Фитопланктон
| Часть серии о |
| Планктон |
|---|
 |
Фитопланктон ( / ˌ f aɪ t oʊ ˈ p l æ ŋ k t ə n / ) — автотрофные (самопитающиеся) компоненты планктонного сообщества и ключевая часть океанических и пресноводных экосистем . Название происходит от греческих слов φυτόν ( фитон ), что означает « растение », и πλαγκτός ( планктос ), что означает «странник» или «бродяга». [1] [2] [3]
Фитопланктон получает энергию посредством фотосинтеза , как это делают деревья и другие растения на суше. Это означает, что фитопланктон должен получать солнечный свет, поэтому они живут в хорошо освещенных поверхностных слоях ( эуфотической зоне ) океанов и озер. По сравнению с наземными растениями фитопланктон распространен на большей площади поверхности, подвержен меньшим сезонным колебаниям и имеет значительно более высокую скорость оборота, чем деревья (дни по сравнению с десятилетиями). В результате фитопланктон быстро реагирует в глобальном масштабе на изменения климата.
Фитопланктон составляет основу морских и пресноводных пищевых сетей и является ключевым игроком в глобальном углеродном цикле . На их долю приходится около половины глобальной фотосинтетической активности и, по крайней мере, половина производства кислорода, несмотря на то, что они составляют лишь около 1% мировой биомассы растений. Фитопланктон очень разнообразен: от фотосинтезирующих бактерий до растительноподобных водорослей и панцирных кокколитофоров . Важные группы фитопланктона включают диатомовые водоросли , цианобактерии и динофлагелляты , хотя представлены и многие другие группы. [2]
Большинство фитопланктона слишком малы, чтобы их можно было увидеть по отдельности невооруженным глазом . Однако, когда они присутствуют в достаточно большом количестве, некоторые разновидности могут быть заметны в виде цветных пятен на поверхности воды из-за присутствия хлорофилла в их клетках и дополнительных пигментов (таких как фикобилипротеины или ксантофиллы ) у некоторых видов.
Типы
[ редактировать ]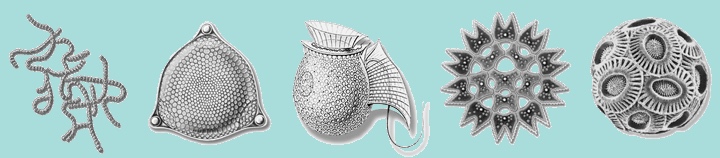
Слева направо: цианобактерии , диатомовые водоросли , динофлагелляты , зеленые водоросли и кокколитофоры.
Фитопланктон – фотосинтезирующие микроскопические протисты и бактерии, населяющие верхний освещенный солнцем слой морских и пресных водоемов на Земле. Параллельно с растениями на суше фитопланктон осуществляет первичную продукцию в воде. [2] создание органических соединений из углекислого газа растворенного в воде . Фитопланктон формирует основу и поддерживает водную пищевую сеть . [4] Земли и являются ключевыми игроками в углеродном цикле . [5]
«В морском фотосинтезе доминируют микроводоросли , которые вместе с цианобактериями называются фитопланктоном». [6] Фитопланктон чрезвычайно разнообразен: от фотосинтезирующих бактерий (цианобактерий) до растительноподобных диатомей и панцирных кокколитофоров. [7] [2]

фитопланктона

бронированы

Экология
[ редактировать ]Фитопланктон получает энергию в процессе фотосинтеза и ) поэтому должен жить в хорошо освещенном поверхностном слое (называемом эвфотической зоной океана , моря , озера или другого водоема. На фитопланктон приходится около половины всей фотосинтетической деятельности на Земле. [9] [10] [11] Их совокупная фиксация энергии в соединениях углерода ( первичное производство ) является основой подавляющего большинства океанических, а также многих пресноводных пищевых сетей ( хемосинтез является заметным исключением).
Хотя почти все виды фитопланктона являются облигатными фотоавтотрофами , есть некоторые миксотрофные и другие, непигментированные виды , которые на самом деле являются гетеротрофными (последние часто рассматриваются как зоопланктон ). [2] [12] Из них наиболее известны динофлагеллят, роды такие как Noctiluca и Dinophys , которые получают органический углерод другие , поглощая организмы или детритный материал.
Фитопланктон обитает в фотической зоне океана, где фотосинтез возможен . В ходе фотосинтеза они усваивают углекислый газ и выделяют кислород. Если солнечная радиация слишком высока, фитопланктон может стать жертвой фотодеградации . Виды фитопланктона обладают большим разнообразием фотосинтетических пигментов , которые в зависимости от вида позволяют им поглощать различные длины волн переменного подводного света. [13] Это означает, что разные виды могут по-разному эффективно использовать длину волны света, и свет является не одним экологическим ресурсом , а множеством ресурсов в зависимости от его спектрального состава. [14] Тем самым было обнаружено, что изменения в спектре света сами по себе могут изменить естественные сообщества фитопланктона, даже если та же интенсивность . доступна [15] Рост клеток фитопланктона дополнительно зависит от питательных веществ, которые поступают в океан с реками, континентальным выветриванием и талой ледниковой водой на полюсах. Фитопланктон выпускает растворенный органический углерод (DOC) в океан. Поскольку фитопланктон является основой морских пищевых сетей , они служат добычей зоопланктона , личинок рыб и других гетеротрофных организмов. Они также могут разрушаться бактериями или вирусным лизисом . Хотя некоторые клетки фитопланктона, например динофлагелляты , способны мигрировать вертикально, они все же неспособны активно двигаться против течений, поэтому медленно тонут и в конечном итоге удобряют морское дно мертвыми клетками и детритом . [16]

Фитопланктон в решающей степени зависит от ряда питательных веществ . В первую очередь это макроэлементы, такие как нитрат , фосфат или кремниевая кислота , которые необходимы для роста в относительно больших количествах. Их наличие на поверхности океана регулируется балансом между так называемым биологическим насосом и подъемом глубинных, богатых питательными веществами вод. Стехиометрический , состав питательных веществ фитопланктона определяет – и определяется – соотношение макронутриентов Редфилда обычно доступных на поверхности океана. Однако на больших территориях океанов, таких как Южный океан , фитопланктон ограничен из-за нехватки микроэлемента железа . Это привело к тому, что некоторые ученые стали выступать за внесение железных удобрений как средство противодействия накоплению антропогенного углекислого газа (CO 2 ) в атмосфере . [17] Крупномасштабные эксперименты добавили железо (обычно в виде солей, таких как сульфат железа ) в океаны, чтобы способствовать росту фитопланктона и привлечь атмосферный CO 2 в океан. Споры по поводу управления экосистемой и эффективности внесения железных удобрений замедлили проведение таких экспериментов. [18]
фитопланктона зависит от витаминов группы B. Выживание Было установлено, что в районах океана наблюдается серьезная нехватка некоторых витаминов группы B и, соответственно, фитопланктона. [19]
Влияние антропогенного потепления на глобальную популяцию фитопланктона является областью активных исследований. Ожидается, что изменения в вертикальной стратификации водной толщи, скорости температурно-зависимых биологических реакций и поступлении питательных веществ из атмосферы окажут важное влияние на будущую продуктивность фитопланктона. [20] [21]

Значительное внимание также уделяется влиянию антропогенного закисления океана на рост фитопланктона и структуру сообществ. Клетки кокколитофорного фитопланктона обычно покрыты оболочкой из карбоната кальция, называемой коккосферой , которая чувствительна к закислению океана. Данные свидетельствуют о том, что из-за короткого времени генерации некоторые фитопланктоны могут адаптироваться к изменениям pH, вызванным увеличением углекислого газа, в короткие сроки (от месяцев до лет). [22] [23]
Фитопланктон служит основой водной пищевой сети, обеспечивая важную экологическую функцию для всей водной жизни. В будущих условиях антропогенного потепления и закисления океана изменения смертности фитопланктона за счет изменения темпов выпаса зоопланктона могут быть значительными. [24] Одна из многих пищевых цепей в океане – примечательная из-за небольшого количества звеньев – это цепь фитопланктона, поддерживающего криль ( ракообразное, похожее на крошечную креветку), который, в свою очередь, поддерживает усатых китов .
Циклы Эль-Ниньо-Южного колебания (ЭНСО) в экваториальной части Тихого океана могут повлиять на фитопланктон. [25] Биохимические и физические изменения во время циклов ЭНСО изменяют структуру сообщества фитопланктона. [25] Кроме того, могут произойти изменения в структуре фитопланктона, такие как значительное снижение биомассы и плотности фитопланктона, особенно во время фаз Эль-Ниньо. [26] Чувствительность фитопланктона к изменениям окружающей среды является причиной того, что его часто используют в качестве индикаторов экологического состояния и здоровья эстуариев и прибрежных районов. [27] Для изучения этих событий используются спутниковые наблюдения за цветом океана. Спутниковые снимки помогают лучше понять их глобальное распространение. [25]
Разнообразие
[ редактировать ]


Термин «фитопланктон» охватывает все фотоавтотрофные микроорганизмы в водных пищевых цепях . Однако, в отличие от наземных сообществ , где большинство автотрофов являются растениями , фитопланктон представляет собой разнообразную группу, включающую протистан -эукариоты , а также эубактериальные и архебактериальные прокариоты . Известно около 5000 видов морского фитопланктона. [28] Как такое разнообразие развивалось, несмотря на скудность ресурсов (ограничивающую дифференциацию ниш ), неясно. [29]
По численности наиболее важные группы фитопланктона включают диатомовые водоросли , цианобактерии и динофлагелляты многие другие группы водорослей , хотя представлены и . Одна группа, кокколитофориды , ответственна (частично) за выброс значительных количеств диметилсульфида (ДМС) в атмосферу . DMS окисляется с образованием сульфата, который в районах с низкой концентрацией аэрозольных частиц в окружающей среде может способствовать заселению ядер конденсации облаков , что в основном приводит к увеличению облачного покрова и альбедо облаков в соответствии с так называемой гипотезой CLAW . [30] [31] Различные типы фитопланктона поддерживают разные трофические уровни в разных экосистемах. В олиготрофных океанических регионах, таких как Саргассово море или южный Тихоокеанский круговорот , в фитопланктоне преобладают клетки небольшого размера, называемые пикопланктоном и нанопланктоном (также называемые пикофлагеллятами и нанофлагеллятами), в основном состоящие из цианобактерий ( Prochromococcus , Synechococcus ) и пикоэукариотов, таких как как Микромонас . В более продуктивных экосистемах, где преобладает апвеллинг или интенсивное воздействие наземных веществ, более крупные динофлагелляты являются более доминирующим фитопланктоном и отражают большую часть биомассы . [32]
Стратегии роста
[ редактировать ]В начале двадцатого века Альфред К. Редфилд обнаружил сходство элементного состава фитопланктона с основными растворенными питательными веществами в глубинах океана. [33] Редфилд предположил, что соотношение углерода, азота и фосфора (106:16:1) в океане контролируется потребностями фитопланктона, поскольку фитопланктон впоследствии выделяет азот и фосфор по мере их реминерализации. Это так называемое « отношение Редфилда » при описании стехиометрии фитопланктона и морской воды стало фундаментальным принципом для понимания морской экологии, биогеохимии и эволюции фитопланктона. [34] Однако коэффициент Редфилда не является универсальной величиной и может отклоняться из-за изменений в доставке экзогенных питательных веществ. [35] и микробный метаболизм в океане, такой как фиксация азота , денитрификация и анаммокс .
Динамическая стехиометрия, показанная у одноклеточных водорослей, отражает их способность хранить питательные вещества во внутреннем пуле, переключаться между ферментами с различными потребностями в питательных веществах и изменять состав осмолитов. [36] [37] Различные клеточные компоненты имеют свои уникальные характеристики стехиометрии. [34] например, механизмы получения ресурсов (света или питательных веществ), таких как белки и хлорофилл, содержат высокую концентрацию азота, но низкую концентрацию фосфора. Между тем, механизмы роста, такие как рибосомальная РНК, содержат высокие концентрации азота и фосфора.
В зависимости от распределения ресурсов фитопланктон подразделяется на три различные стратегии роста: выживаемость, цветение. [38] и универсалист. Фитопланктон, способствующий выживанию, имеет высокое соотношение N:P (>30) и содержит множество механизмов добывания ресурсов для поддержания роста в условиях ограниченных ресурсов. Фитопланктон Bloomer имеет низкое соотношение N:P (<10), содержит высокую долю механизмов роста и адаптирован к экспоненциальному росту. Фитопланктон общего назначения имеет соотношение N:P, аналогичное соотношению Редфилда, и содержит относительно равные механизмы приобретения ресурсов и роста.
Факторы, влияющие на численность
[ редактировать ]Исследование NAAMES представляло собой пятилетнюю программу научных исследований, проводимую в период с 2015 по 2019 год учеными из Университета штата Орегон и НАСА с целью изучения аспектов динамики фитопланктона в океанских экосистемах и того, как такая динамика влияет на атмосферные аэрозоли , облака и климат (NAAMES означает Исследование аэрозолей и морских экосистем Северной Атлантики). Исследование было сосредоточено на субарктическом регионе северной части Атлантического океана, который является местом одного из крупнейших повторяющихся цветений фитопланктона на Земле. Долгая история исследований в этом месте, а также относительная легкость доступа сделали Северную Атлантику идеальным местом для проверки преобладающих научных гипотез. [39] в попытке лучше понять роль аэрозольных выбросов фитопланктона в энергетическом бюджете Земли. [40]
NAAMES был разработан для определения конкретных фаз годового цикла фитопланктона: минимума, кульминации и промежуточного снижения и увеличения биомассы, чтобы разрешить споры о сроках формирования цветения и закономерностях, способствующих возобновлению ежегодного цветения. [40] Проект NAAMES также исследовал количество, размер и состав аэрозолей, образующихся в результате первичного производства , чтобы понять, как циклы цветения фитопланктона влияют на образование облаков и климат. [41]



Факторы, влияющие на производительность
[ редактировать ]
Фитопланктон — ключевые медиаторы биологического насоса . Понимание реакции фитопланктона на изменение условий окружающей среды является необходимым условием для прогнозирования будущих концентраций CO 2 в атмосфере . Температура, освещенность и концентрация питательных веществ, а также CO 2 являются основными факторами окружающей среды, влияющими на физиологию и стехиометрию фитопланктона. [47] Стехиометрия или элементный состав фитопланктона имеет первостепенное значение для вторичных производителей, таких как копеподы, рыба и креветки, поскольку он определяет качество питания и влияет на поток энергии через морские пищевые цепи . [48] Изменение климата может существенно изменить структуру сообществ фитопланктона, что приведет к каскадным последствиям для морских пищевых сетей , тем самым изменяя количество углерода, переносимого в недра океана. [49] [45]
Диаграмма справа дает обзор различных факторов окружающей среды, которые вместе влияют на продуктивность фитопланктона . Ожидается, что все эти факторы претерпят значительные изменения в будущем океане из-за глобальных изменений. [50] Моделирование глобального потепления предсказывает повышение температуры океана; резкие изменения в стратификации океана , циркуляции и изменениях в облачном покрове и морском льду, что приводит к увеличению поступления света на поверхность океана. Кроме того, по прогнозам, сокращение поступления питательных веществ будет происходить одновременно с подкислением и потеплением океана из-за усиления стратификации водной толщи и уменьшения смешивания питательных веществ из глубокой воды с поверхностью. [51] [45]
Роль фитопланктона
[ редактировать ]
На диаграмме справа отсеки, на которые влияет фитопланктон, включают состав атмосферного газа, неорганические питательные вещества и потоки микроэлементов, а также перенос и круговорот органических веществ посредством биологических процессов. Фотосинтетически зафиксированный углерод быстро перерабатывается и повторно используется на поверхности океана, а определенная часть этой биомассы экспортируется в виде тонущих частиц в глубины океана, где подвергается постоянным процессам трансформации, например реминерализации. [52]
Фитопланктон вносит вклад не только в основную пелагическую морскую пищевую сеть, но и в микробный цикл. Фитопланктон является основой морской пищевой сети и, поскольку он не зависит от других организмов в качестве источника питания, он составляет первый трофический уровень. Такие организмы, как зоопланктон, питаются этим фитопланктоном, которым питаются другие организмы, и так далее, пока не будет достигнут четвертый трофический уровень с высшими хищниками. Примерно 90% общего углерода теряется между трофическими уровнями за счет дыхания, детрита и растворенных органических веществ. Это делает процесс реминерализации и круговорот питательных веществ, осуществляемый фитопланктоном и бактериями, важными для поддержания эффективности. [53]
Цветение фитопланктона, при котором вид быстро размножается в условиях, благоприятных для роста, может вызвать вредное цветение водорослей (ВЦВ).
Аквакультура
[ редактировать ]Фитопланктон является ключевым продуктом питания как в аквакультуре , так и в марикультуре . Оба используют фитопланктон в качестве пищи для выращиваемых животных. В марикультуре фитопланктон встречается в природе и попадает в вольеры с нормальной циркуляцией морской воды. В аквакультуре фитопланктон необходимо получать и внедрять напрямую. Планктон можно либо собрать из водоема, либо культивировать, хотя первый метод используется редко. Фитопланктон используется в качестве корма для производства коловраток . [54] которые, в свою очередь, используются для питания других организмов. Фитопланктон также используется в качестве корма для многих разновидностей аквакультурных моллюсков , включая жемчужных устриц и гигантских моллюсков . В исследовании 2018 года была оценена пищевая ценность природного фитопланктона с точки зрения углеводов, белков и липидов в мировом океане с использованием данных о цвете океана, полученных со спутников. [55] и обнаружили, что теплотворная способность фитопланктона значительно различается в разных регионах океана и в разное время года. [55] [56]
Производство фитопланктона в искусственных условиях само по себе является формой аквакультуры. Фитопланктон культивируют для различных целей, в том числе в качестве корма для других аквакультурных организмов. [54] пищевая добавка для беспозвоночных, содержащихся в аквариумах . Размеры культур варьируются от небольших лабораторных культур объемом менее 1 л до нескольких десятков тысяч литров для коммерческой аквакультуры. [54] Независимо от размера культуры необходимо обеспечить определенные условия для эффективного роста планктона. Большая часть культивируемого планктона является морским, и морскую воду с удельным весом в качестве питательной среды можно использовать от 1,010 до 1,026. Эта вода должна быть стерилизована , обычно либо при высоких температурах в автоклаве , либо при воздействии ультрафиолетового излучения , чтобы предотвратить биологическое загрязнение культуры. В питательную среду добавляют различные удобрения , способствующие росту планктона. Культуру необходимо аэрировать или каким-либо образом перемешивать, чтобы планктон оставался во взвешенном состоянии, а также чтобы обеспечить растворенный углекислый газ для фотосинтеза . Помимо постоянной аэрации, большинство культур смешивают вручную или регулярно перемешивают. Необходимо обеспечить свет для роста фитопланктона. Цветовая температура освещения должна составлять примерно 6500 К, но успешно используются значения от 4000 К до более 20 000 К. Продолжительность светового воздействия должна составлять примерно 16 часов ежедневно; это наиболее эффективная искусственная длина дня. [54]
Антропогенные изменения
[ редактировать ]
Морской фитопланктон осуществляет половину глобальной фотосинтетической фиксации CO 2 (чистая глобальная первичная продукция ~ 50 Пг C в год) и половину производства кислорода, несмотря на то, что он составляет лишь ~ 1% глобальной биомассы растений. [57] По сравнению с наземными растениями морской фитопланктон распространен на большей площади поверхности, подвержен меньшим сезонным колебаниям и имеет значительно более высокую скорость оборота, чем деревья (дни по сравнению с десятилетиями). [57] Таким образом, фитопланктон быстро реагирует в глобальном масштабе на изменения климата. Эти характеристики важны при оценке вклада фитопланктона в фиксацию углерода и прогнозировании того, как это производство может измениться в ответ на возмущения. Прогнозирование воздействия изменения климата на первичную продуктивность осложняется циклами цветения фитопланктона, на которые влияет как контроль снизу вверх (например, наличие необходимых питательных веществ и вертикальное смешивание), так и контроль сверху вниз (например, выпас скота и вирусы). [58] [57] [59] [60] [61] [62] Увеличение солнечной радиации, температуры и поступления пресной воды в поверхностные воды усиливает стратификацию океана и, как следствие, уменьшает перенос питательных веществ из глубинных вод в поверхностные, что снижает первичную продуктивность. [57] [62] [63] И наоборот, повышение уровня CO 2 может увеличить первичную продукцию фитопланктона, но только тогда, когда питательные вещества не являются лимитирующими. [64] [65] [66] [24]
Некоторые исследования показывают, что общая глобальная плотность океанического фитопланктона снизилась за последнее столетие. [67] но эти выводы были поставлены под сомнение из-за ограниченной доступности долгосрочных данных по фитопланктону, методологических различий в сборе данных и большой годовой и десятилетней изменчивости продукции фитопланктона. [68] [69] [70] [71] Более того, другие исследования предполагают глобальное увеличение производства океанического фитопланктона. [72] и изменения в конкретных регионах или конкретных группах фитопланктона. [73] [74] Глобальный индекс морского льда снижается. [75] что приводит к более высокому проникновению света и потенциально большему количеству первичной продукции; [76] однако существуют противоречивые прогнозы о влиянии переменных моделей смешивания и изменений в снабжении питательными веществами, а также о тенденциях продуктивности в полярных зонах. [62] [24]
Влияние антропогенного изменения климата на биоразнообразие фитопланктона до конца не изучено. Если выбросы парниковых газов продолжат расти до высоких уровней к 2100 году, некоторые модели фитопланктона предсказывают увеличение видового богатства или количества различных видов на данной территории. Такое увеличение разнообразия планктона связано с повышением температуры океана. Ожидается, что помимо изменения видового богатства места распространения фитопланктона сместятся к полюсам Земли. Такое перемещение может нарушить экосистемы, поскольку фитопланктон потребляется зоопланктоном, который, в свою очередь, поддерживает рыболовство. Этот сдвиг в местоположении фитопланктона может также снизить способность фитопланктона накапливать углерод, выброшенный в результате деятельности человека. Человеческие (антропогенные) изменения фитопланктона влияют как на природные, так и на экономические процессы. [77]
См. также
[ редактировать ]- Альгакультура - аквакультура, связанная с выращиванием водорослей.
- AlgaeBase - База данных видов
- Цветение водорослей - распространение планктонных водорослей в воде.
- Бактериопланктон - бактериальный компонент планктона, дрейфующий в толще воды.
- Биологический насос – Процесс улавливания углерода в океанах
- Гипотеза CLAW - предполагаемая петля отрицательной обратной связи, соединяющая морскую биоту и климат.
- Критическая глубина
- Глубокий хлорофилловый максимум
- Пресноводный фитопланктон - Фитопланктон, встречающийся в пресноводных экосистемах.
- Железное удобрение – Экологическая концепция
- Микрофиты – микроскопические водоросли (микроводорослей).
- ИМЕНА
- Закисление океана – снижение уровня pH в океане.
- Парадокс планктона . Экологическое наблюдение высокого разнообразия планктона, несмотря на конкуренцию за ограниченные ресурсы.
- Фотосинтетический пикопланктон - группа фотосинтезирующего планктона.
- Событие Уайтинга – взвешивание мелкозернистых частиц карбоната кальция в водоемах.
- Тонкие слои (океанография) - Скопления планктона.
Ссылки
[ редактировать ]- ^ Турман, Х.В. (2007). Вводная океанография . Академические интернет-издатели. ISBN 978-1-4288-3314-2 . [ нужна страница ]
- ^ Jump up to: а б с д и Пьерелла Карлушич, Хуан Хосе; Ибарбальц, Федерико М.; Боулер, Крис (3 января 2020 г.). «Фитопланктон в Тарском океане» . Ежегодный обзор морской науки . 12 (1): 233–265. Бибкод : 2020ARMS...12..233P . doi : 10.1146/annurev-marine-010419-010706 . ISSN 1941-1405 . ПМИД 31899671 . S2CID 209748051 .
- ^ Пьерелла Карлушич, Хуан Хосе; Ибарбальц, Федерико М; Боулер, Крис (2020). «Исследование морского фитопланктона: от исторического понимания до эпохи омики» . Журнал исследований планктона . 42 : 595–612. дои : 10.1093/plankt/fbaa049 . hdl : 11336/143676 .
- ^ Госал; Роджерс; Рэй, С.; М.; А. «Влияние турбулентности на фитопланктон» . Предприятие аэрокосмической техники . НТРС . Проверено 16 июня 2011 г.
{{cite web}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Jump up to: а б Смоделированные сообщества фитопланктона в глобальном океане Hyperwall НАСА , 30 сентября 2015 г.
 В данную статью включен текст из этого источника, находящегося в свободном доступе .
В данную статью включен текст из этого источника, находящегося в свободном доступе . - ^ Паркер, Микаэла С.; Мок, Томас; Армбруст, Э. Вирджиния (2008). «Геномный взгляд на морские микроводоросли». Ежегодный обзор генетики . 42 : 619–645. дои : 10.1146/annurev.genet.42.110807.091417 . ПМИД 18983264 .
- ^ Линдси, Р.; Скотт, М.; Симмон, Р. (16 июля 2010 г.). «Что такое фитопланктон?» . Земная обсерватория НАСА .
- ^ «Проект Дарвина Массачусетского технологического института» . Массачусетский технологический институт .
- ^ Майкл Дж. Беренфельд; и др. (30 марта 2001 г.). «Первичное производство биосферы во время перехода ЭНСО» (PDF) . Наука . 291 (5513): 2594–7. Бибкод : 2001Sci...291.2594B . дои : 10.1126/science.1055071 . ПМИД 11283369 . S2CID 38043167 .
- ^ «Спутник НАСА обнаруживает красное свечение для составления карты состояния глобальных океанских растений». Архивировано 10 апреля 2021 года в Wayback Machine NASA , 28 мая 2009 года.
- ^ «Спутник видит рост океанских растений и озеленение побережий» . НАСА . 2 марта 2005 г. Архивировано из оригинала 29 октября 2011 г. Проверено 9 июня 2014 г.
- ^ Митра, Адите; Флинн, Кевин Дж.; Тильманн, Урбан; Рэйвен, Джон А.; Кэрон, Дэвид; Стокер, Дайан К.; Нет, Фабрис; Хансен, Пер Дж.; Халлеграефф, Густав; Сандерс, Роберт; Уилкен, Сюзанна; Макманус, Джордж; Джонсон, Мэтью; Питта, Параскеви; Воге, Селина; Берге, Терье; Кальбет, Альберт; Тингстад, Фреде; Чон, Хэ Джин; Беркхолдер, Джоанн; Глиберт, Патрисия М .; Гранели, Эдна; Лундгрен, Вероника (1 апреля 2016 г.). «Определение функциональных групп планктонных протистов по механизмам получения энергии и питательных веществ: включение разнообразных миксотрофов» . Протист . 167 (2): 106–120. дои : 10.1016/j.protis.2016.01.003 . hdl : 10261/131722 . ISSN 1434-4610 . ПМИД 26927496 .
- ^ Кирк, Джон Т.О. (1994). Свет и фотосинтез в водных экосистемах (2-е изд.). Кембридж: Издательство Кембриджского университета. дои : 10.1017/cbo9780511623370 . ISBN 9780511623370 .
- ^ Стомп, Маайке; Хьюсман, Джефф; де Йонг, Флорис; Вераарт, Аннелис Дж.; Герла, Даан; Райкебур, Махтелд; Ибелингс, Бас В.; Волленцин, Уте, ИА; Стабильный, Лукас Дж. (ноябрь 2004 г.). «Адаптивная дивергенция пигментного состава способствует биоразнообразию фитопланктона» . Природа . 432 (7013): 104–107. Стартовый код : 2004Natur.432..104S . дои : 10.1038/nature03044 . ISSN 1476-4687 . ПМИД 15475947 . S2CID 4409758 .
- ^ Хинц, Нильс Хендрик; Цейзинг, Мориц; Стрибель, Марен (2021). «Изменения спектрального качества подводного света меняют состав сообщества фитопланктона» . Лимнология и океанография . 66 (9): 3327–3337. Бибкод : 2021LimOc..66.3327H . дои : 10.1002/lno.11882 . ISSN 1939-5590 . S2CID 237849374 .
- ^ Jump up to: а б Кезе Л., Гойер Дж.К. (2018) «Реакция фитопланктона на изменение морского климата – введение» . В книге Юнгблут С., Либих В., Боде М. (редакторы) YOUMARES 8 – Океаны через границы: учимся друг у друга , страницы 55–72, Springer. дои : 10.1007/978-3-319-93284-2_5 .
 Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 . - ^ Рихтель, М. (1 мая 2007 г.). «Привлечение планктона для борьбы с глобальным потеплением» . Нью-Йорк Таймс .
- ^ Монастерский, Ричард (1995). «Железо против парника: океанографы осторожно изучают метод лечения глобального потепления». Новости науки . 148 (14): 220–1. дои : 10.2307/4018225 . JSTOR 4018225 .
- ^ Саньюдо-Вильгельми, Серхио (23 июня 2012 г.). «Существование витаминных «пустынь» в океане подтверждено» . ScienceDaily .
- ^ Хенсон, ЮАР; Сармьенто, JL; Данн, JP; Бопп, Л.; Лима, И.; Дони, Южная Каролина ; Джон, Дж.; Болье, К. (2010). «Обнаружение антропогенного изменения климата по спутниковым записям хлорофилла и продуктивности океана» . Биогеонауки . 7 (2): 621–40. Бибкод : 2010BGeo....7..621H . дои : 10.5194/bg-7-621-2010 . hdl : 1912/3208 .
- ^ Штайнахер, М.; Йоос, Ф.; Фрёлихер, ТЛ; Бопп, Л.; Кадуле, П.; Кокко, В.; Дони, Южная Каролина; Гелен, М.; Линдси, К.; Мур, Дж. К.; Шнайдер, Б.; Сегшнайдер, Дж. (2010). «Прогнозируемое снижение продуктивности морской среды в 21 веке: многомодельный анализ» . Биогеонауки . 7 (3): 979–1005. Бибкод : 2010BGeo....7..979S . дои : 10.5194/bg-7-979-2010 . hdl : 11858/00-001M-0000-0011-F69E-5 .
- ^ Коллинз, Шинейд; Рост, Бьёрн; Райнерсон, Татьяна А. (25 ноября 2013 г.). «Эволюционный потенциал морского фитопланктона в условиях закисления океана» . Эволюционные приложения . 7 (1): 140–155. дои : 10.1111/eva.12120 . ISSN 1752-4571 . ПМЦ 3894903 . ПМИД 24454553 .
- ^ Лобек, Кай Т.; Рибезель, Ульф; Ройш, Торстен Б.Х. (8 апреля 2012 г.). «Адаптивная эволюция ключевых видов фитопланктона к закислению океана». Природа Геонауки . 5 (5): 346–351. Бибкод : 2012NatGe...5..346L . дои : 10.1038/ngeo1441 . ISSN 1752-0894 .
- ^ Jump up to: а б с Кавиччиоли, Рикардо; Риппл, Уильям Дж.; Тиммис, Кеннет Н.; Азам, Фарук; Баккен, Ларс Р.; Бэйлис, Мэтью; Беренфельд, Майкл Дж.; Боэций, Антье; Бойд, Филип В.; Классен, Эме Т.; Кроутер, Томас В.; Дановаро, Роберто; Форман Кристин М.; Хьюсман, Джефф; Хатчинс, Дэвид А.; Янссон, Джанет К.; Карл, Дэвид М.; Коскелла, Бритт; Марк Уэлч, Дэвид Б.; Мартини, Дженнифер Б.Х.; Моран, Мэри Энн; Сирота, Виктория Дж.; Рей, Дэвид С.; Ремайс, Джастин В.; Рич, Вирджиния И.; Сингх, Браджеш К.; Штейн, Лиза Ю.; Стюарт, Фрэнк Дж.; Салливан, Мэтью Б.; и др. (2019). «Предупреждение ученых человечеству: микроорганизмы и изменение климата» . Обзоры природы Микробиология . 17 (9): 569–586. дои : 10.1038/s41579-019-0222-5 . ПМЦ 7136171 . ПМИД 31213707 . Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Jump up to: а б с Масотти, И.; Мулен, К.; Альвен, С.; Бопп, Л.; Тальябуэ, А.; Антуан, Д. (4 марта 2011 г.). «Крупномасштабные сдвиги в группах фитопланктона в экваториальной части Тихого океана во время циклов ЭНСО» . Биогеонауки . 8 (3): 539–550. Бибкод : 2011BGeo....8..539M . дои : 10.5194/bg-8-539-2011 . hdl : 20.500.11937/40912 .
- ^ Сатиккаб, Мария Белен; Бауэрак, Делия Елена; Гомес, Нора (15 сентября 2015 г.). «Влияние явления Южного колебания Эль-Ниньо на прибрежный фитопланктон в миксогалинной экосистеме на юго-востоке Южной Америки: устье Рио-де-ла-Плата» . Бюллетень о загрязнении морской среды . 98 (1–2): 26–33. Бибкод : 2015МарПБ..98...26С . дои : 10.1016/j.marpolbul.2015.07.017 . hdl : 11336/112961 . ПМИД 26183307 .
- ^ Сатик, Мария Белен; Бауэр, Делия Елена; Гомес, Нора (15 сентября 2015 г.). «Влияние явления Южного колебания Эль-Ниньо на прибрежный фитопланктон в миксогалинной экосистеме на юго-востоке Южной Америки: устье Рио-де-ла-Плата» . Бюллетень о загрязнении морской среды . 98 (1–2): 26–33. Бибкод : 2015МарПБ..98...26С . дои : 10.1016/j.marpolbul.2015.07.017 . hdl : 11336/112961 . ПМИД 26183307 .
- ^ Халлеграефф, GM (2003). «Вредное цветение водорослей: глобальный обзор» (PDF) . В Халлеграеффе, Густаве М.; Андерсон, Дональд Марк; Чембелла, Аллан Д.; Эневолдсен, Хенрик О. (ред.). Руководство по вредным морским микроводорослям . ЮНЕСКО. стр. 25–49. ISBN 978-92-3-103871-6 .
- ^ Хатчинсон, GE (1961). «Парадокс планктона». Американский натуралист . 95 (882): 137–45. дои : 10.1086/282171 . S2CID 86353285 .
- ^ Чарльсон, Роберт Дж.; Лавлок, Джеймс Э.; Андреэ, Мейнрат О.; Уоррен, Стивен Г. (1987). «Океанический фитопланктон, атмосферная сера, альбедо облаков и климат». Природа . 326 (6114): 655–61. Бибкод : 1987Natur.326..655C . дои : 10.1038/326655a0 . S2CID 4321239 .
- ^ Куинн, ПК ; Бейтс, Т.С. (2011). «Дело против регулирования климата посредством выбросов серы океаническим фитопланктоном» . Природа . 480 (7375): 51–6. Бибкод : 2011Natur.480...51Q . дои : 10.1038/nature10580 . ПМИД 22129724 . S2CID 4417436 .
- ^ Калбет, А. (2008). «Трофическая роль микрозоопланктона в морских системах» . Журнал морских наук ICES . 65 (3): 325–31. doi : 10.1093/icesjms/fsn013 .
- ^ Редфилд, Альфред К. (1934). «О пропорциях органических производных в морской воде и их связи с составом планктона». В Джонстоне, Джеймс; Дэниел, Ричард Джеллико (ред.). Мемориальный том Джеймса Джонстона . Ливерпуль: Университетское издательство Ливерпуля. стр. 176–92. OCLC 13993674 .
- ^ Jump up to: а б Арриго, Кевин Р. (2005). «Морские микроорганизмы и глобальные круговороты питательных веществ». Природа . 437 (7057): 349–55. Бибкод : 2005Natur.437..349A . дои : 10.1038/nature04159 . ПМИД 16163345 . S2CID 62781480 .
- ^ Фаннинг, Кент А. (1989). «Влияние загрязнения атмосферы на ограничение питательных веществ в океане». Природа . 339 (6224): 460–63. Бибкод : 1989Natur.339..460F . дои : 10.1038/339460a0 . S2CID 4247689 .
- ^ Стернер, Роберт Уорнер; Элзер, Джеймс Дж. (2002). Экологическая стехиометрия: биология элементов от молекул до биосферы . Издательство Принстонского университета. ISBN 978-0-691-07491-7 . [ нужна страница ]
- ^ Клаусмайер, Кристофер А.; Личман, Елена ; Левин, Саймон А. (2004). «Рост фитопланктона и стехиометрия в условиях многократного ограничения питательных веществ». Лимнология и океанография . 49 (4 Часть 2): 1463–70. Бибкод : 2004LimOc..49.1463K . дои : 10.4319/lo.2004.49.4_part_2.1463 . S2CID 16438669 .
- ^ Клаусмайер, Кристофер А.; Личман, Елена ; Дофрен, Танги; Левин, Саймон А. (2004). «Оптимальная стехиометрия азота и фосфора фитопланктона». Природа . 429 (6988): 171–4. Бибкод : 2004Natur.429..171K . дои : 10.1038/nature02454 . ПМИД 15141209 . S2CID 4308845 .
- ^ Jump up to: а б Беренфельд М.Дж. и Босс Э.С. (2018) «Учебное пособие для студентов по гипотезам цветения в контексте годовых циклов фитопланктона». Биология глобальных изменений , 24 (1): 55–77. дои : 10.1111/gcb.13858 .
- ^ Jump up to: а б Беренфельд, Майкл Дж.; Мур, Ричард Х.; Хостетлер, Крис А.; Графф, Джейсон; Гаубе, Питер; Рассел, Линн М.; Чен, Гао; Дони, Скотт С.; Джованнони, Стивен; Лю, Хунъюй; Проктор, Кристофер (22 марта 2019 г.). «Исследование аэрозолей и морской экосистемы Северной Атлантики (NAAMES): научный мотив и обзор миссии» . Границы морской науки . 6 : 122. дои : 10.3389/fmars.2019.00122 . ISSN 2296-7745 .
- ^ Энгель, Аня; Банге, Герман В.; Канлифф, Майкл; Берроуз, Сюзанна М.; Фридрихс, Гернот; Гальгани, Луиза; Херрманн, Хартмут; Херткорн, Норберт; Джонсон, Мартин; Лисс, Питер С.; Куинн, Патрисия К. (30 мая 2017 г.). «Жизненная кожа океана: к комплексному пониманию микрослоя морской поверхности» . Границы морской науки . 4 . дои : 10.3389/fmars.2017.00165 . hdl : 10026.1/16046 . ISSN 2296-7745 .
- ^ Беренфельд, Майкл Дж.; Босс, Эммануэль С. (3 января 2014 г.). «Возрождение экологических основ цветения океанского планктона» . Ежегодный обзор морской науки . 6 (1): 167–194. Бибкод : 2014ARMS....6..167B . doi : 10.1146/annurev-marine-052913-021325 . ISSN 1941-1405 . ПМИД 24079309 . S2CID 12903662 .
- ^ NAAMES: Наука - Цели Исследовательский центр Лэнгли, НАСА, Обновлено: 6 июня 2020 г. Дата обращения: 15 июня 2020 г.
- ^ Ригетти Д., Фогт М., Грубер Н., Псомас А. и Циммерманн Н.Э. (2019) «Глобальная картина разнообразия фитопланктона, обусловленная температурой и изменчивостью окружающей среды». Наука продвигается , 5 (5): eaau6253. два : 10.1126/sciadv.aau6253 . Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Jump up to: а б с Бердалл, Джон; Стойкович, Слободанка; Ларсен, Стюарт (2009). «Жизнь в мире с высоким содержанием CO2: влияние глобального изменения климата на морской фитопланктон» . Экология и разнообразие растений . 2 (2): 191–205. Бибкод : 2009PlEcD...2..191B . дои : 10.1080/17550870903271363 . S2CID 83586220 .
- ^ Басу, Самарпита; Макки, Кэтрин (2018). «Фитопланктон как ключевые медиаторы биологического углеродного насоса: их реакция на изменение климата» . Устойчивость . 10 (3): 869. дои : 10.3390/su10030869 . Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Морено, Эллисон Р.; Хагстром, Джордж I .; Примо, Франсуа В.; Левин, Саймон А.; Мартини, Адам С. (2018). «Стехиометрия морского фитопланктона опосредует нелинейные взаимодействия между поступлением питательных веществ, температурой и атмосферным CO 2 » . Биогеонауки . 15 (9): 2761–2779. Бибкод : 2018BGeo...15.2761M . дои : 10.5194/bg-15-2761-2018 .
- ^ Ли, Вэй; Гао, Куньшань; Бердалл, Джон (2012). «Интерактивное воздействие подкисления океана и ограничения содержания азота на диатомовую водоросль Phaeodactylum tricornutum» . ПЛОС ОДИН . 7 (12): e51590. Бибкод : 2012PLoSO...751590L . дои : 10.1371/journal.pone.0051590 . ПМЦ 3517544 . ПМИД 23236517 .
- ^ Ирвин, Эндрю Дж.; Финкель, Зоя В.; Мюллер-Каргер, Франк Э.; Трокколи Гиналья, Луис (2015). «Фитопланктон адаптируется к изменяющейся среде океана» . Труды Национальной академии наук . 112 (18): 5762–5766. Бибкод : 2015PNAS..112.5762I . дои : 10.1073/pnas.1414752112 . ПМЦ 4426419 . ПМИД 25902497 .
- ^ Хадер, Донат-П.; Виллафанье, Вирджиния Э.; Хелблинг, Э. Уолтер (2014). «Продуктивность водных первичных продуцентов в условиях глобального изменения климата» . Фотохим. Фотобиол. Наука . 13 (10): 1370–1392. дои : 10.1039/C3PP50418B . hdl : 11336/24725 . ПМИД 25191675 .
- ^ Сармьенто, JL; Слейтер, Р.; Барбер, Р.; Бопп, Л.; Дони, Южная Каролина; Херст, AC; Клейпас, Дж.; Матеар, Р.; Миколайевич, У.; Монфрей, П.; Солдатов В.; Сполл, ЮАР; Стоуффер, Р. (2004). «Реакция экосистем океана на потепление климата» . Глобальные биогеохимические циклы . 18 (3): н/д. Бибкод : 2004GBioC..18.3003S . дои : 10.1029/2003GB002134 . hdl : 1912/3392 .
- ^ Jump up to: а б Генрихс, Мара Э.; Мори, Коринна; Длугощ, Леон (2020). «Сложные взаимодействия между водными организмами и их химической средой, выясненные с разных точек зрения». YOUMARES 9 – Океаны: наши исследования, наше будущее . стр. 279–297. дои : 10.1007/978-3-030-20389-4_15 . ISBN 978-3-030-20388-7 . S2CID 210308256 . Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Лалли, Кэрол М. (16 мая 1997 г.). Биологическая океанография. Введение . Эльзевир Наука. ISBN 978-0-7506-3384-0 . OCLC 837077589 .
- ^ Jump up to: а б с д Маквей, Джеймс П., Най-Сянь Чао и Ченг-Шэн Ли. Справочник CRC по марикультуре Том. 1: Аквакультура ракообразных. Нью-Йорк: CRC Press LLC, 1993. [ нужна страница ]
- ^ Jump up to: а б Рой, Шовонлал (12 февраля 2018 г.). «Распределение углеводов, белков и липидов фитопланктона в мировом океане по цвету спутникового океана» . Журнал ISME . 12 (6): 1457–1472. Бибкод : 2018ISMEJ..12.1457R . дои : 10.1038/s41396-018-0054-8 . ISSN 1751-7370 . ПМЦ 5955997 . ПМИД 29434313 .
- ^ «Исследование питания выявило нестабильность в наиболее важных рыболовных регионах мира» .
- ^ Jump up to: а б с д Беренфельд, Майкл Дж. (2014). «Климатический танец планктона». Природа Изменение климата . 4 (10): 880–887. Бибкод : 2014NatCC...4..880B . дои : 10.1038/nclimate2349 .
- ^ Хатчинс, Д.А.; Бойд, PW (2016). «Морской фитопланктон и меняющийся цикл железа в океане». Природа Изменение климата . 6 (12): 1072–1079. Бибкод : 2016NatCC...6.1072H . дои : 10.1038/nclimate3147 .
- ^ Де Баар, Хейн Дж.В.; Де Йонг, Джероен ТМ; Баккер, Дороти CE; Лёшер, Беттина М.; Вет, Корнелис; Батманн, Ули; Сметачек, Виктор (1995). «Важность железа для цветения планктона и сокращения выбросов углекислого газа в Южном океане». Природа . 373 (6513): 412–415. Бибкод : 1995Natur.373..412D . дои : 10.1038/373412a0 . S2CID 4257465 .
- ^ Бойд, П.В.; Джикеллс, Т.; Право, КС; Блейн, С.; Бойл, Э.А.; Бюсселер, КО; Коул, КХ; Каллен, Джей-Джей; Де Баар, HJW; Следует, М.; Харви, М.; Ланселот, К.; Левассер, М.; Оуэнс, NPJ; Поллард, Р.; Ривкин, РБ; Сармьенто, Дж.; Шеманн, В.; Сметачек, В.; Такеда, С.; Цуда, А.; Тернер, С.; Уотсон, Эй Джей (2007). «Мезомасштабные эксперименты по обогащению железа 1993–2005 гг.: Синтез и будущие направления» (PDF) . Наука . 315 (5812): 612–617. Бибкод : 2007Sci...315..612B . дои : 10.1126/science.1131669 . ПМИД 17272712 . S2CID 2476669 . Архивировано из оригинала (PDF) 2 ноября 2020 года . Проверено 29 октября 2020 г.
- ^ Беренфельд, Майкл Дж.; о'Мэлли, Роберт Т.; Босс, Эммануэль С.; Вестберри, Тоби К.; Графф, Джейсон Р.; Хэлси, Кимберли Х.; Миллиган, Аллен Дж.; Сигел, Дэвид А.; Браун, Мэтью Б. (2016). «Переоценка воздействия потепления океана на глобальный фитопланктон». Природа Изменение климата . 6 (3): 323–330. Бибкод : 2016NatCC...6..323B . дои : 10.1038/nclimate2838 .
- ^ Jump up to: а б с Беренфельд, Майкл Дж.; Ху, Юнсян; о'Мэлли, Роберт Т.; Босс, Эммануэль С.; Хостетлер, Крис А.; Сигел, Дэвид А.; Сармьенто, Хорхе Л.; Шулиен, Дженнифер; Волосы, Джонатан В.; Лу, Сяомэй; Родье, Шэрон; Скарино, Эми Джо (2017). «Годовые циклы бума и спада биомассы полярного фитопланктона, выявленные с помощью космического лидара». Природа Геонауки . 10 (2): 118–122. Бибкод : 2017NatGe..10..118B . дои : 10.1038/ngeo2861 .
- ^ Беренфельд, Майкл Дж.; о'Мэлли, Роберт Т.; Сигел, Дэвид А.; Макклейн, Чарльз Р.; Сармьенто, Хорхе Л.; Фельдман, Джин С.; Миллиган, Аллен Дж.; Фальковски, Пол Г.; Летелье, Рикардо М.; Босс, Эммануэль С. (2006). «Климатические тенденции в современной продуктивности океана». Природа . 444 (7120): 752–755. Бибкод : 2006Natur.444..752B . дои : 10.1038/nature05317 . ПМИД 17151666 . S2CID 4414391 .
- ^ Левитан О.; Розенберг, Г.; Сетлик И.; Сетликова Е.; Григель, Дж.; Клепетар, Дж.; Прасил, О.; Берман-Франк, И. (2007). «Повышенный уровень CO2 усиливает фиксацию азота и рост морской цианобактерии Trichodesmium». Биология глобальных изменений . 13 (2): 531–538. Бибкод : 2007GCBio..13..531L . дои : 10.1111/j.1365-2486.2006.01314.x . S2CID 86121269 .
- ^ Верспаген, Иоланда М.Х.; Ван Де Ваал, Дедмер Б.; Финке, Ян Ф.; Виссер, Петра М.; Хейсман, Джефф (2014). «Контрастное влияние роста CO2 на первичное производство и экологическую стехиометрию при разных уровнях питательных веществ» (PDF) . Экологические письма . 17 (8): 951–960. Бибкод : 2014EcolL..17..951В . дои : 10.1111/ele.12298 . hdl : 20.500.11755/ecac2c45-7efa-4c90-9e29-f2bafcee1c95 . ПМИД 24813339 .
- ^ Холдинг, Дж. М.; Дуарте, CM; Санс-Мартин, М.; Меса, Э.; Арриета, Дж. М.; Кьеричи, М.; Хендрикс, ИЕ; Гарсиа-Коррал, Лос-Анджелес; Регоди-Де-Жу, А.; Дельгадо, А.; Рейгстад, М.; Вассманн, П.; Агусти, С. (2015). «Температурная зависимость первичной продукции с повышенным содержанием CO 2 в европейском Северном Ледовитом океане». Природа Изменение климата . 5 (12): 1079–1082. Бибкод : 2015NatCC...5.1079H . дои : 10.1038/nclimate2768 . hdl : 10754/596052 .
- ^ Бойс, Дэниел Г.; Льюис, Марлон Р.; Червь, Борис (2010). «Глобальное сокращение фитопланктона за последнее столетие». Природа . 466 (7306): 591–596. Бибкод : 2010Natur.466..591B . дои : 10.1038/nature09268 . ПМИД 20671703 . S2CID 2413382 .
- ^ Маккас, Дэвид Л. (2011). «Влияет ли смешивание данных о хлорофилле на временную тенденцию?». Природа . 472 (7342): Е4–Е5. Бибкод : 2011Natur.472E...4M . дои : 10.1038/nature09951 . ПМИД 21490623 . S2CID 4308744 .
- ^ Рыкачевски, Райан Р.; Данн, Джон П. (2011). «Размеренный взгляд на тенденции содержания хлорофилла в океане» . Природа . 472 (7342): Е5–Е6. Бибкод : 2011Natur.472E...5R . дои : 10.1038/nature09952 . ПМИД 21490624 . S2CID 205224535 .
- ^ Маккуаттерс-Голлоп, Эбигейл; Рид, Филип К.; Эдвардс, Мартин; Беркилл, Питер Х.; Кастеллани, Клаудия; Баттен, Соня; Гискес, Винфрид; Медведь, Дуг; Бидигар, Роберт Р.; Руководитель Эрика; Джонсон, Род; Кару, Мати; Козлоу, Дж. Энтони; Пенья, Анжелика (2011). «Есть ли сокращение морского фитопланктона?». Природа . 472 (7342): Е6–Е7. Бибкод : 2011Natur.472E...6M . дои : 10.1038/nature09950 . ПМИД 21490625 . S2CID 205224519 .
- ^ Бойс, Дэниел Г.; Льюис, Марлон Р.; Червь, Борис (2011). «Бойс и др. Ответ» . Природа . 472 (7342): Е8–Е9. Бибкод : 2011Natur.472E...8B . дои : 10.1038/nature09953 . S2CID 4317554 .
- ^ Антуан, Дэвид (2005). «Объединение наблюдений за цветом океана 1980-х и 2000-х годов в поисках долгосрочных тенденций» . Журнал геофизических исследований . 110 (С6): C06009. Бибкод : 2005JGRC..110.6009A . дои : 10.1029/2004JC002620 .
- ^ Вернан, Марсель Р.; Ван дер Вёрд, Хендрик Дж.; Гискес, Винфрид WC (2013). «Тенденции в цвете океана и концентрации хлорофилла с 1889 по 2000 год во всем мире» . ПЛОС ОДИН . 8 (6): е63766. Бибкод : 2013PLoSO...863766W . дои : 10.1371/journal.pone.0063766 . ПМК 3680421 . ПМИД 23776435 .
- ^ Руссо, Сесиль С.; Грегг, Уотсон В. (2015). «Последние десятилетние тенденции в глобальном составе фитопланктона» . Глобальные биогеохимические циклы . 29 (10): 1674–1688. Бибкод : 2015GBioC..29.1674R . дои : 10.1002/2015GB005139 .
- ^ по индексу морского льда Национальный центр данных по снегу и льду . По состоянию на 30 октября 2020 г.
- ^ Кирчман, Дэвид Л.; Моран, Хосе Анкселу Г.; Даклоу, Хью (2009). «Рост микробов в полярных океанах – роль температуры и потенциальное влияние изменения климата». Обзоры природы Микробиология . 7 (6): 451–459. дои : 10.1038/nrmicro2115 . ПМИД 19421189 . S2CID 31230080 .
- ^ Бенедетти, Фабио; Фогт, Майке; Элизондо, Урс Хофманн; Ригетти, Дамиано; Циммерманн, Никлаус Э.; Грубер, Николас (1 сентября 2021 г.). «Крупная реструктуризация сообществ морского планктона в условиях глобального потепления» . Природные коммуникации . 12 (1): 5226. Бибкод : 2021NatCo..12.5226B . дои : 10.1038/s41467-021-25385-x . ISSN 2041-1723 . ПМЦ 8410869 . ПМИД 34471105 .
Дальнейшее чтение
[ редактировать ]- Грисон, Филип Э. (1982). Аннотированный ключ к идентификации часто встречающихся и доминирующих родов водорослей, наблюдаемых в фитопланктоне США . Вашингтон, округ Колумбия: Типография правительства США. ISBN 978-0-607-68844-3 .
- Кирби, Ричард Р. (2010). Океанские дрифтеры: Тайный мир под волнами . Студия Кактус. ISBN 978-1-904239-10-9 .
- Мартин, Рональд; Куигг, Антониетта (2013). «Крошечные растения, которые когда-то правили морями». Научный американец . 308 (6): 40–5. Бибкод : 2013SciAm.308f..40M . doi : 10.1038/scientificamerican0613-40 . ПМИД 23729069 .
Внешние ссылки
[ редактировать ]- Secchi Disk и приложение Secchi , гражданский научный проект по изучению фитопланктона.
- «Океанские дрифтеры» — короткометражный фильм, рассказанный Дэвидом Аттенборо, о разнообразной роли планктона.
- Планктонные хроники , короткометражный документальный фильм и фотографии.
- DMS и климат , НОАА
- Plankton*Net , изображения планктонных видов


