Фотогетеротроф
Фотогетеротрофы ( греч . фото = свет, гетеро = (ан)другой, троф = питание) — это гетеротрофные фототрофы , то есть это организмы, которые используют свет для получения энергии, но не могут использовать углекислый газ в качестве единственного источника углерода. Следовательно, они используют органические соединения из окружающей среды для удовлетворения своих потребностей в углероде; эти соединения включают углеводы , жирные кислоты и спирты . Примеры фотогетеротрофных организмов включают пурпурные несерные бактерии , зеленые несерные бактерии и гелиобактерии . [ 1 ] Эти микроорганизмы повсеместно распространены в водной среде обитания, занимают уникальные ниши и вносят вклад в глобальный биогеохимический круговорот. Недавние исследования также показали, что восточный шершень и некоторые тли могут использовать свет для пополнения своего запаса энергии. [ 2 ]
Исследовать
[ редактировать ]Исследования показали, что митохондрии млекопитающих также могут улавливать свет и синтезировать АТФ при смешивании с феофорбидом , светоулавливающим метаболитом хлорофилла. [ 3 ] Исследования показали, что тот же метаболит, скармливаемый червю Caenorhabditis elegans, приводит к увеличению синтеза АТФ при воздействии света, а также к увеличению продолжительности жизни. [ 4 ]
Более того, эксперименты по инокуляции показывают, что миксотрофные Ochromonas danica (т.е. золотые водоросли) – и сравнимые с ними эукариоты – предпочитают фотогетеротрофию в олиготрофных (т.е. с ограниченным содержанием питательных веществ) водных средах обитания. [ 5 ] Это предпочтение может повысить эффективность использования энергии и экономический рост за счет сокращения инвестиций в фиксацию неорганического углерода (например, производство автотрофного оборудования, такого как RuBisCo и PSII).
Метаболизм
[ редактировать ]Фотогетеротрофы генерируют АТФ с помощью света одним из двух способов: [ 6 ] [ 7 ] они используют реакционный центр на основе бактериохлорофилла или бактериородопсин . Механизм , основанный на хлорофилле, аналогичен тому, который используется в фотосинтезе, когда свет возбуждает молекулы в реакционном центре и вызывает поток электронов через цепь переноса электронов (ETS). Этот поток электронов через белки заставляет ионы водорода перекачиваться через мембрану. Энергия, запасенная в этом протонном градиенте, используется для синтеза АТФ . В отличие от фотоавтотрофов , электроны движутся только по циклическому пути: электроны, высвобождаемые из реакционного центра, проходят через ЭТС и возвращаются в реакционный центр. Они не используются для восстановления каких-либо органических соединений. Пурпурные несерные бактерии , зеленые несерные бактерии и гелиобактерии являются примерами бактерий, осуществляющих данную схему фотогетеротрофии.
Другие организмы, включая галобактерии и флавобактерии. [ 8 ] и вибрионы [ 9 ] на основе пурпурного родопсина имеют протонные насосы , которые дополняют их запасы энергии. Архейная протеородопсин версия называется бактериородопсин , а эубактериальная называется . версия Насос состоит из одного белка, связанного с производным витамина А — ретиналем . Насос может иметь дополнительные пигменты (например, каротиноиды ), связанные с белком. Когда свет поглощается молекулой сетчатки, молекула изомеризуется. Это заставляет белок менять форму и перекачивать протон через мембрану. Градиент ионов водорода затем можно использовать для генерации АТФ, транспорта растворенных веществ через мембрану или управления жгутиковым мотором . Одна конкретная флавобактерия не может восстанавливать углекислый газ с помощью света, но использует энергию своей системы родопсина для фиксации углекислого газа посредством анаплеротической фиксации. [ 8 ] Флавобактерия по-прежнему является гетеротрофом , поскольку для жизни ей необходимы восстановленные соединения углерода, и они не могут существовать только за счет света и CO 2 . Он не может осуществлять реакции в виде
- n CO 2 + 2n H 2 D + фотоны → (CH 2 O) n + 2n D + n H 2 O,
где H 2 D может быть водой, H 2 S или другим соединением/соединениями, обеспечивающими восстановление электронов и протонов; пара 2D + H 2 O представляет собой окисленную форму.
Однако он может фиксировать углерод в таких реакциях, как:
где малат или другие полезные молекулы получают иным образом путем расщепления других соединений путем
- углеводы + О 2 → малат + СО 2 + энергия.
Этот метод фиксации углерода полезен, когда восстановленных соединений углерода мало и они не могут быть потрачены в виде CO 2 во время взаимопревращений, но энергии много в виде солнечного света.
Экология
[ редактировать ]
Распределение и разделение ниш
[ редактировать ]Фотогетеротрофы - либо 1) цианобактерии (т.е. факультативные гетеротрофы в средах с ограниченным количеством питательных веществ, такие как Synechococcus и Prochromococcus), 2) аэробные аноксигенные фотогетеротрофные бактерии (AAP; с использованием реакционных центров на основе бактериохлорофилла), 3) протеородопсин (PR)-содержащие бактерии и археи, и 4) гелиобактерии (т.е. единственный фототроф с пигменты бактериохлорофилла g , или грамположительные мембраны), встречаются в различных водных средах обитания, включая океаны , стратифицированные озера , рисовые поля и окружающей среды экстремальные условия . [ 10 ] [ 11 ] [ 12 ] [ 13 ]
В фотозонах океанов до 10% бактериальных клеток способны к AAP, тогда как более 50% чистых морских микроорганизмов содержат PR, достигая до 90% в прибрежных биомах. [ 14 ] Как было показано в экспериментах по инокуляции, фотогетеротрофия может обеспечить этим планктонным микробам конкурентные преимущества 1) по сравнению с хемогетеротрофами в олиготрофных (т.е. бедных питательными веществами) средах за счет повышения эффективности использования питательных веществ (т.е. чрезмерного биосинтеза органического углерода по сравнению с производством энергии) и 2) за счет исключения инвестиций в физиологически дорогостоящие автотрофные ферменты/комплексы (РуБисКо и ФСII). [ 15 ] [ 16 ] Кроме того, в Северном Ледовитом океане фотогетеротрофы AAP и PR преобладают в покрытых льдом регионах в зимнее время из-за нехватки света. [ 17 ] Наконец, у морских AAP наблюдался сезонный оборот, поскольку экотипы (т.е. генетически сходные таксоны с разными функциональными признаками и/или экологическими предпочтениями) разделяются во временные ниши. [ 18 ]
В стратифицированных (т. е. эвксиновых) озерах фотогетеротрофы — наряду с другими аноксигенными фототрофами (например, пурпурными/зелеными серными бактериями, фиксирующими углекислый газ с помощью доноров электронов, таких как двухвалентное железо, сульфид и газообразный водород) — часто занимают хемоклин в толще воды и /или отложения. [ 19 ] В этой зоне снижается содержание растворенного кислорода, свет ограничивается длинными волнами (например, красным и инфракрасным), оставшимися от оксигенных фототрофов (например, цианобактерий), и начинается анаэробный метаболизм (т. е. происходящий в отсутствие кислорода). сульфиды и биодоступные питательные вещества (например, органический углерод, фосфат и аммиак) посредством восходящей диффузии. [ 20 ]
Гелиобактерии являются облигатными анаэробами, главным образом обитающими на рисовых полях , где низкие концентрации сульфидов предотвращают конкурентное вытеснение пурпурных/зеленых серобактерий. [ 21 ] Эта заболоченная среда может способствовать симбиотическим отношениям между гелиобактериями и растениями риса, поскольку фиксированный азот из первых обменивается на богатые углеродом корневые выделения.
Наблюдительные исследования охарактеризовали фотогетеротрофы (например, зеленые несерные бактерии, такие как Chloroflexi и AAP) в фотосинтетических матах в экстремальных условиях окружающей среды (например, в горячих источниках и гиперсоленых лагунах). [ 12 ] [ 22 ] Примечательно, что температура и pH определяют состав сообщества аноксигенных фототрофов в Йеллоустонского национального парка . геотермальных объектах [ 12 ] Кроме того, различные светозависимые ниши в гиперсоленых матах озера Грейт-Солк поддерживают фототрофное разнообразие, поскольку микробы оптимизируют производство энергии и борются с осмотическим стрессом. [ 22 ]
Биогеохимический циклизм
[ редактировать ]Фотогетеротрофы влияют на глобальный круговорот углерода, ассимилируя растворенный органический углерод (DOC). [ 23 ] [ 20 ] Следовательно, при сборе световой энергии углерод удерживается в микробном контуре без соответствующего дыхания (т.е. выброс углекислого газа в атмосферу, поскольку DOC окисляется для производства энергии). Это несоответствие, открытие факультативных фотогетеротрофов (например, AAP с гибкими источниками энергии) и предыдущие измерения, проведенные в темноте (т. е. во избежание искажения значений потребления кислорода из-за фотоокисления , УФ-света и кислородного фотосинтеза), приводят к завышенной оценке водного CO. 2 выброса. Например, снижение дыхания на 15,2% наблюдалось в озере Кеп, Чехия, наряду с преимущественным поглощением глюкозы и пирувата, что объясняется факультативными фотогетеротрофами, предпочитающими световую энергию в дневное время, учитывая упомянутые ранее преимущества фитнеса. [ 23 ]
Блок-схема
[ редактировать ]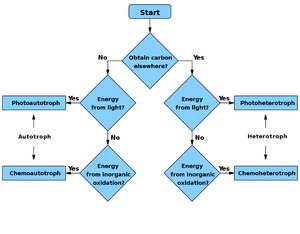
Источник энергии Источник углерода |
Хемотроф | Фототроф |
|---|---|---|
| автотроф | Хемоавтотроф | Фотоавтотроф |
| Гетеротроф | Хемогетеротроф | Фотогетеротроф |
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Брайант, округ Колумбия; Фригаард, Н.-У. (ноябрь 2006 г.). «Освещение фотосинтеза и фототрофии прокариот». Тенденции Микробиол . 14 (11): 488–496. дои : 10.1016/j.tim.2006.09.001 . ПМИД 16997562 .
- ^ Вальмалетт, JC; Домбровский А.; Брат, П.; Мерц, К.; Каповилья, М.; Робишон, А. (2012). «Светоиндуцированный перенос электронов и синтез АТФ у насекомого, синтезирующего каротин» . Научные отчеты . 2 : 579. Бибкод : 2012НатСР...2Э.579В . дои : 10.1038/srep00579 . ПМЦ 3420219 . ПМИД 22900140 .
- ^ Сюй, Чен; Чжан, Цзюньхуа; Михай, Дойна М.; Вашингтон, Ильяс (15 января 2014 г.). «Светособирающие пигменты хлорофилла позволяют митохондриям млекопитающих улавливать фотонную энергию и производить АТФ» . Журнал клеточной науки . 127 (2): 388–399. дои : 10.1242/jcs.134262 . ISSN 0021-9533 . ПМК 6518289 . ПМИД 24198392 .
- ^ Чжан, Дэн; Робинсон, Кира; Михай, Дойна М.; Вашингтон, Ильяс (12 октября 2016 г.). «Секвестрация повсеместно распространенных пигментов пищевого происхождения позволяет митохондриальному светочувствлению» . Научные отчеты . 6 (1): 34320. Бибкод : 2016NatSR...634320Z . дои : 10.1038/srep34320 . ISSN 2045-2322 . ПМК 5059631 . ПМИД 27731322 .
- ^ Уилкен, Сюзанна; Шуурманс, Дж. Мерейн; Маттейс, Ганс КП (декабрь 2014 г.). «Вырастают ли миксотрофы как фотогетеротрофы? Фотофизиологическая акклиматизация хризофита Ochromonas danica после питания» . Новый фитолог . 204 (4): 882–889. дои : 10.1111/nph.12975 . ISSN 0028-646X . ПМИД 25138174 .
- ^ Брайант, Дональд А.; Фригаард, Нильс-Ульрик (ноябрь 2006 г.). «Освещение фотосинтеза и фототрофии прокариот». Тенденции в микробиологии . 14 (11): 488–496. дои : 10.1016/j.tim.2006.09.001 . ISSN 0966-842X . ПМИД 16997562 .
- ^ Зубков, Михаил В. (1 сентября 2009 г.). «Фотогетеротрофия морских прокариот» . Журнал исследований планктона . 31 (9): 933–938. дои : 10.1093/plankt/fbp043 . ISSN 0142-7873 .
- ^ Jump up to: а б Гонсалес, Хосе М.; Фернандес-Гомес, Беатрис; Фернандес-Герра, Энтони; Гомес-Консарнау, Лаура; Санчес, Ольга; Коль-Льядо, Монтсеррат; и др. (24 июня 2008 г.). «Анализ генома протеородопсин-содержащей морской бактерии Polaribacter Sp. MED152 (Flavobacteria)» . Труды Национальной академии наук . 105 (25): 8724–8729. дои : 10.1073/pnas.0712027105 . ISSN 0027-8424 . ПМЦ 2438413 . ПМИД 18552178 .
- ^ Гомес-Консарнау, Лаура; Акрам, Нилам; Линделл, Кристоффер; Педерсен, Андерс; Нойце, Ричард; Милтон, Дебра Л.; Гонсалес, Хосе М.; Пинхасси, Хароне (2010). «Фототрофия протеородопсина способствует выживанию морских бактерий во время голодания» . ПЛОС Биол . 8 (4): e1000358. дои : 10.1371/journal.pbio.1000358 . ПМЦ 2860489 . ПМИД 20436956 .
- ^ Зубков, М.В. (18 июня 2009 г.). «Фотогетеротрофия морских прокариот» . Журнал исследований планктона . 31 (9): 933–938. дои : 10.1093/plankt/fbp043 . ISSN 0142-7873 .
- ^ Джаганнатан, Б.; Гольбек, Дж. Х. (01 января 2009 г.), «Фотосинтез: микробы» , в Шехтере, Моселио (редактор), Энциклопедия микробиологии (третье издание) , Оксфорд: Academic Press, стр. 325–341, ISBN 978-0-12-373944-5 , получено 15 апреля 2023 г.
- ^ Jump up to: а б с Гамильтон, Тринити Л.; Беннетт, Аннастейша К.; Муругапиран, Сентил К.; Хэвиг, Джефф Р. (17 декабря 2019 г.). Флинн, Теодор М. (ред.). «Аноксигенные фототрофы охватывают геохимические градиенты и разнообразную морфологию в наземных геотермальных источниках» . mSystems . 4 (6): e00498–19. дои : 10.1128/mSystems.00498-19 . ISSN 2379-5077 . ПМК 6832021 . ПМИД 31690593 .
- ^ Бёф, Д.; Смиренно, Ф.; Жантон, К. (20 июня 2014 г.). «Разнообразие арктических пелагических <i>Бактерий</i> с акцентом на фотогетеротрофов: обзор» . Биогеонауки . 11 (12): 3309–3322. Бибкод : 2014BGeo...11.3309B . дои : 10.5194/bg-11-3309-2014 . ISSN 1726-4189 .
- ^ Арандия-Горостиди, Нестор; Гонсалес, Хосе М.; Хуэте-Штауффер, Тамара М.; Ансари, Мохд И.; Моран, Хосе Анкселу Г.; Алонсо-Саес, Лаура (2 августа 2020 г.). «Свет поддерживает целостность клеток и скорость роста таксономически разнообразных прибрежных фотогетеротрофов» . Экологическая микробиология . 22 (9): 3823–3837. Бибкод : 2020EnvMi..22.3823A . дои : 10.1111/1462-2920.15158 . hdl : 10754/664249 . ISSN 1462-2912 . ПМИД 32643243 . S2CID 220435352 .
- ^ Уилкен, Сюзанна; Шуурманс, Дж. Мерейн; Маттейс, Ганс КП (19 августа 2014 г.). «Вырастают ли миксотрофы как фотогетеротрофы? Фотофизиологическая акклиматизация хризофита Ochromonas danica после питания» . Новый фитолог . 204 (4): 882–889. дои : 10.1111/nph.12975 . ISSN 0028-646X . ПМИД 25138174 .
- ^ Фенчел, Т.; Кинг, генеральный менеджер; Блэкберн, TH (2012), «Биогеохимический круговорот в почвах» , Бактериальная биогеохимия , Elsevier, стр. 89–120, doi : 10.1016/b978-0-12-415836-8.00006-2 , ISBN 9780124158368 , получено 15 апреля 2023 г.
- ^ Коттрелл, Мэтью Т.; Кирчман, Дэвид Л. (2009). «Фотогетеротрофные микробы в Северном Ледовитом океане летом и зимой» . Прикладная и экологическая микробиология . 75 (15): 4958–4966. Бибкод : 2009ApEnM..75.4958C . дои : 10.1128/aem.00117-09 . ISSN 0099-2240 . ПМЦ 2725502 . ПМИД 19502441 .
- ^ Оладель, Адриа; Санчес, Пабло; Санчес, Ольга; Газоль, Джозеф М.; Феррера, Изабель (2019). «Многолетняя сезонная и межгодовая изменчивость морских аэробных аноксигенных фотогетеротрофных бактерий» . Журнал ISME . 13 (8): 1975–1987. Бибкод : 2019ISMEJ..13.1975A . дои : 10.1038/s41396-019-0401-4 . ISSN 1751-7370 . ПМК 6776013 . ПМИД 30914777 . S2CID 256555570 .
- ^ Пост, Северная Каролина; Бристоу, Луизиана; Кокс, Р.П.; Хабихт, Канзас; Данза, Ф.; Тонолла, М.; Фригаард, Нью-Йорк; Кэнфилд, Делавэр (2017). «Фракционирование изотопов углерода аноксигенными фототрофными бактериями в эвксиновом озере Каданьо» . Геобиология . 15 (6): 798–816. Бибкод : 2017Gbio...15..798P . дои : 10.1111/gbi.12254 . ISSN 1472-4669 . ПМИД 28866873 . S2CID 11132075 .
- ^ Jump up to: а б Федонкин, М.А. (декабрь 2009 г.). «Эукариотизация ранней биосферы: биогеохимический аспект» . Геохимия Интернэшнл . 47 (13): 1265–1333. Бибкод : 2009GeocI..47.1265F . дои : 10.1134/s0016702909130011 . ISSN 0016-7029 . S2CID 98430905 .
- ^ Мэдиган, Майкл Т.; Ормерод, Джон Г. (2004), Таксономия, физиология и экология гелиобактерий , Достижения в области фотосинтеза и дыхания, том. 2, Дордрехт: Kluwer Academic Publishers, стр. 17–30, doi : 10.1007/0-306-47954-0_2 , ISBN. 0-7923-3681-Х , получено 16 апреля 2023 г.
- ^ Jump up to: а б Каник, Мерт; Манро-Эрлих, Мейсон; Фернандес-Мартинс, Мария Клара; Пейн, Девон; Джанулиас, Кэтрин; Келлер, Лиза; Кубацкий, Александр; Линдси, Мелоди Р.; Бакстер, Бонни К.; Ванден Берг, Майкл Д.; Колман, Дэниел Р.; Бойд, Эрик С. (05 мая 2020 г.). «Неожиданная численность и разнообразие фототрофов в матах из морфологически изменчивых микробиалитов в Большом Соленом озере, штат Юта» . Прикладная и экологическая микробиология . 86 (10): e00165–20. Бибкод : 2020ApEnM..86E.165K . дои : 10.1128/AEM.00165-20 . ISSN 1098-5336 . ПМЦ 7205499 . ПМИД 32198176 .
- ^ Jump up to: а б Пивос, Кася; Виллена-Алемани, Кристиан; Муякич, Изабела (2022). «Фотогетеротрофия аэробных аноксигенных бактерий модулирует потоки углерода в пресноводном озере» . Журнал ISME . 16 (4): 1046–1054. Бибкод : 2022ISMEJ..16.1046P . дои : 10.1038/s41396-021-01142-2 . ISSN 1751-7370 . ПМЦ 8941148 . ПМИД 34802055 .
Источники
[ редактировать ]«Микробиология онлайн» (учебник). Университет Висконсина, Мэдисон.