Экология устья Сан-Франциско
Устье Сан-Франциско вместе с дельтой рек Сакраменто-Сан-Хоакин представляют собой сильно измененную экосистему . Регион был серьезно перестроен для удовлетворения потребностей в доставке воды, судоходстве, сельском хозяйстве, а в последнее время и в развитии пригородов. Эти потребности вызвали прямые изменения в движении воды и характере ландшафта, а также косвенные изменения в результате внедрения чужеродных видов . Новые виды изменили архитектуру пищевой сети так же, как дамбы изменили ландшафт островов и каналов, образующих сложную систему, известную как Дельта. [1]
В данной статье конкретно рассматривается экология зоны пониженной солености (ЗСЗ) лимана. Реконструировать историческую пищевую сеть LSZ сложно по ряду причин. Во-первых, нет четких данных о видах, исторически населявших устье реки. Во-вторых, устье и дельта Сан-Франциско на протяжении большей части своей 10-тысячелетней истории находились в геологическом и гидрологическом переходе, и поэтому описание «естественного» состояния устья во многом похоже на «попадание в движущуюся цель». [1] Изменение климата , гидрологическая инженерия, изменение потребностей в воде и появление новых видов будут продолжать изменять конфигурацию пищевой сети устья. Эта модель дает представление о текущем состоянии с примечаниями о недавних изменениях или появлении видов, которые изменили конфигурацию пищевой сети. Понимание динамики нынешней пищевой сети может оказаться полезным для усилий по восстановлению, направленных на улучшение функционирования и видового разнообразия устья.
Физическая география
[ редактировать ]Карта: острова в районе залива Сан-Франциско. |
|---|

Карта: острова залива Суисун и дельта рек Сакраменто – Сан-Хоакин. |
|---|

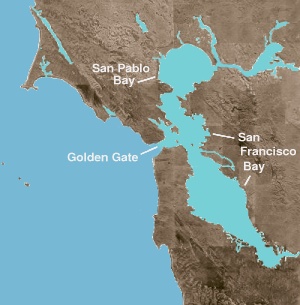
Залив Сан-Франциско является одновременно заливом и устьем . Первый термин относится к любому заливу или бухте, обеспечивающей физическое убежище от открытого океана. Устье — это любой физико-географический объект, где пресная вода встречается с океаном или морем. Северная часть залива представляет собой солоноватое устье, состоящее из ряда заливов, в которых преобладают потоки как морской, так и пресной воды. Этими географическими объектами, перемещающимися от соленого к пресному (или с запада на восток): залив Сан-Пабло , расположенный непосредственно к северу от Центрального залива; пролив Каркинес — узкий глубокий канал, ведущий в залив Суисун ; и дельта рек Сакраменто и Сан-Хоакин .
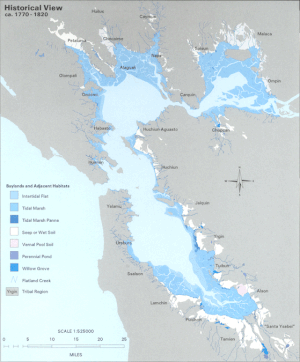
До 20-го века LSZ устья окаймлялся пресноводными с преобладанием туле водно-болотными угодьями . От 80 до 95% этих исторических водно-болотных угодий были засыпаны для облегчения землепользования и развития вокруг района залива. [2] утрата среды обитания на краях пелагической зоны Считается, что приводит к потере местных видов пелагических рыб из-за повышения уязвимости к хищникам.
В приливно - бентическом устье в настоящее время преобладают илистые отмели , которые в основном образовались в результате седиментации, образовавшейся в результате добычи золота в Сьерра-Неваде в конце 19 века. Тенденция к высоким нагрузкам наносов изменилась в 1950-х годах с появлением проекта «Центральная долина» , в результате которого большая часть наносов была заперта за плотинами, что привело к ежегодной чистой потере наносов из устья. [3] Таким образом, илистые отмели, похоже, медленно отступают, хотя мутность остается чрезвычайно высокой. Высокая мутность воды является причиной уникальных условий, существующих в устье Сан-Франциско, где высокая доступность питательных веществ не приводит к высокому производству фитопланктона. Вместо этого большинство фотосинтезирующих организмов из водорослей ограничены светом. [4]
Дельта также претерпела серьезные изменения. Начиная с 19 века, естественные дамбы были укреплены для обеспечения долговечности, чтобы защитить сельскохозяйственные угодья от регулярных наводнений. Многие из этих ферм были созданы на торфяных островах, расположенных посреди водных путей Дельты. Интенсивное земледелие привело к окислению высокого содержания углерода в почве, что привело к значительной потере почвенной массы. В результате эти острова опустились или погрузились почти на 6 метров ниже уровня моря. [5] Сегодня Дельта состоит из сильно изрезанных водными путями, перемежающихся островами, которые выглядят как «плавающие чаши» с бассейнами, расположенными далеко под поверхностью воды. [6] Эти острова подвергаются высокому риску затопления из-за обрушения дамбы. Ожидается, что последующий сдвиг солености на восток резко изменит экологию всей LSZ устья Сан-Франциско. [7]
Гидродинамика
[ редактировать ]
LSZ сосредоточен вокруг 2 psu ( практических единиц солености , измерения солености) и колеблется от примерно 6 psu до 0,5 psu. Основные поступления пресной воды в устье происходят за счет стока региональных осадков, рек Сакраменто и Сан-Хоакин. [8]
Приток реки в значительной степени контролируется попусками из водохранилищ выше по течению. Значительная часть этого притока экспортируется из Дельты в рамках федерального проекта Центральной долины и Государственного водного проекта в южную Калифорнию для сельскохозяйственного и городского использования. Эти изменения устранили большую часть изменений в стоке через устье (т. е. пресной воды, которая выходит из Золотых Ворот ), создавая более низкий отток зимой и более высокий отток летом, чем исторически наблюдалось в устье. В среднем приток пресной воды в устье составляет 50% исторического стока. [9] Фитопланктон , зоопланктон , личинки и взрослая рыба могут попасть в экспортные насосы, оказывая потенциально значительное, но неизвестное влияние на численность этих организмов. Это может быть особенно справедливо в отношении находящейся под угрозой исчезновения дельтовой корюшки , небольшой эндемичной рыбы; не является чем-то исключительным, за исключением того, что в исторических отчетах он описан как чрезвычайно богатый. [10] Считается, что дельтовая корюшка мигрирует и нерестится вверх по течению в дельте в начале лета, подвергая ее икру и личинки высокому риску уноса. [11] Управление корюшкой в настоящее время является источником разногласий, поскольку ее экология приводит к коллизии несопоставимых потребностей в воде для сохранения, развития и сельского хозяйства в Калифорнии.
Движение воды из устья сложное и зависит от ряда факторов. Приливные циклы заставляют воду двигаться к Золотым Воротам и от них четыре раза в течение 24 часов.
Используя 2 psu в качестве маркера зоны низкой солености, направление и величину колебаний можно отслеживать по расстоянию в километрах от Золотых Ворот или X2. Поскольку положение X2 зависит от ряда физических параметров, включая приток, вынос и приливы, его положение смещается на многие километры в суточном и сезонном цикле; в течение года он может варьироваться от залива Сан-Пабло в периоды сильного стока до дельты во время летней засухи. Положение X2 тщательно контролируется и поддерживается путем выпуска воды из резервуаров, расположенных выше по течению, в ожидании экспортного спроса. Это предусмотрено Решением 1641 Совета по водным ресурсам штата и требует, чтобы подача насосов на уровне штата и на федеральном уровне была сокращена, если X2 будет перенесен к востоку от острова Чиппс (75 километров реки вверх по течению от моста Золотые Ворота) в период с февраля по май или к востоку от Коллинсвилля ( 81 километр реки вверх по течению от моста Золотые Ворота) в январе, июне, июле и августе. (Д-1641, стр. 150)
Гравитационная циркуляция заставляет стратифицированную воду с высокой соленостью на глубине течь к суше, в то время как вода с низкой соленостью наверху течет в сторону моря. [12] Эффект гравитационной циркуляции может быть наиболее выражен в периоды большого расхода пресной воды, обеспечивая отрицательную обратную связь для поддержания соляного поля и распределения пелагических организмов в устье.
Перемешивание важно на границе гравитационной циркуляции, обращенной к суше, часто в районе X2, где толща воды становится менее стратифицированной. [13] Фиксированная зона смешивания возникает в районе «выступа Бенисия» на восточной оконечности пролива Каркинес, где глубокий канал резко мелеет по мере входа в залив Суисун. [14] Смешивание имеет решающее значение для поддержания солености, поэтому для перемещения X2 на небольшое расстояние на запад требуются чрезвычайно большие поступления пресной воды. Смешивание также помогает пелагическим организмам сохранять положение в устье. [1] замедление вывода первичного и вторичного производства из системы.
Пелагическая зона
[ редактировать ]Пелагические организмы всю или часть своей жизни проводят в открытой воде, где среда обитания определяется не границами, а физиологической толерантностью к солености и температуре. Зона низкой солености (LSZ) устья Сан-Франциско представляет собой среду обитания для ряда организмов, которые специализируются на выживании в этом уникальном слиянии наземных , пресноводных и морских влияний. Хотя существует множество сред обитания с различной экологией , которые являются частью устья (включая морские, пресноводные, приливные болотные и донные илистые системы), каждая из них связана с LSZ путем экспорта и импорта пресной воды, питательных веществ, углерода и организмов. [1]
Распределение и численность организмов в LSZ зависят как от абиотических, так и от биотических факторов. Абиотические факторы включают физическую географию и гидрологию устья, включая поступление питательных веществ , наносов нагрузку , мутность , стохастичность окружающей среды, климат и антропогенные воздействия. [8] Абиотические факторы, как правило, стимулируют производство в устьевой среде и опосредованы биотическими факторами.
Биотические факторы включают поглощение питательных веществ и первичное производство , вторичное производство зоопланктона , перенос энергии, адвекцию , пищевую сеть и трофическую динамику и распространение внутри и вне системы, выживание и смертность , хищничество и конкуренцию со стороны интродуцированных видов .
Пищевая сеть
[ редактировать ]
Трудно охарактеризовать историческую пищевую сеть устья Сан-Франциско из-за драматических изменений в географии, гидрологии и видовом составе, произошедших в прошлом столетии. Однако мониторинг, начатый в 1970-х годах, дает некоторую информацию об исторической динамике пищевой сети. До 1980-х годов в LSZ доминировала пищевая сеть, управляемая фитопланктоном, стабильная популяция мезопланктона, в которой доминировали копеподы Eurytemora affinis , и крупный макрозоопланктон, типичными представителями которого являются заливные креветки Сан-Франциско и креветки -мизиды . [15] Они обеспечивали питанием и энергией местных фильтраторов, таких как северный анчоус ( Engraulis mordax ), а также планктоноядных, таких как дельта-корюшка и молодь лосося .
Изменение пищевой сети исторически было вызвано увеличением мутности, а в последнее время - интродуцированными видами, как описано в разделах, посвященных первичному и вторичному производству.
Примечательно, что высокая скорость выведения интродуцированной популяции амурского моллюска Potamocorbula amurensis привела к десятикратному снижению плотности планктона, что привело к захвату углерода в бентосе и предполагаемому увеличению производства детрита в отходах. [16] Предполагается, что эти отходы подпитывают микробную петлю, что приводит к увеличению микрозоопланктона, такого как копепод Limnoithona Tetraspina , который использует коловраток и инфузорий .
Эти изменения являются одной из причин сокращения рыбных запасов. Например, северный анчоус Engraulis mordax до 1980-х годов был довольно многочисленным в зоне низкой солености, пока его ареал в устье не стал ограничиваться Центральным и Южным заливами. [17] Вероятно, это связано с поведенческой реакцией на интродукцию моллюска из реки Амур ( Potamocorbula amurensis ) и последующим снижением доступности планктона.
Совсем недавно было описано общее снижение пелагических организмов (POD), и это стало источником большой обеспокоенности в научном, управленческом и политическом сообществах. [18]
Несколько ключевых видов, в том числе дельта-корюшка , длинноперая корюшка , полосатый окунь и нитеплавник , были объявлены «видами, представляющими интерес» из-за постепенного снижения численности, начиная с 2001 года. [19] Это сопровождалось аналогичным снижением вторичной продуктивности и в настоящее время является источником большого количества исследований. Для объяснения POD был предложен ряд гипотез, включая разрушение пищевой сети, экспорт воды из Дельты и токсичные вещества из городских, промышленных или сельскохозяйственных источников.
Продюсеры
[ редактировать ]Первичное производство и потребление питательных веществ
[ редактировать ]Первичное производство фитопланктоном фиксирует энергию и ключевые питательные вещества в биологически доступной форме (т. е. в пище) посредством фотосинтеза . Производство фитопланктона в значительной степени структурировано физическими параметрами: наличием питательных веществ, солнечным светом, мутностью и температурой.
В устье Сан-Франциско имеется множество источников питательных веществ, которые можно использовать для первичного производства, в основном получаемых из очистных сооружений, сельскохозяйственных и городских дренажей, а также океана. [20] [21] Несмотря на это, устье уникально тем, что здесь наблюдается относительно низкий уровень первичной продукции. [22] Вероятно, это связано с двумя факторами: большим поступлением азота в форме аммония , который подавляет поглощение нитратов фитопланктоном, предпочитающим метаболически более дешевый NH 4 +, и высокой мутностью, которая ограничивает свет, необходимый для фотосинтеза, верхними несколькими сантиметрами слоя. столб воды. [21] Эта мутность — наследие гидравлической добычи золота в Сьерра-Неваде в 1850-х годах. [23]
Высокое время пребывания воды в устье способствует накоплению биомассы фитопланктона, увеличивая плотность, тогда как низкое время пребывания удаляет фитопланктон из устья. [1] Последнее типично для основных русел эстуария в периоды сильного стока, когда поверхностные воды имеют тенденцию смывать частицы и планктон вниз по течению.
Травоядные также удаляют фитопланктон из толщи воды. Хотя пелагическая пищевая сеть основана на производстве фитопланктона, большая часть этой продукции перенаправляется в бентос в результате хищничества завезённого амурского моллюска ( Corbula amurensis ). Уровни биомассы фитопланктона снизились на порядок после повсеместного внедрения C. amurensis в середине 1980-х годов и не восстановились. [24]
Фотосинтетическое производство
[ редактировать ]Основным источником энергии, получаемой путем фотосинтеза, является фитопланктон . Вообще говоря, диатомеи и микрофлагелляты производят большую часть биодоступного углерода в устье реки. [25] Другие виды, особенно динофлагелляты , могут вызывать вредное цветение водорослей или красные приливы , которые менее доступны для ассимиляции в пищевую сеть. [26]
Первичная продукция фитопланктона зависит от двух разных факторов: скорости роста и накопления (рис. 1). Хотя LSZ является поглотителем высоких концентраций питательных веществ из городских и сельскохозяйственных источников, темпы производства фитопланктона довольно низкие. [27] Нитрат оптимально используется фитопланктоном для роста, но аммоний (в основном получаемый из сточных вод) подавляет скорость роста. Таким образом, хотя фитопланктон и не ограничен в питательных веществах, он имеет тенденцию расти медленнее из-за присутствующих видов азота. [28] Еще одним фактором, подавляющим скорость роста, является высокая мутность устья, которая ограничивает способность фотосинтетически активной радиации (ФАР) проникать за пределы верхних нескольких сантиметров водного столба. Это ограничивает фотосинтез фитопланктона относительно мелкой фотозоной . Таким образом, при стратификации водной толщи , высокой мутности и присутствии аммония скорость роста фитопланктона обычно подавляется. [29] [30]
Накопление фитопланктона в первую очередь является результатом времени пребывания . [27] Северная дельта и залив Суисун имеют относительно короткое время пребывания из-за большого объема воды, движущейся через регион для нисходящего потока и для экспорта в южную Калифорнию. Поскольку вода движется через эту часть системы быстрее, скорость накопления снижается по мере того, как продуктивность выводится из системы. Напротив, в некоторых частях южной дельты время пребывания выше из-за небольшого объема воды, проходящей через систему; на самом деле вода иногда течет в обратном направлении из-за отсутствия притока из реки Сан-Хоакин и экспортной перекачки. Летом плотность фитопланктона здесь может быть на порядок выше, чем в других частях лимана. [31]
Вредное цветение водорослей (ВЦВ) динофлагеллят или цианобактерий производит токсичные побочные продукты метаболизма, которые делают их вредными для многих организмов. Поддерживаемые сочетанием высоких концентраций питательных веществ и температур, ВЦВ оказывают вдвойне негативное воздействие на пищевую сеть, конкурируя с диатомовыми водорослями и микрофлагеллятами, что еще больше снижает биодоступную первичную продукцию. Хотя некоторые беспозвоночные, такие как двустворчатые моллюски , не могут быть затронуты напрямую, они могут распространять токсины вверх по пищевой цепи, вызывая заболевание или убивая хищников . Не совсем понятно, как это веслоногих ракообразных влияет на . Инвазивные водоросли Microcystis aeruginosa в настоящее время распространены в Дельте в летние месяцы и могут снизить продуктивность копепод (помимо того, что они потенциально канцерогенны для человека). [32]
Обломочное производство
[ редактировать ]Огромное количество осадков и детрита проходит через LSZ. Большая часть из них представляет собой органические остатки в виде растворенных и взвешенных органических веществ (РОВ и ПОВ соответственно). Помимо источников выше по течению, органические вещества могут накапливаться в результате смертности местных организмов и образования отходов. [33] [34]
Детритофаги извлекают выгоду из этого источника энергии, создавая альтернативную и параллельную пищевую сеть потенциально большого значения. Это связано с тем, что фиксация углерода в детритной пищевой сети не ограничивается стратификацией, мутностью или длиной дня, которые ограничивают фотосинтез. Производство обломков происходит непрерывно, ограничиваясь только поступлением и выносом из системы Дельта. [25] [35]
Бактерии являются главными агентами трансформации РОВ и ПОМ в биодоступный углерод через микробную петлю . Этот механизм особенно важен в морских системах с ограниченным количеством питательных веществ, где бактерии выделяют питательные вещества из тонущего детрита, позволяя им возвращаться обратно в фотозону. Функционированию микробной петли в устье Сан-Франциско было посвящено мало исследований, но вполне возможно, что роль бактерий не является решающей в переработке питательных веществ в эвтрофной системе. Скорее, они могут обеспечить альтернативную пищевую цепь за счет прямого выпаса жгутиковых , коловраток и инфузорий . [36]
Высокая численность циклопоидного копепода Limnoithona Tetraspina может быть связана с тем, что он использует инфузории, а не фитопланктон в качестве основного источника пищи. [37] Основные виды каланоидов могут также использовать инфузории в качестве дополнительного или даже основного источника пищи, но в какой степени неизвестно. [38] [39]
Вторичное производство
[ редактировать ]Вторичная продукция относится к организмам, которые питаются первичной продукцией и передают энергию на более высокие трофические уровни эстуарной пищевой сети. Исторически сложилось так, что во вторичном производстве в устье Сан-Франциско доминировало производство креветок-мизид . [40] [41] Однако местный мизид Neomysis mercedis был в значительной степени заменен интродуцированным Acanthomysis Bowmani , который сохраняется при более низких плотностях. Интродуцированный амфипод Gammurus daiberi, возможно, занял часть этой ниши, но его обитание в основном ограничено пресной водой.
Сегодня основным источником вторичной продукции являются копеподы . Считается, что натурализованный местный веслоногий каланоид Eurytemora affinis был завезен примерно в конце 19 века. [42] Он доминировал в зоопланктоне зоны низкой солености до 1980-х годов, когда его в значительной степени заменил другой интродуцированный каланоидный копепод, Pseudodiaptomus forbesi . [16] [43] P. forbesi сохраняется за счет поддержания исходной популяции в пресноводных, густонаселенных районах эстуария, особенно в дельте, за пределами диапазона солеустойчивости моллюска реки Амур. [44] Поскольку некогда доминировавший E. affinis не имеет ареала вверх по течению, он более уязвим для хищников со стороны моллюсков и страдает от явной конкуренции с P. forbesi.
Другими каланоидными копеподами, которые могут иметь значение, являются недавно интродуцированные Sinocalanus doerri и Acartiella sinensis . Мало что известно об истории жизни этих организмов, хотя, судя по их морфологии, они могут охотиться на других копепод. Они появляются в нерегулярных циклах численности, во время которых могут доминировать в зоопланктоне. [45]
Еще один инвазивный копепод, очень маленький циклопид Limnoithona Tetraspina , появился в зоне низкой солености в 1990-х годах. С тех пор L. Tetraspina стала численно доминирующим копеподом, достигнув плотности порядка 10 000/м3. В качестве источника питания он использует микробную петлю , питаясь бактериями, инфузориями и коловратками. [37] Кроме того, по неизвестным причинам он кажется неуязвимым для хищников Амурского моллюска. Из-за своего небольшого размера L. Tetraspina обычно недоступна для потребления более крупными хищниками, особенно рыбами, что делает ее энергетическим тупиком.
Потребители
[ редактировать ]Первичные потребители
[ редактировать ]Первичные потребители полагаются на первичное производство как на основной источник питания. Важнейшими потребителями пелагической сети LSZ являются копеподы, а также упомянутые выше коловратки, жгутиконосцы и инфузории. Все виды веслоногих ракообразных сократились под сильным давлением хищников со стороны недавно завезенного моллюска из реки Амур ( Corbula amurensis ). [16] Из-за этого, а также из-за того, что веслоногие раконогие полагаются как на фотосинтетические, так и на детритные источники пищи, копеподы в LSZ имеют ограниченную обратную связь по первичной продукции, в отличие от морских и лентических систем, где веслоногие раконогие могут поедать цветы за считанные дни.
Pseudodiaptomus forbesi - доминирующий каланоидный копепод LSZ по биомассе. Он обладает достаточно широкой солеустойчивостью, поэтому может сохраняться как при низкой солености, так и в пресной воде. Такое широкое распространение помогает популяции сохранять убежище от хищников вверх по течению, в отличие от других видов с более узкой устойчивостью к солености. [44]
Limnoithona Tetraspina стала численно доминирующим веслоногим циклопоидом с момента своего появления в 1993 году. Он питается в основном инфузориями и микрофлагеллятами, но в отличие от P. forbesi он относительно невосприимчив к хищничеству со стороны моллюсков или рыб, отсюда и его численность. С энергетической точки зрения L. Tetraspina может оказаться тупиком для пищевой сети; эти копеподы либо выносятся из системы приливами и течениями, либо умирают и падают в бентос, где они могут быть доступны микробной петле или детритофагам. [46]
 Щитовое блюдечко ( Lottia pelta ) встречается в центральной и южной части залива Сан-Франциско. [47]
Щитовое блюдечко ( Lottia pelta ) встречается в центральной и южной части залива Сан-Франциско. [47]![Черная улитка-тюрбан (Tegula funebralis), обнаруженная в приливных зонах[47]](//upload.wikimedia.org/wikipedia/commons/thumb/5/5b/Tegula_funebralis.jpg/120px-Tegula_funebralis.jpg) Черная улитка-тюрбан ( Tegula funebralis ) обитает в приливных зонах. [47]
Черная улитка-тюрбан ( Tegula funebralis ) обитает в приливных зонах. [47] Hermissenda crassicornis встречается на доках, камнях и сваях. [47]
Hermissenda crassicornis встречается на доках, камнях и сваях. [47] Кукула Наттолла ( Clinocardium nuttallii ) в приливных зонах
Кукула Наттолла ( Clinocardium nuttallii ) в приливных зонах

![Черная улитка-тюрбан (Tegula funebralis), обнаруженная в приливных зонах[47]](http://upload.wikimedia.org/wikipedia/commons/thumb/5/5b/Tegula_funebralis.jpg/120px-Tegula_funebralis.jpg)


Хищные копеподы
[ редактировать ]По всей дельте обитает ряд хищных копепод, о которых известно относительно мало. Sinocalanus doerri , Acartiella sinensis и Tortanus dextrilobatus, по-видимому, морфологически способны поедать других копепод. Каждый из них был завезен в устье, вероятно, в результате замены балластных вод с 1980-х годов. Как правило, их количество недостаточно, чтобы негативно повлиять на веслоногих потребителей; однако происходит периодическое цветение S. doerri и A. sinensis , которое еще недостаточно изучено. [48]
Макробеспозвоночные
[ редактировать ]
(крошечные существа, похожие на креветок), хотя и способны к фильтрованию , Мизиды в основном плотоядны и питаются взрослыми веслоногими ракообразными. Они обеспечивали энергетический канал между планктоном и планктоноядными рыбами, включая молодь рыб, осетровых , чавычу и американскую шад . Когда-то мизиды были в изобилии, пока в середине 1980-х годов аборигенный вид Neomysis mercedis не был заменен инвазивным видом Acanthomysis Bowmani , который меньше по размеру и менее распространен. Снижение численности мизид было связано с последующим сокращением численности ряда видов рыб в устье реки в 1980-х и 90-х годах. [49] [50]
Креветки — плотоядные животные широкого профиля, которые охотятся в основном на мизид и амфипод . Crangon franciscorum представляет собой один из двух оставшихся коммерческих промыслов в устье реки. Хотя их больше не используют для «коктейлей из креветок из залива Сан-Франциско», их собирают для наживки. Среди других хищников - полосатый окунь , взрослые особи чавычи и смолты . [51]
Рыба
[ редактировать ]
Поскольку рыбы представляют собой таксономически и морфологически разнообразную группу, виды различаются по своей трофической экологии. В целом рыб можно разделить на четыре широкие категории по питанию: фильтраторы , планктофаги, рыбоядные и донные питающиеся животные.
Фильтраторы без разбора фильтруют толщу воды в поисках мелкой добычи, обычно фито- и зоопланктона. К этой категории рыб относятся нитеперая шэд ( Dorosoma petenense ), американская шэд ( Alosa sapidissima ), внутриконтинентальная серебрянка ( Menidia beryllina ) и анчоусы ( Engraulis mordax ). Некоторые данные свидетельствуют о том, что некоторые из этих видов ограничены в еде из-за снижения уровня планктона после завоза моллюсков из реки Амур. Анчоусы покинули LSZ в пользу более продуктивных регионов устья Сан-Пабло и Центрального залива. [17]
![Джексмелт (Atherinopsis Californiensis)[47]](//upload.wikimedia.org/wikipedia/commons/thumb/d/dc/Jack_Silverside.jpg/120px-Jack_Silverside.jpg)
![Дельта-корюшка (Hypomesus transpacificus)[47]](//upload.wikimedia.org/wikipedia/commons/thumb/3/3a/Hypomesus_transpacificus.jpg/120px-Hypomesus_transpacificus.jpg) Дельта-корюшка ( Hypomesus transpacificus ) [47]
Дельта-корюшка ( Hypomesus transpacificus ) [47]![Тихоокеанская сельдь (Clupea pallasii), обычно обитающая у берега, но заходящая в залив на нерест с ноября по март.[47]](//upload.wikimedia.org/wikipedia/commons/thumb/b/b0/Clupea_pallasii_by_OpenCage.jpg/120px-Clupea_pallasii_by_OpenCage.jpg) Тихоокеанская сельдь ( Clupea pallasii ), обычно находится у берега, но заходит в залив на нерест с ноября по март. [47]
Тихоокеанская сельдь ( Clupea pallasii ), обычно находится у берега, но заходит в залив на нерест с ноября по март. [47] Северный анчоус ( Engraulis mordax ) встречается в Сан-Пабло и заливе Суисун. [47]
Северный анчоус ( Engraulis mordax ) встречается в Сан-Пабло и заливе Суисун. [47]![Рыба-игла (Syngnathus leptorhynchus), обитающая в приливных зонах, обычно среди угрей [47]](//upload.wikimedia.org/wikipedia/commons/thumb/c/c7/Bay_pipefish_%28Syngnathus_leptorhynchus%29_01.jpg/120px-Bay_pipefish_%28Syngnathus_leptorhynchus%29_01.jpg) Рыба-игла ( Syngnathus leptorhynchus ), обитающая в приливных зонах, обычно среди угрей. [47]
Рыба-игла ( Syngnathus leptorhynchus ), обитающая в приливных зонах, обычно среди угрей. [47]
![Джексмелт (Atherinopsis Californiensis)[47]](http://upload.wikimedia.org/wikipedia/commons/thumb/d/dc/Jack_Silverside.jpg/120px-Jack_Silverside.jpg)
![Дельта-корюшка (Hypomesus transpacificus)[47]](http://upload.wikimedia.org/wikipedia/commons/thumb/3/3a/Hypomesus_transpacificus.jpg/120px-Hypomesus_transpacificus.jpg)
![Тихоокеанская сельдь (Clupea pallasii), обычно обитающая у берега, но заходящая в залив на нерест с ноября по март.[47]](http://upload.wikimedia.org/wikipedia/commons/thumb/b/b0/Clupea_pallasii_by_OpenCage.jpg/120px-Clupea_pallasii_by_OpenCage.jpg)

![Рыба-игла (Syngnathus leptorhynchus), обитающая в приливных зонах, обычно среди угрей [47]](http://upload.wikimedia.org/wikipedia/commons/thumb/c/c7/Bay_pipefish_%28Syngnathus_leptorhynchus%29_01.jpg/120px-Bay_pipefish_%28Syngnathus_leptorhynchus%29_01.jpg)
Планктофаги выборочно охотятся на отдельные зоопланктоны, такие как копеподы, мизиды и гаммариды . К этой группе относятся большинство личинок рыб, дельта-корюшка ( Hypomesus transpacificus ) и длинноплавниковая корюшка ( Spiinchus thaleichthys ), окунь ( Hysterocarpus traski ) и лососевые смолты . Дельтовая корюшка представляет особый интерес из-за своего статуса под угрозой исчезновения . Это может быть ограничено в еде, но данные несколько противоречивы. Другие факторы, такие как унос яиц и личинок при экспортной перекачке пресной воды из Дельты, также могут объяснить спад. [11]
![Окунь-сёрфингист (Cymatogaster aggregata) встречается в сублиторальных зонах[47]](//upload.wikimedia.org/wikipedia/commons/thumb/8/88/Cymatogaster_aggregata_1zz.jpg/120px-Cymatogaster_aggregata_1zz.jpg) Шайнер-серфингист ( Cymatogaster aggregata ), обитающий в сублиторальных зонах. [47]
Шайнер-серфингист ( Cymatogaster aggregata ), обитающий в сублиторальных зонах. [47] окунь ( Hysterocarpus traskie )
окунь ( Hysterocarpus traskie )![Судак-серфингист (Hyperprosopon argenteum)[47]](//upload.wikimedia.org/wikipedia/commons/thumb/b/b8/Steinhart_Aquarium_13_2013-03-27.jpg/120px-Steinhart_Aquarium_13_2013-03-27.jpg) Судак-серфингист ( Hyperprosopon argenteum ) [47]
Судак-серфингист ( Hyperprosopon argenteum ) [47] коричневый морской окунь ( Sebastes auriculatus )
коричневый морской окунь ( Sebastes auriculatus )![Колибри-жаба (Porichthys notatus), обитающая в сублиторальных зонах[47]](//upload.wikimedia.org/wikipedia/commons/thumb/d/db/Plainfin_Midshipman_-_Porichthys_notatus_%2843465733001%29.jpg/120px-Plainfin_Midshipman_-_Porichthys_notatus_%2843465733001%29.jpg) Колибри-жаба ( Porichthys notatus ), обитающая в сублиторальных зонах. [47]
Колибри-жаба ( Porichthys notatus ), обитающая в сублиторальных зонах. [47]
![Окунь-сёрфингист (Cymatogaster aggregata) встречается в сублиторальных зонах[47]](http://upload.wikimedia.org/wikipedia/commons/thumb/8/88/Cymatogaster_aggregata_1zz.jpg/120px-Cymatogaster_aggregata_1zz.jpg)

![Судак-серфингист (Hyperprosopon argenteum)[47]](http://upload.wikimedia.org/wikipedia/commons/thumb/b/b8/Steinhart_Aquarium_13_2013-03-27.jpg/120px-Steinhart_Aquarium_13_2013-03-27.jpg)

![Колибри-жаба (Porichthys notatus), обитающая в сублиторальных зонах[47]](http://upload.wikimedia.org/wikipedia/commons/thumb/d/db/Plainfin_Midshipman_-_Porichthys_notatus_%2843465733001%29.jpg/120px-Plainfin_Midshipman_-_Porichthys_notatus_%2843465733001%29.jpg)
Основным рыбоядным животным LSZ является полосатый окунь ( Morone saxatilis ), который был завезен в 1890-е годы и активно охотится на местных рыб. Полосатый окунь является важным спортивным промыслом в устье Сан-Франциско и, как таковой, представляет собой незначительный забор биомассы из устья. [52]
К бентосным, или донным, рыбам относятся белый осетр ( Acipenser transmontanus ), белый сом ( Ameiurus catus ) и звездчатая камбала ( Platichthys stellatus ). Из-за своей ориентации на среду обитания они питаются в основном эпибентосными организмами, такими как амфиподы, заливные креветки и двустворчатые моллюски. Известно, что эти рыбы, по крайней мере время от времени, питаются моллюсками реки Амур, которые представляют собой один из немногих каналов потока энергии от этого вида, за исключением производства детрита. [53]
Единственный коммерческий промысел в LSZ – это креветки-наживки. Существует множество видов спортивного рыболовства , которые представляют собой незначительный приток углерода, но приносят значительный приток капитала в местную экономику вокруг устья. Большая часть любительского рыболовства связана с полосатым окунем, осетром и интродуцированным пресноводным окунем в пресноводной дельте. Недостаток рыболовства делает устье Сан-Франциско уникальным. Почти все эстуарии мира поддерживают, по крайней мере, остатки значительного рыболовства. [54] Устье Сан-Франциско одно время поддерживало крупный промысел лосося, анчоусов и крабов Дандженесс до 1950-х годов. Упадок этих промыслов, вероятно, был вызван скорее утратой среды обитания , чем чрезмерным выловом рыбы . [2]
![Леопардовая акула (Triakis semifasciata)[47]](//upload.wikimedia.org/wikipedia/commons/thumb/b/be/Triakis_semifasciata_SI3.jpg/120px-Triakis_semifasciata_SI3.jpg) Леопардовая акула ( Triakis semifasciata ) [47]
Леопардовая акула ( Triakis semifasciata ) [47]![широконосая семижаберная акула (Notorynchus cepedianus)[47]](//upload.wikimedia.org/wikipedia/commons/thumb/e/e9/Notorynchus_cepedianus_head3.jpg/120px-Notorynchus_cepedianus_head3.jpg) Широконосая семижаберная акула ( Notorynchus cepedianus ) [47]
Широконосая семижаберная акула ( Notorynchus cepedianus ) [47]![колючая акула (Squalus acanthias), обитающая в сублиторальной зоне[47]](//upload.wikimedia.org/wikipedia/commons/thumb/2/29/Spiny_dogfish_hokitika.jpg/120px-Spiny_dogfish_hokitika.jpg) колючая морская собака ( Squalus acanthias ), обитающая в сублиторальной зоне. [47]
колючая морская собака ( Squalus acanthias ), обитающая в сублиторальной зоне. [47] Североамериканский зеленый осетр ( Acipenser medirostris )
Североамериканский зеленый осетр ( Acipenser medirostris ) Белый осетр ( Acipenser transmontanus )
Белый осетр ( Acipenser transmontanus )
![Леопардовая акула (Triakis semifasciata)[47]](http://upload.wikimedia.org/wikipedia/commons/thumb/b/be/Triakis_semifasciata_SI3.jpg/120px-Triakis_semifasciata_SI3.jpg)
![широконосая семижаберная акула (Notorynchus cepedianus)[47]](http://upload.wikimedia.org/wikipedia/commons/thumb/e/e9/Notorynchus_cepedianus_head3.jpg/120px-Notorynchus_cepedianus_head3.jpg)
![колючая акула (Squalus acanthias), обитающая в сублиторальной зоне[47]](http://upload.wikimedia.org/wikipedia/commons/thumb/2/29/Spiny_dogfish_hokitika.jpg/120px-Spiny_dogfish_hokitika.jpg)


Птицы
[ редактировать ]Устье Сан-Франциско является основной остановкой на тихоокеанском пролетном пути для мигрирующих водоплавающих птиц. Однако мало что известно о поступлении углерода в устье реки или из него через птиц . Миллионы водоплавающих птиц ежегодно используют мелководье залива как убежище. Большинство птиц — это резвящиеся утки , питающиеся подводной водной растительностью. Ныряющие утки (например, черпаки ) питаются эпибентическими организмами, такими как C. amurensis, что представляет собой возможный поток углерода из этого тупика. [55] [56] два исчезающих вида Здесь водятся птиц: калифорнийская крачка и калифорнийская хлопушка . Обнаженные илы залива обеспечивают важные места кормления куликов , но нижележащие слои ила залива представляют геологическую опасность для сооружений вблизи многих частей периметра залива. В устье также обитают рыбоядные птицы, такие как двухохлатый баклан и бурые пеликаны , но их трофическое влияние остается малоизученным.
В январе 2015 года ученые работали над обнаружением серого, толстого, липкого вещества без запаха на птицах вдоль береговой линии залива Сан-Франциско. Сотни птиц погибли, и еще сотни были покрыты этим веществом. Ученые обеспокоены тем, что другие дикие животные могут подвергаться риску из-за этого вещества. [57]
 Коричневый пеликан ( Pelecanus occidentalis Californicus ) в заливе Сан-Франциско
Коричневый пеликан ( Pelecanus occidentalis Californicus ) в заливе Сан-Франциско Рельс Риджуэя ( Rallus obsoletus ) в Окленде
Рельс Риджуэя ( Rallus obsoletus ) в Окленде двухохлатый баклан
двухохлатый баклан Кольклювая чайка ( Larus delawarensis ) в Окленде
Кольклювая чайка ( Larus delawarensis ) в Окленде





Млекопитающие
[ редактировать ]До 1825 года испанцы, французы, англичане, русские и американцы были привлечены в район залива для добычи огромного количества бобра ( Castor canadensis ), речной выдры, куницы, рыбака, норки, лисицы, ласки, обыкновенного тюленя и морских львов ( Zalophus Californianus). ) и калан ( Enhydra lutris ). Эта ранняя торговля мехом, известная как Калифорнийская меховая лихорадка , была более чем каким-либо другим фактором, ответственным за открытие Запада и района залива Сан-Франциско , в частности, для мировой торговли. [58] К 1817 году каланы в этом районе были практически истреблены. [59] С 1812 по 1840 год русские содержали станцию тюленей на близлежащих Фараллонских островах , вылавливая от 1200 до 1500 морских котиков ежегодно, хотя американские корабли уже эксплуатировали острова. [60] К 1818 году численность тюленей быстро уменьшилась, и в год можно было ловить только около 500 особей, а в течение следующих нескольких лет морские котики были истреблены с островов, пока в 1996 году они не начали повторно заселять острова. [61]
Хотя натуралисты 20-го века скептически относились к тому, что бобр исторически существовал в прибрежных ручьях или в самом заливе, [62] более ранние записи показывают, что калифорнийский золотой бобр ( Castor canadensis ssp. subauratus ) был одним из наиболее ценных пойманных животных и, по-видимому, встречался в большом количестве. [58] Томас Маккей сообщил, что за год компания Гудзонова залива выловила 4000 бобровых шкурок на берегу залива Сан-Франциско. Недавно бобры вновь заселили солоноватое болото Напа-Сонома на севере залива Сан-Пабло и его притоков Сонома-Крик и реки Напа . Бобры также повторно заселяют Южный залив, будучи переселенными в верхнюю часть Лос-Гатос-Крик у Лексингтонского водохранилища в 1980-х годах. Впоследствии они мигрировали вниз по течению к реке Гуадалупе и, достигнув соленой воды, использовали ее для повторного заселения на восток, в Койот-Крик , и на северо-запад, в Матадеро-Крик в Пало-Альто . [63]
Североамериканская речная выдра ( Lontra canadensis ) также повторно колонизирует залив и его притоки. Речная выдра в заливе впервые была зарегистрирована в Редвуд-Крик на пляже Мьюир в 1996 году. [64] а недавно в Корте-Мадера-Крик и в южной бухте Койот-Крик , [65] а также в 2010 году в самом заливе Сан-Франциско у пристани Ричмонд .
Впервые за 65 лет тихоокеанская морская свинья ( Phocoena phocoena ) вернулась в залив в 2009 году. [66] Golden Gate Cetacean Research, некоммерческая организация, занимающаяся исследованиями китообразных , разработала базу данных фотоидентификации, позволяющую ученым идентифицировать конкретных особей морских свиней, и пытается выяснить, привел ли более здоровый залив к их возвращению. [67] Тихоокеанская морская свинья обитает от Пойнт-Консепшен , штат Калифорния, до Аляски, а также до полуострова Камчатка и Японии. Недавние генетические исследования показывают, что существует местная популяция от Сан-Франциско до Русской реки и что прибрежные популяции восточной части Тихого океана редко мигрируют далеко, в отличие от морской свиньи западной части Атлантической гавани. [68]
Обыкновенная афалина ( Tursiops truncatus ) расширяет свой нынешний ареал к северу от залива Южная Калифорния . Первый прибрежный дельфин-афалина в районе залива Сан-Франциско за последнее время был замечен в 1983 году у побережья округа Сан-Матео в 1983 году. В 2001 году афалина была впервые замечена к востоку от моста Золотые Ворота и подтверждена фотографическими данными в 2007 году. Зооархеологические останки Количество афалин указывает на то, что афалина населяла залив Сан-Франциско в доисторические времена, по крайней мере, за 700 лет до настоящего времени, а черепа дельфинов, извлеченные из залива, предполагают случайных посетителей в исторические времена. [69]

 Морские львы ( Zalophus Californianus ) на пирсе 39
Морские львы ( Zalophus Californianus ) на пирсе 39 Тихоокеанский тюлень ( Phoca vitulina richardii )
Тихоокеанский тюлень ( Phoca vitulina richardii ) Речная выдра загорает на камнях в гавани Ричмонда
Речная выдра загорает на камнях в гавани Ричмонда Кит Хамфри — горбатый кит ( Megaptera novaeangliae ), вошедший в залив.
Кит Хамфри — горбатый кит ( Megaptera novaeangliae ), вошедший в залив. Морская свинья ( Phocoena phocoena )
Морская свинья ( Phocoena phocoena ) Мышь, добывающая солончаки ( Reithrodontomys raviventris ), представляет собой исчезающий вид, эндемичный для водно-болотных угодий залива Сан-Франциско и обладающий высокой солеустойчивостью.
Мышь, добывающая солончаки ( Reithrodontomys raviventris ), представляет собой исчезающий вид, эндемичный для водно-болотных угодий залива Сан-Франциско и обладающий высокой солеустойчивостью.







Медуза
[ редактировать ]До недавнего времени медузы не были распространены в устье реки. В эстуариях Восточного побережья, таких как Чесапикский залив , они часто являются хищниками высшего уровня , питающимися без разбора как рыбой, так и зоопланктоном. Несколько мелких инвазивных таксонов были идентифицированы в LSZ и пресноводных регионах. Эти виды летом стробилируют, но сохраняют полипы в бентосе круглый год.
Их влияние на планктон неизвестно, но проводятся исследования для его количественной оценки. При достаточной плотности желе могут играть дополнительную роль по отношению к C. amurensis в подавлении зоопланктона, населяя районы с низкой соленостью за пределами ареала моллюсков, где планктонные виды имели убежище, свободное от хищников.
 Corynactis Californica обнаружена на доках и сваях в приливной зоне.
Corynactis Californica обнаружена на доках и сваях в приливной зоне.

Бентические потребители
[ редактировать ]Бентосное сообщество взяло на себя непропорционально большую роль в экологии пищевой сети эстуария из-за ключевых инвазий двустворчатых моллюсков . [16] Использование этих моллюсков: моллюск реки Амур ( Corbula amurensis ) обладает широкой толерантностью к солености, которая распространяется в зону низкой солености, но не в пресную воду. Он фильтрует и питается фитопланктоном и мелким зоопланктоном, например, науплиями каланоидных копепод . В устье Сан-Франциско у моллюска мало хищников, и это позволяет ему достигать высокой плотности (порядка десятков тысяч/м2). Благодаря высокой скорости очистки он способен очистить всю толщу воды в некоторых частях устья за несколько дней, что приводит к резкому истощению популяций планктона. [70] Считается, что это основная причина снижения продуктивности экосистемы после вторжения моллюсков в середине 1980-х годов. [16] [71] [72]
 Звездчатая камбала ( Platichthys stellatus )
Звездчатая камбала ( Platichthys stellatus )![Крапчатый санддаб (Citharichthys stigmaeus) встречается в сублиторальных зонах с песчаным дном[47]](//upload.wikimedia.org/wikipedia/commons/thumb/4/49/Citharichthys_stigmaeus_%28Speckled_Sanddab%29.jpg/120px-Citharichthys_stigmaeus_%28Speckled_Sanddab%29.jpg) Крапчатый песчаник ( Citharichthys stigmaeus ) встречается в сублиторальных зонах с песчаным дном. [47]
Крапчатый песчаник ( Citharichthys stigmaeus ) встречается в сублиторальных зонах с песчаным дном. [47] Летучая мышь ( Myliobatis Californica ) и Большой скат ( Beringraja binoculata ) в Аквариуме залива обнаружены в сублиторальных районах залива. [47]
Летучая мышь ( Myliobatis Californica ) и Большой скат ( Beringraja binoculata ) в Аквариуме залива обнаружены в сублиторальных районах залива. [47]

![Крапчатый санддаб (Citharichthys stigmaeus) встречается в сублиторальных зонах с песчаным дном[47]](http://upload.wikimedia.org/wikipedia/commons/thumb/4/49/Citharichthys_stigmaeus_%28Speckled_Sanddab%29.jpg/120px-Citharichthys_stigmaeus_%28Speckled_Sanddab%29.jpg)

Это снижение продуктивности по существу связано с перенаправлением пелагической сети на бентосную цепь этим видом. [16] Поскольку моллюск реки Амур питается первичными продуцентами, потребителями и хищниками, он влияет на несколько трофических уровней . Следовательно, почти весь планктон демонстрирует признаки явной конкуренции , поскольку производство на одном трофическом уровне влияет на все остальные за счет увеличения численности моллюсков. отрицательной Это приводит к возникновению петли обратной связи : C. amurensis ограничивает биомассу планктона, что, в свою очередь, ограничивает C. amurensis . Однако поступления из-за пределов системы из-за приливной адвекции или источников, расположенных выше по течению, могут увеличить биомассу C. amurensis , что еще больше приведет к ограничению планктона. Эта петля обратной связи еще больше усиливается, поскольку моллюск может существовать более одного или двух лет, что оказывает дополнительное давление на популяции планктона во время циклов низкой продуктивности.
Перенаправление углерода C. amurensis в бентос привело к образованию ограниченной цепи, в результате чего пелагическая сеть обеднела. Производство детрита в результате выделений и гибели моллюсков может стимулировать производство бактерий, которые могут циркулировать в детритной пищевой сети или микробной петле.
Хотя переработанные питательные вещества могут поддерживать некоторый рост фитопланктона, в конечном итоге это приводит к увеличению популяции C. amurensis. Недавний успех вторжения Limnoithona Tetraspina можно объяснить с точки зрения этого явления. Он питается инфузориями и микрофлагеллятами, которые слишком малы, чтобы их мог съесть моллюск, тем самым избегая конкуренции. Кроме того, L. Tetraspina , по-видимому, неуязвима для хищников со стороны моллюсков или (почти) чего-либо еще. Возникновение микробной пищевой сети и вторжение L. Tetraspina Capitalize являются результатом неиспользованного альтернативного пути потока энергии в пищевой сети, которому способствует C. amurensis. Последующие модели вторжения могут отражать аналогичную картину.
![Краб Дандженесс (Metacarcinus magister)[47]](//upload.wikimedia.org/wikipedia/commons/thumb/d/da/2011-08-14_San_Francisco_043_Pier_39%2C_Aquarium_of_the_Bay_%286092202672%29.jpg/120px-2011-08-14_San_Francisco_043_Pier_39%2C_Aquarium_of_the_Bay_%286092202672%29.jpg) Краб Дандженесс ( Metacarcinus magister ) [47]
Краб Дандженесс ( Metacarcinus magister ) [47] Красный каменный краб ( Cancer Productus )
Красный каменный краб ( Cancer Productus ) Изящный краб ( Metacarcinus gracilis )
Изящный краб ( Metacarcinus gracilis ) Калифорнийский каменный краб ( антеннариум Ромалеона )
Калифорнийский каменный краб ( антеннариум Ромалеона ) Желтый береговой краб ( Hemigrapsus oregonensis ) встречается в мелководных приливных зонах. [47]
Желтый береговой краб ( Hemigrapsus oregonensis ) встречается в мелководных приливных зонах. [47] Американский краб-паук ( Pyromaia tuberculata ), найденный под камнями и возле свай доков. [47]
Американский краб-паук ( Pyromaia tuberculata ), найденный под камнями и возле свай доков. [47] Волосатый рак-отшельник ( Pagurus hirsutiusculus )
Волосатый рак-отшельник ( Pagurus hirsutiusculus )![Охристая морская звезда (Pisaster ochraceus) обитает в приливной зоне[47]](//upload.wikimedia.org/wikipedia/commons/thumb/d/dd/Pisaster_ochraceus_%2823729798342%29.jpg/120px-Pisaster_ochraceus_%2823729798342%29.jpg) Охристая морская звезда ( Pisaster ochraceus ) найдена в приливной зоне. [47]
Охристая морская звезда ( Pisaster ochraceus ) найдена в приливной зоне. [47]
![Краб Дандженесс (Metacarcinus magister)[47]](http://upload.wikimedia.org/wikipedia/commons/thumb/d/da/2011-08-14_San_Francisco_043_Pier_39%2C_Aquarium_of_the_Bay_%286092202672%29.jpg/120px-2011-08-14_San_Francisco_043_Pier_39%2C_Aquarium_of_the_Bay_%286092202672%29.jpg)






![Охристая морская звезда (Pisaster ochraceus) обитает в приливной зоне[47]](http://upload.wikimedia.org/wikipedia/commons/thumb/d/dd/Pisaster_ochraceus_%2823729798342%29.jpg/120px-Pisaster_ochraceus_%2823729798342%29.jpg)
Интродуцированные виды
[ редактировать ]
Интродукция видов увеличивалась, по крайней мере, с 19 века в результате увеличения торговли и перевозок. Интродукции включают многочисленные таксоны, в том числе копеподы, креветки , амфиподы , двустворчатые моллюски , рыбы, а также укорененные и плавающие растения. Многие пелагические виды были завезены совсем недавно в результате сброса балластной воды с крупных судов непосредственно в устье реки. [73] В результате многие из этих интродуцированных видов происходят из эстуариев Тихоокеанского региона, особенно веслоногие раки, такие как P. forbesi и L. Tetraspina. Моллюск реки Амур происходит из Азии и внес значительные и радикальные изменения в экологию LSZ, в первую очередь за счет направления пелагической пищи в бентос и в ускоренный микробный цикл. [74]
Виды также были завезены через прицепы к спортивным лодкам, которые перевозятся между регионами. [73] Это вероятный источник ряда растений с низкой соленостью, таких как Egeria densa и водяной гиацинт ( Eichhornia crassipes ). Эти растения вызвали глубокие изменения в дельте, нарушив поток воды, затеняя фитопланктон и обеспечив среду обитания для рыбоядных рыб, таких как полосатый окунь Morone saxatilis , который сам был намеренно завезен в конце 1800-х годов из Чесапикского залива . [52] [73] Ожидается, что пресноводная мидия квагга , родом из Европы, будет завезена лодочниками в течение следующих двух-десяти лет, несмотря на меры предосторожности. [75] Кроме того, молодой возраст и изолированная среда устья Сан-Франциско привели к успеху инвазивных беспозвоночных из-за отсутствия биоразнообразия. [54]
Будущие вторжения
[ редактировать ]Современная пищевая сеть возникла в результате серии инвазий и трофических замен.
Ожидается, что этот процесс будет продолжаться, поскольку новые организмы появляются в результате случайных или преднамеренных интродукций. Менее ясно, в какой степени предыдущие внедрения прокладывают путь для будущих вторжений. Это может произойти одним из трех способов.
- Ранний захватчик может предоставить ресурс, который не будет использоваться в новой системе до тех пор, пока не будет введен новый хищник ( L. Tetraspina и микробная петля, как описано выше).
- Ранние вселенцы могут способствовать появлению новых, изменяя среду обитания и делая ее пригодной для последующих инвазий (желейные полипы, использующие в качестве субстрата раковины моллюсков реки Амур).
- Очевидная конкуренция между старыми и новыми обитателями может увеличить возможности для вторжения и расселения новых организмов, которые могут извлечь выгоду из неразработанных ресурсов (субсидирование амурских моллюсков верхними по течению популяциями интродуцированных копепод P. forbesi, что создает давление на местных копепод).
Краткое содержание
[ редактировать ]Пищевая сеть LSZ в устье Сан-Франциско действует в двух параллельных и асимметричных направлениях. Основная часть углерода ассимилируется в бентосных и микробных петлях, которые представляют собой энергетические тупики. Меньшая фракция доставляется на более высокие пелагические трофические уровни, которые могут поддерживать веслоногих ракообразных, рыбу, птиц и рыболовство. Такое перенаправление пищевой сети на эти две узкие петли может быть причиной сокращения численности макробеспозвоночных и рыб в устье реки, которые действуют вне этих цепочек. Восстановление эстуария до более высокой степени функциональности зависит от вероятности принести больше пользы пелагической сети без субсидирования бентоса.
Экология будущего
[ редактировать ]Экологию зоны низкой солености устья Сан-Франциско сложно охарактеризовать, поскольку она является результатом сложной синергии как абиотических, так и биотических факторов. Кроме того, он продолжает подвергаться быстрым изменениям в результате появления новых видов, прямого антропогенного воздействия и изменения климата . Будущие экологические изменения будут происходить в масштабах всей экосистемы, особенно по мере того, как повышение уровня моря , тектоническая нестабильность и упадок инфраструктуры приведут к разрушению дамбы в дельте. [76] Ожидается, что возникший в результате обратный поток воды вынудит X2 попасть в дельту, поставив под угрозу пространственно ориентированную среду обитания (например, пресноводные болота), перераспределив зону с низкой соленостью и поставив под угрозу водоснабжение южной Калифорнии с неизвестными и непредвиденными последствиями для природы и человека. экология крупнейшего устья западного побережья.
См. также
[ редактировать ]- Список водотоков в районе залива Сан-Франциско
- Гидрография района залива Сан-Франциско
- Проект «Водораздел»
- Устье Сан-Франциско и наука о водоразделах
Примечания
[ редактировать ]- ^ Jump up to: а б с д и Киммерер 2004 г.
- ^ Jump up to: а б Мы знаем 1979 год.
- ^ Вайсьер 2006
- ^ Джассби 2000
- ^ Деверел 1998 г.
- ^ Филип 2007 г.
- ^ Лунд 2007
- ^ Jump up to: а б Киммерер 2002 г.
- ^ Гюнтер, Эндрю (2011). «Состояние залива Сан-Франциско, 2011 г., Партнерство устья Сан-Франциско» (PDF) . Состояние залива Сан-Франциско, 2011 г. Проверено 6 октября 2017 г.
- ^ Мойл 1992
- ^ Jump up to: а б Беннетт 2006 г.
- ^ Монисмит 1996
- ^ Бурау 1998 г.
- ^ Шеллхамер 2001
- ^ Медведи 1986
- ^ Jump up to: а б с д и ж Киммерер 1996 г.
- ^ Jump up to: а б Киммерер 2006 г.
- ^ Дэвис, Джей (сентябрь 2006 г.). «Пульс устья 2006» (PDF) . sfei.org . Устьевой институт Сан-Франциско . Проверено 2 ноября 2016 г. .
- ^ Тагер 2005 г.
- ^ Смит 2002
- ^ Jump up to: а б Дагдейл 2003 г.
- ^ Джассби 2002
- ^ Николс 1986
- ^ Киммерер 1994 г.
- ^ Jump up to: а б Джассби и Клерн 2000 г.
- ^ Андерсон и др. 2002 г.
- ^ Jump up to: а б Джассби и др. 2002 г.
- ^ Дагдейл и др. 2003 г.
- ^ Клерн 1987
- ^ Коул и Клерн, 1987 г.
- ^ Болл и Артур 1979
- ^ Леман и Уоллер, 2003 г.
- ^ Мюррелл и др. 1999 год
- ^ Мюррелл и Холлибо 2000
- ^ Джассби и др. 1993 год
- ^ Холлибо и Вонг, 1996 г.
- ^ Jump up to: а б Були 2006 г.
- ^ Роллваген-Болленс и Пенри, 2003 г.
- ^ Rollwagen-Bollens et al. 2006 г.
- ^ Модлин 1997 г.
- ^ Киммерер 1998 г.
- ^ Ли 1999
- ^ Медведи 1991
- ^ Jump up to: а б Дюран2006
- ^ Медведи 1999 г.
- ^ Були и Киммерер, 2006 г.
- ^ Jump up to: а б с д и ж г час я дж к л м н тот п д р с т Муи, Рич (2007). Животные залива Сан-Франциско: путеводитель по распространенным донным видам . Сан-Франциско, Калифорния: Калифорнийская академия наук. ISBN 978-0-940228-69-6 . OCLC 86174077 .
- ^ Орси и Оцука 1999 г.
- ^ Орси и Кнутсон, 1979 г.
- ^ Модлин и Орси 1997.
- ^ Ситтс и Найт 1979
- ^ Jump up to: а б Радович 1963 г.
- ^ Петерсон 1997
- ^ Jump up to: а б Николс и др. 1986 год
- ^ Поултон и др. 2002 г.
- ^ Ричман и Ловворн, 2004 г.
- ^ Ученые спешат идентифицировать птиц на берегах залива Сан-Франциско (январь 2015 г.), The Christian Science Monitor.
- ^ Jump up to: а б Скиннер, Джон Э. (1962). Исторический обзор ресурсов рыбы и дикой природы в районе залива Сан-Франциско (ресурсы млекопитающих) . Департамент рыболовства и охоты Калифорнии, Отчет отдела водных проектов №. 1. Сакраменто, Калифорния: Департамент рыболовства и охоты Калифорнии. Архивировано из оригинала 26 июля 2011 г.
- ^ Сюзанна Стюарт и Адриан Прецеллис (ноябрь 2003 г.). Проблемы археологических исследований национального побережья Пойнт-Рейес - национальной зоны отдыха Золотые Ворота (PDF) (Отчет). Центр антропологических исследований Государственного университета Сономы. п. 335 . Проверено 10 января 2010 г.
- ^ Томпсон, Р.А. (1896 г.). Русское поселение в Калифорнии, известное как Форт Росс, основанное в 1812 году... заброшенное в 1841 году: почему они пришли и почему ушли . Санта-Роза, Калифорния: Издательская компания Sonoma Democrat. п. 7. ISBN 0-559-89342-6 .
- ^ Уайт, Питер (1995). Фараллонские острова: стражи Золотых ворот . Сан-Франциско, Калифорния: Scottwall Associates. ISBN 0-942087-10-0 .
- ^ Таппе, Дональд Т. (1942). «Положение бобров в Калифорнии». Игровой бюллетень №3 . Калифорнийский департамент рыбы и дичи.
- ^ Сью Дреманн (4 ноября 2022 г.). «Бобёр вернулся: Пара околоводных грызунов замечена в Пало-Альто» . Пало-Альто Еженедельник . Проверено 18 января 2023 г.
- ^ «Выходи наружу!» . Хроники Сан-Франциско. Апрель 1966 года . Проверено 6 марта 2011 г.
- ^ «Ранчо Блю Оук Резерв» . Калифорнийский университет. Архивировано из оригинала 12 мая 2011 г. Проверено 6 марта 2011 г.
- ^ Дэвид Перлман (8 ноября 2010 г.). «Морские свиньи возвращаются в залив Сан-Франциско – ученые выясняют, почему» . Хроники Сан-Франциско . Проверено 25 июля 2011 г.
- ^ «Проект гавани морской свиньи» . Золотые Ворота Исследования китообразных . Проверено 25 июля 2011 г.
- ^ Морская свинья (Phocoena phocoena): Сан-Франциско-Русский речной запас (PDF) (Отчет). Национальная служба морского рыболовства . 15 октября 2009 года . Проверено 25 июля 2011 г.
- ^ Уильям Кинер; Марк А. Уэббер; Тим М. Марковиц; Марк П. Коттер; Даниэла Мальдини; Р. Х. Дефран; Меган Райс; Аманда Дж. Дебич; Эйми Р. Ланг; Деннис Л. Келли; Алекс Г. Кесарис; Маддалена Беарзи; Кайла Кози; Дэвид Андерсон; Лори Шустер; Дэвид В. Веллер (2023). «Расширение северного ареала калифорнийских прибрежных дельфинов-афалин (Tursiops truncatus)» . Водные млекопитающие . 49 (1): 29–43. дои : 10.1578/AM.49.1.2023.29 . S2CID 255918023 .
- ^ Вернер и Холлибо, 1993 г.
- ^ Карлтон и др. 1990 год
- ^ Николс и др. 1990 год
- ^ Jump up to: а б с Карлтон 1996 г.
- ^ Коул 1992
- ^ Дей 2007
- ^ Эпштейн 2006
Ссылки
[ редактировать ]- Андерсон, DM, премьер-министр Глиберт и Дж. М. Буркхолдер. 2002. Вредное цветение водорослей и эвтрофикация: источники питательных веществ, состав и последствия. Эстуарии 25:704-726.
- Болл, доктор медицинских наук, и Дж. Ф. Артур. 1979. Динамика планктонного хлорофилла в северной части залива и дельты Сан-Франциско. Страницы 265–286 в издательстве TJ Conomos. Залив Сан-Франциско: урбанизированное устье. Американская ассоциация содействия развитию науки, Тихоокеанский отдел, Сан-Франциско, Калифорния.
- Беннетт, Вашингтон (2006). Рост и выживание дельта-корюшки во время недавнего упадка пелагических организмов: что вызывает у них летнее уныние? Научная конференция CALFED. Сакраменто, Калифорния.
- Були П., WJK (2006). «Экология очень многочисленного, интродуцированного циклопоидного копепода в устье умеренного пояса». Серия Marine Ecology Progress Series 324: 219–228.
- Бурау, младший (1998). «Результаты гидродинамического элемента исследования зоны захвата 1994 года в заливе Суисун, Сакраменто, Калифорния». Межведомственная экологическая программа для технического отчета устья залива и дельты Сан-Франциско 56: 13–62.
- Карлтон, Дж.Т., Дж.К. Томпсон, Л.Е. Шемель и Ф.Х. Николс. 1990. Замечательное вторжение в залив Сан-Франциско (Калифорния, США) азиатского моллюска Potamocorbula amurensis. 1. Внедрение и распространение. Серия «Прогресс в области морской экологии». 66:81-94.
- Карлтон, Дж. (1996). «Биологические инвазии и криптогенные виды». Экология 77 (6).
- Коул, Б.Е. и Дж.Э. Клерн. 1987. Эмпирическая модель для оценки продуктивности фитопланктона в эстуариях. Серия «Прогресс в области морской экологии». 36:299-305.
- Коул, Б.Е., Дж.К. Томпсон и Дж.Э. Клерн (1992). «Измерение скорости фильтрации инфаунальными двустворчатыми моллюсками в рециркуляционном лотке». Морская биология 113: 219–225.
- Клоерн, Дж. Э. 1987. Мутность как контроль биомассы и продуктивности фитопланктона в эстуариях. Исследования континентального шельфа 7: 1367-1381.
- Кономос, Т.Дж., Эд. (1979). Залив Сан-Франциско: урбанизированное устье. Сан-Франциско, Калифорния, Тихоокеанское отделение Американской ассоциации содействия развитию науки.
- Деверел, С. (1998). Смягчение последствий оседания в дельте Сакраменто-Сан-Хоакин. Программа CALFED Bay-Delta.
- Дей, Р. (16 января 2007 г., 2007 г.). «Информация об инвазивных мидиях из национальной зоны отдыха на озере Мид». Инициатива «100-й меридиан», с сайта http://100thmeridian. [ постоянная мертвая ссылка ] . орг/мед. жерех.
- Дагдейл, Р.К., В. Хог, А. Марчи, А. Ласситер и Ф. Вилкерсон. 2003. Влияние антропогенного поступления и потока аммония на первичное производство в заливе Сан-Франциско. Научная программа CALFED, Сакраменто, Калифорния.
- Дюран, Дж. (2006). Факторы, определяющие неудачу набора веслоногих ракообразных каланоидов в научной программе по теляту в устье Сан-Франциско. Сакраменто, Калифорния.
- Эпштейн, Э. (2006). Скрипучие дамбы готовы к катастрофе. Хроники Сан-Франциско. Сан-Франциско.
- Холлибо, Дж. Т. и П. С. Вонг. 1996. Распространение и активность бактериопланктона в заливе Сан-Франциско. Страницы 263–288 в книге Дж. Т. Холлибо, редактор. Залив Сан-Франциско: экосистема. Американская ассоциация содействия развитию науки, Сан-Франциско, Калифорния.
- Джассби, А.Д., Дж.Э. Клоерн и Б.Е. Коул. 2002. Ежегодная первичная продукция: закономерности и механизмы изменений в богатой питательными веществами приливной экосистеме. Лимнология и океанография 47:698-712.
- Джассби, А.Д., Дж.Э. Клоерн и Т.М. Пауэлл. 1993. Источники и поглотители органического углерода в заливе Сан-Франциско – изменчивость, вызванная речным стоком. Серия «Прогресс в области морской экологии». 95.
- Джассби, А.Д. и Дж.Э. Клерн. 2000. Источники органических веществ и реабилитация дельты Сакраменто-Сан-Хоакин (Калифорния, США). Охрана водных ресурсов: морские и пресноводные экосистемы 10:323-352.
- Джассби, А.Д., Дж.Э. Клерн и Б.Е. Коул. (2002). «Ежегодная первичная продукция: закономерности и механизмы изменений в богатой питательными веществами приливной экосистеме». Лимнология и океанография 47 (3): 698–712.
- Киммерер, У. Дж., Э. Гартсайд и Дж. Дж. Орси. (1994). «Хищничество завезенного моллюска как вероятная причина существенного сокращения зоопланктона залива Сан-Франциско». Мар. Экол. Прог. Сер. 113: 81–93.
- Киммерер, В.Дж., Дж.Дж. Орси. 1996. Изменения в зоопланктоне устья залива Сан-Франциско с момента вселения моллюска Potamocorbula amurensis. Страницы 403–425 в книге Дж. Т. Холлибо, редактор. Залив Сан-Франциско: экосистема. Американская ассоциация содействия развитию науки, Сан-Франциско, Калифорния.
- Киммерер, В.Дж., Дж.Р. Бурау и В.А. Беннетт. (1998). «Приливно-ориентированная вертикальная миграция и поддержание положения зоопланктона в умеренном устье». Лимнол. Океаногр. 43 (7): 1697–1709.
- Киммерер, WJ (2002). «Физические, биологические и управленческие реакции на переменный приток пресной воды в устье Сан-Франциско». Эстуарии 25: 1275–1290.
- Киммерер, В. (2004). «Процесс открытой воды в устье Сан-Франциско: от физического воздействия к биологическим реакциям». Устье Сан-Франциско и наука о водоразделах 2 (1).
- Киммерер, В. (2006). «Реакция анчоусов ослабляет воздействие инвазивного двустворчатого моллюска Corbula amurensis на пищевую сеть устья Сан-Франциско». Серия Marine Ecology Progress Series 324: 207–218.
- Ли, CE (1999). «Быстрые и повторяющиеся вторжения копепод Eurytermora affinis в пресную воду». Эволюция 53 (5): 1423–1434.
- Леман, П.В. и С. Уоллер. 2003 г. В дельте цветет микроцистис. Информационный бюллетень IEP 16:18-19.
- Лунд Дж., Э. Ханак, В. Флинор, Р. Ховитт, Дж. Маунт, П. Мойл (2007). Представление будущего дельты Сакраменто-Сан-Хоакин. Сан-Франциско, Институт государственной политики.
- Модлин, РФ, и Дж. Дж. Орси (1997). «Acanthomysis Bowmani, новый вид, и A. asperali, Mysidacea, недавно обнаруженная в устье Сакраменто-Сан-Хоакин, Калифорния (Crustacea: Mysidae).» Proceedings of the Biological Society of Washington 110 (3): 439–446.
- Монизмит, С.Г., Дж. Р. Бурау и М. Стейси (1996). Динамика стратификации и гравитационная циркуляция в северной части залива Сан-Франциско. Залив Сан-Франциско: экосистема. Дж. Т. Холлибо. Сан-Франциско, Калифорния, Американская ассоциация содействия развитию науки: 123–153.
- Мойл, П.Б., Б. Гербольд, Д.Е. Стивенс и Л.В. Миллер. (1992). «История жизни и статус дельтовой корюшки в устье Сакраменто-Сан-Хоакин, Калифорния». Пер. Являюсь. Рыба. Соц. 121: 67–77.
- Мюррелл, MC, Дж. Т. Холлибо, М. В. Сильвер и П. С. Вонг. 1999. Динамика бактериопланктона в северной части залива Сан-Франциско: роль ассоциации частиц и сезонного потока пресной воды. Лимнология и океанография. 44:295-308.
- Мюррелл, MC, и Дж. Т. Холлибо. 2000. Распределение и состав растворенного и дисперсного органического углерода в северной части залива Сан-Франциско в условиях низкого расхода. Эстуарий, прибрежные и шельфовые науки 51:75-90.
- Николс, Ф. Х., Дж. Э. Клоерн, С. Луома и Д. Х. Петерсон. 1986. Модификация лимана. Наука 231:567-573.
- Николс, Ф.Х., Дж.К. Томпсон и Л.Е. Шемель. 1990. Замечательное вторжение в залив Сан-Франциско (Калифорния, США) азиатского моллюска Potamocorbula amurensis. 2. Перемещение прежнего сообщества. Серия «Прогресс в области морской экологии». 66:95-101.
- Орси, Джей-Джей и А.С. Натсон. 1979. Роль креветок-мизид в устье Сакраменто-Сан-Хоакин и факторы, влияющие на их численность и распространение. Страницы 401–408 в книге TJ Conomos, редактор. Залив Сан-Франциско: урбанизированный устье. Американская ассоциация содействия развитию науки, Тихоокеанский отдел, Сан-Франциско.
- Орси, Джей-Джей и У.Л. Мекум. (1986). «Распределение и численность зоопланктона в дельте Сакраменто-Сан-Хоакин в зависимости от определенных факторов окружающей среды». Эстуарии 9 (4 B): 326–339.
- Орси, Джей-Джей и Т.С. Уолтер. (1991). «Pseudodiaptomus forbesi и P. marinus (Copepoda: Calanoida), последние иммигранты веслоногих раков в калифорнийское устье Сакраменто-Сан-Хоакин». Бюлл. Планктон Соц. Япония Спец. Том. (Материалы Четвертой Международной конференции по Copepoda): 553–562.
- Орси, Джей-Джей и С. Оцука. (1999). «Интродукция азиатских копепод Acartiella sinensis, Tortanus dextrilobatus (Copepoda: Calanoida) и Limnoithona Tetraspina (Copepoda: Cyclopoida) в устье реки Сан-Франциско, Калифорния, США». Планк. Био. Экол. 46: 128–131.
- Петерсон, Х. 1997. Осетр, фаршированный моллюсками. Информационный бюллетень IEP 9:19-19.
- Филип, Т. (2007). Время доставки водных решений. Сакраменто Би. Сакраменто, Калифорния.
- Поултон, В.К., Дж.Р. Ловворн и Дж.Ю. Такекава. 2002. Плотность моллюсков и пищевое поведение червеца в заливе Сан-Пабло, Калифорния. Кондор. 104:518-527.
- Радович, Дж. (1963). «Влияние температуры океана на перемещение полосатого окуня Roccus saxatilis в сторону моря на побережье Тихого океана». Департамент рыболовства и охоты Калифорнии 49: 191–205.
- Ричман, С.Э. и Дж.Р. Ловворн. 2004. Относительная кормовая ценность местных и экзотических моллюсков из залива Сан-Франциско для черненых уток. Экологические приложения 14:1217-1231.
- Rollwagen-Bollens, GC и DL Penry. 2003. Динамика питания Acartia sp. в большом устье умеренного пояса (залив Сан-Франциско, Калифорния). Серия достижений морской экологии 257:139-158.
- Роллваген-Болленс, Г., С. Гиффорд, А. Слотер и С.М. Болленс. 2006. Протисты в устье реки с умеренным климатом: разнообразие, выпас и потребление многоклеточными животными. Эос, Труды Американского геофизического союза, Приложение к совещанию по наукам об океане. 87:36 ОС11Д-05.
- Шёллхамер, Д.Х. (2001). Влияние солености, топографии дна и приливов на места максимумов мутности эстуариев в северной части залива Сан-Франциско. Прибрежные и устьевые процессы мелкодисперсных отложений. WH McAnally и AJ Mehta. Амстердам, Нидерланды, Elsevier: 343–356.
- Ситтс Р.М., А.В. Найт. 1979. Хищничество эстуарных креветок Crangon franciscorum Stimpson и Palaemon macrodactylus Rathbun. Биологический бюллетень 156:356-368.
- Смит, С.В. и Дж. Т. Холлибо (2002). «Обмен водой, солью и питательными веществами в заливе Сан-Франциско». Межведомственная экологическая программа для технического отчета 66 залива Сан-Франциско и устья дельты Сан-Франциско.
- Тагер, Майк (2005). Экологические сирены в Дельте кричат. Контра Коста Таймс.
- Тротт, Дж.С.С., Уоррен Дж. Фергюсон и Эндрю Дж. Кляйнфельд (2006). Центральное агентство по водным ресурсам дельты против Бюро мелиорации. Апелляционный суд США девятого округа. Сан-Франциско, Калифорния.
- Вайсьер, член парламента (2006). Тенденции изменения бентоса верхнего устья Сан-Франциско. Научная конференция CALFED. Сакраменто, Калифорния.
- Вернер И. и Дж. Т. Холлибо. 1993. Potamocorbula amurensis (Mollusca, Pelecyopoda): Сравнение скорости выведения и эффективности ассимиляции фитопланктона и бактериопланктона. Лимнология и океанография 38:949-964.


