссилии, возраст которых может достигать 4,28 миллиарда лет, что является старейшим свидетельством жизни на Земле , что позволяет предположить «почти мгновенное возникновение жизни» (в геологическом смысле времени), после образования океана 4,41 миллиарда лет назад и вскоре после образования Земли 4,54 миллиарда лет назад. [46] [47] [48] [49] Тем не менее, как утверждают некоторые исследователи, жизнь могла зародиться еще раньше, почти 4,5 миллиарда лет назад. [50] [51]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Гафар, Н.А., Эйр, Б.Д. и Шульц, К.Г. (2019) «Сравнение видовой чувствительности к изменению света и химии карбонатов при кальцинировании морского фитопланктона». Научные отчеты , 9 (1): 1–12. два : 10.1038/s41598-019-38661-0 .
 Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 . - ^ «Группы протистов | Безграничная биология» . Courses.lumenlearning.com . Проверено 16 февраля 2021 г.
- ^ Кавалер-Смит Т. (декабрь 1993 г.). «Царство простейших и его 18 типов» . Микробиологические обзоры . 57 (4): 953–94. doi : 10.1128/mmbr.57.4.953-994.1993 . ПМК 372943 . ПМИД 8302218 .
- ^ Корлисс Дж.О. (1992). «Должен ли быть отдельный кодекс номенклатуры для протистов?». БиоСистемы . 28 (1–3): 1–14. дои : 10.1016/0303-2647(92)90003-H . ПМИД 1292654 .
- ^ Слапета Х., Морейра Д., Лопес-Гарсия П. (2005). «Степень разнообразия протистов: выводы из молекулярной экологии пресноводных эукариот» . Труды Королевского общества B: Биологические науки . 272 (1576): 2073–81. дои : 10.1098/rspb.2005.3195 . ПМЦ 1559898 . ПМИД 16191619 .
- ^ Морейра Д., Лопес-Гарсия П. (2002). «Молекулярная экология микробных эукариот открывает скрытый мир» (PDF) . Тенденции в микробиологии . 10 (1): 31–8. дои : 10.1016/S0966-842X(01)02257-0 . ПМИД 11755083 .
- ^ Фораминиферы: заметки для краткого курса, организованного М.А. Бузасом и Б.К. Сеном Гуптой: подготовлено для краткого курса по фораминиферам, спонсируемого Палеонтологическим обществом, проходившего в Новом Орлеане, штат Луизиана, 17 октября 1982 г. Томас В. Бродхед, Палеонтологическое общество. [Ноксвилл, Теннесси]: Университет Теннесси, факультет геологических наук. 1982. ISBN 0-910249-05-9 . OCLC 9276403 .
{{cite book}}: CS1 maint: другие ( ссылка ) - ^ Рахман, Атта-ур- (2008). «Силикон». Исследования по химии натуральных продуктов . Том. 35. с. 856. ИСБН 978-0-444-53181-0 .
- ^ Эксли, К. (1998). «Кремний в жизни: бионеорганическое решение биоорганической необходимости». Журнал неорганической биохимии . 69 (3): 139–144. дои : 10.1016/S0162-0134(97)10010-1 .
- ^ Эпштейн, Эмануэль (1999). «КРЕМНИЙ». Ежегодный обзор физиологии растений и молекулярной биологии растений . 50 : 641–664. doi : 10.1146/annurev.arplant.50.1.641 . ПМИД 15012222 .
- ^ Ким, Сан Гю; Ким, Ки У; Пак, Ын У; Чой, Дойл (2002). «Индуцированное кремнием укрепление клеточной стенки листьев риса: возможный клеточный механизм повышенной устойчивости хозяина к взрыву» . Фитопатология . 92 (10): 1095–103. дои : 10.1094/PHYTO.2002.92.10.1095 . ПМИД 18944220 .
- ^ Воздух, которым вы дышите? Диатомовая водоросль сделала это
- ^ Трегер, П.; Нельсон, DM; Ван Беннеком, AJ; Демастер, диджей; Лейнарт, А.; Кегинер, Б. (1995). «Баланс кремнезема в Мировом океане: переоценка». Наука . 268 (5209): 375–9. Бибкод : 1995Sci...268..375T . дои : 10.1126/science.268.5209.375 . ПМИД 17746543 . S2CID 5672525 .
- ^ Ирландия, Т., «Инженерия с водорослями» . Биолог , 63 (5):10.
- ^ Васильев, Мэгги (2006) «Планктон — растительный планктон» , Те Ара — Энциклопедия Новой Зеландии . Доступ: 2 ноября 2019 г.
- ^ «Королевский колледж Лондона — озеро Мегачад» . www.kcl.ac.uk. Проверено 5 мая 2018 г.
- ^ Бидл, Кей Д.; Манганелли, Маура; Азам, Фарук (6 декабря 2002 г.). «Регулирование сохранения океанического кремния и углерода путем контроля температуры бактерий» . Наука . 298 (5600): 1980–1984. Бибкод : 2002Sci...298.1980B . дои : 10.1126/science.1076076 . ISSN 0036-8075 . ПМИД 12471255 . S2CID 216994 .
- ^ Дуркин, Коллин А.; Кестер, Джули А.; Бендер, Сара Дж.; Армбруст, Э. Вирджиния (2016). «Эволюция переносчиков кремния у диатомовых водорослей» . Журнал психологии . 52 (5): 716–731. дои : 10.1111/jpy.12441 . ISSN 1529-8817 . ПМК 5129515 . ПМИД 27335204 .
- ^ Jump up to: а б Дагдейл, Колорадо; Вилкерсон, ФП (30 декабря 2001 г.). «Источники и судьба кремния в океане: роль диатомей в климате и ледниковых циклах» . Сциентия Марина . 65 (С2): 141–152. дои : 10.3989/scimar.2001.65s2141 . ISSN 1886-8134 .
- ^ Бейнс, Стивен Б.; Твининг, Бенджамин С.; Бжезинский, Марк А.; Краузе, Джеффри В.; Фогт, Стефан; Ассаэль, Дилан; Макдэниел, Ханна (декабрь 2012 г.). «Значительное накопление кремния морскими пикоцианобактериями» . Природа Геонауки . 5 (12): 886–891. Бибкод : 2012NatGe...5..886B . дои : 10.1038/ngeo1641 . ISSN 1752-0908 .
- ^ Тернер, Джефферсон Т. (январь 2015 г.). «Фекальные гранулы зоопланктона, морской снег, фитодетрит и биологический насос океана» . Прогресс в океанографии . 130 : 205–248. Бибкод : 2015Proce.130..205T . дои : 10.1016/j.pocean.2014.08.005 . ISSN 0079-6611 .
- ^ Юл, Эндрю; Тиррелл, Тоби (2003). «Роль диатомей в регулировании круговорота кремния в океане» . Глобальные биогеохимические циклы . 17 (4): н/д. Бибкод : 2003GBioC..17.1103Y . дои : 10.1029/2002GB002018 . ISSN 1944-9224 . S2CID 16849373 .
- ^ Трегер, Поль Дж.; Де Ла Роша, Кристина Л. (3 января 2013 г.). «Кремнеземный цикл Мирового океана». Ежегодный обзор морской науки . 5 (1): 477–501. doi : 10.1146/annurev-marine-121211-172346 . ПМИД 22809182 .
- ^ Маринов И.; Гнанадэсикан, А.; Тоггвайлер-младший; Сармьенто, JL (июнь 2006 г.). «Биогеохимический разрыв Южного океана». Природа . 441 (7096): 964–967. Бибкод : 2006Natur.441..964M . дои : 10.1038/nature04883 . ПМИД 16791191 . S2CID 4428683 .
- ^ Арсеньев Л.; Саймон, Н.; Риго-Жалабер, Ф.; Ле Галль, Ф.; Шаффрон, С.; Корре, Э.; Приходить.; Бигерд, Э.; Боду, AC (2018). «Первые вирусы, заражающие морскую диатомовую водоросль Guinardia delicatula » . Границы микробиологии . 9 : 3235. дои : 10.3389/fmicb.2018.03235 . ПМК 6334475 . ПМИД 30687251 .
- ^ Чжан, Д.; Ван, Ю.; Кай, Дж.; Пан, Дж.; Цзян, X.; Цзян, Ю. (2012). «Технология биопроизводства на основе диатомовой микро- и наноструктуры» . Китайский научный бюллетень . 57 (30): 3836–3849. Бибкод : 2012ЧСБу..57.3836З . дои : 10.1007/s11434-012-5410-x .
- ^ Биоразлагаемый блеск и микроводоросли, пожирающие загрязнения: новые материалы, вдохновленные природой Horizon , 28 мая 2020 г.
- ^ Jump up to: а б Агирре Л.Е., Оуян Л., Эльфвинг А., Хедблом М., Вульф А. и Инганес О. (2018) «Панцири диатомовых водорослей защищают ДНК от ультрафиолетового света». Научные отчеты , 8 (1): 1–6. два : 10.1038/s41598-018-21810-2 . Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Де Томмаси, Э., Конгестри, Р., Дардано, П., Де Лука, А.С., Манаго, С., Ри, И. и Де Стефано, М. (2018) «УФ-защита и преобразование длины волны центрическими диатомовыми атомами панцири с наноузорами». Научные отчеты , 8 (1): 1–14. doi : 10.1038/s41598-018-34651-w . Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Бут, Британская Колумбия и Марчант, HJ (1987) «Parmales, новый порядок морских хризофитов, с описаниями трех новых родов и семи новых видов». Журнал Phycology , 23 : 245–260. дои : 10.1111/j.1529-8817.1987.tb04132.x .
- ^ Кувата, А., Ямада, К., Ичиномия, М., Ёсикава, С., Трагин, М., Ваулот, Д. и Лопес душ Сантос, А. (2018) «Bolidophyceae, сестринская пикопланктонная группа диатомей - обзор». Границы морской науки , 5 : 370. два : 10.3389/fmars.2018.00370 . Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Jump up to: а б Васильев, Мэгги (2006) «Планктон — животный планктон» , Те Ара — Энциклопедия Новой Зеландии . Доступ: 2 ноября 2019 г.
- ^ Качович, Сара (2018) «Разум важнее методов: связь микрокаменелостей с тектоникой» Блог Отдела тектоники и структурной геологии Европейского союза геонаук.
- ^ Качович С., Шэн Дж. и Эйчисон Дж. К., 2019. Добавление нового измерения к исследованиям ранней эволюции радиолярий. Научные отчеты, 9 (1), стр. 1–10. два : 10.1038/s41598-019-42771-0 . Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Варя, К.; Арагон, JL; Баррио, РА (1999). «Узоры Тьюринга на сфере». Физический обзор E . 60 (4): 4588–4592. Бибкод : 1999PhRvE..60.4588V . дои : 10.1103/PhysRevE.60.4588 . ПМИД 11970318 .
- ^ Васильев, Мэгги (2006) «Кокколитофор» , Те Ара — Энциклопедия Новой Зеландии . Доступ: 2 ноября 2019 г.
- ^ Рост, Б. и Рибеселл, У. (2004) «Кокколитофоры и биологический насос: реакция на изменения окружающей среды». В: Кокколитофоры: от молекулярных процессов к глобальному воздействию , страницы 99–125, Springer. ISBN 9783662062784 .
- ^ Jump up to: а б с д Монтейро Ф.М., Бах Л.Т., Браунли К., Баун П., Рикаби Р.Э., Поултон А.Дж., Тиррелл Т., Бофорт Л., Дуткевич С., Гиббс С. и Гутовска М.А. ( 2016) «Почему кальцинируется морской фитопланктон». Science Advances , 2 (7): e1501822. два : 10.1126/sciadv.1501822 . Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Хемлебен, К.; Андерсон, Орегон; Шпиндлер, М. (1989). Современные планктонные фораминиферы . Спрингер-Верлаг. ISBN 978-3-540-96815-3 .
- ^ Павловский, Ян; Боливар, Игнасио; Фарни, Хосе Ф.; Варгас, Коломбан Де; Баузер, Сэмюэл С. (ноябрь 1999 г.). «Молекулярные доказательства того, что Reticulomyxa Filosa является пресноводным голым фораминифером» . Журнал эукариотической микробиологии . 46 (6): 612–617. дои : 10.1111/j.1550-7408.1999.tb05137.x . ISSN 1066-5234 . ПМИД 10568034 . S2CID 36497475 .
- ^ Favulina hexagona Европейский союз геонаук, 9 ноября 2020 г.
- ^ Фораминиферы: История изучения , Университетский колледж Лондона . Проверено: 18 ноября 2019 г.
- ^ Jump up to: а б Ледбитер Б.С., Томсен Х (2000). «Отряд Хоанофлагеллиды». Иллюстрированный справочник по простейшим, второе издание. Лоуренс: Общество протозоологов . 451 : 14–38.
- ^ Ледбитер Б.С., Келли М. (2001). «Эволюция хоанофлагеллят и губок животных». Вода и атмосфера онлайн . 9 (2): 9–11.
- ^ Древес, Чарли. «Открытие девонских микрокаменелостей» . Университет штата Айова . Проверено 4 марта 2017 г.
- ^ Додд, Мэтью С.; Папино, Доминик; Гренн, Тор; слабый, Джон Ф.; Риттнер, Мартин; Пирайно, Франко; О'Нил, Джонатан; Литтл, Криспин Т.С. (2 марта 2017 г.). «Доказательства ранней жизни в осадках старейших гидротермальных источников Земли» (PDF) . Природа . 543 (7643): 60–64. Бибкод : 2017Natur.543...60D . дои : 10.1038/nature21377 . ПМИД 28252057 .
- ^ Циммер, Карл (1 марта 2017 г.). «Ученые говорят, что окаменелости канадских бактерий могут быть древнейшими на Земле» . Нью-Йорк Таймс . Проверено 2 марта 2017 г.
- ^ Гош, Паллаб (1 марта 2017 г.). «Обнаружены самые ранние свидетельства жизни на Земле» . Новости Би-би-си . Проверено 2 марта 2017 г.
- ^ Данэм, Уилл (1 марта 2017 г.). «Канадские окаменелости, похожие на бактерии, названы древнейшими свидетельствами жизни» . Рейтер . Проверено 1 марта 2017 г.
- ^ Персонал (20 августа 2018 г.). «Временная шкала возникновения и эволюции всей жизни на Земле» . Физика.орг . Проверено 20 августа 2018 г.
- ^ Беттс, Холли С.; Путик, Марк Н.; Кларк, Джеймс В.; Уильямс, Том А.; Донохью, Филип CJ; Пизани, Давиде (20 августа 2018 г.). «Комплексные геномные и ископаемые данные проливают свет на раннюю эволюцию жизни и происхождение эукариот» . Природа . 2 (10): 1556–1562. дои : 10.1038/s41559-018-0644-x . ПМК 6152910 . ПМИД 30127539 .
Дальнейшие ссылки
[ редактировать ]- Сюй К., Хатчинс Д. и Гао К. (2018) «Расположение кокколита следует эйлеровой математике в кокколитофоре Emiliania huxleyi ». PeerJ , 6 : e4608. дои : 10.1126/science.aaa7378 .
- Протистанские скелеты: геологическая история эволюции и ограничений