Принцип конкурентного исключения

2: Более крупный (красный) вид конкурирует за ресурсы.
3: Красный цвет доминирует в середине, поскольку ресурсов больше. Желтый адаптируется к новой нише, ограниченной верхами и низами и избегающей конкуренции .
В экологии действует принцип конкурентного исключения . [1] иногда называемый законом Гаузе , [2] Это предположение, что два вида за , конкурирующие один и тот же ограниченный ресурс, не могут сосуществовать при постоянной численности популяции. Когда один вид имеет хоть малейшее преимущество перед другим, тот, у кого это преимущество, будет доминировать в долгосрочной перспективе. Это ведет либо к вымиранию более слабого конкурента, либо к эволюционному или поведенческому сдвигу в сторону другой экологической ниши . Этот принцип был перефразирован в поговорке: «Полные конкуренты не могут сосуществовать». [1]
История [ править ]
Принцип конкурентного исключения классически приписывают Георгию Гаузе . [3] хотя на самом деле он никогда этого не формулировал. [1] Этот принцип уже присутствует в теории естественного отбора Дарвина. [2] [4]
На протяжении всей своей истории статус этого принципа колебался между априорным («два сосуществующих вида должны иметь разные ниши») и экспериментальной истиной («мы обнаруживаем, что сосуществующие виды действительно имеют разные ниши»). [2]
Экспериментальная база [ править ]

Основываясь на полевых наблюдениях, Джозеф Гриннелл в 1904 году сформулировал принцип конкурентного исключения: «Два вида примерно с одинаковыми пищевыми привычками вряд ли будут долго сохранять равномерную численность в одном и том же регионе. Один вытеснит другой». [5] Георгий Гаузе сформулировал закон конкурентного исключения на основе лабораторных конкурентных экспериментов с использованием двух видов Paramecium , P. aurelia и P. caudatum . Условия заключались в том, чтобы добавлять свежую воду каждый день и вводить постоянный поток пищи. Хотя первоначально P. caudatum доминировал, P. aurelia восстановился и впоследствии привел к вымиранию P. caudatum из-за конкуренции за ресурсы . Однако Гаузе смог позволить P. caudatum выжить, изменив параметры окружающей среды (пища, вода). Таким образом, закон Гаузе справедлив только в том случае, если экологические факторы постоянны.
Прогноз [ править ]
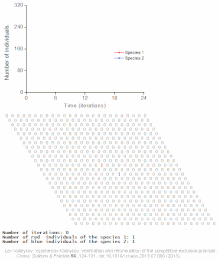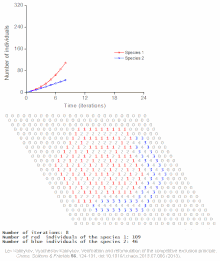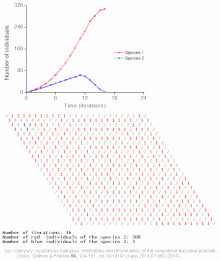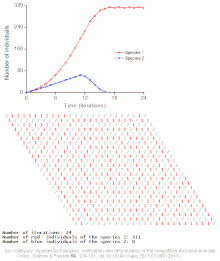
Конкурентное исключение предсказывается математическими и теоретическими моделями, такими как Лотки-Вольтерра модели конкуренции . Однако по плохо понятным причинам конкурентное исключение редко наблюдается в природных экосистемах , и многие биологические сообщества, похоже, нарушают закон Гаузе. Самый известный пример — так называемый « парадокс планктона ». [6] Все виды планктона живут за счет очень ограниченного количества ресурсов, в первую очередь солнечной энергии и минералов, растворенных в воде. Согласно принципу конкурентного исключения, лишь небольшое количество видов планктона должно иметь возможность сосуществовать на этих ресурсах. Тем не менее, большое количество видов планктона сосуществуют в небольших районах открытого моря.
Некоторые сообщества, которые, похоже, поддерживают принцип конкурентного исключения, являются Макартура . славками [7] и дарвиновы зяблики , [8] хотя последние по-прежнему очень сильно перекрываются с экологической точки зрения, а конкуренция оказывает на них негативное влияние только в экстремальных условиях. [9]
Парадоксальные черты [ править ]
Частичное решение парадокса заключается в повышении размерности системы. Пространственная неоднородность , трофические взаимодействия, за многочисленные ресурсы конкуренция , компромисс между конкуренцией и колонизацией и отставание могут предотвратить исключение (игнорируя стохастическое вымирание в более длительных периодах времени). Однако такие системы, как правило, аналитически трудноразрешимы. Кроме того, многие теоретически могут поддерживать неограниченное количество видов. Возникает новый парадокс: большинство известных моделей, допускающих стабильное сосуществование, допускают сосуществование неограниченного числа видов, однако в природе любое сообщество содержит лишь несколько видов.
Переопределение [ править ]
Недавние исследования, посвященные некоторым предположениям, сделанным для моделей прогнозирования конкурентного исключения, показали, что эти предположения необходимо пересмотреть. Например, небольшая модификация предположения о том, как связаны рост и размер тела, приводит к другому выводу, а именно, что в данной экосистеме определенный диапазон видов может сосуществовать, в то время как другие становятся вытесняемыми. [10] [11]
Одним из основных способов сосуществования видов, разделяющих ниши, является компромисс между конкуренцией и колонизацией . Другими словами, виды, которые являются лучшими конкурентами, будут специалистами, тогда как виды, которые являются лучшими колонизаторами, с большей вероятностью будут универсалами. Модели «хозяин-паразит» являются эффективным способом изучения этих отношений с использованием событий передачи хозяина. Кажется, есть два места, где способность к колонизации у экологически близких видов различается. В перьевых вшах Буш и Клейтон [12] представили некоторое подтверждение этого, показав, что два близкородственных рода вшей почти равны по своей способности колонизировать новых голубей-хозяев после переселения. Харбисон [13] продолжили эту линию размышлений, исследуя, различаются ли эти два рода по способности к передаче. Это исследование было сосредоточено в первую очередь на определении того, как происходит колонизация и почему крылатые вши являются лучшими колонизаторами, чем платяные вши. Вертикальный перенос является наиболее распространенным явлением между родителем и потомком, он хорошо изучен и хорошо понятен. Горизонтальный перенос трудно измерить, но у вшей он, по-видимому, происходит посредством фореза или «автостопа» одного вида на другой. Харбисон обнаружил, что платяные вши менее искусны в форезе и превосходят их в конкурентной борьбе, тогда как крылатые вши превосходны в колонизации.
Поддержка модели компромисса конкуренции и колонизации также обнаружена у мелких млекопитающих, связанных с пожарами. В проекте, посвященном долгосрочным последствиям Йеллоустонских пожаров 1988 года, Аллен и др. [14] использовали стабильные изотопы и данные повторного захвата пространственных меток, чтобы показать, что южные красные полевки ( Clethrionomys Gapperi ) ), специалист, исключают мышей-оленей ( Peromyscus maniculatus ) , универсалов, из пищевых ресурсов в старовозрастных лесах. Однако после лесных пожаров мыши-олени становятся более эффективными колонизаторами и способны воспользоваться преимуществами освобождения от конкурентного давления со стороны полевок. Эта динамика устанавливает закономерность экологической преемственности в этих экосистемах, при этом конкурентное исключение полевок определяет количество и качество ресурсов, к которым могут получить доступ мыши-олени.
контекст Филогенетический
Экологическое сообщество — это совокупность видов , поддерживаемая экологическими методами (Хатчинсон, 1959; [15] Лейболд, 1988 г. [16] ) и эволюционный процесс (Weiher and Keddy, 1995; [17] Чейз и др ., 2003). Эти два процесса играют важную роль в формировании существующего сообщества и будут продолжаться и в будущем (Tofts et al ., 2000; Ackerly, 2003; Reich et al ., 2003). В местном сообществе потенциальные члены фильтруются сначала по факторам окружающей среды, таким как температура или наличие необходимых ресурсов, а затем по способности сосуществовать с другими местными видами.
В подходе к пониманию того, как два вида приспосабливаются друг к другу в сообществе или как целое сообщество приживается вместе, в «Происхождении видов» ( Дарвин , 1859) предположено, что в однородных условиях окружающей среды борьба за существование между близкородственными видами сильнее, чем между отдаленнородственными видами. Он также предположил, что функциональные черты могут сохраняться в разных филогениях. Столь сильное филогенетическое сходство между близкородственными видами известно как филогенетические эффекты (Derrickson et al ., 1988. [18] )
С помощью полевых исследований и математических моделей экологи установили связь между сходством функциональных признаков между видами и его влиянием на сосуществование видов. Согласно гипотезе конкурентного родства (Cahil et al ., 2008). [19] ) или гипотеза филогенетического предельного сходства (Violle et al ., 2011 [20] ) межвидовая конкуренция [21] высок среди видов, имеющих схожие функциональные характеристики и конкурирующих за схожие ресурсы и среду обитания. Следовательно, это вызывает сокращение числа близкородственных видов и их равномерное распространение, известное как филогенетическая сверхдисперсия (Webb et al ., 2002). [22] ). Обратной стороной филогенетической сверхдисперсии является филогенетическая кластеризация, в этом случае ожидается, что виды с консервативными функциональными признаками будут встречаться одновременно из-за фильтрации окружающей среды (Weiher et al ., 1995; Webb, 2000). В исследовании, проведенном Уэббом и др ., 2000, они показали, что на небольших участках леса Борнео встречаются близкородственные деревья. Это говорит о том, что близкородственные виды имеют общие черты, которым благоприятствуют конкретные факторы окружающей среды, которые различаются на разных участках, вызывая филогенетическую кластеризацию.
Для обеих филогенетических моделей (филогенетическая сверхдисперсия и филогенетическая кластеризация) базовым предположением является то, что филогенетически родственные виды также экологически схожи (H. Burns et al., 2011). [23] ). Значительного количества экспериментов, позволяющих ответить на вопрос, насколько близкородственные виды схожи и по нише, нет. В связи с этим обе филогенетические закономерности нелегко интерпретировать. Было показано, что филогенетическая сверхдисперсия может быть также результатом конвергенции отдаленно родственных видов (Cavender-Bares et al. 2004; [24] Крафт и др. 2007 год [25] ). В своем исследовании [ нужна ссылка ] , они показали, что черты скорее конвергентны, чем сохраняются. Хотя в другом исследовании [ нужна ссылка ] Было показано, что филогенетическая кластеризация также может быть связана с историческими или биогеографическими факторами, которые не позволяют видам покинуть свои предковые ареалы. Таким образом, для понимания силы взаимодействия видов в собрании сообщества необходимы дополнительные филогенетические эксперименты.
Применение к людям [ править ]
Доказательства того, что принцип конкурентного исключения действует в человеческих группах, были рассмотрены и интегрированы в теорию королевства для объяснения воинственных и мирных обществ. [26] Например, группы охотников-собирателей, окруженные другими группами охотников-собирателей в той же экологической нише, будут воевать, по крайней мере, время от времени, в то время как группы охотников-собирателей, окруженные группами с разными средствами существования, могут мирно сосуществовать. [26]
Еще одно недавнее применение: в своей работе «Историческая динамика» Питер Турчин разработал так называемую теорию метаэтнических границ , согласно которой как расцвет, так и возможное падение империй происходит из-за географически и/или политически конфликтующих популяций. [27] Соответственно, приграничные регионы, в которых действует принцип конкурентного исключения, должны быть ключевыми в этногенезе человека . Подводя итог более широким прогнозам в одном:
Асабия — это концепция из сочинений Ибн Халдуна, которую Турчин определяет как «способность общества к коллективным действиям». Теория метаэтнических границ призвана включить асабию в качестве ключевого фактора в предсказании динамики имперских аграрных обществ – того, как они растут, сокращаются и зарождаются. Турчин утверждает, что многоуровневый отбор может помочь нам выявить динамику асабии в группах. Далее он отмечает три аспекта, в которых логика многоуровневого отбора может иметь значение для понимания изменений в «коллективной солидарности»: межгрупповой конфликт, демографические и ресурсные ограничения и этнические границы.В небольших группах межгрупповой конфликт может усилить асабию, поскольку людям необходимо объединиться, чтобы выжить как группа. И наоборот (опять же для небольших групп), большая численность населения по отношению к доступным ресурсам может уменьшить асабию, поскольку отдельные люди конкурируют за ограниченные ресурсы. Что касается более крупных групп, Турчин предполагает, что этнические границы могут влиять на то, как группы небольших групп с умеренными этническими различиями могут объединяться против людей, которые еще более «этнически дистанцированы» - более «Другие». В этом процессе небольших групп, объединяющихся против народов, более отличных от них самих, они могут сформировать то, что Турчин называет Метаэтнический фронтир … Турчин отмечает, что динамика этнических границ, которая порождает асабию в большой группе (состоящей из меньших групп), слаба, потому что по мере увеличения размера группы центральные регионы менее подвержены межгрупповым конфликтам, и асабия уменьшается, что приводит к к большему внутреннему разделению. Наконец, Турчин отмечает, что все три вышеупомянутые возможности возникают в регионах, составляющих имперские и метаэтнические границы (имперские и метаэтнические границы часто совпадают, отмечает он). Именно в этих регионах интенсивной динамики рождается асабия, наиболее склонная к этногенезу. [28]
См. также [ править ]
Ссылки [ править ]
- ^ Jump up to: Перейти обратно: а б с Гаррет Хардин (1960). «Принцип конкурентного исключения» (PDF) . Наука . 131 (3409): 1292–1297. Бибкод : 1960Sci...131.1292H . дои : 10.1126/science.131.3409.1292 . ПМИД 14399717 . Архивировано из оригинала (PDF) 17 ноября 2017 г. Проверено 24 ноября 2016 г.
- ^ Jump up to: Перейти обратно: а б с Пошевиль, Арно (2015). «Экологическая ниша: история и недавние противоречия» . В Химсе, Томас; Хьюнеман, Филипп; Лекуантр, Гийом; и др. (ред.). Справочник по эволюционному мышлению в науке . Дордрехт: Спрингер. стр. 547–586. ISBN 978-94-017-9014-7 .
- ^ Гаузе, Георгий Францевич (1934). Борьба за существование (1-е изд.). Балтимор: Уильямс и Уилкинс. Архивировано из оригинала 28 ноября 2016 г. Проверено 24 ноября 2016 г.
- ^ Дарвин, Чарльз (1859). О происхождении видов путем естественного отбора, или о сохранении избранных рас в борьбе за жизнь (1-е изд.). Лондон: Джон Мюррей. ISBN 1-4353-9386-4 .
- ^ Гриннелл, Дж. (1904). «Происхождение и распространение каштановой гаички» . Аук . 21 (3). Союз американских орнитологов: 364–382. дои : 10.2307/4070199 . JSTOR 4070199 .
- ^ Хатчинсон, Джордж Эвелин (1961). «Парадокс планктона». Американский натуралист . 95 (882): 137–145. дои : 10.1086/282171 . S2CID 86353285 .
- ^ Макартур, Р.Х. (1958). «Популяционная экология некоторых славок северо-восточных хвойных лесов». Экология . 39 (4): 599–619. Бибкод : 1958Экол...39..599М . дои : 10.2307/1931600 . JSTOR 1931600 . S2CID 45585254 .
- ^ Лак, Д.Л. (1945). «Галапагосские зяблики (Geospizinae); исследование вариаций». Периодические статьи Калифорнийской академии наук . 21 : 36–49.
- ^ Де Леон, LF; Подос, Дж; Гардези, Т; Херрел, А; Хендри, AP (июнь 2014 г.). «Дарвиновы зяблики и их диетические ниши: симпатрическое сосуществование несовершенных универсалов» . Дж Эвол Биол . 27 (6): 1093–104. дои : 10.1111/jeb.12383 . ПМИД 24750315 .
- ^ Растеттер, Е.Б.; Огрен, Г.И. (2002). «Изменения в индивидуальной аллометрии могут привести к сосуществованию без разделения ниш». Экосистемы . 5 : 789–801. дои : 10.1007/s10021-002-0188-3 . S2CID 30089349 .
- ^ Молл, Джей Ди; Браун, Дж. С. (2008). «Конкуренция и сосуществование на нескольких этапах жизненной истории». Американский натуралист . 171 (6): 839–843. дои : 10.1086/587517 . ПМИД 18462131 . S2CID 26151311 .
- ^ Клейтон, Д.Х.; Буш, ЮВ (2006). «Роль размера тела в специфичности хозяина: эксперименты по взаимному переносу перьевых вшей». Эволюция . 60 (10): 2158–2167. дои : 10.1111/j.0014-3820.2006.tb01853.x . ПМИД 17133872 . S2CID 221734637 .
- ^ Харбисон, CW (2008). «Сравнительная динамика передачи конкурирующих видов паразитов». Экология . 89 (11): 3186–3194. Бибкод : 2008Ecol...89.3186H . дои : 10.1890/07-1745.1 . ПМИД 31766819 .
- ^ Аллен, AG; Рёрс, З.П.; Севилья, РС; Ланье, ХК (2022). «Конкурентный выпуск во время пожарной сукцессии влияет на экологический оборот в сообществе мелких млекопитающих» . Экология . 103 (8): 1–12. Бибкод : 2022Ecol..103E3733A . дои : 10.1002/ecy.3733 . ПМЦ 9891167 . ПМИД 35430726 .
- ^ Хатчинсон, GE (1959). «Посвящение Санта-Розалии или почему существует так много видов животных?». Американский натуралист . 93 (870): 145–159. дои : 10.1086/282070 . ISSN 0003-0147 . JSTOR 2458768 . S2CID 26401739 .
- ^ Лейболд, Мэтью А. (1 января 1998 г.). «Сходство и локальное сосуществование видов в региональных биотах». Эволюционная экология . 12 (1): 95–110. Бибкод : 1998EvEco..12...95L . дои : 10.1023/A:1006511124428 . ISSN 1573-8477 . S2CID 6678357 .
- ^ Вейхер, Эван; Кедди, Пол А. (1995). «Ассамблея экспериментальных сообществ водно-болотных растений». Ойкос . 73 (3): 323–335. Бибкод : 1995Oikos..73..323W . дои : 10.2307/3545956 . ISSN 0030-1299 . JSTOR 3545956 .
- ^ Дерриксон, Э.М.; Риклефс, RE (1988). «Таксон-зависимая диверсификация особенностей жизненного цикла и восприятие филогенетических ограничений». Функциональная экология . 2 (3): 417–423. Бибкод : 1988FuEco...2..417D . дои : 10.2307/2389415 . ISSN 0269-8463 . JSTOR 2389415 .
- ^ Кэхилл, Джеймс Ф.; Кембель, Стивен В.; Лэмб, Эрик Г.; Кедди, Пол А. (12 марта 2008 г.). «Влияет ли филогенетическое родство на силу конкуренции среди сосудистых растений?». Перспективы экологии, эволюции и систематики растений . 10 (1): 41–50. Бибкод : 2008PPEES..10...41C . дои : 10.1016/j.ppees.2007.10.001 . ISSN 1433-8319 .
- ^ Виолле, Сирилл; Немергут, Диана Р.; Пу, Чжичао; Цзян, Линь (2011). «Филогенетическое ограничение сходства и конкурентное исключение». Экологические письма . 14 (8): 782–787. Бибкод : 2011EcolL..14..782V . дои : 10.1111/j.1461-0248.2011.01644.x . ISSN 1461-0248 . ПМИД 21672121 .
- ^ Тархуэло, Р.; Моралес, МБ; Арройо, Б.; Маньоса, С.; Бота, Г.; Касас, Ф.; Траба, Дж. (2017). «Внутривидовая и межвидовая конкуренция вызывает сдвиги ниш среды обитания у находящихся под угрозой исчезновения степных птиц в зависимости от плотности» . Экология и эволюция . 7 (22): 9720–9730. Бибкод : 2017EcoEv...7.9720T . дои : 10.1002/ece3.3444 . ПМК 5696386 . ПМИД 29188003 .
- ^ Уэбб, Кэмпбелл О.; Акерли, Дэвид Д.; МакПик, Марк А.; Донохью, Майкл Дж. (2002). «Филогения и экология сообществ». Ежегодный обзор экологии и систематики . 33 (1): 475–505. doi : 10.1146/annurev.ecolsys.33.010802.150448 . S2CID 535590 .
- ^ Бернс, Джин Х.; Штраус, Шэрон Ю. (29 марта 2011 г.). «Более близкие виды более экологически схожи в экспериментальном тесте» . Труды Национальной академии наук . 108 (13): 5302–5307. Бибкод : 2011PNAS..108.5302B . дои : 10.1073/pnas.1013003108 . ISSN 0027-8424 . ПМК 3069184 . ПМИД 21402914 .
- ^ Кавендер-Барес, Дж .; Акерли, Д.Д.; Баум, Д.А.; Баззаз, ФА (июнь 2004 г.). «Филогенетическое перерасселение дубовых сообществ Флориды». Американский натуралист . 163 (6): 823–843. дои : 10.1086/386375 . ISSN 1537-5323 . ПМИД 15266381 . S2CID 2959918 .
- ^ Крафт, Натан Дж.Б.; Корнуэлл, Уильям К.; Уэбб, Кэмпбелл О.; Акерли, Дэвид Д. (август 2007 г.). «Эволюция признаков, сборка сообществ и филогенетическая структура экологических сообществ». Американский натуралист . 170 (2): 271–283. дои : 10.1086/519400 . ISSN 1537-5323 . ПМИД 17874377 . S2CID 7222026 .
- ^ Jump up to: Перейти обратно: а б Туман, Агнер (2017). Воинственные и мирные общества: взаимодействие генов и культуры . Открытое книжное издательство. дои : 10.11647/ОБП.0128 . ISBN 978-1-78374-403-9 .
- ^ Турчин, Петр (2019). Историческая динамика: почему государства растут и падают . Нью-Джерси: Издательство Принстонского университета. стр. 50–77.
- ^ Байм, Мартин (3 декабря 2019 г.). «Реализация теории метаэтнических границ Турчина с помощью NetLogo» . Проверено 21 февраля 2024 г.
