Островной гигантизм

Островной гигантизм , или островной гигантизм , — это биологическое явление, при котором размер вида животных, изолированного на острове, резко увеличивается по сравнению с его материковыми сородичами. Островной гигантизм — это один из аспектов более общего «островного эффекта» или «правила Фостера» , которое утверждает, что, когда материковые животные колонизируют острова, мелкие виды имеют тенденцию развивать более крупные тела, а крупные виды имеют тенденцию развивать меньшие тела ( островная карликовость ). Это само по себе является одним из аспектов более общего явления островного синдрома , который описывает различия в морфологии , экологии , физиологии и поведении островных видов по сравнению с их континентальными аналогами. После прибытия людей и связанных с ними интродуцированных хищников (собак, кошек, крыс, свиней) многие гигантские, а также другие островные эндемики вымерли ( например, додо и пасьянс Родригес , гигантские нелетающие голуби, родственные никобарскому голубю ). Аналогичное увеличение размера, а также повышенная древесность наблюдались у некоторых островных растений, таких как дерево Мапу ( Cyphostemma Mappia ) на Маврикии, которая также известна как «маврикийский баобаб», хотя принадлежит к семейству виноградных ( Vitaceae ).
Возможные причины
[ редактировать ]
Крупные хищные млекопитающие часто отсутствуют на островах из-за недостаточного ареала или трудностей с расселением над водой . В их отсутствие экологические ниши крупных хищников могут быть заняты птицами, рептилиями или более мелкими хищниками, которые затем могут вырасти до размеров, превышающих нормальные. Например, на доисторическом острове Гаргано в миоцен - плиоценовом Средиземноморье , на островах Карибского моря , таких как Куба , а также на Мадагаскаре и Новой Зеландии , некоторые или все высшие хищники были такими птицами, как орлы , соколы и совы , включая некоторые из самых крупных известных экземпляров. этих групп. Однако птицы и рептилии обычно являются менее эффективными крупными хищниками, чем продвинутые хищники .
Поскольку небольшой размер обычно позволяет травоядным убегать или прятаться от хищников, снижение давления хищников на островах может позволить им вырасти больше. [1] [а] Мелкие травоядные также могут выиграть от отсутствия конкуренции со стороны отсутствующих видов крупных травоядных.
Преимущества большого размера, которые были предложены для островных черепах, включают снижение уязвимости к нехватке еды и / или воды за счет способности выживать без них в течение более длительных периодов времени или способности преодолевать большие расстояния, чтобы добыть их. Периоды такого дефицита могут представлять большую угрозу на океанических островах, чем на материке. [4]
Таким образом, островной гигантизм обычно представляет собой эволюционную тенденцию, возникающую в результате устранения ограничений на размер мелких животных, связанных с хищничеством и/или конкуренцией. [5] Однако такие ограничения могут действовать по-разному в зависимости от размера животного; например, в то время как мелкие травоядные могут избежать хищников, спрятавшись, крупные травоядные могут отпугивать хищников, запугивая их. В результате дополнительный феномен островной карликовости может также возникнуть в результате устранения ограничений, связанных с хищничеством и/или конкуренцией за размер крупных травоядных животных. [6] Напротив, островная карликовость среди хищников чаще всего является результатом наложения ограничений, связанных с ограниченными ресурсами добычи, доступными на островах. [6] В отличие от островной карликовости, островной гигантизм встречается у большинства основных групп позвоночных и у беспозвоночных .
Территориализм может способствовать развитию островного гигантизма. Исследование на острове Анахо в Неваде показало, что территориальные виды рептилий, как правило, крупнее на острове по сравнению с материком, особенно более мелкие виды. У территориальных видов больший размер делает особей более способными конкурировать за защиту своей территории. Это дает дополнительный импульс эволюции в сторону увеличения размера островной популяции. [7]
Еще одним средством установления островного гигантизма может быть эффект основателя , действующий, когда более крупные представители материкового населения превосходят свои способности колонизировать острова. [8]
Размер острова играет роль в определении степени гигантизма. Острова меньшего размера обычно ускоряют скорость эволюции изменений в размерах организмов, и организмы на них развиваются до более крайних размеров. [9]
Примеры
[ редактировать ]Примеры островного гигантизма включают:
Млекопитающие
[ редактировать ]Многие грызуны на островах вырастают крупнее, тогда как хищные , хоботные и парнокопытные обычно мельчают.
| Пример | Биномиальное имя | Родной диапазон | Текущий статус | Континентальный родственник |
|---|---|---|---|---|
| Балеарская гигантская землеройка | Незиотиты идальго | Майорка и Менорка | Вымершие (3000-2000 до н.э.) |  Краснозубая землеройка |
| Сардинская гигантская землеройка | как задница | Сардиния и Корсика | Вымершие ( голоцен ) | |
| Сицилийская гигантская землеройка | Asoriculus burgioi | Сицилия | Вымершие (ранний плейстоцен) | |
 Деиногалерикс | Дейногалерикс виды. | Остров Гаргано | Вымершие ( поздний миоцен ) |  Лунные крысы |
Грызуны
[ редактировать ]| Пример | Биномиальное имя | Родной диапазон | Текущий статус | Континентальный родственник | Островной / материковый соотношение длины или массы |
|---|---|---|---|---|---|
| Тупозубая гигантская хутия | Затопленная амблириза | Ангилья и Сен-Мартен | Вымершие ( плейстоцен ) |  Неотропические колючие крысы | |
| Большая ямайская гигантская хутия | Клидомис осборни | Ямайка | Вымершие (поздний плейстоцен) | ||
| Пластинчатозубая гигантская хутия | Косая эласмодонтомия | Пуэрто-Рико | Вымершие (ок. 1 г. н. э.) | ||
| Мышь с искривленными зубами | Тяжелая анемия | Эспаньола | Вымерший | ||
| Древесная гигантская хутия [10] | Тайнотерий Валей | Пуэрто-Рико | Вымерший | ||
| Гигантская хутия Малой Ямайки | Ксаймака fulvopulvis | Ямайка | Вымерший | ||
| Майоркинские гигантские хомяки | Апокритус дардери Трагомис макфей | Майорка | Вымерший | Апокритус Альберти [11] Крицетус коморси [12] | |
 Гигантский хомяк Гаргано | Хаттомис Гаргантюа | Остров Гаргано | Вымерший | ||
 Полевая мышь Сент-Килда | Apodemus sylvaticus hirtensis | Сент-Килда | Наименьшее беспокойство |  Деревянная мышь | МР ≈ 2 [13] |
 Полевая мышь Хенселя | Предварительные сведения об ортодонтии | Корсика и Сардиния | Вымершие (после 1300 г. до н.э.) | ||
 Тенерифская гигантская крыса | Канариомис бравой | Тенерифе | Вымершие (поздний плейстоцен) | Африканские рыженосые крысы | |
 Гран-Канарская гигантская крыса | Канариомис тамарани | Гран-Канария | Вымершие (до 1500 г. н. э.) | ||
| Форментеровская чернохвостая садовая соня | Элиомис quercinus ophiusae | Форментера | Редкий (представлен людьми) [14] |  Садовая соня и другие Leithiinae сони | |
 Балеарская гигантская соня | Hypnomys spp. | Майорка и Менорка | Вымершие (голоцен) | ||
 | Лейтия из книги | Сицилия и Мальта | Вымерший | ||
| Лейтия мелитенсис | |||||
 Оркнейская полевка | Microtus arvalis из фруктового сада | Оркнейские острова | Уязвимый |  Обыкновенная полевка и другие луговые полевки | |
 Гигантские полевки Гаргано | Микротия большая капитал М. М. Парва | Остров Гаргано | Вымершие (ранний плиоцен) | ||
 Домовая мышь Сент-Килда | Мышечная стеночная мышца | Сент-Килда | Вымершие (ок. 1930 г. н.э.) |  Домовая мышь | |
Гигантская крыса Флорес | Папагомис арманвиллей | Флорес | Рядом с угрозой |  Североафриканская черная крыса и другие настоящие крысы | |
| Сулавесская гигантская крыса | Паруромис доминатор | Сулавеси | Наименьшее беспокойство | ||
| Адмиралтейская гигантская крыса | Крыса была задержана | Остров Манус | Неизвестно/вероятно находится под угрозой [15] | ||
| Популяция черных крыс Конгресса [16] | Раттус Раттус | Остров Конгресса | Наименьшее беспокойство | ||
| Оленьи мыши с Нормандских островов | Peromyscus Anyapahensis П. незодиты | Северные Нормандские острова Калифорнии | Вымершие (ок. 6000 г. до н.э.) |  Североамериканская оленьая мышь | |
 Гигантская соня Гаргано | Stertomys laticrestatus [17] | Остров Гаргано | Вымерший |  Глириновые сони |
| Пример | Биномиальное имя | Родной диапазон | Текущий статус | Континентальный родственник |
|---|---|---|---|---|
 Миноркский гигантский зайцеобразный | Нуралагус рекс | Менорка | Вымершие ( средний плиоцен ) | Субподряд (?) Тришизолаг (?) |
| Пролагус империалис | Остров Гаргано | Вымерший |  другой | |
 Сардинская пищуха | Пролагус Сардинский | Корсика , Сардиния и Таволара | Вымершие (ок. 1800 г. н.э.) |
Приматы
[ редактировать ]| Пример | Биномиальное имя | Родной диапазон | Текущий статус | Континентальный родственник |
|---|---|---|---|---|
| Обезьяна Эспаньола | Антиллотрикс берненсис | Эспаньола | Вымершие (до 1600 г. н. э.) |  Керацебус |
| Гаитянская обезьяна | Инсулацебус туссентиана | Юго-Западный Гаити | Вымерший | |
 Кубинские обезьяны | Мариана Паралуатта [18] П. пауки [18] | Куба | Вымершие (плейстоцен) | |
| Ямайская обезьяна | Ксенотрикс Макгрегори | Ямайка | Вымерший | |
 Горилла лемур | Археоиндрис фонтойнонтии | Центральный Мадагаскар | Вымершие (ок. 350 г. до н. э.) |  Лоризоиды |
 Бабуины-лемуры | Виды Archaeolemur . Виды Hadropithecus . | Мадагаскар | Вымершие (до 1280 г. н. э.) | |
 Ленивцы лемуры | Бабакотия виды. Палеопропитек виды. | Западный и Центральный Мадагаскар | Вымершие (ок. 1500 г. н. э.) | |
 Коала лемуры | Мегаладапис Эдвардси М. грандидьери М. мадагаскарский | Мадагаскар | Вымершие (1280–1420 гг. н. э.) |
Хищники
[ редактировать ]| Пример | Биномиальное имя | Родной диапазон | Текущий статус | Континентальный родственник |
|---|---|---|---|---|
 Сардинская гигантская выдра | Мегаленгидрис барбарицина | Сардиния | Вымершие (поздний плейстоцен) |  Выдры |
 Фосса | Криптопрокта Ферокс | Мадагаскар | Уязвимый |  Мангусты |
 Гигантская яма | Криптопрокта спелея | Мадагаскар | Вымершие (до 1400 г. н. э.) |
Птицы
[ редактировать ]Стволовые птицы
[ редактировать ]| Пример | Биномиальное имя | Родной диапазон | Текущий статус | Континентальный родственник |
|---|---|---|---|---|
 Дракон | Б. Бондок | Остров Хатег | Вымершие (поздний мел) |  Джехолорнис [19] |
 Гаргантуавис | Г. филохинос | Иберо-Армориканский остров | Вымершие ( поздний мел ) |  Патагоптерикс (?) |
Ратиты
[ редактировать ]| Пример | Биномиальное имя | Родной диапазон | Текущий статус | Континентальный родственник |
|---|---|---|---|---|
 Киви | Аптеригиды | Новая Зеландия | Переменная | Проаптерикс [б] |
 Большие слоновые птицы | Эпиорнитиды | Мадагаскар | Вымершие (ок. 1700 г. н. э.) | |
 Малые слоновьи птицы | Мюллерорнитиды | Мадагаскар | Вымершие (ок. 1260 г. н. э.) | |
 Гигантские моа | Динорнитиды | Новая Зеландия | Вымершие (ок. 1450 г. н. э.) |  Тинамус |
 Меньше моа | Эмеиды | Новая Зеландия | Вымершие (ок. 1460 г. н. э.) | |
 Нагорное болото | Мегалаптеригиды | Новая Зеландия | Вымершие (ок. 1300 г. н. э.) |
| Пример | Биномиальное имя | Родной диапазон | Текущий статус | Континентальный родственник |
|---|---|---|---|---|
| Новозеландская мускусная утка | Biziura delautouri | Новая Зеландия | Вымершие (после 1500 г. н. э.) |  Австралийская мускусная утка |
 Новозеландские гуси | Кнемиорнис кальцитранс C. gracilis | Новая Зеландия | Вымерший |  Мыс Баррен гусь |
 Гарганорни | Г. Баллмани | Гаргано и Сконтроне Острова | Вымершие ( поздний миоцен ) |  Гуси [21] |
 Черепашья челюсть больше-хуже | Chelychelynechen quassus | Кауаи | Вымершие (ок. 1000 г. н. э.) |  Балующиеся утки |
 Малоклювый моа-нало | Птайохен пау | Мауи | Вымершие (ок. 1000 г. н. э.) | |
Крупноклювая курица потерялась | Тамбеточен хаулиодус | Великий Мауи | Вымершие (ок. 1000 г. н. э.) | |
 Заправка для курицы-неудачника | Тамбетохен ксанион | Одежда | Вымершие (ок. 1000 г. н. э.) | |
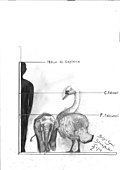 Гигантский лебедь | Лебедь фальконери | Сицилия и Мальта | Вымершие ( средний плейстоцен ) |  Лебедь-шипун |
| утка Скарлетт | Малакоринхус Скарлетти | Новая Зеландия | Вымершие (после 1500 г. н. э.) |  Розовоухая утка |
| Пример | Биномиальное имя | Родной диапазон | Текущий статус | Континентальный родственник |
|---|---|---|---|---|
| Мегапод для сваестроителей | Megapodius molistructor | Новая Каледония и Тонга | Вымершие (ок. 1500 г. до н. э.) |  Кустарник |
| Мега лозы | Мегавитиорнис альтирострис | Фиджи | Вымерший |  курообразные |
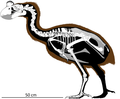 Сильвиорнис | Сильвиорнис неокаледония | Новая Каледония и остров Пайнс | Вымерший |
| Пример | Биномиальное имя | Родной диапазон | Текущий статус | Континентальный родственник |
|---|---|---|---|---|
 Красный рельс | Афанаптерикс боназия | Маврикий | Вымершие (ок. 1700 г. н. э.) |  Рельсы |
 рельс Хокинса | Диафанаптерикс ястреб | Острова Чатем | Вымершие (ок. 1900 г. н.э.) | |
 Антильская пещерная железная дорога | Незотрохис дебоойи | Пуэрто-Рико и Виргинские острова | Вымерший | |
| Кубинская пещерная железная дорога | Незотрохис пикиценсис | Куба | Вымерший | |
| Гаитянская пещерная железная дорога | Несотрохис стеганинос | Эспаньола | Вымерший | |
 Разлом Южного острова | Порфирио Хохштеттери | Южный остров , Новая Зеландия | Находящийся под угрозой исчезновения | |
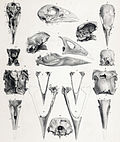 Разлом Северного острова | Порфировая мантия | Северный остров , Новая Зеландия | Вымершие (до 1900 г. н. э.) | |
 Адзебиллы | Экскаватор Апторнис А. отидиформис | Новая Зеландия | Вымерший |  Мадагаскарский пушистик [22] |
 Чатем Кут | Фулика чатаменсис | Острова Чатем | Вымершие (после 1500 г. н. э.) |  Рыжая лысуха и другие лысухи |
 Маскаренская лысуха | лягушка Ньютона | Маврикий и Реюньон | Вымершие (ок. 1700 г. н. э.) | |
| Новозеландская лысуха | Маленькая лысуха | Новая Зеландия | Вымершие (после 1280 г. н. э.) | |
 Болотная встреча | Лазурный порфир | Plaine des Cafres , Реюньон | Вымершие (ок. 1730 г. н. э.) |  Пурпурные болота |
Голуби
[ редактировать ]| Пример | Биномиальное имя | Родной диапазон | Текущий статус | Континентальный родственник |
|---|---|---|---|---|
 Гигантский голубь Вити-Леву | Натунаорнис гигура | Вити-Леву , Фиджи | Вымерший |  Венценосные голуби |
| Человек-голубь | Калоенас канакорум | Новая Каледония | Вымершие (ок. 500 г. до н. э.) |  Никобарский голубь |
 Родригеш одинок | Пезофапс одиночный | Родригес | Вымершие (до 1778 г. н. э.) | |
 Додо | Рафус кукуллатус | Маврикий | Вымершие (ок. 1662 г. н.э.) |
| Пример | Биномиальное имя | Родной диапазон | Текущий статус | Континентальный родственник |
|---|---|---|---|---|
| Пещера Лико беркут | Аквила хризетос симург | Крит | Вымершие (поздний плейстоцен) |  Беркут |
| Гигантский ястреб-краб [23] | Бутеогаллус борраси | Куба | Вымерший |  Большой черный ястреб и другие ястребы |
| Гигантский ястреб | Гигантоиеракс сп. | Куба | Вымерший | |
| Титан-ястреб | Titanohierax gloveralleni | Куба , Эспаньола и Багамы | Вымерший | |
| Ямайская каракара | Каракара наземная | Ямайка | Вымерший |  Каракарас |
| лунь Эйлса | цирковой номер | Новая Зеландия | Вымершие (ок. 1000 г. н. э.) |  Болотный лунь |
 Орлы острова Гаргано | Гарганоэтус Фреудентали Г. муриворус | Остров Гаргано | Вымершие (поздний миоцен) | Аквила дельфинская |
 Орел Хааста | Hieraaetus moorei | Новая Зеландия | Вымершие (ок. 1400 г. н. э.) |  Маленький орел |
 Филиппинский орел | Питекофага Джеффери | Филиппины | Находится под угрозой исчезновения |  Жонглер [24] |
Попугаи
[ редактировать ]| Пример | Биномиальное имя | Родной диапазон | Текущий статус | Континентальный родственник |
|---|---|---|---|---|
 Попугай Геркулес | Геракл неожиданный | Новая Зеландия | Вымершие ( миоцен ) |  Другие попугаи |
 Какапо | Стригопс хаброптилус | Новая Зеландия | Находящийся под угрозой исчезновения | |
 Ширококлювый попугай | Lophopsittacus mauritianus | Маврикий | Вымершие (ок. 1680 г. н. э.) |  Пситтакулиновые попугаи |
Совы
[ редактировать ]| Пример | Биномиальное имя | Родной диапазон | Текущий статус | Континентальный родственник |
|---|---|---|---|---|
 Критская сова | Афины на Крите | Крит | Вымершие (плейстоцен) |  Маленькая сова |
 Кубинские гигантские совы | Орнимегалоникс виды. | Куба | Вымершие (плейстоцен) |  Лесные совы |
 Большая гигантская сова Гаргано | Тито гигантская | Остров Гаргано | Вымершие ( поздний миоцен ) |  Сипухи |
 Сипуха острова Андрос | Эти пыльцы | Остров Андрос , Багамы | Вымершие (до 1600 г. н. э.) | |
 сипуха Риверо | Эти риверой | Куба | Вымерший | |
 Гигантская сова Малая Гаргано | Эти робусты | Остров Гаргано | Вымершие ( ранний плиоцен ) |
| Пример | Биномиальное имя | Родной диапазон | Текущий статус | Континентальный родственник |
|---|---|---|---|---|
| Новозеландский совёнок-козодой | Эготелес novazelandiae | Новая Зеландия | Вымершие (ок. 1200 г. н. э.) |  Австралийский совенок-козодой |
 Новокаледонский совенок-козодой | Эготелес спаси | Новая Каледония | Находится под угрозой исчезновения |
| Пример | Биномиальное имя | Родной диапазон | Текущий статус | Континентальный родственник |
|---|---|---|---|---|
 Чатемский ворон | Ворона мертвых | Острова Чатем | Вымерший | Новозеландский ворон |
 Длинноногая овсянка | Эмбериза альковери | Тенерифе | Вымершие (после 1 г. н.э.) |  овсянка Кабаниса |
| Гигантский нукупуу | Hemignathus vorpalis | Гавайи | Вымершие (после 1000 г. н. э.) |  Зяблики |
| Тасманский превосходный крапивник | Malurus cyaneus cyaneus | Тасмания | Наименьшее беспокойство |  Превосходный крапивник |
| Великолепный крапивник с острова Кенгуру | Malurus cyaneus ashbyi | Остров Кенгуру | Наименьшее беспокойство | |
| Толстоногий крапивник | Pachyplichas yaldwyni | Южный остров Зеландии Новой | Вымерший |  Другие воробьиные |
 Сент-Килда крапивник | Троглодиты troglodytes hirtensis | Сент-Килда, Шотландия | Неизвестный |  Евразийский крапивник |
 Козерог серебристоглазый | Зостеропс латеральный хлороцефал | Козерог и группа бункеров Австралии Большого Барьерного рифа | Неизвестный |  Сильверай |
Рептилии
[ редактировать ]| Пример | Биномиальное имя | Родной диапазон | Текущий статус | Континентальный родственник | Островной / материковый соотношение длины или массы |
|---|---|---|---|---|---|
 Тонганская гигантская игуана [25] | Брахилофус гиббонси | Приехал | Вымершие (ок. 800 г. до н. э.) |  Южноамериканский игуаны | |
 Фиджийская гигантская игуана [26] | Стоимость лапитигуаны | Фиджи | Вымершие (ок. 1000 г. до н. э.) | ||
 Чаквалла с острова Ангела | Савромалус гиспидус | Остров Ангела-Хранителя , Нижняя Калифорния | Рядом с угрозой | Полуостров Чаквалла | МР ≈ 5 [27] |
 Сан-Эстебан Чаквалла | Савромалус вариус | Остров Сан-Эстебан , Нижняя Калифорния | Находящийся под угрозой исчезновения | МР ≈ 5 [27] |
Гекконы
[ редактировать ]| Пример | Биномиальное имя | Родной диапазон | Текущий статус | Континентальный родственник | Островной / материковый соотношение длины или массы |
|---|---|---|---|---|---|
 Гигантский геккон Делькура | Гигарканум делькурти | Новая Каледония | Вымершие (ок. 1870 г. н.э.) |  Диплодактилидные гекконы | ЛР ≈ 6,75 [с] |
 Новокаледонский гигантский геккон | Ракодактилус лечианус | Новая Каледония | Наименьшее беспокойство | ЛР ≈ 4,4 [д] МИ ≈ 60 [и] | |
Гигантский дневной геккон Родригеса | Фелсума Гигас | Родригес | Вымершие (ок. 1850 г. н.э.) |  Дневные гекконы |
Сцинки
[ редактировать ]| Пример | Биномиальное имя | Родной диапазон | Текущий статус | Континентальный родственник |
|---|---|---|---|---|
 Мабуя Вайланта | Хиониния вайланти | Кабо-Верде | Находящийся под угрозой исчезновения |  Материковые мабуйины сцинки- |
 Гигантский сцинк Кабо-Верде | Макросцинкус коктей | Кабо-Верде | Вымершие (после 1900 г. н. э.) | |
Маврикийский гигантский сцинк | Лейолопизма мавританская | Маврикий | Вымершие (после 1600 г. н. э.) | Материковые эугогилиновые сцинки |
| Сцинк террора | Фобосцинкус бокурти | Остров Пайнс у Новой Каледонии | Находящийся под угрозой исчезновения |  Материковые эугогилиновые сцинки |
| Гигантский сцинк Кисиноуэ | Plestiodon kishinouyei | Острова Мияко и острова Яэяма , Япония. | Уязвимый |  Asian Plestiodon spp. |
| Пример | Биномиальное имя | Родной диапазон | Текущий статус | Континентальный родственник |
|---|---|---|---|---|
| Гигантская ящерица Ла-Пальмы | Галлоция аурита | Пальма | Находится под угрозой исчезновения |  Средиземноморские ящерицы-песчанки |
 Гигантская ящерица Ла Гомеры | Галлоция Бравоана | Gomera | Находится под угрозой исчезновения | |
 Тенерифская гигантская ящерица [31] | Галлотия-голиаф | Тенерифе | Вымершие (ок. 1500 г. н. э.) | |
 Гигантская ящерица Эль Йерро | Галлоция симони | Железо | Находится под угрозой исчезновения | |
 Гран-Канарская гигантская ящерица | Галлоция Стелини | Гран-Канария | Наименьшее беспокойство |
Змеи
[ редактировать ]| Пример | Биномиальное имя | Родной диапазон | Текущий статус | Континентальный родственник |
|---|---|---|---|---|
| Крапчатая гремучая змея с острова Анхель-де-ла-Гуарда | Crotalus mitchellii angelensis | Остров Анхель-де-ла-Гуарда у Нижней Калифорнии | Наименьшее беспокойство |  Крапчатая гремучая змея |
| Популяция полосатых змей Таданаэ-дзима [32] | Элапа четырехвиргата | Остров Таданаэ-дзима недалеко от Токио | Неизвестный |  Японская полосатая змея |
 Популяции островных тигровых змей | Обратите внимание на щиты | Острова Маунт-Чаппел ( Тасмания ); Уильямс , Хопкинс и архипелаг Нуйтс (вся Южная Австралия ) [33] | Наименьшее беспокойство [34] |  Тигровая змея |
| Длинноносая змея Исла Серральво | Rhinocheilus lecontei etheridgei | Остров Жака Кусто у южной Нижней Калифорнии | Неизвестный |  Длинноносая змея |
Сомнительные примеры
[ редактировать ]

- Комодский варан на Флоресе и близлежащих островах, крупнейшая из ныне живущих ящериц, и похожий (вымерший) гигантский варан из Тимора считались примерами гигантских островных хищников. Поскольку острова, как правило, предлагают ограниченную пищу и территорию, их хищники-млекопитающие (если они есть) обычно меньше, чем континентальные. Эти случаи касаются экзотермических хищников на островах, слишком маленьких, чтобы поддерживать значительную конкуренцию со стороны млекопитающих . Однако эти ящерицы не такие большие, как их вымершая австралийская родственница мегалания , и на основании ископаемых свидетельств было высказано предположение, что предки этих варанид сначала развили свои крупные размеры в Австралии, а затем рассеялись в Индонезии. [35] Если это правда, то их можно было бы рассматривать не как островных гигантов, а как примеры филетического гигантизма . В пользу этой интерпретации свидетельствуют сведения о существовании в плиоценовой Индии ящерицы Varanus sivalensis , сравнимой по размеру с V. komodoensis . [35] Тем не менее, учитывая, что Австралию часто называют самым большим островом в мире и что родственная ей мегалания, крупнейшая наземная ящерица, известная в летописи окаменелостей, была ограничена Австралией, восприятие крупнейших австралазийских/индонезийских ящериц как островных гигантов все еще может иметь некоторое значение. действительность.
- Гигантские черепахи на Галапагосских и Сейшельских островах , крупнейшие из ныне живущих черепах, а также вымершие черепахи Маскаренских и Канарских островов часто считаются примерами островного гигантизма. Однако во время плейстоцена черепахи сопоставимого размера или более крупные обитали в Австралии ( Meiolania ), Южной Азии ( Megalochelys ), Европе . [36] ( Титанохелон ), Мадагаскар ( Aldabrachelys ), Северная Америка. [37] ( Hesperotestudo ) и Южная Америка [38] ( Chelonoidis , тот же род, который сейчас встречается на Галапагосских островах). [39] ), и на ряде других, более доступных островов Океании и Карибского бассейна . [37] В позднем плиоцене они также присутствовали в Африке (« Geochelone » laetoliensis). [40] ). Нынешняя ситуация, когда крупные черепахи встречаются только на отдаленных островах, по-видимому, отражает то, что эти острова были открыты людьми недавно и не были густо заселены, что делает их черепах менее подверженными чрезмерной эксплуатации .
| Пример | Биномиальное имя | Родной диапазон | Текущий статус | Континентальный родственник | Островной / материковый соотношение длины или массы |
|---|---|---|---|---|---|
| Гигантская древесная лягушка Сан-Томе | Гиперолиус Томенсис [41] | Остров Сан-Томе | Находящийся под угрозой исчезновения |  Африканские тростниковые лягушки | |
| Пальмовая лесная лягушка | Лептопелис пальматус [41] | Принсипи Остров | Уязвимый |  Красная древесная лягушка | ЛР ≈ 1,2 [ф] |
| Гигантская наземная лягушка Фиджи | Распространяющийся мегаботонивит [44] | Вити-Леву , Фиджи | Вымерший |  Азиатские платимантины | |
| Гигантская травяная лягушка Сан-Томе | Птихадена Ньютони [41] | Остров Сан-Томе | Находящийся под угрозой исчезновения |  Маскаренская травяная лягушка |
| Пример | Биномиальное имя | Родной диапазон | Текущий статус | Континентальный родственник |
|---|---|---|---|---|
 Кокосовый краб | Биргус вор | Индийского океана Острова и Полинезия [45] | Уязвимый |  Coenobita Раки-отшельники |
 Гигантская вета | Дейнакрида виды. | Новая Зеландия | Переменная |  Южноафриканские королевские сверчки |
| Гигантский псевдоскорпион [46] | Гарипус титаниус | Боцман Птичий остров | Находящийся под угрозой исчезновения |  Гарипоиды |
 Шипящие тараканы | Gromphadorhini spp. | Мадагаскар | Неизвестный |  Блабериды |
 Уховертка Святой Елены | Лабидура геркулеана | Остров Святой Елены | Вымершие (ок. 1967 г. н.э.) |  Береговая уховертка |
 Гигантская пчела Уоллеса | Мегачилийский Плутон | Северные Молуккские острова | Уязвимый |  Калломегачиле |
 Мегалара | Мегалара Гаруда | Горы Меконга , Сулавеси | Неизвестный |  Краброниновые осы |
 Мадагаскарский гигантские таблетки-многоножки | Виды Microsphaerotherium . Sphaeromymus spp. Zoosphaerium spp. | Мадагаскар | Неизвестный |  Индийские гигантские таблетки-многоножки ( Артросфера ) |
 Орсонвеллес | Орсонвеллес виды. | Гавайи | Неизвестный |  Денежные пауки |
 Гигантский древесный сверчок Конанта Нихоа | Тауматогриллус конанти | Это ушло | Неизвестный |  Древесные сверчки |
 Гигантский фиджийский усач [47] | Герои Ксисутра | Вити-Леву , Фиджи | Находящийся под угрозой исчезновения |  Австралазийский ксишутр |
| Жук Тавеуни | Девятнадцатый ужасный | Тавеуни , Фиджи | Неизвестный |
| Пример | Биномиальное имя | Родной диапазон | Текущий статус | Континентальный родственник |
|---|---|---|---|---|
 Наземные улитки Каури | Парифанта виды. Powelliphanta виды. | Новая Зеландия | Рядом с угрозой |  Другие ритидиды |
Флора
[ редактировать ]Помимо увеличения размеров, островные растения могут также проявлять «островную деревянистость». [48] Наиболее яркими примерами являются мегатравы островов Новой Зеландии субантарктических . [ нужна ссылка ] Увеличенный размер листьев и семян также был зарегистрирован у некоторых островных видов независимо от формы роста (травянистый, кустарниковый или древовидный ). [49]
| Пример | Биномиальное имя | Родной диапазон | Текущий статус | Континентальный родственник |
|---|---|---|---|---|
 Морковь с острова Кэмпбелл | Анизотом широколистный | Кэмпбелл и Окленд Острова | Неизвестный |  Зонтичные |
 Росс Лили | Bulbinella rossii | Кэмпбелл и Окленд Острова | Естественно необычно |  Новозеландская лилия маори |
 Острова Чатем Корокио [49] [50] | Корокия макрокарпа | Острова Чатем | Неизвестный |  Новозеландский таракан [51] |
 Черноглазая ромашка | Дамнамения верникоза | Окленд и острова Кэмпбелл | Естественно необычно |  Астереи |
 Огуречное дерево [52] | Дендросициос сокотранус | Сокотра | Уязвимый |  тыквы |
 Коко де Мер [53] [52] | Лодоицея мальдивская | Сейшельские острова | Находящийся под угрозой исчезновения |  Борассоидные пальмы |
 | Плеврофиллум криниферум | Антиподы , острова Окленд и Кэмпбелл. | Неизвестный |  Цинерария |
 Серебристолистная ромашка | Плеврофиллум хукери | Остров Маккуори , острова Окленд и Кэмпбелл | Неизвестный | |
 Маргаритка с острова Кэмпбелл | Плеврофиллум особенный | Кэмпбелл и Окленд Острова | Естественно необычно | |
 Капуста острова Маккуори | Стильбокарпа полярная | Остров Маккуори и субантарктические острова Новой Зеландии | Уязвимый |  Аралиевые |
См. также
[ редактировать ]- Глубоководный гигантизм
- Островная прирученность
- Островная карликовость
- Мегафауна
- Плейстоценовые вымирания
Примечания
[ редактировать ]- ^ Сокращение хищничества на островах часто также приводит к более укрощенному поведению островных видов добычи - тенденция, которая была проанализирована на ящерицах. [2] [3]
- ^ Самый ранний известный предок новозеландского киви, предположительно недавно прибывший из Австралии. [20]
- ^ Судя по предполагаемой общей длине H. delcourti , ~ 23,6 дюйма, [28] а средняя длина представителя Diplodactylus , наиболее богатого видами рода австралийских диплодактилидных гекконов, составляет ~3,5 дюйма. [29]
- ^ Судя по средней общей длине более крупных подвидов, R. l. leachianus , ~15,5 дюйма, [30] а средняя длина представителя Diplodactylus , наиболее богатого видами рода австралийских диплодактилидных гекконов, составляет ~3,5 дюйма. [29]
- ^ Судя по средней массе более крупных подвидов, R. l. лечианус , ~240 г, [30] при средней массе члена Diplodactylus , наиболее видового рода австралийских диплодактилидных гекконов, ~4 г. [29]
- ^ На основе средней длины от морды до отверстия самки (SVL) L. palmatus , ~ 96 мм, [42] при средней СВЛ самки L. rufus ~80 мм. [43]
Ссылки
[ редактировать ]- ^ Герцег, Великобритания; Гонда, Алабама; Мериля, Дж. (16 июля 2009 г.). «Эволюция гигантизма у девятииглых колюшек». Эволюция . 63 (12): 3190–3200. дои : 10.1111/j.1558-5646.2009.00781.x . ПМИД 19624722 . S2CID 205782326 .
- ^ Купер, МЫ; Пайрон, РА; Гарланд, Т. (8 января 2014 г.). «Островная прирученность: жизнь на островах сокращает расстояние начала полета» . Труды Королевского общества B: Биологические науки . 281 (1777): 20133019. doi : 10.1098/rspb.2013.3019 . ПМК 3896029 . ПМИД 24403345 .
- ^ Йонг, Э. (08 января 2014 г.). «Острова укрощают животных». Природа . дои : 10.1038/nature.2014.14462 . S2CID 183158746 .
- ^ Яффе, Алабама; Слейтер, Дж.Дж.; Альфаро, Мэн (26 января 2011 г.). «Эволюция островного гигантизма и изменение размеров тела черепах и черепах» . Письма по биологии . 7 (4): 558–561. дои : 10.1098/rsbl.2010.1084 . ПМК 3130210 . ПМИД 21270022 .
- ^ Бараона, Ф.; Эванс, SE; Матео, Дж.А.; Гарсиа-Маркес, М.; Лопес-Хурадо, LF (март 2000 г.). «Эндемизм, гигантизм и вымирание островных ящериц: род Gallotia на Канарских островах» . Журнал зоологии . 250 (3): 373–388. дои : 10.1017/s0952836900003101 . HDL : 10553/19918 .
- ^ Jump up to: а б Райя, П.; Мейри, С. (август 2006 г.). «Островное правило у крупных млекопитающих: палеонтология встречается с экологией». Эволюция . 60 (8): 1731–1742. дои : 10.1111/j.0014-3820.2006.tb00516.x . ПМИД 17017072 . S2CID 26853128 .
- ^ Кин, Дж. Э.; Ньето, Северная Каролина; Трейси, ЧР; Гингер, CM; Фельдман, ЧР (27 августа 2013 г.). «Эволюция на необитаемом острове: разница в размерах тела между рептилиями острова Анахо в Неваде и материка вокруг озера Пирамид». Журнал зоологии . 291 (4): 269–278. дои : 10.1111/jzo.12066 .
- ^ Ломолино, М.В. (05.09.2005). «Эволюция размеров тела островных позвоночных: общность правила острова». Журнал биогеографии . 32 (10): 1683–1699. Бибкод : 2005JBiog..32.1683L . дои : 10.1111/j.1365-2699.2005.01314.x . hdl : 2027.42/146565 . S2CID 85866114 .
- ^ Филин И.; Зив, Ю. (2004). «Новая теория островной эволюции: объединение потери дисперсности и изменения массы тела» (PDF) . Исследования в области эволюционной экологии . 6 : 115–124. Архивировано из оригинала (PDF) 25 января 2020 г. Проверено 18 ноября 2014 г.
- ^ Терви, ST (2006). «Новый род и вид гигантской хутии ( Tainotherium valei ) из четвертичного периода Пуэрто-Рико: вымершее древесное четвероногое животное?». Журнал зоологии . 270 (4): 585–594. дои : 10.1111/j.1469-7998.2006.00170.x .
- ^ Торрес-Ройг, Э.; Агусти, Дж.; Бовер, П.; Альковер, JA (2017). «Новый гигантский крицетин из базального плиоцена Майорки (Балеарские острова, западное Средиземноморье): биостратиграфическая связь с континентальными зонами млекопитающих». Историческая биология . 31 (5): 559–573. дои : 10.1080/08912963.2017.1377194 . S2CID 135302585 .
- ^ Фрейденталь, М. (1985). Cricetidae (Rodentia) из неогена Гаргано (пров. Фоджа, Италия) . Национальный музей геологии и минералогии.
- ^ «Изучены огромные полевые мыши Сент-Кильды» . Би-би-си . 3 сентября 2010 г. Проверено 02 марта 2020 г.
- ^ «Ошибка» (PDF) .
- ^ Тимм, РМ; Вейола, В.; Аплин, КП; Доннеллан, Южная Каролина; Фланнери, ТФ; Томсон, В.; Пайн, Р.Х. (12 апреля 2016 г.). «Новый вид Rattus (Rodentia: Muridae) с острова Манус, Папуа-Новая Гвинея» . Журнал маммологии . 97 (3): 861–878. дои : 10.1093/jmammal/gyw034 . hdl : 1808/20678 .
- ^ http://www.raco.cat/index.php/Orsis/article/viewFile/24434/24268 [ пустой URL PDF ]
- ^ Дамс, Р.; Фрейденталь, М. (1985). « Stertomys laticrestatus , новый глирид (соня, Rodentia) из островной фауны Гаргано (провинция Фоджа, Италия)» . Геологический сценарий 77 : 21–27.
- ^ Jump up to: а б Макфи, RDE, Итурральде-Винент, Массачусетс, и Гаффни, ES (февраль 2003 г.). «Домо-де-Заза, местонахождение позвоночных раннего миоцена на юге-центральной части Кубы, с заметками о тектонической эволюции Пуэрто-Рико и проливе Мона» . Американский музей Novitates (3394): 1–42. doi : 10.1206/0003-0082(2003)394<0001:DDZAEM>2.0.CO;2 . HDL : 2246/2820 . S2CID 55615855 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ «Позднемеловые животные румынского острова Хацег - более сложный взгляд» .
- ^ Достойно, Тревор Х.; и др. (2013). Окаменелости миоцена показывают, что киви ( Apteryx , Apterygidae), вероятно, не являются филетическими карликами (PDF) . Палеорнитологические исследования 2013, Материалы 8-го Международного собрания Общества палеонтологии и эволюции птиц . Проверено 16 сентября 2017 г.
- ^ Павия, М.; Мейер, HJM; Росси, Массачусетс; Гёлих, УБ (11 января 2017 г.). «Крайняя островная адаптация Garganornis ballmanni Meijer, 2014: гигантские гусеобразные неогена Средиземноморского бассейна» . Королевское общество открытой науки . 4 (1): 160722. Бибкод : 2017RSOS....460722P . дои : 10.1098/rsos.160722 . ПМК 5319340 . ПМИД 28280574 .
- ^ «Африканское происхождение загадочного Adzebill» .
- ^ Нэйш, Даррен (28 января 2008 г.). «Титаны-ястребы и другие суперхищники» . Блог четвероногих зоологии . ООО «Научные блоги» . Проверено 2 марта 2011 г.
- ^ Лернер, Хизер Р.Л.; Минделл, Дэвид П. (2005). «Филогения орлов, стервятников Старого Света и других Accipitridae на основе ядерной и митохондриальной ДНК». Молекулярная филогенетика и эволюция . 37 (2): 327–346. дои : 10.1016/j.ympev.2005.04.010 . ПМИД 15925523 .
- ^ Прегилл, ГК; Стедман, Д.В. (март 2004 г.). «Игуаны южной части Тихого океана: воздействие человека и новый вид». Журнал герпетологии . 38 (1): 15–21. дои : 10.1670/73-03А . JSTOR 1566081 . S2CID 85627049 .
- ^ Прегилл, ГК; Достойно, TH (март 2003 г.). «Новая ящерица-игуанид (Squamata, Iguanidae) из четвертичного периода Ларе на Фиджи, юго-западная часть Тихого океана». Герпетологика . 59 (1): 57–67. doi : 10.1655/0018-0831(2003)059[0057:ANILSI]2.0.CO;2 . ISSN 0018-0831 . S2CID 85804786 .
- ^ Jump up to: а б Петрен, К.; Кейс, Ти Джей (1997). «Филогенетический анализ эволюции размеров тела и биогеографии чакволл ( Sauromalus ) и других игуан». Эволюция . 51 (1): 206–219. дои : 10.1111/j.1558-5646.1997.tb02402.x . ПМИД 28568786 . S2CID 22032248 .
- ^ Уилсон, К.-Дж. (2004). Полет Гуйи: экология и сохранение новозеландских лягушек, рептилий, птиц и млекопитающих . Издательство Кентерберийского университета. ISBN 0-908812-52-3 . OCLC 937349394 .
- ^ Jump up to: а б с Стюарт, К. (9 мая 2014 г.). « Диплодактилусы Австралии» . Reptilesmagazine.com/ . Проверено 29 февраля 2020 г.
- ^ Jump up to: а б Бергман Дж.; Хампер, Р. (2016). «Инструкция по уходу за новокаледонским гигантским гекконом» . Reptilesmagazine.com/ . Проверено 29 февраля 2020 г.
- ^ Мака-Мейер, Н.; Карранса, С.; Рандо, Джей Си; Арнольд, EN; Кабрера, В.М. (1 декабря 2003 г.). «Статус и взаимоотношения вымершей гигантской ящерицы с Канарских островов Gallotia goliath (Reptilia: Lacertidae), оцененные с использованием древней мтДНК из ее мумифицированных останков». Биологический журнал Линнеевского общества . 80 (4): 659–670. дои : 10.1111/j.1095-8312.2003.00265.x .
- ^ https://www.jstage.jst.go.jp/article/hsj2000/21/1/21_1_43/_pdf [ пустой URL PDF ]
- ^ Кио, Дж.С.; Скотт, IAW; Хейс, К. (январь 2005 г.). «Быстрое и неоднократное возникновение островного гигантизма и карликовости у австралийских тигровых змей» . Эволюция . 59 (1): 226–233. дои : 10.1111/j.0014-3820.2005.tb00909.x . ПМИД 15792242 . S2CID 58524 .
- ^ Майкл, Д.; Клеманн, Н.; Робертсон, П. (2018). « Нотечис скутатус » . Красный список видов, находящихся под угрозой исчезновения МСОП . 2018 : e.T169687A83767147 . Проверено 19 декабря 2019 г.
- ^ Jump up to: а б Хокналл, ЮАР; Пайпер, ПиДжей; ван ден Берг, Грузия; Долг, РА; Морвуд, MJ; Курниаван, И. (2009). «Потерянный рай дракона: палеобиогеография, эволюция и вымирание крупнейших в истории наземных ящериц (varanidae)» . ПЛОС ОДИН . 4 (9): е7241. Бибкод : 2009PLoSO...4.7241H . дои : 10.1371/journal.pone.0007241 . ПМЦ 2748693 . ПМИД 19789642 .
- ^ Перес-Гарсия А., Влахос Э. и Аррибас А. (2017). Последняя гигантская континентальная черепаха Европы: выжившая на стоянке Фонелас P-1 в испанском плейстоцене. Палеогеография, Палеоклиматология, Палеоэкология , 470, 30–39.
- ^ Jump up to: а б Хансен, DM; Донлан, CJ; Гриффитс, CJ; Кэмпбелл, Кей Джей (апрель 2010 г.). «Экологическая история и скрытый природоохранный потенциал: большие и гигантские черепахи как модель для замены таксонов» (PDF) . Экография . 33 (2): 272–284. Бибкод : 2010Экогр..33..272H . дои : 10.1111/j.1600-0587.2010.06305.x . Проверено 2 марта 2012 г.
- ^ Сион, Алабама; Тонни, EP; Сойбельзон, Л. (2003). «Разорванный зигзаг: позднекайнозойское вымирание крупных млекопитающих и черепах в Южной Америке» . Преподобный Муз. Аргентино Сиенс. Нат . НС 5 (1): 1–19. дои : 10.22179/REVMACN.5.26 . ISSN 1514-5158 .
- ^ Фаринья, Р.А., Вискаино, С.Ф. и Де Юлиис, Г. (2013) Мегафауна: гигантские звери Южной Америки. Издательство Индианского университета, 448 страниц.
- ^ Харрисон, Т. (2011). «Черепахи (Chelonii, Testudinidae)». Палеонтология и геология Лаэтоли: эволюция человека в контексте, Том. 2: Ископаемые гоминины и связанная с ними фауна . Палеобиология и палеоантропология позвоночных. Springer Science+Business Media . стр. 479–503. дои : 10.1007/978-90-481-9962-4_17 . ISBN 978-90-481-9961-7 .
- ^ Jump up to: а б с Мизи, Дж.Дж.; Венсес, М.; Древес, Колорадо; Киари, Ю.; Мело, М.; Бурлз, Б. (2006). «Пресноводные пути через океан: молекулярная филогения лягушки Ptychadena newtoni дает представление о колонизации земноводными океанических островов». Журнал биогеографии . 34 (1): 7–20. дои : 10.1111/j.1365-2699.2006.01589.x . S2CID 17562846 .
- ^ « Лептопелис пальматус » . amphibiaweb.org . Калифорнийский университет, Беркли. 2008 год . Проверено 29 февраля 2020 г. .
- ^ « Лептопелис руфус » . amphibiaweb.org . Калифорнийский университет, Беркли. 2008 год . Проверено 29 февраля 2020 г. .
- ^ Достойно, TH (2001). «Новый вид Platymantis (Anura: Ranidae) из четвертичных отложений на Вити-Леву, Фиджи» . Палеонтология . 44 (4): 665–680. Бибкод : 2001Palgy..44..665W . дои : 10.1111/1475-4983.00197 .
- ^ Ни кокосовые крабы, ни их родственники не могут плавать дальше стадии личинки, поэтому взрослые особи на практике высаживают животных. Кокосовые крабы могут весить более 4 кг (9 фунтов); Самый крупный рак-отшельник родственного рода Coenobita , C. brevimanus прибрежной Африки и Азии, достигает всего 230 граммов (0,5 фунта).
- ^ «План действий по сохранению биоразнообразия острова Вознесения: План действий по видам Garypus titanius » (PDF) . Джорджтаун, остров Вознесения: Государственный департамент охраны природы острова Вознесения. 26 февраля 2015 г. Архивировано из оригинала (PDF) 20 сентября 2020 г. Проверено 11 сентября 2019 г.
- ^ Кеппель, Гуннар; Лоу, Эндрю Дж.; Поссингем, Хью П. (2009). «Изменение взглядов на биогеографию тропической части южной части Тихого океана: влияние расселения, наместничества и вымирания». Журнал биогеографии . 36 (6): 1035–1054. Бибкод : 2009JBiog..36.1035K . дои : 10.1111/j.1365-2699.2009.02095.x . ISSN 0305-0270 . S2CID 86478177 .
- ^ Боуэн, Лизабет; Вурен, Дирк Ван (1997). «Островным эндемичным растениям не хватает защиты от травоядных». Биология сохранения . 11 (5): 1249–1254. Бибкод : 1997ConBi..11.1249B . дои : 10.1046/j.1523-1739.1997.96368.x . ISSN 0888-8892 . S2CID 83497517 .
- ^ Jump up to: а б «Маленькие острова порождают большие семена» .
- ^ «Веб-сайт недоступен» .
- ^ «TER:RAIN — Образовательный ресурс Таранаки: сеть исследований, анализа и информации — кизильник Корокия (Корокио)» .
- ^ Jump up to: а б Бернс, КК (май 2019 г.). Эволюция в изоляции: поиск островного синдрома у растений . Издательство Кембриджского университета . дои : 10.1017/9781108379953 . ISBN 978-1108379953 . OCLC 1105218367 . S2CID 186536407 .
- ^ Проктор, Дж. (1984). «Растительность гранитных островов Сейшельских островов». В Стоддарте, доктор медицинских наук (ред.). Биогеография и экология Сейшельских островов . В. Джанк. ISBN 978-90-6193-881-1 . OCLC 906429733 .
Внешние ссылки
[ редактировать ]| Правила |
|  |
|---|---|---|
| Связанный |
| |