Хламидомонада Рейнхардти
| Хламидомонада Рейнхардти | |
|---|---|
 | |
| Научная классификация | |
| (без рейтинга): | Зеленые растения |
| Разделение: | Хлорофита |
| Сорт: | хлорофицеи |
| Заказ: | Хламидомонады |
| Семья: | Хламидомонадовые |
| Род: | Хламидомонада |
| Разновидность: | С. Рейнхардтий |
| Биномиальное имя | |
| Хламидомонада Рейнхардти | |
Chlamydomonas Reinhardtii — одноклеточная зеленая водоросль около 10 микрометров диаметром , плавающая с двумя жгутиками . У него есть клеточная стенка , состоящая из гидроксипролином , богатых гликопротеинов , большой чашеобразный хлоропласт , большой пиреноид и глазной аппарат , воспринимающий свет.
Виды Chlamydomonas широко распространены по всему миру в почве и пресной воде, из которых Chlamydomonas Reinhardtii является одним из наиболее распространенных и распространенных. [1] C. Reinhardtii является особенно хорошо изученным биологическим модельным организмом , отчасти из-за простоты его культивирования и способности манипулировать его генетикой. При освещении C. Reinhardtii может расти фотоавтотрофно, но он также может расти в темноте, если ему снабжают органическим углеродом. С коммерческой точки зрения C. Reinhardtii представляет интерес для производства биофармацевтических препаратов и биотоплива, а также является ценным исследовательским инструментом в производстве водорода .
История
[ редактировать ]Лабораторный штамм C.rainhardtii дикого типа c137 (mt+) происходит от изолята, собранного недалеко от Амхерста, штат Массачусетс , в 1945 году Гилбертом М. Смитом. [2] [3]
Название вида писалось по-разному из-за разной транслитерации названия с русского языка: rainhardi , Reinhardii и Reinhardtii относятся к одному и тому же виду, C. Reinhardtii Dangeard. [4]
Описание
[ редактировать ]Клетки Chlamydomonas reinhardtii в основном сферические, но могут иметь эллипсоидную, яйцевидную, обратнояйцевидную или асимметричную форму. Их длина 10–22 мкм, ширина 8–22 мкм. Клеточная стенка тонкая, без сосочков. Жгутики в 1,5–2 раза длиннее тела клетки. Клетки содержат один чашеобразный хлоропласт, выстилающий дно клетки, с одним базальным пиреноидом. [1]
Пятно под глазом
[ редактировать ]C. Reinhardtii имеет глазной аппарат, сходный с таковым у динофлагеллят . [5] Глазное пятно расположено вблизи экватора клетки. Он состоит из богатого каротиноидами слоя гранул в хлоропласте , который действует как отражатель света. [6] Основная функция глазного пятна — фототаксис , заключающийся в движении (с помощью жгутиков ), связанном со световым раздражителем. [7] Фототаксис имеет решающее значение для водорослей и позволяет локализовать среду с оптимальными световыми условиями для фотосинтеза. [8] Фототаксис может быть положительным или отрицательным в зависимости от интенсивности света. [5] Фототаксический путь состоит из четырех этапов, приводящих к изменению баланса биений между двумя жгутиками (цис-жгутик, который находится ближе всего к глазному пятну, и транс-жгутик, который находится дальше всего от глазного пятна). [7]
Модельный организм
[ редактировать ]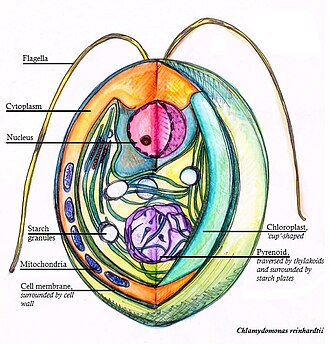
Хламидомонада используется в качестве модельного организма для исследования фундаментальных вопросов клеточной и молекулярной биологии, таких как:
- Как движутся клетки?
- Как клетки реагируют на свет?
- Как клетки узнают друг друга?
- Как клетки генерируют регулярные, повторяющиеся жгутиковые сигналы?
- Как клетки регулируют свой протеом, чтобы контролировать длину жгутиков ?
- Как клетки реагируют на изменение минерального питания? (азот, сера и т. д.)
Известно много мутантов C.rainhardtii . Эти мутанты являются полезными инструментами для изучения различных биологических процессов, включая подвижность жгутиков, фотосинтез и синтез белка . Поскольку виды Chlamydomonas обычно гаплоидны, последствия мутаций проявляются сразу же без дальнейших скрещиваний.
В 2007 году была опубликована полная последовательность ядерного генома C. Reinhardtii . [9]
Каналродопсин -1 и Каналродопсин -2, белки , которые функционируют как светозависимые катионные каналы , первоначально были выделены из C.rainhardtii . [10] [11] Эти и подобные им белки находят все более широкое применение в области оптогенетики . [12]
Митохондриальное значение
[ редактировать ]Геном C. Reinhardtii важен для изучения митохондрий, поскольку это один из видов, у которого гены 6 из 13 белков, кодирующих митохондрии, находятся в ядре клетки, а 7 - в митохондриях. [13] У видов, не относящихся к Chlorophyceae, эти гены присутствуют только в митохондриях и не могут экспрессироваться аллотопически. Это важно для тестирования и разработки методов лечения генетических митохондриальных заболеваний.
Воспроизведение
[ редактировать ]Вегетативные клетки Reinhardtii видов гаплоидны и содержат 17 мелких хромосом . При азотном голодании вегетативные клетки дифференцируются в гаплоидные гаметы. [14] Существует два типа спаривания , идентичных по внешнему виду и, следовательно, изогамных , известных как mt(+) и mt(-) , которые могут сливаться, образуя диплоидную зиготу . Зигота не имеет жгутиков и служит спящей формой вида в почве. На свету зигота подвергается мейозу и высвобождает четыре жгутиковые гаплоидные клетки, которые возобновляют вегетативный жизненный цикл.
В идеальных условиях роста клетки могут иногда подвергаться двум или трем циклам митоза , прежде чем дочерние клетки высвободятся из старой клеточной стенки в среду. Таким образом, один этап роста может привести к образованию 4 или 8 дочерних клеток на одну материнскую клетку.
Клеточный цикл этих одноклеточных зеленых водорослей можно синхронизировать путем чередования периодов света и темноты. Фаза роста зависит от света, тогда как после точки, обозначенной как точка перехода или точка фиксации, процессы становятся независимыми от света. [15]
Генетика
[ редактировать ]Привлекательность водорослей как модельного организма в последнее время возросла после того, как несколько геномных ресурсов стали достоянием общественности. Проект Chlre3 последовательности ядерного генома Chlamydomonas , подготовленный Объединенным институтом генома Министерства энергетики США, включает 1557 каркасов общим размером 120 МБ. Примерно половина генома содержится в 24 каркасах, каждый из которых имеет длину не менее 1,6 МБ. Текущая сборка ядерного генома доступна в Интернете. [16]
Митохондриальный геном размером ~ 15,8 КБ (доступ к базе данных: NC_001638) доступен онлайн в базе данных NCBI. [17] Полный геном хлоропласта размером ~203,8 КБ (доступ к базе данных: NC_005353) доступен в Интернете. [18] [19]
Помимо данных о геномных последовательностях, существует большой запас данных о последовательностях экспрессии, доступных в виде библиотек кДНК и меток экспрессируемых последовательностей (EST). Семь библиотек кДНК доступны в Интернете. [20] Библиотеку BAC можно приобрести в Институте геномики Университета Клемсона. [21] Также имеются две базы данных объемом >50 000 [22] и >160 000 [23] EST доступны в Интернете.
Полногеномная коллекция мутантов с картированными сайтами вставок, охватывающими большинство ядерных генов. [24] [25] доступен: https://www.chlamylibrary.org/ .
геном C. Reinhardtii Было показано, что содержит N 6 -Метиладенозин (м 6 А) признак, распространенный у прокариот, но гораздо реже у эукариот. [26] Некоторые исследования показали, что 6 мА у Chlamydomonas может участвовать в позиционировании нуклеосом, поскольку он присутствует в линкерных областях между нуклеосомами, а также рядом с сайтами начала транскрипции активно транскрибируемых генов. [27]
C. Reinhardtii, по-видимому, способен осуществлять несколько процессов репарации ДНК . [28] К ним относятся рекомбинационная репарация , репарация разрыва цепи и эксцизионная репарация. В частности, C.rainhardtii хлоропласты обладают эффективной системой восстановления двухцепочечных разрывов ДНК . [29] В хлоропластной ДНК гомологичная рекомбинационная репарация сильно стимулируется двухцепочечными разрывами. [29]
Экспериментальная эволюция
[ редактировать ]В этом разделе отсутствует информация о «многоклеточных», дои : 10.1038/s41598-019-39558-8 . ( март 2023 г. ) |
Хламидомонада использовалась для изучения различных аспектов эволюционной биологии и экологии. Это организм, выбранный для многих селекционных экспериментов, потому что:
- у него короткое время генерации,
- одновременно является автотрофом и факультативным гетеротрофом .
- он может размножаться как половым, так и бесполым путем, и
- уже имеется огромное количество генетической информации.
Некоторые примеры (неисчерпывающие) эволюционной работы, проделанной с Chlamydomonas, включают эволюцию полового размножения, [30] фитнес-эффект мутаций, [31] и эффект адаптации к различным уровням CO 2 . [32]
Согласно одной часто цитируемой теоретической гипотезе, [33] половое размножение (в отличие от бесполого размножения) адаптивно поддерживается в благоприятной среде, поскольку оно снижает мутационную нагрузку за счет объединения вредных мутаций из разных линий происхождения и увеличивает среднюю приспособленность. Однако при длительном экспериментальном изучении C.rainhardtii были получены данные, противоречащие этой гипотезе. В половых популяциях не наблюдалось устранения мутаций и не наблюдалось увеличения приспособленности. [34]
Движение
[ редактировать ]
C. Reinhardtii плавает благодаря двум жгутикам. [35] в движении, аналогичном человеческому брассу . Повторяя это элементарное движение 50 раз в секунду, водоросли имеют среднюю скорость 70 мкм/с; [36] генетическое разнообразие различных штаммов приводит к огромному диапазону значений этой величины. После нескольких секунд бега асинхронное биение двух жгутиков приводит к случайному изменению направления — движению, называемому « беги и кувыркайся ». [35] В более широком временном и пространственном масштабе случайное движение водорослей можно описать как явление активной диффузии . [37]
Методы трансформации ДНК
[ редактировать ]Трансформация генов происходит главным образом путем гомологичной рекомбинации в хлоропластах и гетерологичной рекомбинации в ядре. Геном хлоропластов C. reinhardtii можно трансформировать с помощью бомбардировки микрочастицами или перемешивания стеклянными шариками, однако этот последний метод гораздо менее эффективен. Ядерный геном был трансформирован как с помощью перемешивания стеклянными шариками, так и с помощью электропорации. Биолистическая процедура представляется наиболее эффективным способом введения ДНК в геном хлоропластов. Вероятно, это связано с тем, что хлоропласт занимает более половины объема клетки, обеспечивая микроснаряду большую мишень. Показано, что электропорация является наиболее эффективным способом введения ДНК в ядерный геном с максимальными частотами трансформации на два порядка выше, чем при использовании метода стеклянных шариков. [ нужна ссылка ]
Практическое использование
[ редактировать ]Производство биофармацевтических препаратов
[ редактировать ]Генно-инженерный C. Reinhardtii использовался для производства амилоидного белка сыворотки млекопитающих (нужно упоминание), человеческого белка антитела (нужно упоминание), человеческого фактора роста эндотелия сосудов , потенциальной терапевтической вакцины против вируса папилломы человека 16 , [38] потенциальная вакцина от малярии ( вакцина из съедобных водорослей ), [39] и сложный дизайнерский препарат, который можно было бы использовать для лечения рака. [40]
Альтернативный источник белка
[ редактировать ]C. Reinhardtii был предложен в качестве нового источника питания на основе водорослей . Было обнаружено, что по сравнению с хлореллой и спирулиной , C. Reinhardtii содержит больше альфа-линоленовой кислоты и меньшее количество тяжелых металлов, а также содержит все незаменимые аминокислоты и аналогичное содержание белка. [41] Компания Triton Algae Innovations разрабатывала коммерческий альтернативный белковый продукт, изготовленный из Crainhardtii .
Чистый источник производства водорода
[ редактировать ]В 1939 году немецкий исследователь Ганс Гаффрон (1902–1979), работавший в то время в Чикагском университете, открыл водородный обмен одноклеточных зеленых водорослей. C Reinhardtii и некоторые другие зеленые водоросли при определенных обстоятельствах могут перестать производить кислород и вместо этого перейти к производству водорода. Эта реакция гидрогеназы , фермента, активного только в отсутствие кислорода, является кратковременной. В течение следующих тридцати лет Гаффрон и его команда разработали основную механику фотосинтетического производства водорода водорослями. [42]
Чтобы увеличить производство водорода, исследователи идут по нескольким направлениям.
- Первый путь — отделение гидрогеназы от фотосинтеза. Таким образом, накопление кислорода больше не может препятствовать выработке водорода. А если пойти еще дальше и изменить структуру фермента гидрогеназы, становится возможным сделать гидрогеназу нечувствительной к кислороду. Это делает возможным непрерывное производство водорода. В этом случае поток электронов, необходимый для этого производства, больше не исходит от производства сахаров, а поступает от распада собственных запасов крахмала . [43]
- Второй путь — временно прервать процесс фотосинтеза посредством генетических манипуляций с гидрогеназой. Это препятствует достижению кислородом уровня, на котором он способен остановить выработку водорода. [44]
- Третий путь, главным образом исследованный исследователями в 1950-х годах, — это химические или механические методы удаления О2, образующегося в результате фотосинтетической деятельности клеток водорослей. Они включали добавление поглотителей O2, использование дополнительных восстановителей и очистку культур инертными газами. [45] Однако эти методы по своей сути не масштабируемы и могут быть неприменимы к прикладным системам. Появились новые исследования по вопросу удаления кислорода из культур водорослей и могут устранить проблемы с образованием накипи.
- Был исследован четвертый путь, а именно использование солей меди для отделения действия гидрогеназы от производства кислорода. [46]
- Пятый путь был предложен для перенаправления потока фотосинтетических электронов с CO 2 фиксации в цикле Кальвина на гидрогеназу путем воздействия короткими световыми импульсами на анаэробные водоросли. [47] или путем истощения культуры CO 2 . [48]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Перейти обратно: а б Эттл, Х. (1983). Эттл, Х.; Герлофф, Дж.; Хейниг, Х.; Молленхауэр, Д. (ред.). Хлорофита. 1. Часть/Часть 1: Фитомонадины . Пресноводная флора Центральной Европы. Том. 9. ВЭБ Густав Фишер Верлаг. стр. XIV + 808. ISBN 978-3-8274-2659-8 .
- ^ «CC-125 дикого типа mt+ 137c» . Список основной коллекции Центра Хламидомонады . Архивировано из оригинала 27 июля 2009 г. Проверено 9 марта 2009 г.
- ^ Харрис, Стерн и Уитман, 2009 г.
- ^ «Таксономия хламидомонады» . Университет Монреаля.
- ^ Перейти обратно: а б Уэки, Норико; Кобаяси, Юки; Ямагути, Сюдзи; Танака, Дзюн; Хисабори, Тору; Кен-Ичи (2016). «Определение фототаксического признака у Chlamydomonas reinhardtii» . Труды Национальной академии наук . 113 (19) Bibcode : 2016PNAS..113.5299U . . : 5299–5304 пнас.1525538113 . ПМЦ 4868408 .
- ^ Фостер К.В., Смит Р.Д. (декабрь 1980 г.). «Световые антенны в фототаксических водорослях» . Микробиол Рев . 44 (4): 572–630. дои : 10.1128/мр.44.4.572-630.1980 . ПМК 373196 . ПМИД 7010112 .
- ^ Перейти обратно: а б Хегеманн, П.; Бертольд, П. « 13. Сенсорные фоторецепторы и световой контроль жгутиковой активности стр. 395–430 ». В Харрис, Стерн и Уитман (2009) .
- ^ Деммиг-Адамс, Б.; Адамс, WW (1992). «Фотозащита и другие реакции растений на сильный световой стресс». Ежегодный обзор физиологии растений и молекулярной биологии растений . 43 : 599–626. дои : 10.1146/annurev.pp.43.060192.003123 .
- ^ Торговец; Прочник, С.Э.; Валлон, О; Харрис, Э. Х.; Карпович, С.Дж.; Уитман, Великобритания; Терри, А; Саламов А; и др. (2007). «Геном хламидомонады раскрывает эволюцию ключевых функций животных и растений» . Наука . 318 (5848): 245–250. Бибкод : 2007Sci...318..245M . дои : 10.1126/science.1143609 . ПМЦ 2875087 . ПМИД 17932292 .
- ^ Нагель Г., Оллиг Д., Фурманн М. и др. (28 июня 2002 г.). «Канал родопсин-1: светозапираемый протонный канал в зеленых водорослях». Наука . 296 (5577): 2395–8. Бибкод : 2002Sci...296.2395N . дои : 10.1126/science.1072068 . ПМИД 12089443 . S2CID 206506942 .
- ^ Лагали П.С., Баля Д., Аватрамани ГБ, Мюнх Т.А., Ким Д.С., Бускамп В., Чепко С.Л., Роска Б (июнь 2008 г.). «Светоактивируемые каналы, нацеленные на биполярные клетки ON, восстанавливают зрительную функцию при дегенерации сетчатки». Природа Нейронауки . 11 (6): 667–75. дои : 10.1038/nn.2117 . ПМИД 18432197 . S2CID 6798764 .
- ^ Бойден ES и др. (3 мая 2011 г.). «История оптогенетики: разработка инструментов для управления цепями мозга с помощью света» . Отчеты о биологии F1000 . 3 (11): 11. дои : 10.3410/B3-11 . ПМК 3155186 . ПМИД 21876722 .
- ^ Фигероа-Мартинес Ф., Фунес С., Франсен Л.Г., Гонсалес-Хальфен Д. (май 2008 г.). «Реконструкция механизма импорта митохондриального белка Chlamydomonas Reinhardtii» . Генетика . 179 (1): 149–155. doi : 10.1534/genetics.108.087965 . ПМК 2390594 . ПМИД 18493047 .
- ^ Сагер Р., Граник С. (июль 1954 г.). «Питательный контроль сексуальности у Chlamydomonas Reinhardi» . Дж. Генерал Физиол . 37 (6): 729–42. дои : 10.1085/jgp.37.6.729 . ПМК 2147466 . ПМИД 13174779 .
- ^ Ольденхоф Х.; Захледер В.; ден Энде Х. (2006). «Регуляция клеточного цикла синим и красным светом у Chlamydomonas reinhardtii (Chlorophyta)» . Евро. Дж. Фикол . 41 (3): 313–320. Бибкод : 2006EJPhy..41..313O . дои : 10.1080/09670260600699920 .
- ^ « Хламидомонада Рейнхардта v3.0» .
- ^ « Митохондрии Chlamydomonas reinhardtii , полный геном» . Нуклеотид . Национальная медицинская библиотека. 3 апреля 2023 г. Справочная последовательность NCBI: NC_001638.1.
- ^ « Хламидомонада Рейнхардтии хлоропласт, полный геном» . Нуклеотид . Национальная медицинская библиотека. 3 апреля 2023 г. Справочная последовательность NCBI: NC_005353.1.
- ^ «Портал генома хлоропластов хламидомонады» .
- ^ «Центр Хламидомонады — Библиотеки» . Архивировано из оригинала 19 октября 2004 г. Проверено 28 сентября 2006 г.
- ^ «КУГИ» . Архивировано из оригинала 26 декабря 2014 г. Проверено 3 апреля 2006 г.
- ^ «[KDRI] Индекс EST Chlamydomonas reinhardtii» .
- ^ «Поиск ХламиЕСТ» . Хлами Центр. Архивировано из оригинала 4 февраля 2005 г. Проверено 28 сентября 2006 г.
- ^ Ли, Сяобо; Чжан, Ру; Патена, Вероника; Банда, Спенсер С.; Блюм, Шон Р.; Иванова, Нина; Ю, Ребекка; Робертсон, Джейкоб М.; Лефевр, Поль А.; Фитц-Гиббон, Сорел Т.; Гроссман, Артур Р.; Йоникас, Мартин К. (01 февраля 2016 г.). «Индексированная картированная библиотека мутантов позволяет проводить обратные генетические исследования биологических процессов у Chlamydomonas Reinhardtii» . Растительная клетка . 28 (2): 367–387. дои : 10.1105/tpc.15.00465 . ISSN 1040-4651 . ПМЦ 4790863 . ПМИД 26764374 .
- ^ Ли, Сяобо; Патена, Вероника; Фаузер, Фридрих; Джинкерсон, Роберт Э.; Сарусси, Шай; Мейер, Мориц Т.; Иванова, Нина; Робертсон, Джейкоб М.; Ю, Ребекка; Чжан, Ру; Виларраса-Блази, Хосеп; Виткопп, Тайлер М.; Рамундо, Сильвия; Блюм, Шон Р.; Ох, Одри; Лаудон, Мэтью; Шрикумар, Таран; Лефевр, Поль А.; Гроссман, Артур Р.; Йоникас, Мартин К. (апрель 2019 г.). «Полногеномная библиотека мутантов водорослей и функциональный скрининг идентифицируют гены, необходимые для фотосинтеза эукариот» . Природная генетика . 51 (4): 627–635. дои : 10.1038/s41588-019-0370-6 . ISSN 1546-1718 . ПМК 6636631 . ПМИД 30886426 .
- ^ Хаттман, С; Кенни, К; Бергер, Л; Пратт, К. (сентябрь 1978 г.). «Сравнительное исследование метилирования ДНК у трех одноклеточных эукариот» . Журнал бактериологии . 135 (3): 1156–7. дои : 10.1128/JB.135.3.1156-1157.1978 . ПМК 222496 . ПМИД 99431 .
- ^ Фу, Е; Гуань-Чэн; Дэн, Синь; Хань, Хао, Цзянь; Лу, Синъюй, Луи С.; Цюаньцзян, Лоуренс; Хэ, Чуан (май 2015 г.). Метилдезоксиаденозин отмечает активные сайты начала транскрипции в хламидомонады» . клетках - 161 4): 879–892 ( « N6 . PMC 4427561. PMID 25936837 .
- ^ Влчек Д., Шевцовикова А., Свезена Б., Галова Е., Мядокова Е. (январь 2008 г.). «Chlamydomonas Reinhardtii: удобная модельная система для изучения репарации ДНК у фотоавтотрофных эукариот». Карр Жене . 53 (1): 1–22. дои : 10.1007/s00294-007-0163-9 . ПМИД 17992532 .
- ^ Перейти обратно: а б Дюрренбергер Ф., Томпсон А.Дж., Херрин Д.Л., Роше Дж.Д. (сентябрь 1996 г.). «Рекомбинация, индуцированная двунитевым разрывом, в хлоропластах Chlamydomonas reinhardtii» . Нуклеиновые кислоты Рез . 24 (17): 3323–31. дои : 10.1093/нар/24.17.3323 . ПМК 146090 . ПМИД 8811085 .
- ^ Коулгрейв Н. (2002). «Секс снимает ограничение скорости эволюции». Природа . 420 (6916): 664–6. Бибкод : 2002Natur.420..664C . дои : 10.1038/nature01191 . hdl : 1842/692 . ПМИД 12478292 . S2CID 4382757 .
- ^ Де Виссер Дж.А., Хукстра Р.Ф., Ван ден Энде Х. (1996). «Влияние пола и вредных мутаций на приспособленность хламидомонады ». Учеб. Р. Сок. Лонд. Б. 263 (1367): 193–200. дои : 10.1098/rspb.1996.0031 . JSTOR 50473 .
- ^ Коллинз, Белл (2004). «Фенотипические последствия 1000 поколений отбора при повышенном уровне CO 2 у зеленой водоросли». Природа . 431 (7008): 566–9. Бибкод : 2004Natur.431..566C . дои : 10.1038/nature02945 . ПМИД 15457260 . S2CID 4354542 .
- ^ Кондрашов А.С. (октябрь 1984 г.). «Вредные мутации как эволюционный фактор. 1. Преимущество рекомбинации» . Жене. Рез . 44 (2): 199–217. дои : 10.1017/s0016672300026392 . ПМИД 6510714 .
- ^ Ренаут С., Реплански Т., Хепплстон А., Белл Г. (ноябрь 2006 г.). «Экология и генетика приспособленности Chlamydomonas. XIII. Приспособленность долгоживущих сексуальных и бесполых популяций в благоприятной среде». Эволюция . 60 (11): 2272–9. doi : 10.1554/06-084.1 (неактивен 30 июля 2024 г.). ПМИД 17236420 . S2CID 18977144 .
{{cite journal}}: CS1 maint: DOI неактивен по состоянию на июль 2024 г. ( ссылка ) - ^ Перейти обратно: а б Полин, Марко; Тувал, Идан; Дрешер, Кнут; Голлуб, JP; Гольдштейн, Раймонд Э. (24 июля 2009 г.). «Хламидомонада плавает с двумя «шестеренками» в эукариотической версии бегущего движения» . Наука . 325 (5939): 487–490. Бибкод : 2009Sci...325..487P . дои : 10.1126/science.1172667 . ISSN 0036-8075 . ПМИД 19628868 . S2CID 10530835 .
- ^ Гарсия, Микаэль (9 июля 2013 г.). Гидродинамика микропловцов (доктор философии) (на французском языке). Университет Гренобля. тел-00952771.
- ^ Гольдштейн, Раймонд Э (23 июля 2018 г.). «Являются ли теоретические результаты «результатами»?» . электронная жизнь . 7 : е40018. doi : 10.7554/eLife.40018 . ISSN 2050-084X . ПМК 6056240 . ПМИД 30033910 .
- ^ Демуртас О.К.; Масса С; Ферранте П; Венути А; Франкони Р; и др. (2013). «Вакцина против вируса папилломы человека 16 E7, полученная из хламидомонады, вызывает специфическую защиту от опухоли» . ПЛОС ОДИН . 8 (4): e61473. Бибкод : 2013PLoSO...861473D . дои : 10.1371/journal.pone.0061473 . ПМК 3634004 . ПМИД 23626690 .
- ^ «Биологи производят потенциальную вакцину от малярии из водорослей» . ФизОрг. 16 мая 2012 г.
- ^ «Инженерные водоросли для создания сложного противоракового «дизайнерского» лекарства» . ФизОрг. 10 декабря 2012 г.
- ^ Дарвиш, Ранда; Геди, Мохамед; Акепач, Патчания; Ассайе, Хирут; Заки, Абдерлахман; Грей, Дэвид (26 сентября 2020 г.). «Chlamydomonas reinhardtii — потенциальная пищевая добавка, способная превзойти хлореллу и спирулину» . Прикладные науки . 10 (19): 6736. дои : 10.3390/app10196736 . hdl : 20.500.11820/20a558d8-9745-4613-8203-d86234aa4762 . Проверено 26 августа 2021 г.
- ^ Анастасиос Мелис; Томас Хаппе (2004). «Пути исследования водорода в зеленых водорослях — от Ганса Гаффрона к новым рубежам» (PDF) . Исследования фотосинтеза . 80 (1–3): 401–9. Бибкод : 2004PhoRe..80..401M . doi : 10.1023/B:PRES.0000030421.31730.cb . ПМИД 16328836 . S2CID 7188276 .
- ^ Лоран Курнак; Флоренс Муса; Летиция Бернарда; Женевьева Гуденея; Полетт Виньеб; Жиль Пельти (2002). «Ограничивающие этапы производства водорода у Chlamydomonas reinhardtii и Synechocystis PCC 6803 по данным анализа переходных процессов газообмена, индуцированного светом». Международный журнал водородной энергетики . 27 (12.11): 1229–37. Бибкод : 2002IJHE...27.1229C . дои : 10.1016/S0360-3199(02)00105-2 .
- ^ Анастасиос Мелис. «Производство водорода и углеводородного биотоплива путем фотосинтеза микроводорослей» . Архивировано из оригинала 3 апреля 2008 г. Проверено 7 апреля 2008 г.
- ^ Косоуров С.; Цыганов А.; Зайберт, М.; Жирарди, М. (июнь 2002 г.). «Устойчивое фотопроизводство водорода Chlamydomonas Reinhardtii: Влияние параметров культуры». Биотехнология. Биоинж . 78 (7): 731–40. дои : 10.1002/бит.10254 . ПМИД 12001165 .
- ^ Фернандес В.М., Руа М.Л., Рейес П., Каммак Р., Хачикян ЕС (ноябрь 1989 г.). «Ингибирование гидрогеназы Desulfovibrio gigas солями меди и ионами других металлов» . Евро. Дж. Биохим . 185 (2): 449–54. дои : 10.1111/j.1432-1033.1989.tb15135.x . ПМИД 2555191 .
- ^ Косоуров С.; Йокель, М.; Аро, Э.-М.; Аллахвердиева, Ю. (март 2018 г.). «Новый подход к устойчивому и эффективному фотопроизводству H 2 Chlamydomonas Reinhardtii» . Энергетика и экология . 11 (6): 1431–6. дои : 10.1039/C8EE00054A .
- ^ Надь, В.; Подманички А.; Видаль-Мейрелеш, А.; Тенголичс, Р.; Ковач, Л.; Ракхели, Г.; Скома, А.; Тот СЗ. (март 2018 г.). в зеленых водорослях на основе расщепления воды «Устойчивое и эффективное производство H 2 , достигнутое за счет ограничения субстрата в цикле Кальвина-Бенсона-Бэсшема» . Биотехнология для биотоплива . 11:69 . дои : 10.1186/s13068-018-1069-0 . ПМЦ 5858145 . ПМИД 29560024 .
Дальнейшее чтение
[ редактировать ]- Аояма, Х.; Куроива, Т.; Накамура, С. (2009). «Динамическое поведение митохондрий в живых зиготах во время созревания и мейоза у Chlamydomonas Reinhardtii ». Евро. Дж. Фикол . 44 (4): 497–507. Бибкод : 2009EJPhy..44..497A . дои : 10.1080/09670260903272599 .
- Джеймерс, А.; Ленжу, М.; Дерэдт, П.; ван Бокстале, Д.; Бласт, Р.; де Коэн, В. (2009). «Проточный цитометрический анализ зеленых водорослей Chlamydomonas reinhadtii (Chlorophyceae), подвергшихся воздействию кадмия». Евро. Дж. Фикол. . 44 (4): 541–550. Бибкод : 2009EJPhy..44..541J . дои : 10.1080/09670260903118214 .
- Харрис, Э. Х.; Стерн, Д.Б.; Уитман, ГБ, ред. (2009). Справочник по хламидомонаде . Том. 1–3 (2-е изд.). Академический. ISBN 978-0-12-370873-1 .
Внешние ссылки
[ редактировать ]- Ресурсный центр Chlamydomonas — «Центральное хранилище для получения, каталогизации, сохранения и распространения высококачественных и надежных культур дикого типа и мутантных культур зеленой водоросли Chlamydomonas Reinhardtii , а также полезных молекулярных реагентов и наборов для образования и исследований».
- Портал сравнительной геномики растений — ресурсы Chlamydomonas Reinhardtii от Объединенного института генома Министерства энергетики
- Гири, доктор медицины; Гири, ГМ « Хламидомонас Рейнхардти » . База водорослей . Всемирное электронное издание, Национальный университет Ирландии, Голуэй.
- Клетка Chlamydomonas reinhardtii , жизненный цикл, штаммы, типы спаривания — архивная база данных.
| Базы данных органов управления : Национальные |
|---|