Домовая мышь
| Домовая мышь | |
|---|---|
 | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Сорт: | Млекопитающие |
| Заказ: | Грызуны |
| Семья: | Муриды |
| Род: | Идти |
| Подрод: | Идти |
| Разновидность: | М. мышца |
| Биномиальное имя | |
| Мускулистые мышцы | |
| Подвиды | |
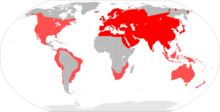 | |
| Ассортимент домовых мышей (примечание: распространение неполное) | |
| Синонимы | |
Мусс вниз | |

Домовая мышь ( Mus musculus ) — небольшое млекопитающее отряда Rodentia , характеризующееся заостренной мордой, большими закругленными ушами и длинным и почти безволосым хвостом. Это один из самых распространенных видов рода Mus . Несмотря на то, что домовая мышь является диким животным , она получила значительную выгоду от общения с человеком до такой степени, что по-настоящему дикие популяции встречаются значительно реже, чем полуручные популяции вблизи человеческой деятельности.
Домовую мышь одомашнили как домашнюю или причудливую мышь , а также как лабораторную мышь , которая является одним из наиболее важных модельных организмов в биологии и медицине. Полный эталонный геном мыши был секвенирован в 2002 году. [3] [4]
Характеристики
[ редактировать ]
Домовые мыши имеют длину тела взрослой особи (от носа до основания хвоста) 7,5–10 см (3–4 дюйма) и длину хвоста 5–10 см (2–4 дюйма). Вес обычно составляет 11–30 г ( 3 / 8–1 унции). В дикой природе они различаются по цвету от серого и светло-коричневого до черного (отдельные волоски на самом деле имеют цвет агути ), но домашние причудливые мыши и лабораторные мыши производятся во многих цветах: от белого до цвета шампанского и черного. [5] У них короткая шерсть, а у некоторых, но не у всех, подвидов светлый живот. [5] Уши и хвост имеют мало шерсти. Задние лапы короткие по сравнению с мышами Apodemus , только 15–19 mm (9⁄16–3⁄4 in) long; the normal gait is a run with a stride of about 4,5 см ( 1 + 3 ⁄ дюйма ), хотя они могут прыгать вертикально на высоту до 45 см (18 дюймов). [6] Голос – пронзительный писк. [7] [8] Домовые мыши процветают в самых разных условиях; они встречаются внутри и вокруг домов и коммерческих построек, а также на открытых полях и сельскохозяйственных угодьях. [ нужна ссылка ]
Новорожденных мальчиков и девочек можно отличить при внимательном рассмотрении, поскольку аногенитальное расстояние у мальчиков примерно вдвое больше, чем у женщин. [9] С возраста около 10 дней у самок имеется пять пар молочных желез и сосков ; у самцов нет сосков. [10] В половозрелом возрасте наиболее ярким и очевидным отличием является наличие яичек у самцов. Они большие по сравнению с остальной частью тела и могут быть втянуты в тело. [ нужна ссылка ]
Хвост, который используется для баланса, [11] [12] [13] имеет лишь тонкий волосяной покров, так как является основным периферическим органом теплоотдачи при терморегуляции. [12] а также — в меньшей степени — безволосые части лап и ушей. Приток крови к хвосту можно точно контролировать в ответ на изменения температуры окружающей среды, используя систему артериовенозных анастомозов для повышения температуры кожи на хвосте на целых 10 ° C (10 K; 18 ° F) для потери тела. нагревать. [14] Длина хвоста варьируется в зависимости от температуры окружающей среды мыши во время постнатального развития, поэтому мыши, живущие в более холодных регионах, как правило, имеют более короткие хвосты. [5] Хвост также используется для баланса, когда мышь карабкается или бежит, или в качестве основы, когда животное стоит на задних лапах (поведение, известное как трипод ), а также для передачи информации о статусе доминирования особи при встречах с другими животными. мыши. [15]
В дополнение к обычному органу тимуса размером с горошину в груди у домашних мышей есть второй функциональный орган тимуса размером с булавочную головку, расположенный на шее рядом с трахеей. [16]
Таксономия и подвиды
[ редактировать ]| Эуархонтоглирес |
| |||||||||||||||||||||||||||

Glires Мыши — млекопитающие клады , что означает, что они входят в число ближайших родственников человека, за исключением зайцеобразных , землеройок , летающих лемуров и других приматов .
три широко распространенных подвида как отдельные виды: Некоторые все чаще рассматривают [17] [18]
- Домовая мышь Юго-Восточной Азии ( Mus musculus castaneus ) (Южная и Юго-Восточная Азия)
- Западноевропейская домовая мышь ( Mus musculus Domesticus ); включает причудливую мышь и лабораторную мышь (Западная Европа, Северная Америка, Южная Америка, Африка и Океания)
- Восточноевропейская домовая мышь ( Mus musculus musculus ) (Восточная Европа и Северная Азия)
Совсем недавно были признаны два дополнительных подвида: [18]
- Домовая мышь Юго-Западной Азии ( Mus musculus bactrianus ) (Юго-Западная и Средняя Азия). Однако из-за значительного генетического сходства, наблюдаемого между ( Mus musculus bactrianus ) и Mus musculus castaneus , обозначение подвида Mus musculus bactrianus теперь подвергается сомнению. [2]
- карликовая домовая мышь ( Mus musculus gentilulus ) (Аравийский полуостров и Мадагаскар) [19]
Домашним мышам были даны названия многих других подвидов, но теперь они считаются синонимами пяти подвидов. Некоторые популяции представляют собой гибриды разных подвидов, в том числе японская домовая мышь ( M. m. molossinus ). [18] Примечательным регионом гибридизации является регион в Европе, где M. m. Domesticus и М. м. musculus часто гибридизуются. [20] Однако самцы гибридных мышей обычно испытывают гибридную стерильность, которая поддерживает репродуктивное разделение между двумя подвидами. [21]
Хромосомные расы
[ редактировать ]Стандартный видовой кариотип состоит из 40 хромосом . В Западной Европе существуют многочисленные популяции – хромосомные расы – с уменьшенным количеством хромосом, возникающим в результате робертсоновского слияния .
Эволюция
[ редактировать ]Сузуки и др. , 2013 подтверждает теорию о M. musculus происхождении в Юго-Западной Азии и выделяет 5 подвидов и их происхождение: musculus в Северной Евразии , castaneus в Индии и Юго-Восточной Азии , ранее неизвестный подвид из Непала , Domesticus в Западной Европе и gentilulus в Йемене . [22]
Недавнее исследование с использованием 89 полногеномных последовательностей показало, что современный Mus musculus castaneus произошел от предковой популяции Mus musculus на Индийском субконтиненте где-то около 700 тысяч лет назад. Оттуда эта предковая популяция мигрировала в Иран около 360 тыс. лет назад, чтобы сформировать Mus musculus Domesticus , а затем в Афганистан около 260 тыс. лет назад, чтобы сформировать Mus musculus musculus . [23]
Поведение
[ редактировать ]
Домашние мыши обычно бегают, ходят или стоят на четвереньках, но во время еды, борьбы или ориентации они встают на задние лапы с дополнительной поддержкой со стороны хвоста - поведение, известное как «стояние на треноге». Мыши — хорошие прыгуны, альпинисты и пловцы, и их обычно считают тигмотактическими , то есть обычно пытаются поддерживать контакт с вертикальными поверхностями. [ нужна ссылка ]
Мыши в основном ведут сумеречный или ночной образ жизни ; они не любят яркий свет. Сообщается, что среднее время сна домашней мыши в неволе составляет 12,5 часов в день. [ нужна ссылка ] Они живут в самых разных укромных местах рядом с источниками пищи и строят гнезда из различных мягких материалов. Мыши территориальны, и один доминирующий самец обычно живет вместе с несколькими самками и детенышами. друг друга Доминирующие самцы уважают территории и обычно заходят на территорию другого, только если она свободна. Если двух или более самцов содержать вместе в клетке, они часто становятся агрессивными, если только их не воспитывали вместе с рождения. [ нужна ссылка ]
Домовые мыши в основном питаются растительной пищей, но всеядны . [ нужна ссылка ] Они едят собственные фекалии , чтобы получить питательные вещества, вырабатываемые бактериями в кишечнике. [24] Домовые мыши, как и большинство других грызунов, не рвут. [25]
Мыши обычно боятся крыс , которые часто их убивают и едят. Такое поведение известно как убийство. Несмотря на это, свободноживущие популяции крыс и мышей все же существуют вместе в лесных районах Новой Зеландии, Северной Америки и других стран. Домовые мыши, как правило, являются плохими конкурентами и в большинстве районов не могут выжить вдали от населенных пунктов в районах, где другие мелкие млекопитающие, такие как лесные мыши . присутствуют [26] Однако в некоторых регионах (например, в Австралии) мыши могут сосуществовать с другими мелкими видами грызунов. [27]
Социальное поведение
[ редактировать ]Социальное поведение домовой мыши не жестко закреплено в видоспецифичных моделях, а вместо этого адаптируется к условиям окружающей среды, таким как наличие пищи и пространства. [28] [29] Эта приспособляемость позволяет домовым мышам обитать в самых разных местах — от песчаных дюн до многоквартирных домов. [28]
Домовые мыши обладают двумя формами социального поведения, выражение которых зависит от контекста окружающей среды. Домовые мыши, обитающие в зданиях и других урбанизированных районах в непосредственной близости от людей, известны как комменсалы . [28] Популяции комменсальных мышей часто имеют чрезмерный источник пищи, что приводит к высокой плотности популяции и небольшим участкам обитания. Это вызывает переход от территориального поведения к иерархии особей. [28] [30] Когда население имеет избыток еды, меньше агрессии между женщинами, которая обычно возникает для получения доступа к пище или предотвращения детоубийства. [28] Агрессия между самцами возникает в комменсальных популяциях, в основном для защиты самок и небольшой территории. [28] [29] популяциях распространен высокий уровень агрессии между мужчинами и женщинами при низком уровне агрессии между женщинами и женщинами В полигамных . [31] Социальная единица популяций комменсальных домовых мышей обычно состоит из одного самца и двух или более самок, обычно связанных между собой. [31] [32] Эти группы размножаются совместно, а самки кормят грудью вместе. Такое совместное разведение и выращивание родственными самками помогает повысить репродуктивный успех. Когда родственных самок нет, племенные группы могут формироваться из неродственных самок. [32]
На открытых территориях, таких как кустарники и поля, популяция домовых мышей известна как некомменсальная. Эти популяции часто ограничены запасами воды или продовольствия и имеют большие территории. [29] Агрессия между самками в популяциях некомменсальных домашних мышей намного выше, достигая уровня, обычно приписываемого видам, живущим на свободном выгуле. Мужская агрессия также выше в некомменсальных популяциях. В комменсальных популяциях самцы довольно часто вступают в контакт с другими самцами из-за высокой плотности популяции, и агрессия должна быть опосредована, иначе риск травм становится слишком большим. [28]
Самцы как комменсальных, так и некомменсальных домовых мышей агрессивно защищают свою территорию и стараются исключить всех злоумышленников. Самцы метят свою территорию запахом мочи. На отмеченных территориях злоумышленники проявляли значительно меньшую агрессию, чем жители территорий. [29] Домовые мыши демонстрируют предпочтение самцов; самцы обычно покидают места своего рождения и мигрируют, образуя новые территории, тогда как самки обычно остаются и являются оппортунистическими заводчиками , а не сезонными. [33]
Чувства и общение
[ редактировать ]Зрение
[ редактировать ]Зрительный аппарат мышей в основном аналогичен зрительному аппарату человека, но отличается тем, что они дихроматы и имеют только два типа колбочек , тогда как люди являются трихроматами и имеют три. Это означает, что мыши не воспринимают некоторые цвета зрительного спектра человека. [34] Однако вентральная мыши область сетчатки имеет гораздо большую плотность чувствительных к ультрафиолету колбочек, чем другие области сетчатки, хотя биологическое значение этой структуры неизвестно. [35] [36] [37] В 2007 году было показано, что мыши, генетически модифицированные для получения колбочек третьего типа, способны различать ряд цветов, аналогичный тому, который воспринимается тетрахроматами. [34]
обоняние
[ редактировать ]Домовые мыши также используют феромоны для социального общения, некоторые из которых производятся препуциальными железами обоих полов. Слезная жидкость и моча самцов мышей также содержат феромоны, такие как основные белки мочи . [38] [39] Мыши обнаруживают феромоны главным образом с помощью сошниково-носового органа (органа Якобсона), расположенного в нижней части носа.
Моча домовых мышей, особенно самцов, имеет характерный резкий запах. не менее 10 различных соединений, таких как алканы , спирты В моче обнаруживаются и т. д. Среди них пять соединений специфичны для мужчин, а именно 3-циклогексен-1-метанол, аминотриазол (3-амино-s-триазол), 4-этилфенол, 3-этил-2,7-диметилоктан и 1-йодундекан. [40]
Запахи взрослых самцов, беременных или кормящих самок могут ускорять или задерживать половое созревание молодых самок и синхронизировать репродуктивные циклы у зрелых самок (т.е. эффект Уиттена ). Запахи незнакомых самцов мышей могут прервать беременность, то есть эффект Брюса .
Тактильный
[ редактировать ]Мыши могут ощущать поверхности и движения воздуха с помощью своих усов , которые также используются во время тигмотаксиса . Если мыши слепы от рождения, сверхнормальный рост вибрисс происходит, по-видимому, как компенсаторная реакция. [41] И наоборот, если вибриссы отсутствуют, использование зрения усиливается. [42]
Жизненный цикл и размножение
[ редактировать ]

У самок домовой мыши эстральный цикл длится от четырех до шести дней, а сама течка длится менее суток. Если несколько самок содержатся вместе в скученных условиях, у них часто вообще не бывает течки. Если затем они подвергнутся воздействию мужской мочи, у них наступит течка через 72 часа. [ нужна ссылка ]
Самцы домовых мышей ухаживают за самками, издавая характерные ультразвуковые звуки в диапазоне 30–110 кГц. [ нужна ссылка ] диапазон. Крики наиболее часты во время ухаживания, когда самец принюхивается и следует за самкой; однако крики продолжаются и после начала спаривания, и в это время крики совпадают с поведением при подъеме. Самцов можно побудить издавать эти крики с помощью женских феромонов. Вокализации, по-видимому, различаются у разных людей, и их сравнивают с песнями птиц из-за их сложности. [43] Хотя самки обладают способностью издавать ультразвуковые сигналы, они обычно не делают этого во время спаривания. [ нужна ссылка ]
После совокупления у самок мышей обычно образуется пробка для спаривания , которая предотвращает дальнейшее совокупление. Вилка не обязательна для начала беременности, так как это произойдет и без вилки. Наличие или отсутствие пробки также не влияет на размер помета. [44] Эта пробка остается на месте около 24 часов. Период беременности составляет около 19–21 дня, в помете рождается 3–14 детенышей (в среднем от шести до восьми). Одна самка может приносить от 5 до 10 пометов в год, поэтому популяция мышей может очень быстро увеличиваться. Размножение происходит в течение всего года. (Однако животные, живущие в дикой природе, не размножаются в более холодные месяцы, хотя и не впадают в спячку .) [ нужна ссылка ]
Щенки рождаются слепыми, без шерсти и ушей. Уши полностью развиваются к четвертому дню, шерсть начинает появляться примерно через шесть дней, а глаза открываются примерно через 13 дней после рождения; щенков отнимают от груди примерно в 21 день. Самки достигают половой зрелости примерно в шесть недель, а самцы — примерно в восемь недель, но оба могут совокупляться уже через пять недель. [45]
Полигамия
[ редактировать ]Хотя домовые мыши могут быть как моногамными, так и полигамными, чаще всего они полигамны . Обычно они демонстрируют характеристики полигинии , защищающей партнера , в том смысле, что самцы очень территориальны и защищают своих партнеров, в то время как самки менее агонистичны . [46] Общинные группы медсестер, возникающие в результате такого поведения, приводят к меньшему количеству детоубийств, поскольку больше женщин способны защитить большее количество потомков. [47]
Эволюционные и поведенческие последствия
[ редактировать ]Полигамная природа домовой мыши приводит к как эволюционным, так и поведенческим последствиям. Одним из последствий является отцовский вклад , который ниже у полигамных мышей, чем у моногамных мышей. [48] Это происходит из-за того, что самцы тратят больше времени на сексуальную конкуренцию , чем самки, оставляя меньше времени на отцовскую заботу. [48] Полигамные домовые мыши-самцы проводят меньше времени наедине со щенками. [48] Они также реже и медленнее находят потерянных детенышей, чем самцы моногамных мышей. [48] Напротив, материнские инвестиции одинаковы у самок мышей, которые спаривались один раз, и у размноженных мышей. [48]
Полигамное поведение самок домашних мышей способствует конкуренции сперматозоидов как самцов, так и самок , что влияет на эволюционную приспособленность . [44] Самки, спаривающиеся с несколькими самцами, как правило, производят на свет обоих детенышей в большем количестве. [44] и с более высокими показателями выживаемости, [49] повышение женской физической формы. Конкуренция спермы, возникающая в результате полигамии, благоприятствует мужчинам с более быстрыми и подвижными сперматозоидами в большем количестве, что повышает мужскую физическую форму. [44] Конкурентный аспект осеменения увеличивает частоту полиандрических событий и оплодотворений. Полиандрия развилась для увеличения репродуктивного успеха. [50] На брачное поведение самцов также влияет практика полигамного поведения. По сравнению с моногамными домашними мышами, полигамные домашние мыши спариваются в течение более длительных периодов времени. [51] Такое поведение позволяет увеличить как передачу спермы, так и успешность отцовства, что, в свою очередь, повышает мужскую приспособленность. [51]
Полиандрия
[ редактировать ]В отличие от полигинии, полиандрическое поведение самок представляет собой акт размножения с несколькими самцами в течение одного сезона. [52] В популяции происходит различие в количестве самцов, с которыми спариваются самки. Полиандрическое поведение является распространенным паттерном спаривания у подвида Mus musculus musculus , а также у его родственника Mus musculus Domesticus . [52]
Полиандрия встречается у 30% всех диких популяций домашних мышей. [53] Пометы от нескольких быков, как правило, более генетически разнообразны, чем пометы от одиночных быков. [52] Множественное отцовство также чаще встречается в больших популяциях, чем в небольших, потому что существует большее количество партнеров и более разнообразные партнеры на выбор. [53] Внутри популяции самцы и самки демонстрируют разную степень множественного спаривания. Во время полового отбора самки проявляют предвзятость к неродственным самцам, а не к родственным самцам, что приводит к более генетически разнообразному потомству и уменьшению инбридинговой депрессии. [50] Инбредная депрессия увеличивает генетическую несовместимость, уровень гомозиготности и вероятность проявления вредных рецессивных аллелей. [50] Было показано, что полиандрия увеличивает выживаемость потомства по сравнению с моандрией.
Эволюционные последствия
[ редактировать ]Приспособленность самок увеличивается у полиандрических линий за счет большего генетического разнообразия и увеличения размера помета. [44]
Из-за полиандрии самцы могут сбить с толку личность нового потомства. [54] Множественные спаривания самок и путаница в отцовстве могут снизить уровень детоубийств. [54] Если самцы не уверены, принадлежит ли потомство им, у них меньше шансов убить потомство. [54]
Внутриматочная инсеминация вызывает эволюционные последствия, возникающие в результате полиандрического поведения. [55] образуется несколько наборов сперматозоидов . Когда несколько самцов спариваются с одной самкой, у самки мыши Потомство, оплодотворенное несколькими самцами, может более активно конкурировать за ресурсы матери и может привести к уменьшению размера тела и изменению размера тела. [55]
Избегание инбридинга
[ редактировать ]Поскольку инбридинг вреден, его стараются избегать. У домашней мыши кластер генов главного белка мочи ( MUP ) обеспечивает высокополиморфный сигнал запаха генетической идентичности, который, по-видимому, лежит в основе распознавания родственников и предотвращения инбридинга. Таким образом, между мышами, имеющими общие гаплотипы MUP, происходит меньше спариваний, чем можно было бы ожидать, если бы спаривание было случайным. [56] Другой механизм предотвращения инбридинга очевиден, когда самка домовой мыши спаривается с несколькими самцами. В таком случае, по-видимому, происходит отбор сперматозоидов, управляемый яйцеклетками, против спермы родственных самцов. [57]
Генетика
[ редактировать ]Область мышиной хромосомы 16 связана с функцией щитовидной железы у мышей. Однако мыши с нокаутом 16 генов (550 т.п.н.) в этом регионе имели нормальный фенотип, исключая именно эти гены из дисфункции, рассматриваемой в этом исследовании. [58]
Ожидаемая продолжительность жизни
[ редактировать ]
Домовые мыши обычно живут менее одного года в дикой природе из-за высокого уровня хищничества и воздействия суровых условий окружающей среды. Однако в защищенной среде они часто живут два-три года. Премия Мафусаила Мауса — это конкурс на выведение или создание чрезвычайно долгоживущих лабораторных мышей. По состоянию на 2005 год [update]Рекордсменом стала генно-инженерная мышь, прожившая 1819 дней (7 дней меньше 5 лет). [59] Другой рекордсмен, который содержался в обогащенной среде, но не получал никакого генетического, фармакологического или диетического лечения, прожил 1551 день (4 года 90 дней). [60] [61]
Старение
[ редактировать ]У нескольких различных линий мышей с возрастом наблюдалось значительное увеличение уровней 8-оксо-2'-дезоксигуанозина (8-оксо-dG) в ядерной ДНК печени , сердца , головного мозга , почек , скелетных мышц и селезенки . [62] Это увеличение повреждения ДНК было связано с возрастным увеличением чувствительности этих тканей к окислительному стрессу . [62] ограничения в питании Известно, что увеличивают продолжительность жизни грызунов и замедляют старение . Было обнаружено, что диетические ограничения значительно снижают возрастное накопление уровней 8-оксо-dG в ядерной ДНК всех тканей, исследованных на мышах. [62] Таким образом, было высказано предположение, что окислительные повреждения ДНК, возникающие в результате нормального клеточного метаболизма, могут иметь большое значение для старения и болезней старения. [62] В другом исследовании было обнаружено, что два типа повреждений ДНК (8-гидрокси-2'-дезоксигуанозин и перекрестные связи ДНК-белок) увеличиваются с возрастом в мозге и печени мышей. [63]
Мыши и люди
[ редактировать ]История
[ редактировать ]Домовые мыши обычно живут рядом с людьми, в домах или полях или вокруг них. Они родом из Индии , [64] [65] а позже они распространились на восточное Средиземноморье около 13 000 г. до н. э., а на остальную Европу распространились только около 1 000 г. до н. э. [66] Считается, что эта временная задержка связана с тем, что мышам необходимы аграрные населенные пункты, превышающие определенный размер. [66] Домовая мышь впервые прибыла в Америку в начале шестнадцатого века. Его возили на борт корабли испанских исследователей и конкистадоров . Примерно сто лет спустя он прибыл в Северную Америку вместе с французскими торговцами мехом и английскими колонистами. С тех пор люди распространили их во все части земного шара. [ нужна ссылка ]
Было проведено множество исследований филогении мышей с целью реконструкции ранних движений человека. Например, одно исследование предполагает возможность ранее не подозревавшейся ранней связи между Северной Европой и Мадейрой на основе происхождения мадейрских мышей. [67] Считалось, что домовые мыши были основной причиной приручения кошек . [ нужна ссылка ]
Как домашние животные
[ редактировать ]
Первое письменное упоминание о мышах, содержащихся в качестве домашних животных, встречается в Эрии , старейшем из сохранившихся китайских словарей, из упоминания в версии 1100 г. до н. э. [68] Одомашнивание человека привело к появлению многочисленных линий « модных » или хобби-мышей с разнообразной окраской и послушным темпераментом . [69] Домашние разновидности домовой мыши разводятся в качестве источника пищи для некоторых хищных домашних рептилий , птиц , членистоногих и рыб . [69] Эффекты одомашнивания могут быть быстрыми: согласно недавним исследованиям, мыши, выращенные в неволе, отличаются смелостью и характером активности по сравнению с мышами, пойманными в дикой природе, после 4–5 поколений. [70] [71]
Мыши как вредители
[ редактировать ]
Мыши — широко распространенные организмы- вредители и одни из наиболее распространенных грызунов, заселяющих человеческие постройки. Весной и летом они обычно добывают корм на открытом воздухе, но осенью и зимой уходят в здания в поисках тепла и еды. Обычно они питаются оставленной без присмотра пищей, остатками и садовыми продуктами. Их добывание пищи сопряжено с риском загрязнения и ухудшения запасов продовольствия, а также может способствовать распространению других вредителей, таких как блохи , клещи , вши и клещи . [72]
Заражая дома, домашние мыши могут представлять опасность повреждения и нарушения конструкции мебели и самого здания. Они грызут различные материалы, чтобы подпилить растущие зубы и контролировать их длину. К распространенным повреждениям относятся обгрызенные электрические провода, следы на деревянной мебели и опорных элементах конструкций, а также повреждения текстиля. [72]
Мыши и болезни
[ редактировать ]Домашние мыши иногда могут передавать болезни, загрязнять продукты питания и повреждать их упаковку. Хотя Центры по контролю и профилактике заболеваний предоставляют список болезней, передающихся грызунами, [73] лишь немногие из болезней передаются через домовую мышь. [ нужна ссылка ]
Лимфоцитарный хориоменингит (LCMV) может передаваться мышами, но не является часто встречающейся инфекцией у людей, хотя большинство инфекций протекают в легкой форме и часто никогда не диагностируются. [74] [75] [76] Существует некоторая обеспокоенность тем, что женщины не должны заражаться LCMV во время беременности. [77] [78]
Домашние мыши обычно не являются переносчиками человеческой чумы ( бубонной чумы ), потому что они меньше заражены блохами, чем крысы, а также потому, что блохи, которые обычно переносят домашние мыши, мало склонны кусать людей, а не своего естественного хозяина. [79]
Риккетсиальная оспа , вызываемая бактерией Rickettsia akari и сходная с ветряной оспой , в целом передается мышами, но встречается очень редко, обычно протекает в легкой форме и при отсутствии лечения проходит в течение двух-трех недель. Смертельных случаев в результате этой болезни не зарегистрировано. Мышиный тиф (также называемый эндемическим тифом), вызываемый бактерией Rickettsia typhi , передается блохами, поражающими крыс. В то время как крысиные блохи являются наиболее распространенными переносчиками, кошачьи блохи и мышиные блохи являются менее распространенными способами передачи инфекции. [80] Эндемический тиф хорошо поддается лечению антибиотиками. Центры по контролю и профилактике заболеваний США в настоящее время не упоминают на своем веб-сайте риккетсиозную оспу или мышиный тиф в отношении болезней, непосредственно передающихся грызунами (в целом). [73]
Лептоспироз переносится различными дикими и домашними животными, включая собак, крыс, свиней, крупный рогатый скот, мышей в целом, и может передаваться через мочу инфицированного животного и заразен, пока моча еще влажная. [81]
В Центральной Европе последовательность хантавируса Dobriva была обнаружена у домашних мышей. Это самый серьезный вид ханты , способный заразить человека. [82]
Инвазивные виды
[ редактировать ]Мыши стали инвазивным видом на островах, куда они распространились в период европейских исследований и колонизации. [ нужна ссылка ]
До прихода человека в Новой Зеландии не было наземных млекопитающих, кроме двух видов летучих мышей, а домовая мышь — один из многих интродуцированных видов. Мыши несут ответственность за сокращение численности местных видов птиц, поскольку они едят ту же пищу, что и птицы. Также известно, что они убивают ящериц и оказывают большое влияние на местных насекомых. [83]
Остров Гоф в Южной Атлантике используется для размножения 20 видами морских птиц, включая почти всех мировых тристанских альбатросов ( Diomedea dabbenena ) и атлантических буревестников ( Pterodroma incerta ). До тех пор, пока в 19 веке с моряками на остров не прибыли домовые мыши, у птиц не было хищников-млекопитающих. С тех пор мыши выросли необычайно большими и научились нападать на птенцов альбатросов , которые могут достигать 90 см в высоту, но в основном неподвижны, работая группами и грызя их, пока они не истечет кровью. [84]
В зерновом поясе юго-восточной Австралии интродуцированный подвид Mus musculus Domesticus размножается настолько успешно, что каждые три года или около того они достигают масштабов чумы, достигая плотности 1000 особей на гектар и нанося огромные разрушения общинам, а также убытки для сельского хозяйства в размере 36 миллионов австралийских долларов в год. . [85]
Как модельный организм
[ редактировать ]
Мыши являются наиболее часто используемыми млекопитающими лабораторными животными из-за их относительно близкого родства и связанной с ними высокой гомологии с человеком, простоты их содержания и обращения, а также высокой скорости воспроизводства. Лабораторные мыши обычно принадлежат к стандартизированным инбредным линиям, отобранным по стабильности или ясности конкретных вредных мутаций. Это позволяет исследованиям на лабораторных мышах легко ограничивать генетические и биологические переменные, что делает их очень полезными модельными организмами в генетических и медицинских исследованиях. [86] Мышей использовали в научных исследованиях с 1650-х годов. [87]
В народной культуре
[ редактировать ]Важность мышей как домашних и сельскохозяйственных вредителей привела к развитию множества ритуалов и историй, связанных с мышами, в мировых культурах. У древних египтян была история о «Мышке как визире ». [88]
У многих южных славян традиционно ежегодно отмечался «День мыши». На восточных Балканах (большая часть Болгарии, Северная Македония , Торлакские районы Сербии ) «День Мыши» ( болг . Миши ден, Мишин ден ) отмечался 9 октября по юлианскому календарю (соответствует 27 октября по григорианскому календарю). календаря в XX и XXI веках), на следующий день после праздника Святого Димитрия . На западных Балканах ( Босния , Хорватия ) День мыши обычно празднуется весной, во время Масленичной недели или в начале Великого поста . [89]
См. также
[ редактировать ]- Мышиный мозг
- Myobia musculi , обыкновенный клещ , обнаруженный в шерсти домовой мыши
Ссылки
[ редактировать ]- ^ Мюссер, Г.; Хаттерер, Р.; Криштуфек, Б.; Йигит, Н.; Мицаинас, Г. (2021 г.) [дополненная версия оценки 2016 г.]. « Мышцы мускулы » . Красный список исчезающих видов МСОП . 2021 : e.T13972A197519724. doi : 10.2305/IUCN.UK.2021-1.RLTS.T13972A197519724.en . Проверено 17 февраля 2022 г.
- ^ Перейти обратно: а б Лаваль, РА; и др. (2022). «Таксономическая оценка двух подвидов диких домашних мышей с использованием полногеномного секвенирования» . Научные отчеты . 12 (20866): 20866. Бибкод : 2022NatSR..1220866L . дои : 10.1038/s41598-022-25420-x . ПМЦ 9718808 . ПМИД 36460842 .
- ^ Грегори С.Г., Сехон М., Шейн Дж., Чжао С., Осоэгава К., Скотт С.Э. и др. (август 2002 г.). «Физическая карта генома мыши». Природа . 418 (6899): 743–50. Бибкод : 2002Natur.418..743G . дои : 10.1038/nature00957 . ПМИД 12181558 . S2CID 4325788 .
- ^ Уотерстон Р.Х., Линдблад-Тох К., Бирни Э., Роджерс Дж., Абриль Дж.Ф., Агарвал П. и др. (декабрь 2002 г.). «Первичное секвенирование и сравнительный анализ генома мыши» . Природа . 420 (6915): 520–62. Бибкод : 2002Natur.420..520W . дои : 10.1038/nature01262 . PMID 12466850 .
- ^ Перейти обратно: а б с Берри, Р.Дж. (1970). «Естественная история домовой мыши» . Полевые исследования . 3 : 219–62 . Проверено 18 декабря 2013 г.
- ^ Бейкер Р.О., Бодман Г.Р., Тимм Р.М. (1994). «Методы строительства и защиты от грызунов» . В Hygnstrom SE, Timm RM, Larson GE (ред.). Предотвращение и контроль ущерба дикой природе . Университет Небраски-Линкольн. [ нужна страница ]
- ^ Линеборг, Л. (1971). Млекопитающие Европы . Блэндфорд Пресс. [ нужна страница ]
- ^ Лоуренс М.Дж., Браун Р.В. (1974). Млекопитающие Британии: их следы, тропы и знаки . Блэндфорд Пресс. [ нужна страница ]
- ^ Хочкисс А.К., Ванденберг Дж.Г. (июль 2005 г.). «Индекс аногенитального расстояния мышей ( Mus musculus Domesticus ): анализ». Современные темы лабораторных зоотехники . 44 (4): 46–8. ПМИД 16050669 .
- ^ Майер Дж.А., Фоли Дж., Де Ла Круз Д., Чуонг С.М. , Виделиц Р. (ноябрь 2008 г.). «Преобразование соска в эпителий, несущий волосы, путем снижения активности пути костного морфогенетического белка на границе дермы и эпидермиса» . Американский журнал патологии . 173 (5): 1339–48. дои : 10.2353/ajpath.2008.070920 . ПМК 2570124 . ПМИД 18832580 .
- ^ Грин, Юнис Чейс (1935). «Анатомия крысы». Труды Американского философского общества, проводимые в Филадельфии для распространения полезных знаний . Труды Американского философского общества. 27 . Американское философское общество: iii–370. дои : 10.2307/1005513 . JSTOR 1005513 . OCLC 685221899 .
- ^ Перейти обратно: а б Сигел, Майкл И. (1970). «Хвост, передвижение и равновесие у мышей». Американский журнал физической антропологии . 33 : 101–2. дои : 10.1002/ajpa.1330330113 .
- ^ Бак, CW, Толман Н., Толман В. (ноябрь 1925 г.). «Хвост как балансирующий орган у мышей». Журнал маммологии . 6 (4): 267–71. дои : 10.2307/1373415 . JSTOR 1373415 .
- ^ Ле Барс Д., Гозариу М., Кэдден С.В. (декабрь 2001 г.). «Животные модели ноцицепции». Фармакологические обзоры . 53 (4): 597–652. ПМИД 11734620 .
- ^ Дрикамер, Ли К. (2005). «Использование хвоста для общения у домашних мышей» . В Викторе Санчес-Кордеро; Медельин, Родриго А. (ред.). в Коллекция млекопитающих честь Бернардо Вильи (на испанском языке). УНАМ. стр. 157–62. ISBN 978-970-32-2603-0 .
- ^ Тершовский Г., Мюллер С.М., Блеул CC, Блюм С., Ширмбек Р., Рейманн Дж., Паскье Л.Д., Амагай Т., Бём Т., Родевальд Х.Р. (апрель 2006 г.). «Доказательства функционального второго тимуса у мышей» . Наука . 312 (5771): 284–7. Бибкод : 2006Sci...312..284T . дои : 10.1126/science.1123497 . ПМИД 16513945 . S2CID 24553384 .
- ^ Митчелл-Джонс Эй Джей; Амори Г.; Богданович В.; Кристуфек Б.; Рейндерс П.Дж.Х.; Шпитценбергер Ф.; Стуббе М; Тиссен JBM; Вогралик В.; Зима Дж. (1999). Атлас млекопитающих Европы . Т. и А.Д. Пойзер. ISBN 978-0-85661-130-8 . [ нужна страница ]
- ^ Перейти обратно: а б с Массер, Гай Г.; Карлтон, Майкл Д. (2005). «Надсемейство Муроидеа» . В Уилсоне, Дон Э.; Ридер, ДиЭнн М. (ред.). Виды млекопитающих мира: таксономический и географический справочник (3-е изд.). Балтимор: Издательство Университета Джонса Хопкинса. стр. 894–1531. ISBN 978-0-8018-8221-0 .
- ^ Прагер Э.М., Оррего С., Сейдж Р.Д. (октябрь 1998 г.). «Генетические вариации и филогеография среднеазиатских и других домашних мышей, включая важную новую митохондриальную линию в Йемене» . Генетика . 150 (2): 835–61. дои : 10.1093/генетика/150.2.835 . ПМК 1460354 . ПМИД 9755213 .
- ^ Тернер, Лесли М; Харр, Беттина (9 декабря 2014 г.). «Полногеномное картирование в гибридной зоне домовой мыши выявляет локусы гибридной стерильности и взаимодействия Добжанского-Мюллера» . электронная жизнь . 3 : e02504. дои : 10.7554/eLife.02504.001 . hdl : 11858/00-001M-0000-0024-5B29-0 . ПМЦ 4359376 . ПМИД 25487987 .
- ^ Форейт, Иржи (октябрь 1996 г.). «Гибридная стерильность у мышей». Тенденции в генетике . 12 (10): 412–417. дои : 10.1016/0168-9525(96)10040-8 . ПМИД 8909138 .
- ^ Чайчан, П.; Мерсье, А.; Галал, Л.; Махиттикорн, А.; Ари, Ф.; Моран, С.; Бумедьен, Ф.; Удонсом, Р.; Хамидович А.; Мюрат, Дж.Б.; Суктана, Ю.; Дарде, ML (2017). «Географическое распространение генотипов Toxoplasma gondii в Азии: связь с соседними континентами». Инфекция, генетика и эволюция . 53 . Эльзевир Б.В .: 227–238. Бибкод : 2017InfGE..53..227C . дои : 10.1016/j.meegid.2017.06.002 . ISSN 1567-1348 . ПМИД 28583867 . S2CID 4698999 .
- Этот обзор цитирует это исследование.
- Сузуки, Х.; Нуноме, М.; Киношита, Г.; Аплин, КП; Фогель, П.; Крюков А.П.; Джин, ML; Хан, Ш.; Марианто, И.; Цучия, К.; Икеда, Х.; Сироиси, Т.; Ёнекава, Х.; Мориваки, К. (2013). «Эволюционная история и история расселения евразийских домовых мышей Mus musculus , уточненная с помощью более обширного географического отбора митохондриальной ДНК» . Наследственность . 111 (5). Портфолио природы : 375–390. дои : 10.1038/hdy.2013.60 . ISSN 0018-067X . ПМК 3806020 . ПМИД 23820581 . S2CID 25657111 . Общество генетиков .
- ^ Лаваль, РА; Дюмон, БЛ (2023). «Наследственная изменчивость и ее влияние на геномы диких домашних мышей» . биоRxiv . дои : 10.1101/2023.11.09.566486 .
- ^ Хилшер-Конклин, Кэрил (1998). «Rattus Biologicus: копрофагия: здоровое поведение ваших крыс» . Газета «Крыса и мышь» .
- ^ Хорн CC, Кимбалл Б.А., Ван Х, Каус Дж., Диенел С., Надь А., Гатрайт Г.Р., Йейтс Б.Дж., Эндрюс П.Л. (2013). «Почему грызунов не может рвать? Сравнительное поведенческое, анатомическое и физиологическое исследование» . ПЛОС ОДИН . 8 (4): е60537. Бибкод : 2013PLoSO...860537H . дои : 10.1371/journal.pone.0060537 . ПМЦ 3622671 . ПМИД 23593236 .
- Чарльз К. Чой (16 апреля 2013 г.). «Почему сурки не чихают» . Живая наука .
- ^ Таттерсолл, Франсуаза Х.; Смит, Р.Х.; Ноуэлл, Ф. (1997). «Экспериментальное заселение контрастных местообитаний домовыми мышами» . Журнал маммологии . 62 (6): 350–8.
- ^ Моро, Дориан; Моррис, Кейт (2000). «Передвижение и рефугиумы короткохвостых мышей Лейкленд-Даунс, Leggadina Lakedownensis , и домашних мышей Mus Domesticus на острове Тевенард, Западная Австралия». Исследования дикой природы . 27 (1): 11–20. дои : 10.1071/WR99016 .
- ^ Перейти обратно: а б с д и ж г Фринта, Дэниел; Слабова, Маркета; Вахова, Хана; Вольфова, Радка; Манклингер, Павел (2005). «Агрессия и комменсализм домашней мыши: сравнительное исследование в Европе и на Ближнем Востоке». Агрессивное поведение . 31 (3): 283–93. дои : 10.1002/ab.15555 .
- ^ Перейти обратно: а б с д Грей, Саманта Дж; Херст, Джейн Л. (1997). «Поведенческие механизмы, лежащие в основе пространственного расселения комменсалов Mus Domesticus и пастбищных Mus spretus ». Поведение животных . 53 (3): 511–24. дои : 10.1006/anbe.1996.0301 . S2CID 54989751 .
- ^ Вольф, Роберт Дж. (1985). «Брачное поведение и выбор самок: их отношение к социальной структуре у пойманных в дикой природе домашних мышей ( Mus musculus ), содержащихся в полуестественной среде». Журнал зоологии . 207 : 43–51. дои : 10.1111/j.1469-7998.1985.tb04914.x .
- ^ Перейти обратно: а б Сенци П., Бансеги О., Гроо З., Альтбекер В. (2012). «Развитие социального поведения двух видов мышей с контрастирующими социальными системами». Агрессивное поведение . 38 (4): 288–97. дои : 10.1002/ab.21431 . ПМИД 25363698 .
- ^ Перейти обратно: а б Добсон, Ф. Стивен; Бодуэн, Клод (2002). «Экспериментальные тесты пространственной ассоциации и родства на моногамных мышах ( Mus spicilegus ) и полигинных мышах ( Mus musculus Domesticus )». Канадский журнал зоологии . 80 (6): 980–6. дои : 10.1139/z02-055 .
- ^ Герлах, Габриэле (1996). «Механизмы эмиграции диких домашних мышей - лабораторное исследование влияния социальной структуры, плотности популяции и агрессии». Поведенческая экология и социобиология . 39 (3): 159–70. дои : 10.1007/s002650050277 . JSTOR 4601248 . OCLC 299796761 . S2CID 24161932 .
- ^ Перейти обратно: а б Одлинг Сми, Люси (2007). «Мыши созданы, чтобы видеть радугу цветов» . Новости@природа . дои : 10.1038/news070319-12 . OCLC 605144848 . S2CID 211729271 .
- ^ Кальдероне Дж.Б., Джейкобс Г.Х. (2009). «Региональные различия относительной чувствительности к ультрафиолетовому излучению сетчатки мыши». Визуальная нейронаука . 12 (3): 463–8. дои : 10.1017/s0952523800008361 . ПМИД 7654604 . S2CID 45068446 .
- ^ Ёкояма С., Ши Ю (декабрь 2000 г.). «Генетика и эволюция ультрафиолетового зрения у позвоночных». Письма ФЭБС . 486 (2): 167–72. Бибкод : 2000FEBSL.486..167Y . дои : 10.1016/s0014-5793(00)02269-9 . ПМИД 11113460 . S2CID 28891403 .
- ^ Нейтц М., Нейтц Дж. (май 2001 г.). «Необычная сетчатка обычной домовой мыши». Тенденции в нейронауках . 24 (5): 248–50. дои : 10.1016/s0166-2236(00)01773-2 . ПМИД 11311361 . S2CID 3756078 .
- ^ Кимото Х., Хага С., Сато К., Тохара К. (октябрь 2005 г.). «Полоспецифичные пептиды экзокринных желез стимулируют вомероназальные сенсорные нейроны мышей». Природа . 437 (7060): 898–901. Бибкод : 2005Natur.437..898K . дои : 10.1038/nature04033 . ПМИД 16208374 . S2CID 4388164 .
- ^ Чамеро П., Мартон Т.Ф., Логан Д.В., Фланаган К., Круз Дж.Р., Сагателян А., Краватт Б.Ф., Стоуэрс Л. (декабрь 2007 г.). «Идентификация белковых феромонов, способствующих агрессивному поведению». Природа . 450 (7171): 899–902. Бибкод : 2007Natur.450..899C . дои : 10.1038/nature05997 . ПМИД 18064011 . S2CID 4398766 .
- ^ Ачираман С., Арчунан Г. (декабрь 2002 г.). «Характеристика летучих веществ в моче у швейцарских мышей-самцов ( Mus musculus ): биоанализ идентифицированных соединений». Журнал биологических наук . 27 (7): 679–86. дои : 10.1007/BF02708376 . ПМИД 12571373 . S2CID 8533630 .
- ^ Раушекер Дж. П., Тиан Б., Корте М., Эгерт У. (июнь 1992 г.). «Кроссмодальные изменения в соматосенсорной системе вибрисса/бочонок у животных с депривацией зрения» . Труды Национальной академии наук Соединенных Штатов Америки . 89 (11): 5063–7. Бибкод : 1992PNAS...89.5063R . дои : 10.1073/pnas.89.11.5063 . JSTOR 2359588 . ПМК 49229 . ПМИД 1594614 .
- ^ Соколов В.Е., Тихонова Г.Н., Тихонов И.А. (1996). «[Роль сенсорных систем в поведении мышей Рюкю ( Mus caroli Banhote, 1902)]». Известия Академии Наук. Серия Биологическая (2): 169–75. ПМИД 8723619 .
- ^ Святой ТЭ, Го Цзы (декабрь 2005 г.). «Ультразвуковые песни мышей-самцов» . ПЛОС Биология . 3 (12): е386. дои : 10.1371/journal.pbio.0030386 . ПМЦ 1275525 . ПМИД 16248680 .
- Майкл Парди (31 октября 2005 г.). «Исследователи добавляют мышей в список существ, которые поют в присутствии партнеров» . Вашингтонский университет в Сент-Луисе .
- ^ Перейти обратно: а б с д и Фирман RC, Симмонс LW (май 2010 г.). «Экспериментальная эволюция качества спермы посредством посткопулятивного полового отбора у домашних мышей» . Эволюция; Международный журнал органической эволюции . 64 (5): 1245–56. дои : 10.1111/j.1558-5646.2009.00894.x . ПМИД 19922447 .
- ^ «Мышиоводство, разведение и развитие» . Университет Каролины, Ирвин, Рекомендации по работе с трансгенными мышами . Университет Каролины. Архивировано из оригинала 4 июля 2007 года.
- ^ Добсон, Ф. Стивен; Жако Катрин (июнь 2002 г.). «Экспериментальные тесты пространственной ассоциации и родства у моногамных и полигинных мышей». Канадский журнал зоологии . 80 (6): 980–986. дои : 10.1139/Z02-055 .
- ^ Добсон, Ф. Стивен; Жако Катрин; Бодуэн, Клод (октябрь 2000 г.). «Экспериментальный тест родственных ассоциаций у домовой мыши». Канадский журнал зоологии . 78 (10): 1807–1812. дои : 10.1139/z00-100 .
- ^ Перейти обратно: а б с д и Патрис Б., Бодуэн С. (октябрь 2000 г.). «Сравнительное исследование родительской заботы между двумя видами грызунов: последствия для системы спаривания мыши Mus spicilegus , строящей насыпи ». Поведенческие процессы . 51 (1–3): 35–43. дои : 10.1016/S0376-6357(00)00117-0 . ПМИД 11074310 . S2CID 12674813 .
- ^ Фирман RC, Симмонс LW (7 марта 2008 г.). «Полиандрия, конкуренция сперматозоидов и репродуктивный успех у мышей» . Поведенческая экология . 19 (4): 695–702. дои : 10.1093/beheco/arm158 .
- ^ Перейти обратно: а б с Дин, доктор медицины; Ардли, КГ; Нахман, М.В. (2006). «Частота множественного отцовства позволяет предположить, что конкуренция сперматозоидов распространена у домашних мышей ( Mus Domesticus )» . Молекулярная экология . 15 (13): 4141–4151. Бибкод : 2006MolEc..15.4141D . дои : 10.1111/j.1365-294x.2006.03068.x . ПМК 2904556 . ПМИД 17054508 .
- ^ Перейти обратно: а б Клемме, Инес; Фирман, Рене Клер (апрель 2013 г.). «Самцы домашних мышей, которые эволюционировали в результате конкуренции сперматозоидов, увеличили продолжительность спаривания и успешность отцовства». Поведение животных . 85 (4): 751–758. дои : 10.1016/j.anbehav.2013.01.016 . S2CID 17428265 .
- ^ Перейти обратно: а б с Тонхаузер К.Е., Тосс М., Мусольф К., Клаус Т., Пенн DJ (январь 2014 г.). «Множественное отцовство у диких домашних мышей ( Mus musculus musculus ): влияние на генетическое разнообразие потомства и массу тела» . Экология и эволюция . 4 (2): 200–9. Бибкод : 2014EcoEv...4..200T . дои : 10.1002/ece3.920 . ПМЦ 3925383 . ПМИД 24558575 .
- ^ Перейти обратно: а б Фирман Р.К., Симмонс Л.В. (март 2008 г.). Снук, Р. (ред.). «Полиандрия помогает избежать посткопулятивного инбридинга у домашних мышей» . Эволюция; Международный журнал органической эволюции . 62 (3): 603–11. дои : 10.1111/j.1558-5646.2007.00307.x . ПМИД 18081715 . S2CID 23933418 .
- ^ Перейти обратно: а б с Оклер И., Кениг Б., Линдхольм А.К. (ноябрь 2014 г.). «Социально опосредованная полиандрия: новое преимущество совместного гнездования млекопитающих» . Поведенческая экология . 25 (6): 1467–1473. дои : 10.1093/beheco/aru143 . ПМЦ 4235584 . ПМИД 25419087 .
- ^ Перейти обратно: а б Фирман, Р.; Симмонс, Л. (2007). «Полиандрия, конкуренция сперматозоидов и репродуктивный успех у мышей» . Поведенческая экология . 19 (4): 695–702. дои : 10.1093/beheco/arm158 .
- ^ Шерборн А.Л., Том М.Д., Патерсон С., Джури Ф., Оллиер В.Е., Стокли П., Бейнон Р.Дж., Херст Дж.Л. (декабрь 2007 г.). «Генетическая основа предотвращения инбридинга у домашних мышей» . Современная биология . 17 (23): 2061–6. Бибкод : 2007CBio...17.2061S . дои : 10.1016/j.cub.2007.10.041 . ПМК 2148465 . ПМИД 17997307 .
- ^ Фирман RC, Симмонс LW (сентябрь 2015 г.). «Гаметические взаимодействия способствуют предотвращению инбридинга у домашних мышей». Экологические письма . 18 (9): 937–43. Бибкод : 2015EcolL..18..937F . дои : 10.1111/ele.12471 . ПМИД 26154782 .
- ^ Гарфилд, Наташа; Караплис, Эндрю К. (2001). «Генетика и животные модели гипопаратиреоза». Тенденции в эндокринологии и обмене веществ . 12 (7). Cell Press : 288–294. дои : 10.1016/s1043-2760(01)00435-0 . ISSN 1043-2760 . ПМИД 11504667 . S2CID 23117178 .
- ^ Гобель, Д. (2013) [2003]. «Последние победители Mprize» . Премия Анджея Бартке за долголетие . Фонд Мафусаила . Проверено 06 февраля 2019 г.
- ^ Коннор, Стив (31 октября 2004 г.). «Самая старая мышь в неволе получила высшую научную награду» . «Индепендент» (Великобритания) . Проверено 30 июля 2013 г.
- ^ «Реверсивный приз» . Фонд Мафусаила. Архивировано из оригинала 30 августа 2008 г. Проверено 14 марта 2009 г.
- ^ Перейти обратно: а б с д Гамильтон М.Л., Ван Реммен Х., Дрейк Дж.А., Ян Х., Го З.М., Кьюитт К., Уолтер К.А., Ричардсон А. Увеличивается ли окислительное повреждение ДНК с возрастом? Proc Natl Acad Sci, США, 28 августа 2001 г.; 98 (18): 10469-74. дои: 10.1073/pnas.171202698. Электронная публикация 2001 г., 21 августа. PMID 11517304; PMCID: PMC56984
- ^ Изотти А., Картилья С., Танингер М., Де Флора С., Балански Р. Возрастное увеличение количества 8-гидрокси-2'-дезоксигуанозина и перекрестных связей ДНК-белка в органах мыши. Мутат Рес. 13 декабря 1999 г.; 446 (2): 215–23. дои: 10.1016/s1383-5718(99)00189-8. PMID 10635344
- ^ Музейные заметки, том 36, выпуск 1 – том 46, выпуск 33 . Университет Небраски.
Домовая мышь родом из Индии.
- ^ Бурсо П., Дин В., Ананд Р., Дарвич Д., Дод Б., фон Даймлинг Ф., Талвар Г.П., Боном Ф. (1996). «Происхождение и распространение домовой мыши: филогения митохондриальной ДНК». Журнал эволюционной биологии . 9 (4): 391–415. дои : 10.1046/j.1420-9101.1996.9040391.x . S2CID 84895257 .
- ^ Перейти обратно: а б Кукки Т., Винье Дж.Д., Оффрэ Дж.К. (2005). «Первое появление домовой мыши ( Mus musculus Domesticus Schwarz & Schwarz, 1943) в Западном Средиземноморье: зооархеологический пересмотр находок субфоссилий» (PDF) . Биологический журнал Линнеевского общества . 84 (3): 429–45. дои : 10.1111/j.1095-8312.2005.00445.x .
- ^ Гюндюз И., Оффрэ Х.К., Бриттон-Дэвидиан Дж., Каталан Дж., Ганем Дж., Рамальиньо М.Г., Матиас М.Л., Сирл Дж.Б. (август 2001 г.). «Молекулярные исследования заселения Мадейрского архипелага домашними мышами». Молекулярная экология . 10 (8): 2023–9. Бибкод : 2001МолЭк..10.2023Г . дои : 10.1046/j.0962-1083.2001.01346.x . ПМИД 11555245 . S2CID 19068030 .
- ^ «История необычных мышей» . Американская ассоциация необычных крыс и мышей. Архивировано из оригинала 5 октября 2009 года . Проверено 29 июля 2013 г.
- ^ Перейти обратно: а б Клуб крыс и мышей Америки
- ^ Кортни Джонс, Стефани К.; Бирн, Филип Г. (декабрь 2017 г.). «Какую роль наследственность играет в трансгенерационных фенотипических реакциях на содержание в неволе? Последствия для управления популяциями в неволе». Зоопарковая биология . 36 (6): 397–406. дои : 10.1002/zoo.21389 . ПМИД 29193268 .
- ^ Кортни Джонс, Стефани К.; Манн, Адам Дж.; Бирн, Филип Г. (апрель 2017 г.). «Влияние неволи на поведение домашних мышей в новой среде: последствия для природоохранной практики» . Прикладная наука о поведении животных . 189 : 98–106. дои : 10.1016/j.applanim.2017.01.007 .
- ^ Перейти обратно: а б «Ущерб имуществу, причиненный заражением домовыми мышами» . Круглосуточная борьба с вредителями . 25 июля 2019 г.
- ^ Перейти обратно: а б «Болезни, передающиеся непосредственно грызунами» . Центры по контролю и профилактике заболеваний (последнее обновление страницы: 7 июня 2011 г.). 04.09.2018.
- ^ «Лимфоцитарный хориоменингит» (PDF) . Центр продовольственной безопасности и общественного здравоохранения Университета штата Айова. Март 2010.
- ^ Верхаег Э.М., Мудроус В., Буйтинг АГ, ван дер Эйк А.А., Тийссен CC (2014). «[Менингит после укуса мыши]» [Менингит после укуса мыши]. Голландский медицинский журнал (на голландском языке). 158 :A7033. ПМИД 25017980 .
- ^ Центры по профилактике заболеваний (CDC) (август 2005 г.). «Временное руководство по минимизации риска заражения вирусом лимфоцитарного хориоменингита человека, связанного с грызунами». ММВР. Еженедельный отчет о заболеваемости и смертности . 54 (30): 747–9. ПМИД 16079740 .
- ^ Джеймисон DJ, Куртис А.П., Белл М., Расмуссен С.А. (июнь 2006 г.). «Вирус лимфоцитарного хориоменингита: новый акушерский патоген?» . Американский журнал акушерства и гинекологии . 194 (6): 1532–6. дои : 10.1016/j.ajog.2005.11.040 . ПМИД 16731068 .
- ^ Бонтиус, диджей (сентябрь 2012 г.). «Вирус лимфоцитарного хориоменингита: недооцененная причина неврологических заболеваний у плода, ребенка и взрослого» . Семинары по детской неврологии . 19 (3): 89–95. дои : 10.1016/j.spen.2012.02.002 . ПМЦ 4256959 . ПМИД 22889536 .
- ^ Шрусбери, JFD (1970). История бубонной чумы на Британских островах . Издательство Кембриджского университета. п. 15.
- ^ «В предыдущем исследовании [1943] сообщалось о домашних мышах, естественным образом зараженных R. typhi в штате Джорджия; однако в нашем исследовании не было обнаружено ни одной ПЦР-положительной мыши. Высыпания популяций мышей в отсутствие крыс были причастны к нескольким вспышки мышиного тифа, однако эти наблюдения не были подтверждены лабораторными данными». Еремеева М.Е., Варашина В.Р., Стерджен М.М., Буххольц А.Е., Олмстед Г.К., Парк С.Ю., Эффлер П.В., Карпаты С.Е. (октябрь 2008 г.). «Rickettsia typhi и R. felis у крысиных блох (Xenopsylla cheopis), Оаху, Гавайи» . Новые инфекционные заболевания . 14 (10): 1613–5. дои : 10.3201/eid1410.080571 . ПМК 2609893 . ПМИД 18826827 .
- ^ Браун К., Прескотт Дж. (февраль 2008 г.). «Лептоспироз у домашних собак: перспективы общественного здравоохранения» . CMAJ . 178 (4): 399–401. дои : 10.1503/cmaj.071097 . ПМЦ 2228361 . ПМИД 18268265 .
- ^ Вайдманн, Манфред; Шмидт, П.; Вачкова, М.; Криванец, К.; Манклинджер, П.; Хуферт, FT (февраль 2005 г.). «Идентификация генетических доказательств распространения вируса Добрава у грызунов с помощью вложенной обратной транскрипции (ОТ)-ПЦР и TaqMan RT-PCR» . Журнал клинической микробиологии . 43 (2): 808–812. doi : 10.1128/JCM.43.2.808-812.2005 . ПМК 548048 . ПМИД 15695684 .
- ^ Кинг, Кэролайн, изд. (1995). Справочник млекопитающих Новой Зеландии . Окленд, Новая Зеландия: Издательство Оксфордского университета. ISBN 978-0-19-558320-5 . [ нужна страница ]
- ^ Ванлесс Р.М., Анхель А., Катберт Р.Дж., генеральный менеджер Хилтон, Райан П.Г. (июнь 2007 г.). «Может ли хищничество агрессивных мышей привести к вымиранию морских птиц?» . Письма по биологии . 3 (3): 241–4. дои : 10.1098/rsbl.2007.0120 . ПМК 2464706 . ПМИД 17412667 .
- ^ «Мыши: пример» . Биотехнология Австралии . Содружество Австралии. Архивировано из оригинала 4 апреля 2015 года . Проверено 25 апреля 2015 г.
- ^ «МГИ — Биология лабораторной мыши» . Информатика.jax.org . Проверено 06 февраля 2019 г.
- ^ д'Иса Р., Фазано С., Брамбилла Р. (июнь 2024 г.). «Редакционная статья: Безвредные для животных методы тестирования поведения грызунов в нейробиологических исследованиях» . Границы поведенческой нейронауки . 18 : 1431310. дои : 10.3389/fnbeh.2024.1431310 . ПМЦ 11232432 . ПМИД 38983871 .
- ↑ Мышь в роли визиря , источник: Эмма Бруннер-Траут, Истории животных из страны фараона , Майнц, Цаберн, 2000.
- ^ Plotnikova, Anna Arkadievna (Анна Аркадьевна Плотникова) (2004). Этнолингвистическая география Южной Славии [ Ethnolinguistic Geography of the South Slav Lands ] (in Russian). Moscow: Indrik. pp. 64–68. ISBN 978-5857592878 .
Дальнейшее чтение
[ редактировать ]- Найби, Джон Г. (2001). «Слуховое общение взрослых» . В Уиллотте, Джеймс Ф. (ред.). Справочник по слуховым исследованиям мышей: от поведения до молекулярной биологии . ЦРК Пресс. стр. 3–18. ISBN 978-1-4200-3873-6 .
Внешние ссылки
[ редактировать ] в этом разделе Использование внешних ссылок может не соответствовать политике и рекомендациям Википедии . ( январь 2017 г. ) |
Таксономия
[ редактировать ]Генетика
[ редактировать ]- Ensembl Mus musculus Браузер генома из Ensembl проекта
- Vega Mus musculus Браузер генома , включает последовательность мыши NOD и аннотацию.
- Просмотрите сборку генома mm10 в браузере генома UCSC .
СМИ
[ редактировать ]- Картинки, видеоролики и апплеты, показывающие анатомию Mus musculus , с сайта www.digimorph.org.
- Архивные фотографии. Короткий текст.
Дальнейшее чтение
[ редактировать ]- Природная мышь, специальный выпуск 2002 г.
- Биология лабораторных грызунов Дэвида Г. Бессельсена
- Информационный бюллетень о домовых мышах от Национальной ассоциации по борьбе с вредителями с информацией о привычках, среде обитания и угрозах для здоровья.
- Исчерпывающая информация о домовых мышах, включая изображения , предоставлена Зоологическим музеем Мичиганского университета.
- «Необычные мыши» включает в себя много поведенческой и физиологической информации.
- Немного информации об убийстве
- Вокализации во время совокупления