Морские протисты
Морские протисты определяются по их среде обитания как протисты , которые живут в морской среде , то есть в соленой воде морей или океанов или в солоноватой воде прибрежных эстуариев . Жизнь зародилась как морские одноклеточные прокариоты (бактерии и археи), а затем развилась в более сложные эукариоты . Эукариоты — это более развитые формы жизни, известные как растения, животные, грибы и протисты. Протисты — это эукариоты, которых нельзя отнести к растениям, грибам или животным. Они в основном одноклеточные и микроскопические. Термин «протист» исторически вошёл в употребление как удобный термин для обозначения эукариот, которых нельзя строго отнести к растениям, животным или грибам. Они не являются частью современной кладистики, поскольку являются парафилетическими (не имеют общего предка для всех потомков).
Большинство протистов слишком малы, чтобы их можно было увидеть невооруженным глазом. Это очень разнообразные организмы, которые в настоящее время разделены на 18 типов, но их нелегко классифицировать. [1] [2] Исследования показали, что большое разнообразие протистов существует в океанах, глубоких морских жерлах и речных отложениях, что позволяет предположить, что большое количество эукариотических микробных сообществ еще не обнаружено. [3] [4] протистов мало исследовали Миксотрофных , но недавние исследования в морской среде показали, что миксотрофные протисты составляют значительную часть биомассы протистов . [5] Поскольку протисты являются эукариотами (а не прокариотами), они обладают внутри своей клетки по крайней мере одним ядром , а также органеллами , такими как митохондрии и тельца Гольджи . Многие виды протистов могут переключаться между бесполым размножением и половым размножением, включающим мейоз и оплодотворение . [6]
В отличие от клеток прокариот клетки эукариот высокоорганизованы. Растения, животные и грибы обычно многоклеточные и обычно макроскопические . Большинство протистов одноклеточные и микроскопические. Но есть исключения. Некоторые одноклеточные морские простейшие макроскопичны. Некоторые морские слизевики имеют уникальный жизненный цикл, включающий переключение между одноклеточными, колониальными и многоклеточными формами. [7] Другие морские протисты не являются ни одноклеточными, ни микроскопическими, например морские водоросли .
Протистов описывают как таксономическую кучку неудачников, куда все, что не вписывается в одно из основных биологических царств . можно поместить [8] Некоторые современные авторы предпочитают исключать многоклеточные организмы из традиционного определения простейшего, ограничивая простейших одноклеточными организмами. [9] [10] Это более ограниченное определение исключает все бурые , многоклеточные красные и зеленые водоросли и, иногда, слизевики (слизевики исключаются, когда многоклеточность определяется как «сложная»). [11]
| Часть серии обзоров по |
| Морская жизнь |
|---|
 |
Фон
[ редактировать ]«Морские протисты — это полифилетическая группа организмов, играющих важную роль в экологии и биогеохимии океанов, в том числе выполняющих большую часть фотосинтеза Земли и управляющих циклами углерода, азота и кремния. Кроме того, морские протисты занимают ключевые позиции на дереве жизнь, в том числе как ближайшие родственники многоклеточных животных [животных]... Одноклеточных эукариот часто называют «протистами», термин, который полезен, несмотря на его таксономическую неуместность и происхождение как определение путем исключения: простейшим считается любой эукариот, не являющийся растение, животное или гриб». [12]
Океан представляет собой крупнейшую непрерывную планетарную экосистему, в которой обитает огромное разнообразие организмов, в том числе микроскопическая биота, такая как одноклеточные эукариоты (протисты). Несмотря на свой небольшой размер, протисты играют ключевую роль в морских биогеохимических циклах и обладают огромным эволюционным разнообразием. [13] [14] Несмотря на их значение для понимания эволюции жизни на Земле и их роли в морских пищевых сетях , а также управления биогеохимическими циклами для поддержания обитаемости, мало что известно об их клеточной биологии, включая размножение , метаболизм и передачу сигналов . [12] Большая часть имеющихся биологических знаний основана на сравнении белков с культивируемых видов гомологами генетически поддающихся модели таксонов. [15] [16] [17] [18] Основным препятствием для понимания клеточной биологии этих разнообразных эукариот является то, что протоколы генетической модификации доступны лишь для небольшого числа видов. [19] [20] которые не представляют ни наиболее экологически значимых протистов, ни широты эукариотического разнообразия. Несмотря на это, в десятилетие до 2020 г. геном [15] [16] [17] и транскриптома инициативы по секвенированию [18] привели к тому, что почти 120 миллионов унигенов , у протистов было идентифицировано [21] что способствует разработке генетических инструментов для модельных видов. [22]


Трофические режимы
[ редактировать ]Простейших можно разделить на четыре группы в зависимости от того, какое у них питание: растительное, животное, грибковое и [23] или их смесь. [24]
Протисты по способу добывания пищи | |||||||
|---|---|---|---|---|---|---|---|
| Тип простейшего | Описание | Пример | Некоторые другие примеры | ||||
| Растительный | Автотрофные простейшие, которые производят себе пищу без необходимости потреблять другие организмы, обычно путем фотосинтеза (иногда путем хемосинтеза). |  | Зеленые водоросли Пирамимоны. | Красные и бурые водоросли , диатомовые водоросли , кокколитофоры и некоторые динофлагелляты . Растительноподобные протисты являются важными компонентами фитопланктона, обсуждаемыми ниже . | |||
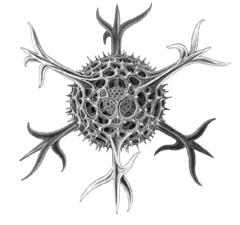
| Животный | Гетеротрофные протисты, добывающие пищу, потребляя другие организмы (бактерии, археи и мелкие водоросли). |  | Радиолярий- протист, рисунок Геккеля | Фораминиферы , а также некоторые морские амебы , инфузории и жгутиконосцы . | |||
| Грибковый | Сапротрофные протисты, получающие пищу из останков разложившихся и разложившихся организмов. |  | Сети морской слизи образуют лабиринтные сети трубок, по которым могут перемещаться амебы без ложноножек. | Морские лишайники | |||
| Миксотрофы | Различный ( см. ниже ) | Миксотрофные и осмотрофные протисты, которые получают пищу за счет комбинации вышеперечисленных веществ. |  | Euglena mutabilis — фотосинтезирующий жгутиконосец . | Многие морские миксотропы встречаются среди простейших, особенно среди инфузорий и динофлагеллят. [5] | ||


- Одноклеточные и микроскопические протисты
![Диатомовые водоросли являются основной группой водорослей, производящей около 20% мирового производства кислорода.[26]](//upload.wikimedia.org/wikipedia/commons/thumb/3/31/Diatoms_through_the_microscope.jpg/297px-Diatoms_through_the_microscope.jpg) Диатомовые водоросли — основная группа водорослей, производящая около 20% мирового производства кислорода. [26]
Диатомовые водоросли — основная группа водорослей, производящая около 20% мирового производства кислорода. [26] Ископаемые панцири диатомей от 32 до 40 млн лет назад
Ископаемые панцири диатомей от 32 до 40 млн лет назад
 Одноклеточная водоросль Gephyrocapsa Oceanica.
Одноклеточная водоросль Gephyrocapsa Oceanica. Две динофлагелляты
Две динофлагелляты

 Эта инфузория переваривает цианобактерии . Цитостом . или рот находится внизу справа
Эта инфузория переваривает цианобактерии . Цитостом . или рот находится внизу справа
![Диатомовые водоросли являются основной группой водорослей, производящей около 20% мирового производства кислорода.[26]](http://upload.wikimedia.org/wikipedia/commons/thumb/3/31/Diatoms_through_the_microscope.jpg/297px-Diatoms_through_the_microscope.jpg)







| Внешние видео | |
|---|---|
- Инфузория, поедающая диатомовую водоросль
- Амеба поглощает диатомовую водоросль
Грибоподобные сапробоны-протисты специализируются на поглощении питательных веществ из неживой органической материи, например, мертвых организмов или их отходов. Например, многие виды оомицетов растут на мертвых животных или водорослях. Морские сапробиотические протисты выполняют важную функцию по возврату неорганических питательных веществ в воду. Этот процесс обеспечивает рост новых водорослей, что, в свою очередь, обеспечивает пищу для других организмов в пищевой цепи. Действительно, без сапробных видов, таких как протисты, грибы и бактерии, жизнь прекратила бы свое существование, поскольку весь органический углерод оказался «связанным» в мертвых организмах. [27] [28]
Миксотрофы
[ редактировать ]

У миксотрофов нет единого трофического режима. Миксотроф — это организм, который может использовать смесь различных источников энергии и углерода вместо одного трофического режима в континууме от полной автотрофии на одном конце до гетеротрофии на другом. Подсчитано, что миксотрофы составляют более половины всего микроскопического планктона. [29] Существует два типа эукариотических миксотрофов: те, которые имеют собственные хлоропласты , и те, которые имеют эндосимбионты , а также другие, которые приобретают их посредством клептопластики или путем порабощения всей фототрофной клетки. [30]
Различие между растениями и животными часто нарушается у очень маленьких организмов. Возможные комбинации: фото- и хемотрофия , лито- и органотрофия , ауто- и гетеротрофия или другие их комбинации. Миксотрофы могут быть как эукариотами , так и прокариотами . [31] Они могут использовать различные условия окружающей среды. [32]
Недавние исследования морского микрозоопланктона показали, что 30–45% численности инфузорий являются миксотрофными, а до 65% биомассы амебоидов, форамов и радиолярий - миксотрофными. [5]
Phaeocystis — важный род водорослей, встречающийся в составе морского фитопланктона по всему миру. Он имеет полиморфный жизненный цикл: от свободноживущих клеток до крупных колоний. [33] Он обладает способностью образовывать плавающие колонии, где сотни клеток встроены в гелевую матрицу, которая может значительно увеличиваться в размерах во время цветения . [34] В результате Phaeocystis вносит важный вклад в содержание морского углерода. [35] и циклы серы . [36] Виды Phaeocystis являются эндосимбионтами акантарианских радиолярий. [37] [38]
Миксотрофный планктон, сочетающий фототрофию и гетеротрофию – таблица на основе Stoecker et al., 2017. [39] | |||||||
|---|---|---|---|---|---|---|---|
| Общие типы | Описание | Пример | Дальнейшие примеры | ||||
| Бактериопланктон | Фотогетеротрофный бактериопланктон |  | Холерный вибрион | Розеобактер виды. Эритробактерии виды. Гаммапротеобактериальная клада ОМ60 Распространен среди бактерий и архей. | |||
| Фитопланктон | назвал их конститутивными миксотрофами . Mitra et al., 2016 [40] Питающийся фитопланктон: фотосинтезирующие протисты с наследственными пластидами и способностью поедать добычу. |  | охромонас Виды | Ochromonas spp. Малый Примнезиум Примеры динофлагеллят: Fragilidium subglobosum , Heterocapsa triquetra , Karlodinium ядовитый , Neoceratium Furca , Prorocentrum minimus. | |||
| Зоопланктон | назвал их неконститутивными миксотрофами . Mitra et al., 2016 [40] Зоопланктон, обладающий фотосинтезом: микрозоопланктон или многоклеточный зоопланктон, приобретающий фототрофию за счет удержания хлоропластов. а или поддержание эндосимбионтов водорослей. | ||||||
| Генералисты | Протисты, сохраняющие хлоропласты и редко другие органеллы многих таксонов водорослей. |  | Большинство инфузорий - олиготрихов, сохраняющих пластиды. а | ||||
| Специалисты | 1. Протисты, сохраняющие хлоропласты, а иногда и другие органеллы одного вида водорослей или очень близких видов водорослей. |  | Динофиз остроконечный | Динофизис виды. Мезодиний красный | |||
| 2. Протисты или зоопланктон с водорослевыми эндосимбионтами только одного вида водорослей или очень близких видов водорослей. |  | Ноктилука сцинтилланс | Метазоопланктон с эндосимбионтами водорослей Наиболее миксотрофные ризарии ( Acantharea , Polycystinea и Foraminifera ). Зеленая Noctiluca scintillans | ||||
| а Удержание хлоропластов (или пластид) = секвестрация = порабощение. Некоторые виды, сохраняющие пластиды, также сохраняют другие органеллы и цитоплазму добычи. | |||||||
- Миксопланктон
 Тинтинниды инфузории Фавелла
Тинтинниды инфузории Фавелла Euglena mutabilis — фотосинтезирующий жгутиконосец .
Euglena mutabilis — фотосинтезирующий жгутиконосец . Зоохлореллы (зеленые), живущие внутри инфузории Stichotricha secunda .
Зоохлореллы (зеленые), живущие внутри инфузории Stichotricha secunda .



Протистское передвижение
[ редактировать ]Другой способ классификации протистов — по способу их передвижения. Многие одноклеточные протисты, особенно простейшие, подвижны и могут совершать движения с помощью жгутиков , ресничек или ложноножек . Клетки, использующие жгутики для движения, обычно называются жгутиками , клетки, использующие реснички, обычно называются инфузориями , а клетки, использующие ложноножки, обычно называются амебами или амебоидами . Другие протисты не подвижны и, следовательно, не имеют механизма движения.
Протисты в зависимости от того, как они двигаются | ||||||||
|---|---|---|---|---|---|---|---|---|
| Тип простейшего | Механизм движения | Описание | Пример | Другие примеры | ||||
| Для Моти | Жгутиконосцы |  | Жгутик хлыст (лат. « » ) — это похожий на плеть придаток, который выступает из тела клетки некоторых простейших (а также некоторых бактерий). Жгутиконосцы используют от одного до нескольких жгутиков для передвижения, а иногда и как органеллы питания и чувств . |  | Криптофиты | Все динофлагелляты и нанофлагелляты ( хоанофлагелляты , силикофлагелляты , большинство зеленых водорослей ) [41] [42] (Другие протисты проходят фазу гамет , когда у них появляется временный жгутик — некоторые радиолярии , фораминиферы и Apicomplexa ). | ||
| Инфузории |  | Ресничка ресница (лат. ) — это крошечный жгутик. Инфузории используют несколько ресничек, число которых может исчисляться сотнями, чтобы перемещаться по воде. |  | Парамеция бурсария нажмите, чтобы увидеть реснички | Фораминиферы , а также некоторые морские амебы , инфузории и жгутиконосцы . | |||
| Амебы (амебоиды) |  | Псевдоподии (по-гречески « ложные ноги» ) — это лопастные придатки, которые амебы используют для прикрепления к твердой поверхности и вытягивания себя вперед. Они могут менять свою форму, выдвигая и втягивая ложноножки. [43] |  | Амеба | Встречается во всех основных линиях протистов . Амебоидные клетки встречаются среди простейших , а также в водорослях и грибах . [44] [45] | |||
| Не подвижный | никто |  | Диатомовая водоросль | Кокколитофоры , большинство диатомей и неподвижные виды Phaeocystis. [42] Среди простейших паразитические Apicomplexa неподвижны. | ||||
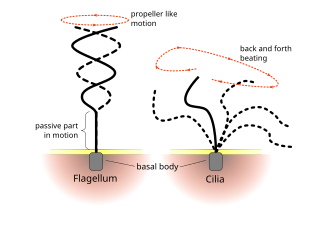
Жгутики используются у прокариот (архей и бактерий), а также у протистов. Кроме того, и жгутики, и реснички широко используются в эукариотических клетках (растительных и животных), помимо протистов.
Регулярные ритмы работы ресничек и жгутиков эукариот вызывают движение на клеточном уровне. Примеры варьируются от продвижения отдельных клеток, например, плавания сперматозоидов , до транспортировки жидкости вдоль стационарного слоя клеток, например, в дыхательных путях . Хотя эукариотические жгутики и подвижные реснички ультраструктурно идентичны, характер движения двух органелл может быть разным. В случае жгутиков движение часто плоское и волнообразное, тогда как подвижные реснички часто совершают более сложное трехмерное движение с силовым и восстановительным ходом.
Эукариотические жгутики — жгутики животных, растений и простейших — представляют собой сложные клеточные отростки, которые перемещаются вперед и назад. классифицируются Эукариотические жгутики вместе с эукариотическими подвижными ресничками как ундулиподии. [46] чтобы подчеркнуть их особую роль волнистых придатков в клеточных функциях или подвижности . Первичные реснички неподвижны и не являются ундулиподиями.
Криптаулакс , Аболлифер , Бодо , Ринхомонада , Спасибо , Аллас и Метромонас. [47]
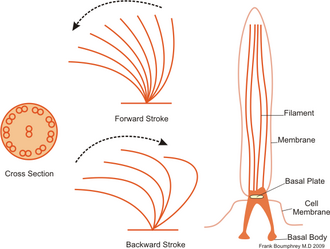
Инфузории обычно имеют от сотен до тысяч ресничек, которые плотно упакованы в массивы. Как и жгутики, реснички приводятся в действие специализированными молекулярными моторами . Эффективный гребок вперед выполняется с застывшим жгутиком, за которым следует неэффективный гребок назад с расслабленным жгутиком. Во время движения отдельная ресничка деформируется, поскольку она использует силовые удары с высоким коэффициентом трения и восстановительные движения с низким коэффициентом трения. Поскольку в отдельном организме имеется множество ресничек, упакованных вместе, они демонстрируют коллективное поведение в метахрональном ритме . Это означает, что деформация одной реснички синфазна с деформацией соседней, вызывая волны деформации, распространяющиеся по поверхности организма. Эти распространяющиеся волны ресничек позволяют организму скоординировано использовать реснички для движения. Типичным примером реснитчатого микроорганизма является Paramecium , одноклеточное реснитчатое простейшее, покрытое тысячами ресничек. Сбивающиеся вместе реснички позволяют Парамеция движется по воде со скоростью 500 микрометров в секунду. [48]
- Жгутиковые, инфузории и амебы

 Парамеции питаются бактериями
Парамеции питаются бактериями Инфузория Oxytricha trifallax с хорошо заметными ресничками.
Инфузория Oxytricha trifallax с хорошо заметными ресничками. Амеба с проглоченными диатомовыми водорослями
Амеба с проглоченными диатомовыми водорослями




| Внешние видео | |
|---|---|
Морские водоросли
[ редактировать ]| Часть серии о |
| Планктон |
|---|
 |
Водоросли — неофициальный термин для широко распространенной и разнообразной группы фотосинтезирующих протистов , которые не обязательно тесно связаны между собой и, таким образом, являются полифилетическими . Морские водоросли можно разделить на шесть групп: зеленые , красные и бурые водоросли , эвгленофиты , динофлагелляты и диатомовые водоросли .
Динофлагелляты и диатомеи являются важными компонентами морских водорослей и имеют отдельные разделы ниже. Эвгленофиты — это тип одноклеточных жгутиконосцев, состоящий всего из нескольких морских представителей.
Не все водоросли микроскопические. Зеленые, красные и бурые водоросли имеют многоклеточные макроскопические формы, которые составляют знакомые морские водоросли . Зеленые водоросли , неофициальная группа, насчитывает около 8000 признанных видов. [49] Многие виды живут большую часть своей жизни в виде одиночных клеток или являются нитевидными, тогда как другие образуют колонии, состоящие из длинных цепочек клеток, или представляют собой высокодифференцированные макроскопические водоросли. Красные водоросли — (спорный) тип, насчитывающий около 7000 признанных видов. [50] в основном многоклеточные и включают множество известных морских водорослей. [50] [51] Бурые водоросли образуют класс, содержащий около 2000 признанных видов. [52] в основном многоклеточные и включают множество морских водорослей, таких как ламинария .В отличие от высших растений, у водорослей отсутствуют корни, стебли и листья. По размеру их можно разделить на микроводоросли и макроводоросли .
Микроводоросли — это микроскопические виды водорослей, невидимые невооруженным глазом. В основном это одноклеточные виды, существующие по отдельности, цепочками или группами, хотя некоторые из них являются многоклеточными . Микроводоросли являются важными компонентами морских протистов, обсуждавшихся выше , а также фитопланктона, обсуждаемого ниже . Они очень разнообразны . По оценкам, существует 200 000–800 000 видов, из которых описано около 50 000 видов. [53] В зависимости от вида их размеры колеблются от нескольких микрометров (мкм) до нескольких сотен микрометров. Они специально адаптированы к среде, в которой преобладают силы вязкости.
 Chlamydomonas globosa , одноклеточная зеленая водоросль с двумя жгутиками , едва заметными внизу слева.
Chlamydomonas globosa , одноклеточная зеленая водоросль с двумя жгутиками , едва заметными внизу слева.
 Центрическая диатомовая водоросль
Центрическая диатомовая водоросль Динофлагелляты
Динофлагелляты




Макроводоросли — это более крупные, многоклеточные и более заметные виды водорослей, обычно называемые морскими водорослями . Морские водоросли обычно растут на мелководье в прибрежных водах, где они прикреплены к морскому дну с помощью крепления . Как и микроводоросли, макроводоросли (водоросли) можно рассматривать как морские протисты, поскольку они не являются настоящими растениями. Но они не являются микроорганизмами, поэтому не входят в задачу данной статьи.
Одноклеточные организмы обычно микроскопические, длиной менее одной десятой миллиметра. Есть исключения. Бокал Русалки , род субтропических зеленых водорослей , является одноклеточным, но удивительно большим и сложным по форме с одним большим ядром, что делает его модельным организмом для изучения клеточной биологии . [55] Другая одноклеточная водоросль, Caulerpa Taxifolia , имеет вид сосудистого растения с «листьями», аккуратно расположенными на стеблях, как у папоротника. Селективное разведение в аквариумах с целью получения более выносливых штаммов привело к случайному попаданию в Средиземное море, где они превратились в инвазивный вид, известный в просторечии как водоросли-убийцы . [56]
Диатомовые водоросли
[ редактировать ]
Диатомеи — это фотосинтезирующие одноклеточные водоросли, населяющие океаны и другие воды по всему земному шару. Они образуют (спорный) тип, содержащий около 100 000 признанных видов. Ежегодно диатомовые водоросли производят около 20 процентов всего кислорода, производимого на планете. [26] поглощают более 6,7 миллиардов тонн кремния из вод, в которых они обитают. и ежегодно [57] Они производят 25–45% всей первичной продукции органического материала в океанах. [58] [59] [60] из-за их распространенности в районах открытого океана, когда общая биомасса фитопланктона максимальна. [61] [62]
Диатомовые водоросли заключены в защитные кремнеземные (стеклянные) оболочки, называемые панцирями . Их классифицируют по форме стеклянных клеток, в которых они живут и которые строят по мере роста. Каждая панцирь состоит из двух взаимосвязанных частей, покрытых крошечными отверстиями, через которые диатомовые водоросли обмениваются питательными веществами и отходами. [63] Мертвые диатомовые водоросли дрейфуют на дно океана, где за миллионы лет остатки их панцирей могут накапливаться на глубине до полумили . [64] Диатомовые водоросли имеют относительно высокие скорости погружения по сравнению с другими группами фитопланктона, и на их долю приходится около 40% твердых частиц углерода, экспортируемых в глубины океана. [60] [65] [62]
 Диатомовые водоросли — один из наиболее распространенных видов фитопланктона.
Диатомовые водоросли — один из наиболее распространенных видов фитопланктона. Их защитные оболочки (фрустлы) изготовлены из кремния.
Их защитные оболочки (фрустлы) изготовлены из кремния.








| Внешние видео | |
|---|---|

Физически обусловленное сезонное обогащение поверхностных питательных веществ благоприятствует цветению диатомовых водорослей . Антропогенное изменение климата напрямую повлияет на эти сезонные циклы, изменив сроки цветения и уменьшив их биомассу, что приведет к сокращению первичного производства и поглощения CO 2 . [67] [62] Данные дистанционного зондирования показывают, что в период с 1998 по 2012 год произошло глобальное сокращение численности диатомовых водорослей, особенно в северной части Тихого океана, что связано с обмелением поверхностного смешанного слоя и снижением концентрации питательных веществ. [68] [62]
 Кварцованная панцирь пеннатной диатомеи с двумя перекрывающимися половинками.
Кварцованная панцирь пеннатной диатомеи с двумя перекрывающимися половинками. Guinardia delicatula , диатомовая водоросль, ответственная за цветение диатомей в Северном море. [69]
Guinardia delicatula , диатомовая водоросль, ответственная за цветение диатомей в Северном море. [69] Существует более 100 000 видов диатомовых водорослей, на которые приходится 25–45% первичной продукции океана.
Существует более 100 000 видов диатомовых водорослей, на которые приходится 25–45% первичной продукции океана.



 Связанные диатомовые водоросли
Связанные диатомовые водоросли![Пеннатная диатомовая водоросль из арктического талого пруда, зараженная двумя хитридоподобными грибковыми патогенами. Масштабная линейка = 10 мкм.[70]](//upload.wikimedia.org/wikipedia/commons/thumb/b/b5/Pennate_diatom_infected_with_two_chytrid-like_fungal_pathogens.png/280px-Pennate_diatom_infected_with_two_chytrid-like_fungal_pathogens.png) Пеннатная диатомовая водоросль из арктического плавильного пруда , зараженная двумя хитридоподобными грибковыми патогенами. Масштабная линейка = 10 мкм. [70]
Пеннатная диатомовая водоросль из арктического плавильного пруда , зараженная двумя хитридоподобными грибковыми патогенами. Масштабная линейка = 10 мкм. [70]

![Пеннатная диатомовая водоросль из арктического талого пруда, зараженная двумя хитридоподобными грибковыми патогенами. Масштабная линейка = 10 мкм.[70]](http://upload.wikimedia.org/wikipedia/commons/thumb/b/b5/Pennate_diatom_infected_with_two_chytrid-like_fungal_pathogens.png/280px-Pennate_diatom_infected_with_two_chytrid-like_fungal_pathogens.png)
Кокколитофоры
[ редактировать ]

Кокколитофоры — это мелкие одноклеточные фотосинтезирующие протисты с двумя жгутиками для передвижения. Большинство из них защищены раковинами из карбоната кальция, покрытыми декоративными круглыми пластинками или чешуйками, называемыми кокколитами . Термин кокколитофор происходит от греческого слова « камень, несущий семя» , что связано с их небольшим размером и камнями кокколита, которые они несут. При правильных условиях они цветут, как и другой фитопланктон, и могут сделать океан молочно-белым. [72]

 Кокколитофор . Emiliania huxleyi
Кокколитофор . Emiliania huxleyi Цветение водорослей Emiliania huxleyi у южного побережья Англии.
Цветение водорослей Emiliania huxleyi у южного побережья Англии.



Динофлагелляты
[ редактировать ]

Динофлагелляты обычно относят к группе водорослей и образуют тип одноклеточных жгутиконосцев, насчитывающий около 2000 морских видов. [75] Название происходит от греческого «динос», что означает вращение , и латинского «жгутик», означающего кнут или плеть . Это относится к двум хлыстообразным приспособлениям (жгутикам), используемым для движения вперед. Большинство динофлагеллят защищены красно-коричневой целлюлозной броней. Как и другой фитопланктон, динофлагелляты являются r-стратегами , которые при правильных условиях могут цвести и вызывать красные приливы . Раскопки могут быть самой базальной линией жгутиковых. [41]
По трофической направленности динофлагелляты встречаются повсеместно. Известно, что некоторые динофлагелляты фотосинтезируют , но большая часть из них на самом деле миксотрофны , сочетая фотосинтез с поеданием добычи ( фаготрофия ). [76] Некоторые виды являются эндосимбионтами морских животных и других протистов и играют важную роль в биологии коралловых рифов . Другие предшествуют другим простейшим, а некоторые формы являются паразитическими. Многие динофлагелляты миксотрофны и их также можно отнести к фитопланктону.
Токсичная динофлагеллята Dinophys acuta приобретает хлоропласты от своей добычи. «Он не может поймать криптофиты сам по себе и вместо этого полагается на заглатывание инфузорий, таких как красный Mesodinium Rubrum , которые изолируют свои хлоропласты от определенной клады криптофитов (Geminigera/Plagioselmis/Teleaulax)». [39]
 Гиродиний , одна из немногих голых динофлагеллят, лишенных панциря.
Гиродиний , одна из немногих голых динофлагеллят, лишенных панциря. Динофлагеллята Protoperidinium выдвигает большую кормовую вуаль для захвата добычи.
Динофлагеллята Protoperidinium выдвигает большую кормовую вуаль для захвата добычи. Радиолярии насселлярий могут находиться в симбиозе с динофлагеллятами.
Радиолярии насселлярий могут находиться в симбиозе с динофлагеллятами. Динофлагеллята Dinophys acuta.
Динофлагеллята Dinophys acuta.








Динофлагелляты часто живут в симбиозе с другими организмами. Многие насселлярий радиолярии динофлагеллят . симбионты содержат в своих раковинах [78] Насселлярия обеспечивает аммонием и углекислым газом , а динофлагеллята обеспечивает насселлярию слизистой оболочкой, полезной для охоты и защиты от вредных захватчиков. динофлагелляту [79] свидетельствует Анализ ДНК о том, что симбиоз динофлагеллят с радиоляриями развился независимо от других симбиозов динофлагеллят, например, с фораминиферами . [80]
Некоторые динофлагелляты биолюминесцентны . Ночью океанская вода может светиться изнутри и сверкать синим светом из-за этих динофлагеллят. [81] [82] Биолюминесцентные динофлагелляты обладают сцинтиллонами , отдельными цитоплазматическими тельцами, содержащими динофлагеллятлюциферазу , основной фермент, участвующий в люминесценции. Люминесценция, иногда называемая фосфоресценцией моря , возникает в виде коротких (0,1 секунды) синих вспышек или искр при стимуляции отдельных сцинтиллонов, обычно в результате механических воздействий, например, от лодки, пловца или прибоя. [83]
 Tripos muelleri можно узнать по рогам U-образной формы.
Tripos muelleri можно узнать по рогам U-образной формы.![Oodinium, род паразитических динофлагеллят, вызывает у рыб бархатную болезнь.[84]](//upload.wikimedia.org/wikipedia/commons/thumb/8/87/Archives_de_zoologie_exp%C3%A9rimentale_et_g%C3%A9n%C3%A9rale_%281920%29_%2820299351186%29.jpg/260px-Archives_de_zoologie_exp%C3%A9rimentale_et_g%C3%A9n%C3%A9rale_%281920%29_%2820299351186%29.jpg)
 Karenia brevis вызывает красные приливы, очень токсичные для человека. [85]
Karenia brevis вызывает красные приливы, очень токсичные для человека. [85]

![Oodinium, род паразитических динофлагеллят, вызывает у рыб бархатную болезнь.[84]](http://upload.wikimedia.org/wikipedia/commons/thumb/8/87/Archives_de_zoologie_exp%C3%A9rimentale_et_g%C3%A9n%C3%A9rale_%281920%29_%2820299351186%29.jpg/260px-Archives_de_zoologie_exp%C3%A9rimentale_et_g%C3%A9n%C3%A9rale_%281920%29_%2820299351186%29.jpg)


 Noctiluca scintillans — биолюминесцентная динофлагеллята. [86]
Noctiluca scintillans — биолюминесцентная динофлагеллята. [86] Ornithocercus гетеропорус - выставлены выдающиеся списки
Ornithocercus гетеропорус - выставлены выдающиеся списки



Морские простейшие
[ редактировать ]Простейшие — это протисты, которые питаются органическими веществами, такими как другие микроорганизмы или органические ткани и мусор. [87] [88] Исторически простейшие считались «одноклеточными животными», поскольку они часто обладают животным поведением, например, подвижностью и хищничеством , и лишены клеточной стенки , как это наблюдается у растений и многих водорослей . [89] [90] Хотя традиционная практика группирования простейших с животными больше не считается обоснованной, этот термин продолжает использоваться в широком смысле для обозначения одноклеточных организмов, которые могут передвигаться независимо и питаться путем гетеротрофии .
К морским простейшим относятся зоофлагелляты , фораминиферы , радиолярии и некоторые динофлагелляты .
Радиолярии
[ редактировать ]

Радиолярии — это одноклеточные хищные протисты , заключенные в сложные шаровидные оболочки, обычно размером от 0,1 до 0,2 миллиметра, обычно сделанные из кремнезема и пронизанные отверстиями. Их название происходит от латинского слова «радиус». Они ловят добычу, вытягивая части своего тела через отверстия. Как и в случае с кремнеземными панцирями диатомей, раковины радиолярий могут опускаться на дно океана, когда радиолярии умирают, и сохраняются как часть океанских отложений . Эти останки в виде микрокаменелостей предоставляют ценную информацию о прошлых условиях океана. [91]
 Как и диатомеи, радиолярии бывают разных форм.
Как и диатомеи, радиолярии бывают разных форм. Раковины радиолярий, как и диатомеи, обычно состоят из силиката.
Раковины радиолярий, как и диатомеи, обычно состоят из силиката. Однако акантарианские радиолярии имеют раковины из кристаллов сульфата стронция .
Однако акантарианские радиолярии имеют раковины из кристаллов сульфата стронция . Схематическая диаграмма сферической раковины радиолярий в разрезе
Схематическая диаграмма сферической раковины радиолярий в разрезе





близко повторяют структуру раковин некоторых радиолярий. [92]
| Внешние видео | |
|---|---|
 Кладококк абиетинус
Кладококк абиетинус Клевейплегма северная
Клевейплегма северная


Фораминиферы
[ редактировать ]

Как и радиолярии, фораминиферы ( сокращенно форамы ) представляют собой одноклеточные хищные протисты, также защищенные раковинами с отверстиями. Их название происходит от латинского слова «носители дыр». Их раковины, часто называемые панцирями , имеют камеры (по мере роста у них появляются дополнительные камеры). Раковины обычно состоят из кальцита, но иногда из склеенных частиц осадка или хитона и (редко) кремнезема. Большинство форамов донные, но около 40 видов — планктонные. [93] Они широко исследуются с использованием хорошо известных летописей окаменелостей, которые позволяют ученым делать много выводов о окружающей среде и климате прошлого. [91]


| Внешние видео | |
|---|---|
 Разрез, показывающий камеры спирального отверстия.
Разрез, показывающий камеры спирального отверстия. Живая аммиачная тепида, струящая гранулированную эктоплазму для улавливания пищи
Живая аммиачная тепида, струящая гранулированную эктоплазму для улавливания пищи Группа планктонных форм.
Группа планктонных форм. Ископаемые образования нуммулитид разных размеров из эоцена.
Ископаемые образования нуммулитид разных размеров из эоцена.![Египетские пирамиды были построены из известняка, содержащего нуммулиты.[94]](//upload.wikimedia.org/wikipedia/commons/thumb/a/af/All_Gizah_Pyramids.jpg/325px-All_Gizah_Pyramids.jpg)




![Египетские пирамиды были построены из известняка, содержащего нуммулиты.[94]](http://upload.wikimedia.org/wikipedia/commons/thumb/a/af/All_Gizah_Pyramids.jpg/325px-All_Gizah_Pyramids.jpg)
Ряд форумов миксотрофны ( см. ниже ). У них есть одноклеточные водоросли как эндосимбионты , принадлежащие к различным линиям, таким как зеленые водоросли , красные водоросли , золотые водоросли , диатомовые водоросли и динофлагелляты . [93] Миксотрофные фораминиферы особенно распространены в бедных питательными веществами океанических водах. [95] Некоторые отверстия являются клептопластическими , сохраняя хлоропласты из проглоченных водорослей для проведения фотосинтеза . [96]
Амеба
[ редактировать ]

 Обнаженная амеба с пищевыми вакуолями и проглоченной диатомовой водорослью.
Обнаженная амеба с пищевыми вакуолями и проглоченной диатомовой водорослью.
 Ксеногенная раковинная амеба, покрытая диатомовыми водорослями (из коллекции амеб Пенарда )
Ксеногенная раковинная амеба, покрытая диатомовыми водорослями (из коллекции амеб Пенарда )



| Внешние видео | |
|---|---|
Инфузории
[ редактировать ]
Морские инфузории являются основными пожирателями фитопланктона. [97] [98]
Первичная продукция фитопланктона поддерживает более высокие трофические уровни и способствует микробной реминерализации . [99] [100] Доминирующие пелагические поедающие фитопланктон обычно связаны с различными режимами работы отделов пищевой сети и круговоротом питательных веществ . Гетеротрофные травоядные и доминирование микрозоопланктона обычно связаны с микробной петлей и регенерированной продукцией; в то время как мезозоопланктон связан с линейной пищевой цепью и экспортным производством . [101] [102] Потребление первичной продукции твердых частиц на поверхности мирового океана составляет ~ 10–15% для мезозоопланктона и 59–75% для микрозоопланктона. [103] [104] [105] [106] с оценками для прибрежных и устьевых систем обычно в более низком диапазоне. [106] [98]
Инфузории составляют важный компонент сообщества микрозоопланктона, отдавая предпочтение жертвам мелких размеров, в отличие от мезозоопланктона, а многие виды инфузорий поедаются также мезозоопланктоном. [107] Таким образом, инфузории могут быть важным связующим звеном между мелкими клетками и высшими трофическими уровнями. [108] Помимо своей значительной роли в переносе углерода, инфузории также считаются высококачественной пищей как источник белковых соединений с низким соотношением C:N по сравнению с фитопланктоном. [109] [110] [98]
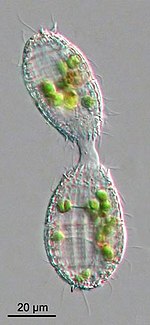
Хотя многие инфузории являются гетеротрофами, ряд пелагических видов являются миксотрофами , сочетающими в себе как фаготрофное , так и фототрофное питание (Stoecker, 1998). Признание миксотрофии в пищевой сети морского планктона поставило под сомнение классическое понимание пелагических пищевых сетей, поскольку автотрофия и гетеротрофия не обязательно являются двумя отдельными функциональными компартментами. [111] Классическое понимание экологических взаимодействий между планктоном, таких как конкуренция за питательные вещества, указывает на то, что сродство к поглощению питательных веществ уменьшается с размером организма. [112] предпочтение меньшим размерам в условиях ограниченности ресурсов. Миксотрофия выгодна для организмов в условиях ограниченности питательных веществ, позволяя им уменьшить прямую конкуренцию за счет выпаса более мелкой добычи и увеличить прямое потребление питательных веществ. [113] Результаты моделирования показывают, что миксотрофия благоприятствует более крупным организмам и, следовательно, повышает эффективность трофического переноса. [113] [114] Вдобавок ко всему, миксотрофия, по-видимому, важна как в пространстве, так и во времени в морских системах. [115] подчеркивая необходимость экологических полевых исследований для дальнейшего выяснения роли миксотрофии. [98]
 Тинтиннопсис колокольчик
Тинтиннопсис колокольчик

 Морская инфузория Strombidium rassoulzadegani .
Морская инфузория Strombidium rassoulzadegani .




 голофиры Яйцо
голофиры Яйцо


- Несколько таксонов инфузорий взаимодействуют
- Blepharisma americanum плавает в капле прудовой воды вместе с другими микроорганизмами.
| Внешние видео | |
|---|---|
Макроскопические протисты
[ редактировать ]- Макроскопические протисты (см. также одноклеточные макроводоросли → )
 Одноклеточная гигантская амеба имеет до 1000 ядер и достигает длины 5 мм.
Одноклеточная гигантская амеба имеет до 1000 ядер и достигает длины 5 мм.![Gromia sphaerica — крупная сферическая раковинная амеба, оставляющая грязевые следы. Его диаметр до 3,8 см.[116]](//upload.wikimedia.org/wikipedia/commons/thumb/9/99/Gromia_in_situ_closeup.png/266px-Gromia_in_situ_closeup.png) Gromia sphaerica — крупная сферическая раковинная амеба , оставляющая грязевые следы. Его диаметр до 3,8 см. [116]
Gromia sphaerica — крупная сферическая раковинная амеба , оставляющая грязевые следы. Его диаметр до 3,8 см. [116] Spiculosiphon Oceana — одноклеточный фораминифер , внешним видом и образом жизни имитирующий губку , вырастает до 5 см в длину.
Spiculosiphon Oceana — одноклеточный фораминифер , внешним видом и образом жизни имитирующий губку , вырастает до 5 см в длину.![Ксенофиофор, еще один одноклеточный фораминифер, обитает в абиссальных зонах. Имеет гигантскую раковину диаметром до 20 см.[117]](//upload.wikimedia.org/wikipedia/commons/thumb/5/5d/Xenophyophore.jpg/292px-Xenophyophore.jpg) Ксенофиофор абиссальных , еще один одноклеточный фораминифер, обитает в зонах . Имеет гигантскую раковину диаметром до 20 см. [117]
Ксенофиофор абиссальных , еще один одноклеточный фораминифер, обитает в зонах . Имеет гигантскую раковину диаметром до 20 см. [117] Гигантская ламинария , бурая водоросль , не является настоящим растением, однако она многоклеточная и может достигать 50 м в высоту.
Гигантская ламинария , бурая водоросль , не является настоящим растением, однако она многоклеточная и может достигать 50 м в высоту.

![Gromia sphaerica — крупная сферическая раковинная амеба, оставляющая грязевые следы. Его диаметр до 3,8 см.[116]](http://upload.wikimedia.org/wikipedia/commons/thumb/9/99/Gromia_in_situ_closeup.png/266px-Gromia_in_situ_closeup.png)

![Ксенофиофор, еще один одноклеточный фораминифер, обитает в абиссальных зонах. Имеет гигантскую раковину диаметром до 20 см.[117]](http://upload.wikimedia.org/wikipedia/commons/thumb/5/5d/Xenophyophore.jpg/292px-Xenophyophore.jpg)

Планктонные протисты
[ редактировать ]Интерактом
[ редактировать ]
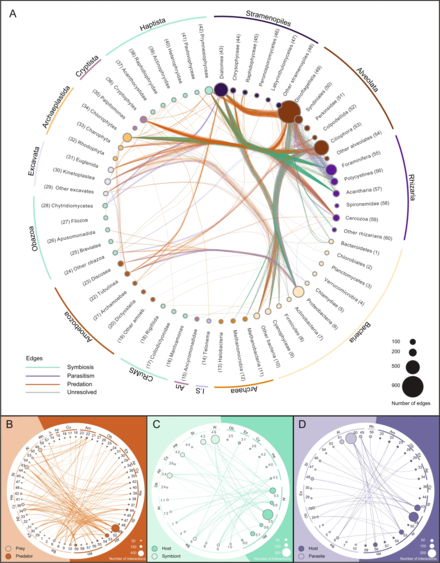
Двусторонние сети, предоставляющие обзор взаимодействий, представленных вручную курируемой базой данных взаимодействия протистов (PIDA).
Взаимодействие между видами микробов сыграло важную роль в эволюции и видообразовании. [118] Одним из лучших примеров является то, что происхождение эукариот основано на взаимодействиях эндосимбиоза ; создавая митохондрии , хлоропласты и другие метаболические способности в эукариотической клетке, [119] [120] [121] [122] Взаимодействия микробов гарантируют функционирование экосистемы, играя решающую роль, например, в передаче углерода в фотосимбиозе, контроле цветения микроводорослей паразитами и связанных с фитопланктоном бактерий, влияющих на рост и здоровье своего хозяина. [118]
Несмотря на их важность, понимание микробных взаимодействий в океане и других водных системах находится в зачаточном состоянии, и большинство из них до сих пор неизвестны. [13] [123] [124] [125] Самые ранние исследования взаимодействия водных микробов относятся к XIX веку. В 1851 году, находясь на борту HMS Rattlesnake в Тихом океане, Томас Хаксли обнаружил маленькие желто-зеленые клетки внутри заметных планктонных радиолярий, которые, по его мнению, были органеллами . [126] Позже Карл Брандт установил, что желтоватые клетки являются симбиотическими водорослями, и назвал их зооксантеллой . [127] Со времени этих ранних исследований сотни других сообщили о микробных взаимодействиях, используя классические инструменты, в основном микроскопию, но эти знания еще не собраны в одну доступную базу данных. В последние годы высокопроизводительное секвенирование (HTS) [128] [129] [130] изменили ДНК или РНК окружающей среды понимание микробного разнообразия [131] и эволюция, [132] а также создание гипотез о микробных взаимодействиях на основе корреляций предполагаемой численности микробов в пространственно-временных масштабах. [133] [134] [135] [136] [118]
Диаграмма справа представляет собой обзор взаимодействий между планктонными протистами, записанных в вручную создаваемой базе данных взаимодействия протистов (PIDA). Сеть основана на 2422 экологических взаимодействиях в PIDA, зарегистрированных на основе ~500 публикаций за последние 150 лет. Номенклатура и таксономический порядок эукариот основаны на Adl et al. 2019. [137] Номенклатура и таксономический порядок бактерий основаны на Schultz et al. 2017. [138] [118]
Узлы сгруппированы (внешний круг) в соответствии с супергруппами эукариот (или Incertae sedis ), бактериями и архей. Все основные линии протистанов участвовали во взаимодействиях в качестве хозяев, симбионтов (мутуалистов и комменсалистов), паразитов, хищников и/или жертв. Хищничество было наиболее распространенным взаимодействием (39%), за ним следовали симбиоз (29%), паразитизм (18%) и неразрешенные взаимодействия (14%, когда неясно, является ли взаимодействие полезным или антагонистическим). Узлы представляют эукариотические и прокариотические таксоны и окрашены соответствующим образом. Размер узла указывает количество ребер/ссылок, подключенных к этому узлу. Каждому узлу/таксону присвоен номер, который соответствует номерам таксонов в B, C и D. Края представляют взаимодействия между двумя таксонами и окрашены в соответствии с типом экологического взаимодействия: хищничество (оранжевый), симбиоз (зеленый) и паразитизм. (фиолетовый). [118]
Сеть ненаправленная, то есть узел может содержать как паразитов/симбионтов/добычу, так и хозяев/хищников. Чтобы не загромождать рисунок, «самопетли», представляющие случаи, когда оба взаимодействующих организма принадлежат к одному и тому же таксону (например, динофлагеллята, поедающая другую динофлагелляту), не показаны на этом рисунке в виде ребер/звеньев, а рассматриваются в размер узлов. Самый внешний круг группирует таксоны в различных «супергруппах» эукариот или прокариотических доменах Bacteria и Archaea. Ancryomonadidae сокращенно An. Телонема не отнесена ни к одной из супергрупп, а классифицируется как Incertae sedis (на рисунке сокращенно IS). В B, B и D используются следующие сокращения супергрупп: Ar Archaea, Ba Bacteria, Rh Rhizaria , Al Alveolata , St Stramenopiles , Ha Haptista , Cy Cryptista , Ap Archaeplastida , Ex Excavata , Ob Obazoa , Am Amoebozoa , Cu CRuMS. , Ancryomonadidae, Is Incertae sedis . [118]
B: Взаимодействие хищник-жертва в PIDA. Номера узлов соответствуют номерам узлов таксонов в a. Сокращения супергрупп описаны выше. Фон и узлы окрашены в соответствии с функциональной ролью во взаимодействии: жертвы окрашены в светло-оранжевый цвет (левая часть рисунка), а хищники изображены в темно-оранжевый цвет (правая часть рисунка). Размер каждого узла представляет собой количество ребер, соединенных с этим узлом. [118]
C. Взаимодействия симбионт-хозяин, включенные в PIDA. Номера узлов соответствуют номерам узлов в A. Сокращения для супергрупп описаны выше. Симбионты слева окрашены в светло-зеленый цвет, а их хозяева справа - в темно-зеленый. Размер каждого узла представляет собой количество ребер, соединенных с этим узлом. [118]
D: Взаимодействия паразит-хозяин, включенные в PIDA. Номера узлов соответствуют номерам узлов в A. Сокращения для супергрупп описаны выше. Таксоны паразитов изображены светло-фиолетовым цветом (слева), хозяева - темно-фиолетовым (справа). [118]
Было обнаружено, что хищники-протисты кажутся «многоядными», в то время как взаимодействия паразит-хозяин и симбионт-хозяин имеют умеренную степень специализации. Супергруппа SAR (т. е. Stramenopiles, Alveolata и Rhizaria) в значительной степени доминировала в PIDA, и сравнение с молекулярным исследованием глобального океана ( экспедиция Тары ) показало, что несколько линий SAR, которые богаты и разнообразны в морской сфере, были недостаточно представлены среди записанные взаимодействия. [118]
Раковины протистов
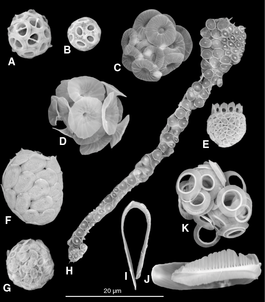
[ редактировать ]Многие протисты имеют защитные панцири или панцири . [139] обычно изготавливается из карбоната кальция (мела) или кремнезема (стекла). Протисты в основном одноклеточные и микроскопические. Их панцири часто представляют собой твердые минерализованные формы, которые устойчивы к разложению и могут пережить смерть протистов как микроископаемые . Хотя протисты очень малы, они повсеместно распространены. Их численность такова, что их раковины играют огромную роль в формировании океанских отложений , а также в глобальном круговороте элементов и питательных веществ.
Панцири диатомей называются панцирями и состоят из кремнезема. Эти стеклянные структуры накапливались более 100 миллионов лет, оставив по всему земному шару богатые залежи нано- и микроструктурированного оксида кремния в виде диатомита . Эволюционные причины образования нано- и микроструктурированного кремнезема фотосинтезирующими водорослями еще не ясны. Однако в 2018 году было показано, что отражение ультрафиолетового света наноструктурированным кремнеземом защищает ДНК в клетках водорослей, и это может быть эволюционной причиной образования стеклянных клеток. [140] [141]
Кокколитофороры защищены панцирем, состоящим из декоративных круглых пластинок или чешуек, называемых кокколитами . Кокколиты состоят из карбоната кальция или мела. Термин кокколитофор происходит от греческого слова « камень, несущий семя» , что связано с их небольшим размером и камнями кокколита, которые они несут. [72]




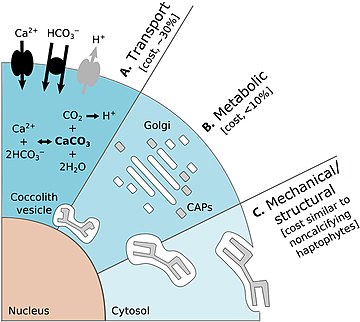
Есть преимущества для протистов, имеющих защитный панцирь. На диаграмме слева выше показаны некоторые преимущества, которые кокколитофоры получают от ношения кокколитов. На диаграмме (A) представлен ускоренный фотосинтез, включая механизмы концентрации углерода (CCM) и повышенное поглощение света за счет рассеяния редких фотонов глубоко обитающими видами. (B) представляет собой защиту от фотоповреждений, включая защиту солнцезащитного козырька от ультрафиолетового света (УФ) и фотосинтетического активного излучения (ФАР), а также рассеивание энергии в условиях яркого освещения. (C) представляет собой броневую защиту, включающую защиту от вирусных/бактериальных инфекций и выпаса селективными и неселективными травоядными. [144]
Есть также затраты на простейших, имеющих защитные панцири. Диаграмма справа выше показывает некоторые энергетические затраты, которые несет кокколитофор при переноске кокколитов. На диаграмме энергетические затраты указаны в процентах от общего фотосинтетического бюджета. (А) представляет собой транспортные процессы, включающие транспорт в клетку из окружающей морской воды субстратов первичной кальцификации Ca 2 + и HCO 3 - (черные стрелки) и удаление конечного продукта H+ из клетки (серая стрелка). Транспорт Ca 2+ через цитоплазму к везикуле кокколита (CV) является доминирующей затратой, связанной с кальцификацией. (B) представляет собой метаболические процессы, включающие синтез связанных с кокколитом полисахаридов (CAP – серые прямоугольники) комплексом Гольджи (белые прямоугольники), которые регулируют зарождение и геометрию кристаллов CaCO 3 . Завершенный кокколит (серая пластинка) представляет собой сложную структуру из сложно расположенных CAP и CaCO 3 кристаллов . (C) Механические и структурные процессы ответственны за секрецию завершенных кокколитов, которые транспортируются из своего исходного положения, прилегающего к ядру, на периферию клетки, где они переносятся на поверхность клетки. [144]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Кавалер-Смит Т. (декабрь 1993 г.). «Царство простейших и его 18 типов» . Микробиологические обзоры . 57 (4): 953–94. doi : 10.1128/mmbr.57.4.953-994.1993 . ПМК 372943 . ПМИД 8302218 .
- ^ Корлисс Дж.О. (1992). «Должен ли быть отдельный кодекс номенклатуры для протистов?». БиоСистемы . 28 (1–3): 1–14. Бибкод : 1992BiSys..28....1C . дои : 10.1016/0303-2647(92)90003-H . ПМИД 1292654 .
- ^ Слапета Х., Морейра Д., Лопес-Гарсия П. (2005). «Степень разнообразия протистов: выводы из молекулярной экологии пресноводных эукариот» . Труды Королевского общества B: Биологические науки . 272 (1576): 2073–81. дои : 10.1098/rspb.2005.3195 . ПМЦ 1559898 . ПМИД 16191619 .
- ^ Морейра Д., Лопес-Гарсия П. (2002). «Молекулярная экология микробных эукариот открывает скрытый мир» (PDF) . Тенденции в микробиологии . 10 (1): 31–8. дои : 10.1016/S0966-842X(01)02257-0 . ПМИД 11755083 .
- ^ Jump up to: а б с Лелес, С.Г.; Митра, А.; Флинн, К.Дж.; Стокер, ДК; Хансен, П.Дж.; Кальбет, А.; Макманус, Великобритания; Сандерс, RW; Кэрон, Д.А.; Нет, Ф.; Халлеграефф, генеральный директор (2017). «Океанические протисты с различными формами приобретенной фототрофии демонстрируют контрастную биогеографию и численность» . Труды Королевского общества B: Биологические науки . 284 (1860): 20170664. doi : 10.1098/rspb.2017.0664 . ПМЦ 5563798 . ПМИД 28768886 .
- ^ Характеристики протистов в: Рожь, Конни; Ависар, Яэль; Чхве, Юнг Хо; ДеСэ, Жан; Жруковский, Владимир; Уайз, Роберт Р. (2013). Биология . Хьюстон, Техас. ISBN 978-1-938168-09-3 . OCLC 896421272 .
{{cite book}}: CS1 maint: отсутствует местоположение издателя ( ссылка ) Измененный текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
Измененный текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 . - ^ Девреотес П. (1989). «Dictyostelium discoideum: модельная система межклеточных взаимодействий в развитии». Наука . 245 (4922): 1054–8. Бибкод : 1989Sci...245.1054D . дои : 10.1126/science.2672337 . ПМИД 2672337 .
- ^ Нил AC, Рис JB, Саймон EJ (2004) Основная биология с физиологией Пирсон / Бенджамин Каммингс, страница 291. ISBN 9780805375039
- ^ О'Мэлли М.А., Симпсон А.Г., Роджер А.Дж. (2012). «Другие эукариоты в свете эволюционной протистологии». Биология и философия . 28 (2): 299–330. дои : 10.1007/s10539-012-9354-y . S2CID 85406712 .
- ^ Адл С.М., Симпсон А.Г., Фармер М.А., Андерсен Р.А., Андерсон О.Р., Барта Дж.Р., Баузер С.С., Брюгеролль Дж., Фенсом Р.А., Фредерик С., Джеймс Т.Я., Карпов С., Кугренс П., Круг Дж., Лейн CE, Льюис Л.А., Лодж Дж. , Линн Д.Х., Манн Д.Г., МакКорт Р.М., Мендоса Л., Моэструп О., Мозли-Стэндридж С.Е., Нерад Т.А., Ширер К.А., Смирнов А.В., Шпигель Ф.В., Тейлор М.Ф. (2005). «Новая классификация эукариот более высокого уровня с упором на таксономию протистов» . Журнал эукариотической микробиологии . 52 (5): 399–451. дои : 10.1111/j.1550-7408.2005.00053.x . ПМИД 16248873 . S2CID 8060916 .
- ^ Маргулис Л., Чепмен М.Дж. (19 марта 2009 г.). Королевства и владения: Иллюстрированный путеводитель по типам жизни на Земле . Академическая пресса. ISBN 9780080920146 .
- ^ Jump up to: а б Кольер, Джеки Л.; Отдых, Джошуа С. (2019). «Плавание, скольжение и скатывание в сторону мейнстрима: клеточная биология морских простейших» . Молекулярная биология клетки . 30 (11): 1245–1248. дои : 10.1091/mbc.E18-11-0724 . ПМК 6724603 . ПМИД 31084566 .
- ^ Jump up to: а б Уорден, Аризона; Следует, MJ; Джованнони, С.Дж.; Уилкен, С.; Циммерман, А.Е.; Килинг, Пи Джей (2015). «Переосмысление морского углеродного цикла: учет разнообразия образа жизни микробов» . Наука . 347 (6223). дои : 10.1126/science.1257594 . ПМИД 25678667 . S2CID 206560125 .
- ^ Де Варгас, К.; и др. (2015). «Разнообразие эукариотического планктона в освещенном солнцем океане» . Наука . 348 (6237). дои : 10.1126/science.1261605 . hdl : 10261/117736 . ПМИД 25999516 . S2CID 12853481 .
- ^ Jump up to: а б Кертис, Брюс А.; и др. (2012). «Геномы водорослей раскрывают эволюционный мозаицизм и судьбу нуклеоморфов» . Природа . 492 (7427): 59–65. Бибкод : 2012Natur.492...59C . дои : 10.1038/nature11681 . ПМИД 23201678 . S2CID 4380094 .
- ^ Jump up to: а б Армбруст, EV; и др. (2004). «Геном диатомовой водоросли Thalassiosira Pseudonana: экология, эволюция и метаболизм» . Наука . 306 (5693): 79–86. Бибкод : 2004Sci...306...79A . дои : 10.1126/science.1101156 . ПМИД 15459382 . S2CID 8593895 .
- ^ Jump up to: а б Прочтите, Бетси А.; и др. (2013). «Пангеном фитопланктона Emiliania лежит в основе его глобального распространения» . Природа . 499 (7457): 209–213. Бибкод : 2013Natur.499..209. . дои : 10.1038/nature12221 . hdl : 1854/LU-4120924 . ПМИД 23760476 . S2CID 4428297 .
- ^ Jump up to: а б Килинг, Патрик Дж.; и др. (2014). «Проект секвенирования транскриптома морских микробов эукариот (MMETSP): освещение функционального разнообразия эукариотической жизни в океанах посредством секвенирования транскриптома» . ПЛОС Биология . 12 (6): e1001889. дои : 10.1371/journal.pbio.1001889 . ПМК 4068987 . ПМИД 24959919 .
- ^ Наймарк, Марианна; Шарма, Амит Кумар; Спарстад, Торфинн; Боунс, Атл М.; Винге, Пер (2016). «Система CRISPR/Cas9, адаптированная для редактирования генов морских водорослей» . Научные отчеты . 6 : 24951. Бибкод : 2016NatSR...624951N . дои : 10.1038/srep24951 . ПМЦ 4842962 . ПМИД 27108533 .
- ^ Надежды, Аманда; Некрасов Владимир; Камун, Софиен; Мок, Томас (2016). «Редактирование гена уреазы с помощью CRISPR-Cas у диатомовой водоросли Thalassiosira pseudonana» . Растительные методы . 12:49 . дои : 10.1186/s13007-016-0148-0 . ПМК 5121945 . ПМИД 27904648 .
- ^ Каррадек, Квентин; и др. (2018). «Глобальный океанический атлас генов эукариот» . Природные коммуникации . 9 (1): 373. Бибкод : 2018NatCo...9..373C . дои : 10.1038/s41467-017-02342-1 . ПМЦ 5785536 . ПМИД 29371626 .
- ^ Jump up to: а б Факторова, Драгомира; и др. (2020). «Разработка генетических инструментов морских протистов: новые модельные организмы для экспериментальной клеточной биологии» . Природные методы . 17 (5): 481–494. дои : 10.1038/s41592-020-0796-x . ПМК 7200600 . ПМИД 32251396 . Измененный текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Уиттакер, Р.Х.; Маргулис, Л. (1978). «Классификация протистов и царства организмов». Биосистемы . 10 (1–2): 3–18. Бибкод : 1978BiSys..10....3W . дои : 10.1016/0303-2647(78)90023-0 . ПМИД 418827 .
- ^ Фор, Э; Нет, Ф; Бенуастон, AS; Лабади, К; Биттнер, Л; Аята, SD (2019). «Миксотрофные протисты демонстрируют контрастную биогеографию в мировом океане» . Журнал ИСМЕ . 13 (4): 1072–1083. Бибкод : 2019ISMEJ..13.1072F . дои : 10.1038/s41396-018-0340-5 . ПМК 6461780 . ПМИД 30643201 .
- ^ Бадд, Грэм Э; Йенсен, Сорен (2017). «Происхождение животных и гипотеза «саванны» ранней двусторонней эволюции» . Биологические обзоры . 92 (1): 446–473. дои : 10.1111/brv.12239 . ПМИД 26588818 .
- ^ Jump up to: а б Воздух, которым вы дышите? Диатомовая водоросль сделала это
- ^ Кларк М.А., Дуглас М. и Чой Дж. (2018) Биология 2e , 23.4 «Экология протистов» , OpenStax, Хьюстон, Техас. Измененный текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Валлет, морской пехотинец; Баумайстер, Тим У.Х.; Кафтан, Филип; Грабе, Вейт; Буайя, Энтони; Тайнс, Марко; Сватош, Алеш; Понерт, Георг (2019). «Оомицет Lagenisma coscinodisci захватывает синтез алкалоидов хозяина во время заражения морской диатомовой водорослью» . Природные коммуникации . 10 (1): 4938. Бибкод : 2019NatCo..10.4938V . doi : 10.1038/s41467-019-12908-w . ПМК 6821873 . ПМИД 31666506 .
- ^ Остерегайтесь миксотрофов — они могут уничтожить целые экосистемы «за считанные часы».
- ^ Микроскопические похитители тел заполонили наши океаны - Phys.org
- ^ Эйлер А. (декабрь 2006 г.). «Доказательства повсеместного распространения миксотрофных бактерий в верхних слоях океана: последствия и последствия» . Appl Environ Microbiol . 72 (12): 7431–7. Бибкод : 2006ApEnM..72.7431E . дои : 10.1128/АЕМ.01559-06 . ПМК 1694265 . ПМИД 17028233 .
- ^ Катечакис А., Стибор Х. (июль 2006 г.). «Миксотроф Ochromonas tuberculata может вторгаться и подавлять специализированные фаго- и фототрофные планктонные сообщества в зависимости от условий питания». Экология . 148 (4): 692–701. Бибкод : 2006Oecol.148..692K . дои : 10.1007/s00442-006-0413-4 . ПМИД 16568278 . S2CID 22837754 .
- ^ Шеманн, Вероника; Беквор, Сильви; Стефельс, Жаклин; Руссо, Вероника; Ланселот, Кристиана (1 января 2005 г.). «Цветение Phaeocystis в мировом океане и механизмы его контроля: обзор». Журнал морских исследований . Ресурсы железа и океанические питательные вещества – развитие глобального экологического моделирования. 53 (1–2): 43–66. Бибкод : 2005JSR....53...43S . CiteSeerX 10.1.1.319.9563 . дои : 10.1016/j.seares.2004.01.008 .
- ^ «Добро пожаловать на домашнюю страницу проекта секвенирования генома Phaeocystis antarctica» .
- ^ ДиТуллио, Греция; Гребмайер, Дж. М .; Арриго, КР; Лизотт, член парламента; Робинсон, Д.Х.; Левентер, А.; Барри, JP; ВанВорт, ML; Данбар, РБ (2000). «Быстрый и ранний экспорт цветения Phaeocystis antarctica в море Росса, Антарктида». Природа . 404 (6778): 595–598. Бибкод : 2000Natur.404..595D . дои : 10.1038/35007061 . ПМИД 10766240 . S2CID 4409009 .
- ^ Дж, Стефельс; Л, Дейхуизен; WWC, Гискес (20 июля 1995 г.). «Активность DMSP-лиазы в весеннем цветении фитопланктона у голландского побережья, связанная с численностью Phaeocystis sp.» (PDF) . Серия «Прогресс в области морской экологии» . 123 : 235–243. Бибкод : 1995MEPS..123..235S . дои : 10.3354/meps123235 .
- ^ Десель, Йохан; Симо, Рафель; Гали, Марти; Варгас, Коломбан де; Колен, Себастьян; Дедевиз, Ив; Биттнер, Люси; Проберт, Ян; Нет, Фабрис (30 октября 2012 г.). «Оригинальный способ симбиоза планктона открытого океана» . Труды Национальной академии наук . 109 (44): 18000–18005. Бибкод : 2012PNAS..10918000D . дои : 10.1073/pnas.1212303109 . ISSN 0027-8424 . ПМЦ 3497740 . ПМИД 23071304 .
- ^ Марс Брисбин, Маргарет; Гроссманн, Мэри М.; Месроп, Лиза Ю.; Митарай, Сатоши (2018). «Разнообразие симбионтов внутри хозяина и расширенное поддержание симбионтов в фотосимбиотических акантареях (клада F)» . Границы микробиологии . 9 : 1998. doi : 10.3389/fmicb.2018.01998 . ISSN 1664-302X . ПМК 6120437 . ПМИД 30210473 .
- ^ Jump up to: а б Стокер, ДК; Хансен, П.Дж.; Кэрон, Д.А.; Митра, А. (2017). «Миксотрофия морского планктона» . Ежегодный обзор морской науки . 9 : 311–335. Бибкод : 2017ARMS....9..311S . doi : 10.1146/annurev-marine-010816-060617 . ПМИД 27483121 .
- ^ Jump up to: а б Митра, А; Флинн, К.Дж.; Тильманн, Ю; Рэйвен, Дж; Кэрон, Д; и др. (2016). «Определение функциональных групп планктонных протистов по механизмам получения энергии и питательных веществ; включение разнообразных миксотрофных стратегий» . Протист . 167 (2): 106–20. дои : 10.1016/j.protis.2016.01.003 . hdl : 10261/131722 . ПМИД 26927496 .
- ^ Jump up to: а б Доусон, Скотт С; Паредес, Александр Р. (2013). «Альтернативные цитоскелетные ландшафты: новизна цитоскелета и эволюция простейших с базальными выемками» . Современное мнение в области клеточной биологии . 25 (1): 134–141. дои : 10.1016/j.ceb.2012.11.005 . ПМЦ 4927265 . ПМИД 23312067 .
- ^ Jump up to: а б Аткинсон, А.; Полимен, Л.; Файлман, Э.С.; Виддикомб, CE; МакЭвой, AJ; Смит, Ти Джей; Джегри, Н.; Сайли, Сан-Франциско; Корнуэлл, Луизиана (2018). « Комментарий. Что определяет сезонность планктона в стратифицированном шельфовом море? Некоторые конкурирующие и дополняющие друг друга теории"]" (PDF) . Лимнология и океанография . 63 (6): 2877–2884. Бибкод : 2018LimOc..63.2877A . дои : 10.1002/lno.11036 . S2CID 91380765 .
- ^ Синглтон, Пол (2006). Словарь по микробиологии и молекулярной биологии, 3-е издание, переработанное . Чичестер, Великобритания: John Wiley & Sons. стр. 32 . ISBN 978-0-470-03545-0 .
- ^ Дэвид Дж. Паттерсон. «Амебы: протисты, которые передвигаются и питаются с помощью псевдоподий» . Веб-проект «Древо жизни».
- ^ «Амебы» . Эдинбургский университет. Архивировано из оригинала 10 июня 2009 года.
- ^ Биологический словарь , 2004 г., по состоянию на 1 января 2011 г.
- ^ Паттерсон, Дэвид Дж. (2000) «Жгутиконосцы: гетеротрофные протисты со жгутиками» Древо жизни .
- ^ Лауга, Эрик; Томас Р. Пауэрс (25 августа 2009 г.). «Гидродинамика плавающих микроорганизмов». Отчеты о прогрессе в физике . 72 (9): 096601. arXiv : 0812.2887 . Бибкод : 2009RPPH...72i6601L . дои : 10.1088/0034-4885/72/9/096601 . S2CID 3932471 .
- ^ Гири, доктор медицинских наук (октябрь 2012 г.). «Сколько существует видов водорослей?». Журнал психологии . 48 (5): 1057–63. Бибкод : 2012JPcgy..48.1057G . дои : 10.1111/j.1529-8817.2012.01222.x . ПМИД 27011267 . S2CID 30911529 .
- ^ Jump up to: а б Гири, доктор медицины; Гири, генеральный менеджер (2016). «Альгебаза» . www.algaebase.org . Проверено 20 ноября 2016 г. .
- ^ Д. Томас (2002). Морские водоросли . Серия «Жизнь». Музей естественной истории , Лондон. ISBN 978-0-565-09175-0 .
- ^ Хук, Кристиан; ден Хёк, Хёк Ван; Чувак, Дэвид; Янс, Х.М. (1995). Водоросли: введение в психологию . Издательство Кембриджского университета. стр. 166. ИСБН 9780521316873 . OCLC 443576944 .
- ↑ Старкс, Сенне (31 октября 2012 г.) Место под солнцем: водоросли — это урожай будущего, по мнению исследователей из Geel. Архивировано 7 ноября 2017 г. в Wayback Machine Flanders Today, проверено 8 декабря 2012 г.
- ^ Дюваль, Б.; Маргулис, Л. (1995). «Микробное сообщество разносторонних колоний Ophrydium: эндосимбионты, резиденты и арендаторы». Симбиоз . 18 : 181–210. ПМИД 11539474 .
- ^ Мандоли, Д.Ф. (1998). «Разработка плана тела и фазового перехода во время развития вертлужной впадины: как строится сложная архитектура гигантской одноклеточной клетки?». Ежегодный обзор физиологии растений и молекулярной биологии растений . 49 : 173–198. дои : 10.1146/annurev.arplant.49.1.173 . ПМИД 15012232 . S2CID 6241264 .
- ^ Пьер Мадль; Марисела Йип (2004). «Обзор литературы Caulerpa Taxifolia» . БУФУС-Инфо . 19 (31). Архивировано из оригинала 8 октября 2022 года . Проверено 12 мая 2020 г.
- ^ Трегер, П.; Нельсон, DM; Ван Беннеком, AJ; Демастер, диджей; Лейнарт, А.; Кегинер, Б. (1995). «Баланс кремнезема в Мировом океане: переоценка». Наука . 268 (5209): 375–9. Бибкод : 1995Sci...268..375T . дои : 10.1126/science.268.5209.375 . ПМИД 17746543 . S2CID 5672525 .
- ^ Нельсон, Дэвид М.; Трегер, Поль; Бжезинский, Марк А.; Лейнарт, Од; Кегинер, Бернар (1995). «Производство и растворение биогенного кремнезема в океане: пересмотренные глобальные оценки, сравнение с региональными данными и связь с биогенным седиментированием». Глобальные биогеохимические циклы . 9 (3): 359–372. Бибкод : 1995GBioC...9..359N . дои : 10.1029/95GB01070 .
- ^ Мальвия, Шрути; Скалько, Элеонора; Аудик, Стефан; Винсент, Флора; Велучами, Алагурадж; Пулен, Жюли; Винкер, Патрик; Иудиконе, Даниэле; Де Варгас, Колумбан; Биттнер, Люси; Зингоне, Адриана; Боулер, Крис (2016). «Понимание глобального распределения и разнообразия диатомовых водорослей в мировом океане» . Труды Национальной академии наук . 113 (11): Е1516–Е1525. Бибкод : 2016PNAS..113E1516M . дои : 10.1073/pnas.1509523113 . ПМК 4801293 . ПМИД 26929361 . S2CID 22035749 .
- ^ Jump up to: а б Трегер, Поль; Боулер, Крис; Морисо, Бриваэла; Дуткевич, Стефани; Гелен, Мэрион; Омон, Оливье; Биттнер, Люси; Дагдейл, Ричард; Финкель, Зоя; Иудиконе, Даниэле; Ян, Оливер; Гуиди, Лайонел; Ласблеиз, морской пехотинец; Леблан, Карин; Леви, Марина; Пондавен, Филипп (2018). «Влияние разнообразия диатомей на биологический углеродный насос океана» (PDF) . Природа Геонауки . 11 (1): 27–37. Бибкод : 2018NatGe..11...27T . дои : 10.1038/s41561-017-0028-x . S2CID 134885922 .
- ^ Махадеван, Амала; д'Асаро, Эрик; Ли, Крейг; Перри, Мэри Джейн (2012). «Стратификация, управляемая вихрями, инициирует весеннее цветение фитопланктона в Северной Атлантике». Наука . 337 (6090): 54–58. Бибкод : 2012Наука...337...54М . дои : 10.1126/science.1218740 . ПМИД 22767922 . S2CID 42312402 .
- ^ Jump up to: а б с д Кавиччиоли, Рикардо; Риппл, Уильям Дж.; Тиммис, Кеннет Н.; Азам, Фарук; Баккен, Ларс Р.; Бэйлис, Мэтью; Беренфельд, Майкл Дж.; Боэций, Антье; Бойд, Филип В.; Классен, Эме Т.; Кроутер, Томас В.; Дановаро, Роберто; Форман Кристин М.; Хьюсман, Джефф; Хатчинс, Дэвид А.; Янссон, Джанет К.; Карл, Дэвид М.; Коскелла, Бритт; Марк Уэлч, Дэвид Б.; Мартини, Дженнифер Б.Х.; Моран, Мэри Энн; Сирота, Виктория Дж.; Рей, Дэвид С.; Ремайс, Джастин В.; Рич, Вирджиния И.; Сингх, Браджеш К.; Штейн, Лиза Ю.; Стюарт, Фрэнк Дж.; Салливан, Мэтью Б.; и др. (2019). «Предупреждение ученых человечеству: микроорганизмы и изменение климата» . Обзоры природы Микробиология . 17 (9): 569–586. дои : 10.1038/s41579-019-0222-5 . ПМЦ 7136171 . ПМИД 31213707 . Измененный текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Васильев, Мэгги (2006) «Планктон — растительный планктон» , Те Ара — Энциклопедия Новой Зеландии . Доступ: 2 ноября 2019 г.
- ^ «Королевский колледж Лондона — озеро Мегачад» . www.kcl.ac.uk. Проверено 5 мая 2018 г.
- ^ Бойд, Филип В.; Клаустр, Эрве; Леви, Марина; Сигел, Дэвид А.; Вебер, Томас (2019). «Многогранные насосы для частиц способствуют улавливанию углерода в океане» (PDF) . Природа . 568 (7752): 327–335. Бибкод : 2019Natur.568..327B . дои : 10.1038/s41586-019-1098-2 . ПМИД 30996317 . S2CID 119513489 .
- ^ Чжан, Д.; Ван, Ю.; Кай, Дж.; Пан, Дж.; Цзян, X.; Цзян, Ю. (2012). «Технология биопроизводства на основе диатомовой микро- и наноструктуры» . Китайский научный бюллетень . 57 (30): 3836–3849. Бибкод : 2012ЧСБу..57.3836З . дои : 10.1007/s11434-012-5410-x .
- ^ Беренфельд, Майкл Дж.; Дони, Скотт С.; Лима, Иван; Босс, Эммануэль С.; Сигел, Дэвид А. (2013). «Годовые циклы экологического нарушения и восстановления, лежащие в основе субарктического весеннего цветения планктона в Атлантике» . Глобальные биогеохимические циклы . 27 (2): 526–540. Бибкод : 2013GBioC..27..526B . дои : 10.1002/gbc.20050 . HDL : 1912/6250 .
- ^ Руссо, Сесиль С.; Грегг, Уотсон В. (2015). «Последние десятилетние тенденции в глобальном составе фитопланктона» . Глобальные биогеохимические циклы . 29 (10): 1674–1688. Бибкод : 2015GBioC..29.1674R . дои : 10.1002/2015GB005139 .
- ^ Арсеньев Л.; Саймон, Н.; Риго-Жалабер, Ф.; Ле Галль, Ф.; Шаффрон, С.; Корре, Э.; Приходить.; Бигерд, Э.; Боду, AC (2018). «Первые вирусы, заражающие морскую диатомовую водоросль Guinardia delicatula» . Границы микробиологии . 9 : 3235. дои : 10.3389/fmicb.2018.03235 . ПМК 6334475 . ПМИД 30687251 .
- ^ Килиас, Эстель С.; Юнгес, Леандро; Шупраха, Лука; Леонард, Гай; Метфис, Катя; Ричардс, Томас А. (2020). «Распространение хитридиевых грибов и их совместное присутствие с диатомовыми водорослями коррелируют с таянием морского льда в Северном Ледовитом океане» . Коммуникационная биология . 3 (1): 183. дои : 10.1038/s42003-020-0891-7 . ПМЦ 7174370 . ПМИД 32317738 . S2CID 216033140 . Измененный текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Рост, Б. и Рибеселл, У. (2004) «Кокколитофоры и биологический насос: реакция на изменения окружающей среды». В: Кокколитофоры: от молекулярных процессов к глобальному воздействию , страницы 99–125, Springer. ISBN 9783662062784 .
- ^ Jump up to: а б Васильев, Мэгги (2006) «Кокколитофор» , Те Ара — Энциклопедия Новой Зеландии . Доступ: 2 ноября 2019 г.
- ^ Янг, младший; Пративи, С.; Су, X. (2017). «Отчет о данных: отбор проб планктона поверхностной морской воды на наличие кокколитофоров, взятый во время 359-й экспедиции IODP» . Труды Международной программы открытия океана. дои : 10.14379/iodp.proc.359.111.2017 .
{{cite journal}}: Для цитирования журнала требуется|journal=( помощь ) - ^ Хагино К., Онума Р., Кавачи М. и Хоригучи Т. (2013) «Открытие эндосимбиотической азотфиксирующей цианобактерии UCYN-A у Braarudosphaera bigelowii (Prymnesiophyceae)». PLoS One , 8 (12): e81749. дои : 10.1371/journal.pone.0081749 .
- ^ Гомес Ф (2012). «Контрольный список и классификация живых динофлагеллят (Dinoflagellata, Alveolata)» . СИКИМАР Океаниды . 27 (1): 65–140. doi : 10.37543/oceanides.v27i1.111 .
- ^ Стокер Д.К. (1999). «Миксотрофия среди динофлагеллят». Журнал эукариотической микробиологии . 46 (4): 397–401. дои : 10.1111/j.1550-7408.1999.tb04619.x . S2CID 83885629 .
- ↑ Предлагаемое объяснение светящихся морей, включая Национальный научный фонд светящихся морей Калифорнии , 18 октября 2011 г.
- ^ Болтовской, Деметрио; Андерсон, О. Роджер; Корреа, Нэнси М. (2017). «Радиолярия и Феодария». Справочник протистов . Спрингер, Чам. стр. 731–763. дои : 10.1007/978-3-319-28149-0_19 . ISBN 9783319281476 .
- ^ Андерсон, Орегон (1983). Радиолярии . Springer Science & Business Media.
- ^ Гаст, Р.Дж.; Кэрон, Д.А. (1 ноября 1996 г.). «Молекулярная филогения симбиотических динофлагеллят планктонных фораминифер и радиолярий» . Молекулярная биология и эволюция . 13 (9): 1192–1197. doi : 10.1093/oxfordjournals.molbev.a025684 . ISSN 0737-4038 . ПМИД 8896371 .
- ^ Кастро П., Хубер М.Е. (2010). Морская биология (8-е изд.). МакГроу Хилл. стр. 95 . ISBN 978-0071113021 .
- ^ Гастингс Дж.В. (1996). «Химия и цвета биолюминесцентных реакций: обзор». Джин . 173 (1 номер спецификации): 5–11. дои : 10.1016/0378-1119(95)00676-1 . ПМИД 8707056 .
- ^ Хэддок С.Х., Молин М.А., Кейс Дж.Ф. (2009). «Биолюминесценция в море». Ежегодный обзор морской науки . 2 : 443–93. Бибкод : 2010ARMS....2..443H . doi : 10.1146/annurev-marine-120308-081028 . ПМИД 21141672 . S2CID 3872860 .
- ^ «Простейшие, поражающие жабры и кожу» . Ветеринарное руководство компании Merck . Архивировано из оригинала 3 марта 2016 года . Проверено 4 ноября 2019 г.
- ^ Брэнд, Ларри Э.; Кэмпбелл, Лиза; Бреснан, Эйлин (2012). « Карения : Биология и экология ядовитого рода» . Вредные водоросли . 14 : 156–178. Бибкод : 2012HAlga..14..156B . дои : 10.1016/j.hal.2011.10.020 . ПМЦ 9891709 . ПМИД 36733478 .
- ^ Баски, Э.Дж. (1995). «Рост и биолюминесценция Noctiluca scintillans на различных водорослевых диетах». Журнал исследований планктона . 17 (1): 29–40. дои : 10.1093/планкт/17.1.29 .
- ^ Панно, Джозеф (14 мая 2014 г.). Клетка: эволюция первого организма . Издательство информационной базы. ISBN 9780816067367 .
- ^ Бертран, Жан-Клод; Кометт, Пьер; Лебарон, Филипп; Мэтерон, Роберт; Норманд, Филипп; Симе-Нгандо, Телесфор (26 января 2015 г.). Экологическая микробиология: основы и приложения: Микробная экология . Спрингер. ISBN 9789401791182 .
- ^ Мэдиган, Майкл Т. (2012). Брок Биология микроорганизмов . Бенджамин Каммингс. ISBN 9780321649638 .
- ^ Йегер, Роберт Г. (1996). Простейшие: строение, классификация, рост и развитие . НКБИ. ISBN 9780963117212 . ПМИД 21413323 . Проверено 23 марта 2018 г.
- ^ Jump up to: а б Васильев, Мэгги (2006) «Планктон — животный планктон» , Те Ара — Энциклопедия Новой Зеландии . Доступ: 2 ноября 2019 г.
- ^ Варя, К.; Арагон, JL; Баррио, РА (1999). «Узоры Тьюринга на сфере». Физический обзор E . 60 (4): 4588–92. Бибкод : 1999PhRvE..60.4588V . дои : 10.1103/PhysRevE.60.4588 . ПМИД 11970318 .
- ^ Jump up to: а б Хемлебен, К.; Андерсон, Орегон; Шпиндлер, М. (1989). Современные планктонные фораминиферы . Спрингер-Верлаг. ISBN 978-3-540-96815-3 .
- ^ Фораминиферы: История изучения , Университетский колледж Лондона . Проверено: 18 ноября 2019 г.
- ^ Достижения в области микробной экологии, Том 11.
- ^ Бернхард, Дж. М.; Баузер, С.М. (1999). «Бентосные фораминиферы дизоксичных отложений: секвестрация хлоропластов и функциональная морфология». Обзоры наук о Земле . 46 (1): 149–165. Бибкод : 1999ESRv...46..149B . дои : 10.1016/S0012-8252(99)00017-3 .
- ^ Кальбет, Альберт; Лэндри, Майкл Р. (2004). «Рост фитопланктона, выпас микрозоопланктона и круговорот углерода в морских системах». Лимнология и океанография . 49 (1): 51–57. Бибкод : 2004LimOc..49...51C . дои : 10.4319/lo.2004.49.1.0051 . hdl : 10261/134985 . S2CID 22995996 .
- ^ Jump up to: а б с д Харагути, Луми; Якобсен, Ханс Х.; Лундхольм, Нина; Карстенсен, Джейкоб (2018). «Динамика сообщества фитопланктона: движущая сила трофических стратегий инфузорий» . Границы морской науки . 5 . дои : 10.3389/fmars.2018.00272 . S2CID 51925344 . Измененный текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Азам, Ф.; Фенчел, Т.; Филд, Дж. Г.; Грей, Дж. С.; Мейер-Рейл, Луизиана; Вингстад, Ф. (1983). «Экологическая роль микробов водной толщи моря» . Серия «Прогресс в области морской экологии» . 10 : 257–263. Бибкод : 1983MEPS...10..257A . дои : 10.3354/meps010257 .
- ^ Шерр, Эвелин; Шерр, Барри (1988). «Роль микробов в пелагических пищевых сетях: пересмотренная концепция» . Лимнология и океанография . 33 (5): 1225–1227. Бибкод : 1988LimOc..33.1225S . дои : 10.4319/lo.1988.33.5.1225 .
- ^ Фенчел, Т. (1988). «Пищевые цепи морского планктона». Ежегодный обзор экологии и систематики . 19 :19–38. doi : 10.1146/annurev.es.19.110188.000315 .
- ^ Буйтенхейс, Эрик; Ле Кере, Коринн; Омон, Оливье; Богранд, Грегори; Бункер, Адриан; Херст, Эндрю; Икеда, Цутому; О'Брайен, Тодд; Пионтковский, Сергей; Стрейл, Дитмар (2006). «Биогеохимические потоки через мезозоопланктон» . Глобальные биогеохимические циклы . 20 (2): н/д. Бибкод : 2006GBioC..20.2003B . дои : 10.1029/2005GB002511 . hdl : 2115/13694 .
- ^ Беренфельд, Майкл Дж.; Фальковски, Пол Г. (1997). «Скорость фотосинтеза определяется на основе концентрации хлорофилла, полученной со спутников» . Лимнология и океанография . 42 (1): 1–20. Бибкод : 1997LimOc..42....1B . дои : 10.4319/lo.1997.42.1.0001 . S2CID 15857675 .
- ^ Кальбет, Альберт (2001). «Влияние выпаса мезозоопланктона на первичную продукцию: глобальный сравнительный анализ морских экосистем». Лимнология и океанография . 46 (7): 1824–1830. Бибкод : 2001LimOc..46.1824C . дои : 10.4319/lo.2001.46.7.1824 . hdl : 10261/49263 . S2CID 85461746 .
- ^ Лэндри, Майкл Р.; Кальбет, Альберт (2004). «Продукция микрозоопланктона в Мировом океане» . Журнал морских наук ICES . 61 (4): 501–507. Бибкод : 2004ICJMS..61..501L . дои : 10.1016/j.icesjms.2004.03.011 .
- ^ Jump up to: а б Бютенхейс, Эрик Т.; Ривкин, Ричард Б.; Сайли, Севрин; Ле Кере, Корин (2010). «Биогеохимические потоки через микрозоопланктон» . Глобальные биогеохимические циклы . 24 (4): н/д. Бибкод : 2010GBioC..24.4015B . дои : 10.1029/2009GB003601 . S2CID 131413083 .
- ^ Хансен, Пер Жюэль; Бьёрнсен, Питер Кёфед; Хансен, Бенни Виндинг (2000). «Выпас и рост зоопланктона: масштабирование в диапазоне размеров тела 2–2000 мкм» . Лимнология и океанография . 45 (8): 1891. Бибкод : 2000LimOc..45.1891H . дои : 10.4319/lo.2000.45.8.1891 .
- ^ Нильсен, Торкель Гиссель; Кирбо, Томас (1994). «Регуляция биомассы и продукции зоопланктона в прибрежной экосистеме умеренного климата. 2. Инфузории» . Лимнология и океанография . 39 (3): 508–519. Бибкод : 1994LimOc..39..508N . дои : 10.4319/lo.1994.39.3.0508 .
- ^ Стокер, Дайан К.; Капуццо, Джудит Макдауэлл (1990). «Хищничество простейших: его значение для зоопланктона» . Журнал исследований планктона . 12 (5): 891–908. дои : 10.1093/планкт/12.5.891 .
- ^ Гиффорд, Дайан Дж. (1991). «Трофическая связь простейших и метазоев в пелагических экосистемах». Журнал протозоологии . 38 : 81–86. дои : 10.1111/j.1550-7408.1991.tb04806.x .
- ^ Флинн, Кевин Дж.; Стокер, Дайан К.; Митра, Адите; Рэйвен, Джон А.; Глиберт, Патрисия М .; Хансен, Пер Жюэль; Гранели, Эдна; Буркхолдер, Джоан М. (2013). «Неправильное использование дихотомии фитопланктона и зоопланктона: необходимость относить организмы к миксотрофам внутри функциональных типов планктона» . Журнал исследований планктона . 35 : 3–11. дои : 10.1093/plankt/fbs062 .
- ^ Эдвардс, Кайл Ф.; Томас, Мридул К.; Клаусмайер, Кристофер А.; Личман, Елена (2012). «Аллометрическое масштабирование и таксономические различия в характеристиках использования питательных веществ и максимальной скорости роста фитопланктона». Лимнология и океанография . 57 (2): 554–566. Бибкод : 2012LimOc..57..554E . дои : 10.4319/lo.2012.57.2.0554 . S2CID 13376583 .
- ^ Jump up to: а б Митра, А.; Флинн, К.Дж.; Беркхолдер, Дж. М.; Берге, Т.; Кальбет, А.; Рэйвен, Дж.А.; Гранели, Э.; Глиберт, премьер-министр; Хансен, П.Дж.; Стокер, ДК; Тингстад, Ф.; Тильманн, У.; Воге, С.; Уилкен, С.; Зубков, М.В. (2014). «Роль миксотрофных протистов в биологическом углеродном насосе» . Биогеонауки . 11 (4): 995–1005. Бибкод : 2014BGeo...11..995M . дои : 10.5194/bg-11-995-2014 . hdl : 10261/93693 .
- ^ Уорд, Бен А.; Далее следует Майкл Дж. (2016). «Морская миксотрофия увеличивает эффективность трофического переноса, средний размер организма и вертикальный поток углерода» . Труды Национальной академии наук . 113 (11): 2958–2963. Бибкод : 2016PNAS..113.2958W . дои : 10.1073/pnas.1517118113 . ПМК 4801304 . ПМИД 26831076 .
- ^ Лелес, С.Г.; Митра, А.; Флинн, К.Дж.; Стокер, ДК; Хансен, П.Дж.; Кальбет, А.; Макманус, Великобритания; Сандерс, RW; Кэрон, Д.А.; Нет, Ф.; Халлеграефф, генеральный менеджер; Питта, П.; Рэйвен, Дж.А.; Джонсон, доктор медицины; Глиберт, премьер-министр ; Воге, С. (2017). «Океанические протисты с различными формами приобретенной фототрофии демонстрируют контрастную биогеографию и численность» . Труды Королевского общества B: Биологические науки . 284 (1860). дои : 10.1098/rspb.2017.0664 . ПМЦ 5563798 . ПМИД 28768886 .
- ^ Мац, Михаил В.; Тамара М. Франк; Н. Джастин Маршалл; Эдит А. Виддер; Сонке Йонсен (9 декабря 2008 г.). «Гигантский глубоководный протист оставляет следы, похожие на двусторонние» (PDF) . Современная биология . 18 (23). Elsevier Ltd: 1849–1854 гг. Бибкод : 2008CBio...18.1849M . дои : 10.1016/j.cub.2008.10.028 . ПМИД 19026540 . S2CID 8819675 .
- ^ Добрый день, Эй Джей; Аранда да Силва, А.; Павловский, Дж. (1 декабря 2011 г.). «Ксенофиофоры (Rhizaria, Foraminifera) из каньона Назаре (окраина Португалии, северо-восток Атлантики)». Глубоководные исследования. Часть II: Актуальные исследования в океанографии . Геология, геохимия и биология подводных каньонов к западу от Португалии. 58 (23–24): 2401–2419. Бибкод : 2011DSRII..58.2401G . дои : 10.1016/j.dsr2.2011.04.005 .
- ^ Jump up to: а б с д и ж г час я дж к Бьорбекмо, Марит Ф. Маркуссен; Эвенстад, Андреас; Рёсег, Лайн Либлейн; Крабберёд, Андерс К.; Логарес, Рамиро (2020). «Интерактом планктонных протистов: где мы находимся после столетия исследований?» . Журнал ISME . 14 (2): 544–559. Бибкод : 2020ISMEJ..14..544B . дои : 10.1038/s41396-019-0542-5 . ПМЦ 6976576 . ПМИД 31685936 . Измененный текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Маргулис, Линн ; Фестер, Рене (1991). Симбиоз как источник эволюционных инноваций: видообразование и морфогенез . МТИ Пресс. ISBN 9780262132695 .
- ^ Лопес-Гарсия, Purificación; Эме, Лаура; Морейра, Дэвид (2017). «Симбиоз в эволюции эукариот» . Журнал теоретической биологии . 434 : 20–33. Бибкод : 2017JThBi.434...20L . дои : 10.1016/j.jtbi.2017.02.031 . ПМК 5638015 . ПМИД 28254477 .
- ^ Арчибальд, Джон М. (2015). «Эндосимбиоз и эволюция эукариотических клеток» . Современная биология . 25 (19): Р911–Р921. Бибкод : 2015CBio...25.R911A . дои : 10.1016/j.cub.2015.07.055 . ПМИД 26439354 . S2CID 16089231 .
- ^ Кавальер-Смит, Томас (2013). «Симбиогенез: механизмы, эволюционные последствия и систематические последствия». Ежегодный обзор экологии, эволюции и систематики . 44 : 145–172. doi : 10.1146/annurev-ecolsys-110411-160320 .
- ^ Маэ, Фредерик; Де Варгас, Колумбан; Басс, Дэвид; Чех, Лукас; Стаматакис, Александрос; Лара, Энрике; Певец, Дэвид; Мэр Иордании; Бунге, Джон; Сернакер, Сара; Сименсмейер, Тобиас; Траутманн, Изабель; Ромак, Сара; Берни, Седрик; Козлов, Алексей; Митчелл, Эдвард А.Д.; Сеппи, Кристоф Фольксваген; Эгге, Элианна; Лентенду, Гийом; Вирт, Райнер; Труба, Габриэль; Данторн, Мика (2017). «Паразиты доминируют в гиперразнообразных почвенных сообществах протистов в неотропических тропических лесах». Экология и эволюция природы . 1 (4): 91. Бибкод : 2017NatEE...1...91M . дои : 10.1038/s41559-017-0091 . ПМИД 28812652 . S2CID 2631960 .
- ^ Биар, Тристан; Стемманн, Ларс; Пишераль, Марк; Майо, Николя; Вандромм, Питер; Хаусс, Хелена; Горский, Габриэль; Гуиди, Лайонел; Кико, Райнер; Нет, Фабрис (2016). «Визуализация in situ показывает биомассу гигантских простейших в мировом океане» (PDF) . Природа . 532 (7600): 504–507. Бибкод : 2016Natur.532..504B . дои : 10.1038/nature17652 . ПМИД 27096373 . S2CID 205248710 .
- ^ Финли, Би Джей; Эстебан, Г.Ф. (1998). «Пресноводные простейшие: биоразнообразие и экологическая функция». Биоразнообразие и сохранение . 7 (9): 1163–1186. Бибкод : 1998BiCon...7.1163F . дои : 10.1023/А:1008879616066 . S2CID 10702795 .
- ^ Хаксли, Томас Х. (1851). «XXXIV. — Зоологические заметки и наблюдения, сделанные на борту HMS Rattlesnake» . Анналы и журнал естественной истории . 8 (48): 433–442. дои : 10.1080/03745486109495002 .
- ^ Брандт К. (1881) «О сосуществовании животных и водорослей». Верх Физиол Гесь , 1 : 524–527.
- ^ Логарес, Рамиро; Хаверкамп, Томас Х.А.; Кумар, Сурендра; Ланцен, Андерс; Недербрагт, Александр Дж.; Айва, Кристофер; Каузеруд, Ховард (2012). «Экологическая микробиология через призму высокопроизводительного секвенирования ДНК: обзор современных платформ и подходов биоинформатики». Журнал микробиологических методов . 91 (1): 106–113. дои : 10.1016/j.mimet.2012.07.017 . ПМИД 22849829 .
- ^ Согин, М.Л.; Моррисон, ХГ; Хубер, Дж.А.; Уэлч, DM; Хаус, С.М.; Нил, PR; Арриета, Дж. М.; Херндл, Г.Дж. (2006). «Микробное разнообразие в морских глубинах и малоизученной «редкой биосфере» » . Труды Национальной академии наук . 103 (32): 12115–12120. Бибкод : 2006PNAS..10312115S . дои : 10.1073/pnas.0605127103 . ПМЦ 1524930 . ПМИД 16880384 .
- ^ Гудвин, Сара; Макферсон, Джон Д.; МакКомби, В. Ричард (2016). «Взросление: десять лет технологий секвенирования нового поколения» . Обзоры природы Генетика . 17 (6): 333–351. дои : 10.1038/nrg.2016.49 . ПМЦ 10373632 . ПМИД 27184599 . S2CID 8295541 .
- ^ Педрос-Алио С., Ацинас С.Г., Логарес Р., Массана Р. Морское микробное разнообразие, наблюдаемое с помощью высокопроизводительного секвенирования. В: Газоль, Хосеп М.; Кирчман, Дэвид Л. (27 марта 2018 г.). Микробная экология океанов . Джон Уайли и сыновья. ISBN 9781119107187 . , стр. 47–87.
- ^ Спанг, Аня; Пила, Джимми Х.; Йоргенсен, Штеффен Л.; Заремба-Недзведска, Катажина; Мартейн, Йоран; Линд, Андерс Э.; Ван Эйк, Роэл; Шлепер, Криста; Гай, Лайонел; Эттема, Тийс Дж.Г. (2015). «Сложные археи, заполняющие пропасть между прокариотами и эукариотами» . Природа . 521 (7551): 173–179. Бибкод : 2015Natur.521..173S . дои : 10.1038/nature14447 . ПМЦ 4444528 . ПМИД 25945739 .
- ^ Фауст, Каролина; Лахти, Лео; Гонзе, Дидье; Де Вос, Виллем М.; Раес, Йерун (2015). «Метагеномика сочетается с анализом временных рядов: разгадка динамики микробного сообщества» . Современное мнение в микробиологии . 25 : 56–66. дои : 10.1016/j.mib.2015.04.004 . ПМИД 26005845 .
- ^ Фауст, Каролина; Раес, Йерун (2012). «Микробные взаимодействия: от сетей к моделям». Обзоры природы Микробиология . 10 (8): 538–550. дои : 10.1038/nrmicro2832 . ПМИД 22796884 . S2CID 22872711 .
- ^ Лима-Мендес, Г.; и др. (2015). «Детерминанты структуры сообщества в глобальном интерактоме планктона». Наука . 348 (6237). дои : 10.1126/science.1262073 . hdl : 10261/117702 . ПМИД 25999517 . S2CID 10326640 .
- ^ Лайегифард, Мехди; Хван, Дэвид М.; Гуттман, Дэвид С. (2017). «Распутывание взаимодействий в микробиоме: сетевая перспектива» . Тенденции в микробиологии . 25 (3): 217–228. дои : 10.1016/j.tim.2016.11.008 . ПМЦ 7172547 . ПМИД 27916383 .
- ^ Адл, Сина М.; и др. (2019). «Пересмотр классификации, номенклатуры и разнообразия эукариот» . Журнал эукариотической микробиологии . 66 (1): 4–119. дои : 10.1111/jeu.12691 . ПМК 6492006 . ПМИД 30257078 .
- ^ Шульц, Фредерик; Элоэ-Фадрош, Эмили А.; Бауэрс, Роберт М.; Джаретт, Джессика; Нильсен, Торбен; Иванова Наталья Н.; Кирпидес, Никос К.; Войке, Таня (2017). «На пути к сбалансированному взгляду на бактериальное древо жизни» . Микробиом . 5 (1): 140. дои : 10.1186/s40168-017-0360-9 . ПМЦ 5644168 . ПМИД 29041958 .
- ^ «Группы протистов | Безграничная биология» . Courses.lumenlearning.com . Проверено 16 февраля 2021 г.
- ^ Jump up to: а б Агирре Л.Е., Оуян Л., Эльфвинг А., Хедблом М., Вульф А. и Инганес О. (2018) «Панцири диатомовых водорослей защищают ДНК от ультрафиолетового света». Научные отчеты , 8 (1): 1–6. два : 10.1038/s41598-018-21810-2 . Измененный текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Де Томмаси, Э., Конгестри, Р., Дардано, П., Де Лука, А.С., Манаго, С., Ри, И. и Де Стефано, М. (2018) «УФ-защита и преобразование длины волны с помощью центрических диатомей панцири с наноузорами». Научные отчеты , 8 (1): 1–14. doi : 10.1038/s41598-018-34651-w . Измененный текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Биоразлагаемый блеск и микроводоросли, пожирающие загрязнения: новые материалы, вдохновленные природой Horizon , 28 мая 2020 г.
- ^ Гафар, Н.А., Эйр, Б.Д. и Шульц, К.Г. (2019) «Сравнение видовой чувствительности к изменению света и химии карбонатов при кальцинировании морского фитопланктона». Научные отчеты , 9 (1): 1–12. два : 10.1038/s41598-019-38661-0 . Измененный текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Jump up to: а б с д Монтейро Ф.М., Бах Л.Т., Браунли К., Баун П., Рикаби Р.Э., Поултон А.Дж., Тиррелл Т., Бофорт Л., Дуткевич С., Гиббс С. и Гутовска М.А. ( 2016) «Почему кальцинируется морской фитопланктон». Science Advances , 2 (7): e1501822. два : 10.1126/sciadv.1501822 . Измененный текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
Дальнейшие ссылки
[ редактировать ]- Бьорбекмо, Марит Ф. Маркуссен; Эвенстад, Андреас; Рёсег, Лайн Либлейн; Крабберёд, Андерс К.; Логарес, Рамиро (2020). «Интерактом планктонных протистов: где мы находимся после столетия исследований?» . Журнал ISME . 14 (2): 544–559. Бибкод : 2020ISMEJ..14..544B . дои : 10.1038/s41396-019-0542-5 . ПМЦ 6976576 . ПМИД 31685936 . Доступно по международной лицензии Creative Commons Attribution 4.0 .
- Флинн, Кевин Дж.; Митра, Адите; Анестис, Константинос; Аншюц, Анна А.; Кальбет, Альберт; Феррейра, Гильерме Дуарте; Гипенс, Натали; Хансен, Пер Дж.; Джон, Уве; Мартин, Джон Лапейра; Мансур, Йост С.; Маселли, Майра; Медич, Никола; Норлин, Андреас; Нет, Фабрис; Питта, Параскеви; Романо, Филомена; Саиз, Энрик; Шнайдер, Лиза К.; Столте, Виллем; Трабони, Клаудия (2019). «Миксотрофные протисты и новая парадигма морской экологии: куда сейчас идут исследования планктона?» . Журнал исследований планктона . 41 (4): 375–391. дои : 10.1093/plankt/fbz026 . hdl : 10261/192145 .
- Лелеш, Сузана Гонсалвес; Полимен, Лука; Брюггеман, Йорн; Блэкфорд, Джереми; Чаватта, Стефано; Митра, Адите; Флинн, Кевин Джон (2018). «Моделирование миксотрофного функционального разнообразия и последствий для функционирования экосистемы». Журнал исследований планктона . 40 (6): 627–642. дои : 10.1093/plankt/fby044 .
- Килинг, Патрик Дж.; Кампо, Хавьер дель (2017). «Морские протисты — это не просто большие бактерии» . Современная биология . 27 (11). Эльзевир Б.В.: R541–R549. Бибкод : 2017CBio...27.R541K . дои : 10.1016/j.cub.2017.03.075 . ISSN 0960-9822 . ПМИД 28586691 . S2CID 207052528 .
