Одомашнивание


Одомашнивание отношения между несколькими поколениями — это мутуалистические , в которых вид животных, например люди или муравьи-листорезы , берет на себя контроль и заботу о других видах, таких как овцы или грибы, чтобы получать от них постоянный запас ресурсов, таких как мясо, молоко или труд. Этот процесс является постепенным и географически разбросанным, основанным на методе проб и ошибок.
Первым животным, одомашненным человеком, была собака как комменсал , по крайней мере 15 000 лет назад. Другие животные, включая коз , овец и коров, были одомашнены примерно 11 000 лет назад. Среди птиц курицу впервые одомашнили в Восточной Азии, по-видимому, для петушиных боев, около 7000 лет назад. Лошадь была одомашнена около 5500 лет назад в Центральной Азии как рабочее животное. Среди беспозвоночных шелкопряд соответственно и западная медоносная пчела были одомашнены более 5000 лет назад для получения и меда шелка .
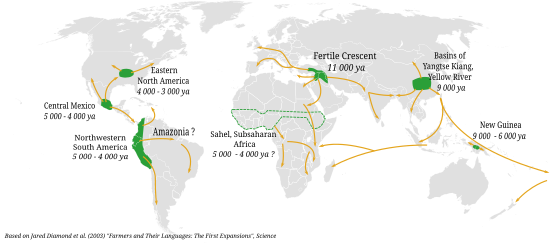
Одомашнивание растений началось около 13 000–11 000 лет назад с таких зерновых , как пшеница и ячмень на Ближнем Востоке , наряду с такими культурами, как чечевица , горох , нут и лен . Впервые рис начали выращивать в Китае примерно 13 500–8 200 лет назад. Примерно 10 000 лет назад коренные народы Америки начали выращивать арахис , тыкву , кукурузу , картофель , хлопок и маниоку . В Африке такие культуры, как сорго были одомашнены . Сельское хозяйство развивалось примерно в 13 центрах по всему миру, где были одомашнены различные сельскохозяйственные культуры и животные.
Одомашнивание повлияло на гены поведения животных, сделав их менее агрессивными. У растений одомашнивание повлияло на морфологические гены, такие как увеличение размера семян и прекращение разрушения семенных головок, как у пшеницы. Такие изменения одновременно облегчают обращение с одомашненными организмами и снижают их способность выживать в дикой природе.
Определения [ править ]
Одомашнивание (не путать с приручением отдельного животного [3] [4] [5] ), происходит от латинского Domesticus , «принадлежащий дому». [6] Этот термин оставался неопределенным до 21 века, когда американский археолог Мелинда А. Зедер определила его как долгосрочные отношения, в которых люди берут на себя контроль и заботу о другом организме, чтобы получить предсказуемый запас ресурса, что приводит к взаимной выгоде. . Далее она отметила, что это не синоним сельского хозяйства, поскольку сельское хозяйство зависит от одомашненных организмов, но не является автоматически результатом одомашнивания. [7]

Майкл Д. Пурудганан отмечает, что одомашнивание трудно дать определение, несмотря на «инстинктивный консенсус» в том, что оно означает «растения и животные, находящиеся под опекой людей, которые приносят нам пользу и которые развивались под нашим контролем». [8] Он отмечает, что такие насекомые, как термиты , жуки-амброзии и муравьи-листорезы , одомашнили некоторые виды грибов , а также отмечает, что другие группы, такие как сорняки и комменсалы, ошибочно называются одомашненными. [8] Отталкиваясь от определения Зедера, Пуругганан предлагает «широкое» определение: «коэволюционный процесс, возникающий в результате мутуализма, в котором один вид (домашний) конструирует среду, в которой он активно управляет как выживанием, так и воспроизводством другого вида (домашненного). для того, чтобы обеспечить первых ресурсами и/или услугами». [8] Он комментирует, что это добавляет создание ниши . к деятельности домоседателя [8]
Синдром одомашнивания — это набор фенотипических признаков, возникших во время первоначального процесса одомашнивания и отличающих сельскохозяйственные культуры от их диких предков . [9] [10] Это также может означать набор различий, наблюдаемых сейчас у одомашненных животных, не обязательно отражающих первоначальный процесс одомашнивания. Изменения включают повышенную послушность и прирученность, окраску шерсти, уменьшение размера зубов, морфологию черепно-лицевой области, форму ушей и хвоста (например, висячие уши), циклы течки, уровни адренокортикотропного гормона и нейротрансмиттеров, пролонгацию ювенильного поведения и уменьшение размера мозга. и отдельных областей мозга. [11]
Причина и время [ править ]
Одомашнивание животных и растений было спровоцировано изменениями климата и окружающей среды, произошедшими после пика Последнего ледникового максимума и продолжающимися по сей день. Эти изменения затруднили добычу пищи путем охоты и собирательства . [12] Первым животным, которое было одомашнено, была собака, по крайней мере, 15 000 лет назад. [1] Младший дриас 12 900 лет назад был периодом сильного холода и засушливости, который заставил людей активизировать свою стратегию добывания пищи, но не благоприятствовал сельскому хозяйству. К началу голоцена 11700 лет назад более теплый климат и увеличение численности населения привели к мелкомасштабному одомашниванию животных и растений, а также к увеличению запасов продовольствия. [13]
| Событие | Центр происхождения | Цель | Дата/лет назад |
|---|---|---|---|
| Собирание дикорастущих зерновых | Азия | Еда | > 23 000 [14] |
| Собака | Евразия | Комменсал | > 15 000 [1] |
| Рис | Китай | Еда | 13,500–8,200 [2] |
| Пшеница , Ячмень | Ближний Восток | Еда | 13,000–11,000 [14] |
| Лен | Ближний Восток | Текстиль | 13,000–11,000 [15] |
| Коза , Овца , Свинья , Корова | Ближний Восток, Южная Азия | Еда | 11,000–10,000 [1] |
| Курица | Восточная Азия | петушиные бои | 7,000 [16] |
| Лошадь | Центральная Азия | Драфт , езда | 5,500 [1] |
За появлением домашней собаки в археологических записях, по крайней мере, 15 000 лет назад, последовало одомашнивание скота и таких культур, как пшеница и ячмень , изобретение сельского хозяйства и переход людей от собирательства к земледелию в разных местах и странах. раз по всей планете. [1] [17] [18] [19] Например, небольшое пробное выращивание зерновых началось около 28 000 лет назад на участке Охало II в Израиле. [20]
показывает, что в Плодородном полумесяце 11 000–10 000 лет назад Зооархеология козы, свиньи, овцы и тауриновый крупный рогатый скот первыми одомашненными домашними животными были . Две тысячи лет спустя горбатый скот зебу был одомашнен на территории современного Белуджистана в Пакистане. В Восточной Азии 8000 лет назад свиньи были одомашнены от диких кабанов, генетически отличающихся от тех, что обитали в Плодородном полумесяце. [1] Кошку одомашнили на территории Плодородного полумесяца примерно 10 000 лет назад. [21] от европейских диких кошек , возможно, для борьбы с грызунами , которые повреждали хранящиеся продукты. [22]
Животные [ править ]
Желательные качества [ править ]

Одомашнивание животных — это отношения между животными, не являющимися людьми, и людьми, которые влияют на их уход и воспроизводство. [7] В своей книге 1868 года «Изменение животных и растений при одомашнивании » Чарльз Дарвин признал небольшое количество черт, которые отличают домашние виды от их диких предков. Он также был первым, кто осознал разницу между сознательным селекционным разведением , при котором люди напрямую отбирают желаемые признаки, и бессознательным отбором, при котором признаки развиваются как побочный продукт естественного отбора или отбора по другим признакам. [25] [26] [27]
Существует разница между домашними и дикими популяциями; некоторые из этих различий составляют синдром приручения — черты, предположительно важные на ранних стадиях приручения, тогда как другие представляют собой черты более позднего улучшения. [9] [28] [29] Домашние животные, как правило, меньше и менее агрессивны, чем их дикие собратья; другие общие черты — висячие уши, меньший мозг и более короткая морда. [24] Признаки одомашнивания обычно фиксируются у всех домашних животных и были выбраны во время начального эпизода одомашнивания этого животного или растения, тогда как признаки улучшения присутствуют только у части домашних животных, хотя они могут быть зафиксированы в отдельных породах или региональных популяциях . [28] [29] [30]
Определенные виды животных и отдельные особи этих видов являются лучшими кандидатами на приручение из-за их поведенческих характеристик: [31] [32] [33] [34]
- Размер и организация их социальной структуры [31]
- Доступность и степень избирательности в выборе партнёра. [31]
- Легкость и скорость, с которой родители связываются со своими детенышами, а также зрелость и подвижность детенышей при рождении. [31]
- Степень гибкости диеты и толерантности к среде обитания. [31]
- Реакция на людей и новую окружающую среду, включая снижение реакции на полет и реакцию на внешние раздражители. [31]
Млекопитающие [ править ]

Начало одомашнивания животных представляло собой длительный коэволюционный процесс, состоявший из нескольких стадий разных путей. Есть три предполагаемых основных пути, по которым большинство одомашненных животных пошли к одомашниванию: [31] [29] [35]
- комменсалы , приспособленные к человеческой нише (например, собаки , кошки , возможно, свиньи ) [31]
- животные-жертвы, ищущие пищу (например, овцы , козы , крупный рогатый скот , водяной буйвол , як , свинья, северный олень , лама и альпака ) [31]
- животные, предназначенные для тягловой и верховой езды (например, лошадь , осел , верблюд ). [31]
Люди не собирались приручать животных ни по комменсальному, ни по пути добычи, или, по крайней мере, они не предполагали, что в результате этого получится одомашненное животное. В обоих случаях люди запутались в этих видах по мере того, как отношения между ними усиливались, а роль человека в их выживании и воспроизводстве постепенно привела к формализованному животноводству . [29] Хотя направленный путь тягловых и верховых животных проходил от отлова к приручению, два других пути не столь целенаправленны, и археологические данные позволяют предположить, что они происходили в течение гораздо более длительных периодов времени. [36]
В отличие от других домашних видов, отобранных в первую очередь по производственным характеристикам, собаки изначально отбирались по их поведению. [37] [38] Собака была приручена задолго до других животных. [39] [40] они утвердились по всей Евразии до конца эпохи позднего плейстоцена , задолго до развития сельского хозяйства . [39]
Археологические и генетические данные позволяют предположить, что долгосрочный двунаправленный поток генов между дикими и домашними животными – например, у ослов , лошадей , верблюдов Нового и Старого Света, коз, овец и свиней – был обычным явлением. [29] [35] Человеческий отбор по домашним признакам, вероятно, противодействовал гомогенизирующему эффекту потока генов от диких кабанов к свиньям и создал острова одомашнивания в геноме . Тот же процесс может применяться и к другим домашним животным. [41] [42]
в 2023 году, Гипотеза о приручении млекопитающих, опосредованном паразитами предполагает, что эндопаразиты , такие как гельминты и простейшие, могли способствовать приручению млекопитающих. Одомашнивание предполагает приручение, имеющее эндокринный компонент; а паразиты могут изменять эндокринную активность и микроРНК . Гены устойчивости к паразитам могут быть связаны с генами синдрома одомашнивания; прогнозируется, что домашние животные менее устойчивы к паразитам, чем их дикие сородичи. [43] [44]
Птицы [ править ]


Под домашними птицами в основном подразумевают домашнюю птицу , выращиваемую ради мяса и яиц: [45] некоторые курообразные ( куры , индейки , цесарки ) и гусеобразные (водоплавающие птицы: утки , гуси , лебеди ). Также широко одомашнены клеточные птицы , такие как певчие птицы и попугаи ; они хранятся как для удовольствия, так и для использования в исследованиях. [46] Домашний голубь использовался как в пищу, так и в качестве средства сообщения между отдаленными местами за счет использования голубиного инстинкта возвращения домой; Исследования показывают, что он был одомашнен еще 10 000 лет назад. [47] Окаменелости курицы в Китае датируются 7400 лет назад. Диким предком курицы является Gallus Gallus , красная лесная птица Юго-Восточной Азии. Судя по всему, изначально этот вид содержался для петушиных боев, а не для еды. [16]
Беспозвоночные [ править ]
Два насекомых , тутовый шелкопряд и западная медоносная пчела , были одомашнены более 5000 лет, часто для коммерческого использования. Шелкопряда выращивают ради шелковых нитей, обматывающих его куколочный кокон; западная медоносная пчела для меда , а с 20 века для опыления сельскохозяйственных культур. [48] [49]
Некоторые другие беспозвоночные, как наземные, так и водные, были одомашнены, в том числе такие, как плодовые мухи Drosophila melanogaster и пресноводная книдария Hydra, для исследований в области генетики и физиологии. Лишь немногие имеют долгую историю одомашнивания. Большинство из них используются для производства продуктов питания или других продуктов, таких как шеллак и кошениль . относятся К этим типам Cnidaria , Platyhelminthes ( для биологической борьбы с вредителями ), Annelida , Mollusca , Arthropoda (морские ракообразные, а также насекомые и пауки) и Echinodermata . Хотя многие морские моллюски используются в пищу, лишь немногие из них были одомашнены, в том числе кальмары , каракатицы и осьминоги , и все они используются в исследованиях поведения и неврологии . Наземных улиток рода Helix выращивают в пищу. Некоторые паразитические или паразитоидные насекомые, включая муху Eucelatoria , жука Chrysolina и осу Aphytis , выращиваются для биологической борьбы. Сознательный или бессознательный искусственный отбор оказывает множество воздействий на одомашненные виды; изменчивость может быть легко утрачена в результате инбридинга, отбора против нежелательных признаков или генетического дрейфа. У дрозофилы вариабельность времени вылета (когда вылупляются взрослые особи) увеличилась. [50]


 Лак-жук Kerria lacca сохранился для шеллаковой смолы.
Лак-жук Kerria lacca сохранился для шеллаковой смолы.




Растения [ править ]
Люди добывали дикие злаки, семена и орехи за тысячи лет до того, как они были одомашнены; Например, дикая пшеница и ячмень были собраны в Леванте по крайней мере 23 000 лет назад. [51] [14] Неолитические общества в Западной Азии впервые начали культивировать, а затем одомашнивать некоторые из этих растений примерно 13–11 000 лет назад. [14] западноазиатского Основополагающими культурами неолита были зерновые ( полба , однозернянка , ячмень ), бобовые ( чечевица , горох , нут , вика горькая ) и лен . [15] [52] Другие растения были независимо одомашнены в 13 центрах происхождения (подразделенных на 24 района) Америки, Африки и Азии (Ближний Восток, Южная Азия, Дальний Восток, Новая Гвинея и Уоллесия); примерно в тринадцати из этих регионов люди начали выращивать травы и зерновые культуры. [53] [54] Впервые рис начали выращивать в Восточной Азии. [2] [55] Сорго широко культивировалось в странах Африки к югу от Сахары. [56] в то время как арахис, [57] давить, [57] [58] хлопок, [57] кукуруза , [59] картофель , [60] и маниока [61] были одомашнены в Америке. [57]
По свидетельствам археологии и генетики, продолжающееся одомашнивание было постепенным и географически разбросанным – происходило множеством маленьких шагов и распространялось на обширную территорию. [62] Это был процесс периодических проб и ошибок, который часто приводил к расхождениям в чертах и характеристиках. [63]
В то время как одомашнивание животных больше всего повлияло на гены, контролирующие поведение, одомашнивание растений больше всего повлияло на гены, которые контролировали морфологию (размер семян, архитектуру растений, механизмы распространения) и физиологию (время прорастания или созревания). [31] [18] как и при одомашнивании пшеницы . Дикая пшеница разбивается и падает на землю, чтобы засеять себя, когда созреет, но одомашненная пшеница остается на стебле, что облегчает сбор урожая. Такое изменение стало возможным из-за случайной мутации в диких популяциях в начале выращивания пшеницы . Пшеница с этой мутацией собиралась чаще и становилась семенами следующего урожая. Поэтому, сами того не осознавая, первые фермеры выбрали эту мутацию. Результатом является одомашненная пшеница, воспроизводство и распространение которой зависит от фермеров. [14]
 Фермеры, выращивающие пшеницу и крупный рогатый скот – древнеегипетское искусство 3400 лет назад.
Фермеры, выращивающие пшеницу и крупный рогатый скот – древнеегипетское искусство 3400 лет назад. Колосья дикой пшеницы при созревании разламываются, но одомашненную пшеницу приходится обмолачивать и провеивать (как показано), чтобы высвободить и отделить зерна. Фотография Гарольда Уэстона , Иран, 1920-е годы.
Колосья дикой пшеницы при созревании разламываются, но одомашненную пшеницу приходится обмолачивать и провеивать (как показано), чтобы высвободить и отделить зерна. Фотография Гарольда Уэстона , Иран, 1920-е годы.


Отличия от дикорастущих растений [ править ]

Одомашненные растения во многом отличаются от своих диких сородичей, в том числе
- отсутствие растрескивания, например, початков злаков (созревших кочанов), [14] потеря плода, опадение [65]
- менее эффективная система размножения (например, без нормальных органов опыления , что требует вмешательства человека), более крупные семена с меньшим успехом в дикой природе, [14] или даже стерильность (например, плоды без косточек ) и поэтому только вегетативное размножение [66] [67]
- лучшие вкусовые качества (например, более высокое содержание сахара, уменьшенная горечь), лучший запах и более низкая токсичность [68] [69]
- съедобная часть крупнее, например, зерна злаков [70] или фрукты [65]
- съедобную часть легче отделить от несъедобной части [70]
- увеличение количества фруктов или зерен [65]
- измененный цвет, вкус и текстура [65]
- от светового дня независимость [65]
- определенный рост [65]
- снижена или отсутствует яровизация [65]
- меньше покоя семян . [65]
Защита растений от травоядных животных , таких как шипы, шипы и колючки , яды, защитные покровы и прочность, возможно, была снижена у одомашненных растений. Это повысит вероятность того, что их съедят травоядные животные, если они не будут защищены людьми, но большая часть этих утверждений имеет лишь слабую поддержку. [68] Фермеры сделали выбор в пользу снижения горечи и токсичности, а также качества продуктов питания, что, вероятно, повысило вкусовые качества сельскохозяйственных культур для травоядных животных, а также для людей. [68] Однако исследование 29 одомашненных растений показало, что сельскохозяйственные культуры были так же хорошо защищены от двух основных насекомых-вредителей ( свекловичная совка и зеленая персиковая тля ) как химически (например, горькими веществами), так и морфологически (например, благодаря стойкости), как и их дикие предки. [71]
Изменения в геноме растений [ править ]

Во время одомашнивания виды сельскохозяйственных культур подвергаются интенсивному искусственному отбору, который изменяет их геномы, устанавливая основные черты, которые определяют их как одомашненные, такие как увеличенный размер зерна. [14] [73] Сравнение кодирующей ДНК хромосомы 8 риса ароматных и неароматных сортов показало, что ароматные и ароматные рисы, включая басмати и жасмин , произошли от предков одомашненного риса, у которого была делеция в экзоне 7, которая изменила кодировку бетаин-альдегида. дегидрогеназа (BADH2). [74] Сравнение генома картофеля с геномом других растений выявило гены устойчивости к фитофторозу картофеля, вызываемому Phytophthora infestans . [75]
В кокосе геномный анализ 10 микросателлитных локусов ( некодирующей ДНК ) обнаружил два эпизода одомашнивания, основанные на различиях между особями Индийского и Тихого океана . [76] [77] Кокос испытал эффект основателя , когда небольшое количество особей с низким разнообразием основало современную популяцию, навсегда потеряв большую часть генетических вариаций дикой популяции. [76] Популяционные узкие места , которые уменьшили вариации по всему геному на более позднем этапе после одомашнивания, очевидны в таких культурах, как просо , хлопок , фасоль обыкновенная и фасоль лимская . [77]
Одомашнивание пшеницы включало повторную гибридизацию и полиплоидию . Эти шаги представляют собой масштабные и, по сути, мгновенные изменения в геноме и эпигеноме , позволяющие быстро эволюционно ответить на искусственный отбор. Полиплоидия увеличивает количество хромосом, создавая новые комбинации генов и аллелей, которые, в свою очередь, делают возможным дальнейшие изменения , например, путем хромосомного скрещивания . [72]
Влияние на микробиом растений [ править ]
Одомашнивание влияет на микробиом населяющих , совокупность микроорганизмов, поверхность и внутренние ткани растений. Сюда входят изменения в видовом составе микробов. [78] [79] [80] и разнообразие. [81] [80] Линия растений, включая видообразование , одомашнивание и селекцию , сформировала эндофиты растений ( филосимбиоз ) по тем же закономерностям, что и гены растений. [80] [82] [83] [84]
Грибы [ править ]

Некоторые виды грибов были одомашнены для использования непосредственно в пищу или в процессе ферментации для производства продуктов питания и лекарств. Культурный гриб Agaricus bisporus широко выращивается в пищу. [85] Дрожжи Saccharomyces cerevisiae на протяжении тысячелетий использовались для брожения пива и вина , а также для закваски хлеба . [86] Плесневые грибы, включая Penicillium, используются для созревания сыров и других молочных продуктов, а также для производства лекарств, таких как антибиотики . [87]
Эффекты [ править ]
О домашних животных [ править ]
Отбор животных по видимым признакам может иметь нежелательные последствия для генетики домашних животных. [88] Побочным эффектом одомашнивания стали зоонозные заболевания. Например, крупный рогатый скот заразил человечество различными вирусными оспами , корью и туберкулезом ; свиньи и утки стали причиной гриппа ; и лошади принесли риновирусы . Многие паразиты также происходят от домашних животных. [89] Наряду с этим, появление одомашнивания привело к увеличению плотности человеческих популяций, что обеспечило зрелые условия для размножения, мутации, распространения патогенов и, в конечном итоге, поиска нового хозяина в организме человека. [90]
Об обществе [ править ]
Ученые выразили самые разные точки зрения на влияние одомашнивания на общество. Анархо-примитивизм критикует приручение как разрушение предполагаемого примитивного состояния гармонии с природой в обществах охотников-собирателей и замену его, возможно, насильственным путем или путем порабощения, социальной иерархией с появлением собственности и власти. [91] Натуралист -диалектал Мюррей Букчин утверждал, что приручение животных, в свою очередь, означало приручение человечества, причем обе стороны неизбежно меняются в результате своих отношений друг с другом. [92] Социолог Дэвид Ниберт утверждает, что приручение животных подразумевало насилие над животными и нанесение ущерба окружающей среде. Это, в свою очередь, утверждает он, развратило человеческую этику и проложило путь к «завоеванию, истреблению, перемещению, репрессиям, принудительному и порабощенному рабству, гендерному подчинению, сексуальной эксплуатации и голоду». [93]
О разнообразии [ править ]

Одомашненные экосистемы обеспечивают пищу, уменьшают хищников и природные опасности, а также способствуют торговле, но их создание привело к изменению или утрате среды обитания, а также к многочисленным вымираниям, начавшимся в позднем плейстоцене. [94]
Одомашнивание снижает генетическое разнообразие одомашненной популяции, особенно аллелей генов, на которые направлен отбор. [95] Одной из причин является узкое место в популяции , созданное искусственным отбором наиболее желательных особей для размножения. Большая часть одомашненных штаммов тогда рождается всего от нескольких предков, создавая ситуацию, подобную эффекту основателя . [96] Одомашненные популяции, такие как собаки, рис, подсолнечник, кукуруза и лошади, имеют повышенную мутационную нагрузку , как и ожидалось в узком популяционном горлышке, где генетический дрейф усиливается из-за небольшого размера популяции. Мутации также можно зафиксировать в популяции путем выборочной проверки . [97] [98] Мутационная нагрузка может быть увеличена за счет снижения селективного давления на умеренно вредные признаки, когда репродуктивная приспособленность контролируется человеком. [24] Однако есть свидетельства против узких мест в таких культурах, как ячмень, кукуруза и сорго, где генетическое разнообразие медленно сокращалось, а не демонстрировало быстрое первоначальное снижение в момент одомашнивания. [97] [96] В дальнейшем генетическое разнообразие этих культур регулярно пополнялось за счет естественной популяции. [97] Аналогичные данные существуют для лошадей, свиней, коров и коз. [24]
Одомашнивание насекомыми [ править ]
По крайней мере, три группы насекомых, а именно жуки-амброзии, муравьи-листорезы и термиты, имеют одомашненные виды грибов . [8]
Жуки-амброзии [ править ]
Жуки-амброзии из долгоносиков подсемейств Scolytinae и Platypodinae роют туннели в мертвых или находящихся в стрессе деревьях, в которых они создают грибные сады, свой единственный источник питания. Приземлившись на подходящее дерево, жук-амброзия выкапывает туннель, в котором выпускает своего грибного симбионта . растения Гриб проникает в ткань ксилемы , извлекает из нее питательные вещества и концентрирует питательные вещества на поверхности хода жука и вблизи нее. Грибы-амброзии обычно плохо разлагают древесину и вместо этого используют менее требовательные питательные вещества. [99] Симбиотические грибы производят и обезвреживают этанол, который является аттрактантом для жуков-амброзии и, вероятно, предотвращает рост антагонистических патогенов и отбирает других полезных симбионтов. [100] Жуки-амброзии преимущественно заселяют древесину недавно погибших деревьев. [101]
Муравьи-листорезы [ править ]
Муравьи-листорезы — это любой из 47 видов листогрызущих муравьев родов Acromyrmex и Atta . Муравьи несут срезанные ими диски листьев обратно в свое гнездо, где они скармливают листовой материал грибам, за которыми они ухаживают. Некоторые из этих грибов не полностью одомашнены: грибы, выращиваемые Mycocepurus smithii, постоянно производят споры, бесполезные для муравьев, которые вместо этого поедают грибковые гифы . С другой стороны, процесс одомашнивания муравьев Атта завершен; это заняло 30 миллионов лет. [102]
Термиты [ править ]
Термиты поедают разлагающийся растительный материал, такой как древесина, опавшие листья и перегной почвы . У многих видов термитов есть специализированная средняя кишка с ферментами, способными расщеплять целлюлозные волокна. [103] Термиты полагаются в первую очередь на симбиотическое микробное сообщество, которое включает бактерии, жгутиковые протисты, такие как метамонады и гипермастигиды . Это сообщество обеспечивает ферменты, которые переваривают целлюлозу, позволяя насекомым поглощать конечные продукты для собственного использования. [104] [105]
- Одомашнивание насекомыми
 Галерея жука-амброзии Xylosandrus crassiusculus раскололась, с куколками и черным грибом. Гриб разлагает материалы древесины, обеспечивая пищу жукам.
Галерея жука-амброзии Xylosandrus crassiusculus раскололась, с куколками и черным грибом. Гриб разлагает материалы древесины, обеспечивая пищу жукам. Муравьи-листорезы Atta cephalotes несут диски листового материала обратно в гнездо, чтобы накормить свой домашний гриб.
Муравьи-листорезы Atta cephalotes несут диски листового материала обратно в гнездо, чтобы накормить свой домашний гриб. Гнездо лесоядного термита Reticulitermes
Гнездо лесоядного термита Reticulitermes трихонимфид Жгутиконосцы из кишечника термита Reticulitermes помогают переваривать древесину.
трихонимфид Жгутиконосцы из кишечника термита Reticulitermes помогают переваривать древесину.




См. также [ править ]
Ссылки [ править ]
- ↑ Перейти обратно: Перейти обратно: а б с д и ж г МакХью, Дэвид Э.; Ларсон, Грегер; Орландо, Людовик (2017). «Укрощение прошлого: древняя ДНК и изучение приручения животных». Ежегодный обзор биологических наук о животных . 5 : 329–351. doi : 10.1146/annurev-animal-022516-022747 . ПМИД 27813680 . S2CID 21991146 .
- ↑ Перейти обратно: Перейти обратно: а б с Нормил, Деннис (1997). «Янцзы считается самым ранним местом выращивания риса». Наука . 275 (5298): 309–310. дои : 10.1126/science.275.5298.309 . S2CID 140691699 .
- ^ Прайс, Эдвард О. (2008). Принципы и применение поведения домашних животных: вводный текст . Издательство Кембриджского университета. ISBN 9781780640556 . Проверено 21 января 2016 г.
- ^ Дрисколл, Калифорния; Макдональд, Д.В.; О'Брайен, SJ (2009). «От диких животных к домашним животным: эволюционный взгляд на приручение» . Труды Национальной академии наук . 106 (Приложение 1): 9971–9978. Бибкод : 2009PNAS..106.9971D . дои : 10.1073/pnas.0901586106 . ПМК 2702791 . ПМИД 19528637 .
- ^ Даймонд, Джаред (2012). «Глава 1». В Гептс, П. (ред.). Биоразнообразие в сельском хозяйстве: одомашнивание, эволюция и устойчивость . Издательство Кембриджского университета . п. 13.
- ^ «Домашний» . Оксфордские словари . Издательство Оксфордского университета . 2014. Архивировано из оригинала 20 июля 2012 года.
- ↑ Перейти обратно: Перейти обратно: а б Зедер, Мелинда А. (2015). «Основные вопросы исследований приручения» . Труды Национальной академии наук Соединенных Штатов Америки . 112 (11): 3191–3198. Бибкод : 2015PNAS..112.3191Z . дои : 10.1073/pnas.1501711112 . ПМЦ 4371924 . ПМИД 25713127 .
- ↑ Перейти обратно: Перейти обратно: а б с д и ж Пуруганан, Майкл Д. (2022). «Что такое приручение?» . Тенденции в экологии и эволюции . 37 (8): 663–671. дои : 10.1016/j.tree.2022.04.006 . ПМИД 35534288 .
- ↑ Перейти обратно: Перейти обратно: а б Олсен, КМ; Вендел, Дж. Ф. (2013). «Обильный урожай: геномный взгляд на фенотипы одомашнивания сельскохозяйственных культур». Ежегодный обзор биологии растений . 64 : 47–70. doi : 10.1146/annurev-arplant-050312-120048 . ПМИД 23451788 . S2CID 727983 .
- ^ Хаммер, К. (1984). «Синдром одомашнивания». Культурное растение (на немецком языке). 32 :11–34. дои : 10.1007/bf02098682 . S2CID 42389667 .
- ^ Уилкинс, Адам С.; Рэнгэм, Ричард В.; Fitch, В. Текумсе (июль 2014 г.). «Синдром одомашнивания» млекопитающих: единое объяснение, основанное на поведении и генетике клеток нервного гребня» (PDF) . Генетика . 197 (3): 795–808. дои : 10.1534/genetics.114.165423 . ПМК 4096361 . ПМИД 25024034 .
- ^ Заллуа, Пьер А.; Матису-Смит, Элизабет (6 января 2017 г.). «Картирование послеледниковой экспансии: заселение Юго-Западной Азии» . Научные отчеты . 7 : 40338. Бибкод : 2017NatSR...740338P . дои : 10.1038/srep40338 . ISSN 2045-2322 . ПМК 5216412 . ПМИД 28059138 .
- ^ МакХьюго, Джиллиан П.; Довер, Майкл Дж.; МакХью, Дэвид Э. (2 декабря 2019 г.). «Раскрытие происхождения и биологии домашних животных с помощью древней ДНК и палеогеномики» . БМК Биология . 17 (1): 98. дои : 10.1186/s12915-019-0724-7 . ПМК 6889691 . ПМИД 31791340 .
- ↑ Перейти обратно: Перейти обратно: а б с д и ж г час Пуруганан, Майкл Д.; Фуллер, Дориан К. (1 февраля 2009 г.). «Природа отбора при одомашнивании растений» (PDF) . Природа . 457 (7231): 843–848. Бибкод : 2009Natur.457..843P . дои : 10.1038/nature07895 . ISSN 0028-0836 . ПМИД 19212403 . S2CID 205216444 .
- ↑ Перейти обратно: Перейти обратно: а б Зохари, Хопф и Вайс 2012 , с. 139.
- ↑ Перейти обратно: Перейти обратно: а б с Лоулер, Эндрю; Адлер, Джерри (июнь 2012 г.). «Как курица завоевала мир» . Журнал Смитсоновского института (июнь 2012 г.).
- ^ Фуллер, Дориан К.; Уиллкокс, Джордж; Аллаби, Робин Г. (2011). «Культивирование и одомашнивание имели множественное происхождение: аргументы против основной гипотезы происхождения сельского хозяйства на Ближнем Востоке». Мировая археология . 43 (4): 628–652. дои : 10.1080/00438243.2011.624747 . S2CID 56437102 .
- ↑ Перейти обратно: Перейти обратно: а б Зедер, Мелинда А. (2006). «Археологические подходы к документированию приручения животных». В Зедере, Массачусетс; Брэдли, генеральный директор; Эмшвиллер, Э.; Смит, Б.Д. (ред.). Документирование одомашнивания: новые генетические и археологические парадигмы . Беркли: Издательство Калифорнийского университета . стр. 209–227.
- ^ Галиберт, Фрэнсис; Киньон, Паскаль; Хитте, Кристоф; Андре, Кэтрин (1 марта 2011 г.). «На пути к пониманию истории эволюции и одомашнивания собак» . Comptes Rendus Biologies . По следам одомашнивания, миграций и вторжений в сельское хозяйство. 334 (3): 190–196. дои : 10.1016/j.crvi.2010.12.011 . ПМИД 21377613 .
- ^ Снир, Айнит; Надель, Дэни; Громан-Ярославский, Ирис; Меламед, Йоэль; Штернберг, Марсело; Бар-Йосеф, Офер; Вайс, Эхуд (22 июля 2015 г.). «Происхождение культивирования и протосорняков задолго до неолитического земледелия» . ПЛОС ОДИН . 10 (7): e0131422. дои : 10.1371/journal.pone.0131422 . ПМК 4511808 . ПМИД 26200895 .
- ^ Дрисколл, Карлос (2009). «Укрощение кошки. Генетические и археологические находки указывают на то, что дикие кошки стали домашними кошками раньше и в другом месте, чем считалось ранее» . Научный американец . 300 (6): 68–75. Бибкод : 2009SciAm.300f..68D . doi : 10.1038/scientificamerican0609-68 (неактивен 14 апреля 2024 г.). ПМЦ 5790555 . ПМИД 19485091 .
{{cite journal}}: CS1 maint: DOI неактивен по состоянию на апрель 2024 г. ( ссылка ) - ^ Дрисколл, Карлос А.; Менотти-Рэймонд, Мэрилин; Рока, Альфред Л.; Хьюп, Карстен; Джонсон, Уоррен Э.; и др. (27 июля 2007 г.). «Ближневосточное происхождение приручения кошек» . Наука . 317 (5837): 519–523. Бибкод : 2007Sci...317..519D . дои : 10.1126/science.1139518 . ПМК 5612713 . ПМИД 17600185 .
- ^ Даймонд, Джаред ; Беллвуд, П. (2003). «Фермеры и их языки: первые расширения». Наука . 300 (5619): 597–603. Бибкод : 2003Sci...300..597D . CiteSeerX 10.1.1.1013.4523 . дои : 10.1126/science.1078208 . ПМИД 12714734 . S2CID 13350469 .
- ↑ Перейти обратно: Перейти обратно: а б с д Франц, Лоран А.Ф.; Брэдли, Дэниел Г.; Ларсон, Грегер; Орландо, Людовик (2020). «Одомашнивание животных в эпоху древней геномики» . Обзоры природы Генетика . 21 (8): 449–460. дои : 10.1038/s41576-020-0225-0 . ПМИД 32265525 . S2CID 214809393 .
- ^ Дарвин, Чарльз (1868). Изменение животных и растений при одомашнивании . Лондон: Джон Мюррей. OCLC 156100686 .
- ^ Даймонд 2005 , с. 130.
- ^ Ларсон, Г.; Пиперно, ДР; Аллаби, Р.Г.; Пуруганан, доктор медицины; Андерссон, Л.; и др. (2014). «Современные перспективы и будущее исследований одомашнивания» . Труды Национальной академии наук . 111 (17): 6139–6146. Бибкод : 2014PNAS..111.6139L . дои : 10.1073/pnas.1323964111 . ПМК 4035915 . ПМИД 24757054 .
- ↑ Перейти обратно: Перейти обратно: а б Дуст, АН; Люкенс, Л.; Олсен, КМ; Мауро-Эррера, М.; Мейер, А.; Роджерс, К. (2014). «За пределами одного гена: как эпистаз и воздействие генов на окружающую среду влияют на одомашнивание сельскохозяйственных культур» . Труды Национальной академии наук . 111 (17): 6178–6183. Бибкод : 2014PNAS..111.6178D . дои : 10.1073/pnas.1308940110 . ПМК 4035984 . ПМИД 24753598 .
- ↑ Перейти обратно: Перейти обратно: а б с д и Ларсон, Грегер; Фуллер, Дориан К. (2014). «Эволюция приручения животных» (PDF) . Ежегодный обзор экологии, эволюции и систематики . 45 : 115–136. doi : 10.1146/annurev-ecolsys-110512-135813 . S2CID 56381833 . Архивировано из оригинала (PDF) 13 мая 2019 года . Проверено 19 января 2016 г.
- ^ Мейер, Рэйчел С.; Пуруганан, Майкл Д. (2013). «Эволюция видов сельскохозяйственных культур: генетика одомашнивания и диверсификации». Обзоры природы Генетика . 14 (12): 840–852. дои : 10.1038/nrg3605 . ПМИД 24240513 . S2CID 529535 .
- ↑ Перейти обратно: Перейти обратно: а б с д и ж г час я дж к л Зедер, Мелинда А. (2012). «Одомашнивание животных». Журнал антропологических исследований . 68 (2): 161–190. дои : 10.3998/jar.0521004.0068.201 . S2CID 85348232 .
- ^ Хейл, Э.Б. (1969). «Одомашнивание и эволюция поведения». В Хафезе, ESE (ред.). Поведение домашних животных (2-е изд.). Лондон: Байер, Тиндалл и Касселл. стр. 22–42.
- ^ Прайс, Эдвард О. (1984). «Поведенческие аспекты приручения животных». Ежеквартальный обзор биологии . 59 (1): 1–32. дои : 10.1086/413673 . JSTOR 2827868 . S2CID 83908518 .
- ^ Прайс, Эдвард О. (2002). Одомашнивание и поведение животных (PDF) . Уоллингфорд, Великобритания: Издательство CABI. Архивировано из оригинала (PDF) 17 мая 2017 года . Проверено 29 февраля 2016 г.
- ↑ Перейти обратно: Перейти обратно: а б Маршалл, Ф. (2013). «Оценка роли направленного размножения и потока генов в приручении животных» . Труды Национальной академии наук Соединенных Штатов Америки . 111 (17): 6153–6158. Бибкод : 2014PNAS..111.6153M . дои : 10.1073/pnas.1312984110 . ПМК 4035985 . ПМИД 24753599 .
- ^ Ларсон, Грегер (2013). «Взгляд на приручение животных с точки зрения популяционной генетики» (PDF) . Тенденции в генетике . 29 (4): 197–205. дои : 10.1016/j.tig.2013.01.003 . ПМИД 23415592 . Архивировано из оригинала (PDF) 8 июня 2019 года . Проверено 2 января 2016 г.
- ^ Серпелл, Дж.; Даффи, Д. (2014). «Породы собак и их поведение». В Горовице, Александра (ред.). Познание и поведение домашней собаки . Берлин / Гейдельберг: Springer.
- ^ Кейган, Алекс; Бласс, Торстен (2016). «Идентификация геномных вариантов, предположительно являющихся целью отбора во время приручения собаки» . Эволюционная биология BMC . 16 (1): 10. Бибкод : 2016BMCEE..16...10C . дои : 10.1186/s12862-015-0579-7 . ПМК 4710014 . ПМИД 26754411 .
- ↑ Перейти обратно: Перейти обратно: а б Ларсон, Грегер (2012). «Переосмысление приручения собак путем интеграции генетики, археологии и биогеографии» . Труды Национальной академии наук Соединенных Штатов Америки . 109 (23): 8878–8883. Бибкод : 2012PNAS..109.8878L . дои : 10.1073/pnas.1203005109 . ПМЦ 3384140 . ПМИД 22615366 .
- ^ Перри, Анджела (2016). «Волк в собачьей шкуре: первоначальное приручение собаки и плейстоценовая вариация волка». Журнал археологической науки . 68 : 1–4. Бибкод : 2016JArSc..68....1P . дои : 10.1016/j.jas.2016.02.003 .
- ^ Франц, Л. (2015). «Доказательства долгосрочного потока генов и отбора во время одомашнивания на основе анализа геномов диких и домашних свиней Евразии». Природная генетика . 47 (10): 1141–1148. дои : 10.1038/ng.3394 . ПМИД 26323058 . S2CID 205350534 .
- ^ Пенниси, Э. (2015). «Укрощение свиньи приняло несколько диких оборотов». Наука . дои : 10.1126/science.aad1692 .
- ^ Скок, Дж. (2023a). «Гипотеза о приручении паразитов» . Сельскохозяйственная наука . 20 (1): 1–7. дои : 10.18690/agricsci.20.1.1 .
- ^ Скок, Дж. (2023b). «Дополнение к «Гипотезе о приручении, опосредованном паразитами» » . ОСФ . дои : 10.31219/osf.io/f92aj .
- ^ «Птица» . Американское наследие: Словарь английского языка (4-е изд.). Компания Хоутон Миффлин. 2009.
- ^ «Наша история» . Общество птицеводов Америки . Проверено 3 декабря 2023 г.
- ^ Блехман, Эндрю (2007). Голуби – захватывающая сага о самой почитаемой и порицаемой птице в мире . Университет Квинсленда Пресс. ISBN 9780702236419 .
- ^ Айзен, Марсело А.; Хардер, Лоуренс Д. (2009). «Глобальные запасы одомашненных медоносных пчел растут медленнее, чем сельскохозяйственный спрос на опыление» . Современная биология . 19 (11): 915–918. дои : 10.1016/j.cub.2009.03.071 . ПМИД 19427214 . S2CID 12353259 .
- ^ Поттс, Саймон Г.; и др. (2010). «Глобальное сокращение количества опылителей: тенденции, последствия и движущие силы» . Тенденции в экологии и эволюции . 25 (6): 345–353. CiteSeerX 10.1.1.693.292 . дои : 10.1016/j.tree.2010.01.007 . ПМИД 20188434 .
- ^ Гон III, Сэмюэл М.; Прайс, Эдвард О. (октябрь 1984 г.). «Одомашнивание беспозвоночных: поведенческие соображения» (PDF) . Бионаука . 34 (9): 575–579. дои : 10.2307/1309600 . JSTOR 1309600 .
- ^ Вайс, Э., Кислев, М.Э., Симхони, О. и Надель, Д. Мелкозернистые дикие травы как основной продукт питания на 23000-летнем участке Экономической ботаники Охало II 58: s125-s134.
- ^ Запрет 2002 года .
- ^ Зохари, Хопф и Вайс 2012 .
- ^ Харрис, Дэвид Р. (1996). Происхождение и распространение земледелия и скотоводства в Евразии . Лондон: Издательство Университетского колледжа Лондона. стр. 142–158. ISBN 9781857285376 .
- ^ «Новые археоботанические данные для изучения истоков сельского хозяйства в Китае», Чжицзюнь Чжао, Текущая антропология, том. 52, № С4, (октябрь 2011 г.), стр. С295-С306.
- ^ Карни, Джудит (2009). В тени рабства . Беркли и Лос-Анджелес, Калифорния: Издательство Калифорнийского университета. п. 16. ISBN 9780520269965 .
- ↑ Перейти обратно: Перейти обратно: а б с д Диллехей, Том Д.; Россен, Джек; Андрес, Томас С.; Уильямс, Дэвид Э. (29 июня 2007 г.). «Докерамическое использование арахиса, тыквы и хлопка в Северном Перу». Наука . 316 (5833). Американская ассоциация развития науки (AAAS): 1890–1893 гг. Бибкод : 2007Sci...316.1890D . дои : 10.1126/science.1141395 . ISSN 0036-8075 . ПМИД 17600214 . S2CID 43033764 .
- ^ Смит, Брюс Д. (15 августа 2006 г.). «Восточная часть Северной Америки как независимый центр одомашнивания растений» . Труды Национальной академии наук Соединенных Штатов Америки . 103 (33): 12223–12228. Бибкод : 2006PNAS..10312223S . дои : 10.1073/pnas.0604335103 . ПМЦ 1567861 . ПМИД 16894156 .
- ^ Пиперно, Долорес Р. (октябрь 2011 г.). «Истоки выращивания и одомашнивания растений в тропиках Нового Света: закономерности, процесс и новые разработки». Современная антропология . 52 (С4): С453–С470. дои : 10.1086/659998 . S2CID 83061925 .
Центральная долина реки Бальсас в Мексике, предполагаемая колыбель происхождения кукурузы ... распространилась в нижнюю часть Центральной Америки к 7600 году до нашей эры.
- ^ Спунер, Дэвид М.; Маклин, Карен; Рамзи, Гэвин; Во, Робби; Брайан, Гленн Дж. (29 сентября 2005 г.). «Единственное одомашнивание картофеля на основе генотипирования полиморфизма длины мультилокусного амплифицированного фрагмента» . Труды Национальной академии наук . 102 (41): 14694–14699. Бибкод : 2005PNAS..10214694S . дои : 10.1073/pnas.0507400102 . ПМЦ 1253605 . ПМИД 16203994 .
- ^ Олсен, Кеннет М.; Шааль, Барбара А. (11 мая 1999 г.). «Свидетельства о происхождении маниоки: филогеография Manihot esculenta» . Труды Национальной академии наук . 96 (10): 5586–5591. Бибкод : 1999PNAS...96.5586O . дои : 10.1073/pnas.96.10.5586 . ISSN 0027-8424 . ПМК 21904 . ПМИД 10318928 .
- ^ Гросс, Бриана Л.; Олсен, Кеннет М. (2010). «Генетические перспективы одомашнивания сельскохозяйственных культур» . Тенденции в науке о растениях . 15 (9): 529–537. doi : 10.1016/j.tplants.2010.05.008 . ISSN 1360-1385 . ПМЦ 2939243 . ПМИД 20541451 .
- ^ Хьюз, Аойф; Оливейра, HR; Фрэдгли, Н.; Корк, Ф.; Кокрам, Дж.; Дунан, Дж. Х.; Нибау, К. (14 марта 2019 г.). «Анализ признаков μCT выявляет морфометрические различия между одомашненными мелкозернистыми злаками умеренного пояса и их дикими родственниками» . Заводской журнал . 99 (1): 98–111. дои : 10.1111/tpj.14312 . ПМК 6618119 . PMID 30868647 .
- ^ Снир, Айнит и Вайс, Эхуд, 2014. Новый морфометрический метод дифференциации дикого и домашнего ячменя посредством внутрирахисных измерений, Journal of Archaeological Science 44: 69-75, https://doi.org/10.1016/j.jas.2014.01 .014
- ↑ Перейти обратно: Перейти обратно: а б с д и ж г час я Ленсер, Тереза; Тайсен, Гюнтер (2013). «Молекулярные механизмы, участвующие в конвергентном одомашнивании сельскохозяйственных культур». Тенденции в науке о растениях . 18 (12). Cell Press : 704–714. doi : 10.1016/j.tplants.2013.08.007 . ISSN 1360-1385 . ПМИД 24035234 .
- ^ Августин, Мануэль; Кузен-Милло, Эдвард (2020). Род Цитрусовые Издательство Вудхед. стр. 100-1 219–244. ISBN 978-0-12-812163-4 .
- ^ Перье, Ксавье; Бакри, Фредерик; Каррель, Франсуаза; и др. (2009). «Объединение биологических подходов, чтобы пролить свет на эволюцию съедобных бананов» . Этноботаника: исследования и приложения . 7 : 199–216. дои : 10.17348/era.7.0.199-216 . hdl : 10125/12515 . Архивировано из оригинала 16 ноября 2019 года . Проверено 27 октября 2019 г.
- ↑ Перейти обратно: Перейти обратно: а б с Милла, Рубен; Осборн, Колин П.; Тюркотт, Мартин М.; Виолле, Сирил (2015). «Одомашнивание растений через экологическую призму». Тенденции в экологии и эволюции . 30 (8). Эльзевир Б.В.: 463–469. дои : 10.1016/j.tree.2015.06.006 . ISSN 0169-5347 . ПМИД 26138385 .
- ^ Ву, Юе; Го, Тинтин; Му, Ци; и др. (декабрь 2019 г.). «Аллелохимикаты призваны сбалансировать конкурирующие виды в африканских агроэкосистемах» . Природные растения . 5 (12): 1229–1236. дои : 10.1038/s41477-019-0563-0 . ISSN 2055-0278 . ПМИД 31792396 . S2CID 208539527 .
- ↑ Перейти обратно: Перейти обратно: а б Кантар, Майкл Б.; Тил, Катрин Э.; Дорн, Кевин М.; и др. (29 апреля 2016 г.). «Многолетние зерновые и масличные культуры» . Ежегодный обзор биологии растений . 67 (1). Годовые обзоры : 703–729. doi : 10.1146/annurev-arplant-043015-112311 . ISSN 1543-5008 . ПМИД 26789233 .
- ^ Тюркотт, Мартин М.; Терли, Нэш Э.; Джонсон, Марк ТиДжей (18 июля 2014 г.). «Влияние одомашнивания на устойчивость к двум травоядным универсальным животным в 29 независимых событиях одомашнивания» . Новый фитолог . 204 (3). Уайли: 671–681. дои : 10.1111/nph.12935 . ISSN 0028-646X . ПМИД 25039644 .
- ↑ Перейти обратно: Перейти обратно: а б Головнина К.А.; Глушков С.А.; Блинов А.Г.; Майоров, В.И.; Адкисон, ЛР; Гончаров Н.П. (12 февраля 2007 г.). «Молекулярная филогения рода Triticum L». Систематика и эволюция растений . 264 (3–4). Спрингер: 195–216. Бибкод : 2007PSyEv.264..195Г . дои : 10.1007/s00606-006-0478-x . ISSN 0378-2697 . S2CID 39102602 .
- ^ Гептс, Пол (2004). «Одомашнивание сельскохозяйственных культур как долгосрочный селекционный эксперимент» (PDF) . Обзоры селекции растений . 2. 24 .
- ^ Шао, Г.; Тан, А.; Тан, ЮК; Луо, Дж.; Цзяо, Джорджия; Ву, Дж.Л.; Ху, PS (апрель 2011 г.). «Новая делеционная мутация ароматного гена и разработка трех молекулярных маркеров аромата риса». Селекция растений . 2. 130 (2): 172–176. дои : 10.1111/j.1439-0523.2009.01764.x .
- ^ Консорциум по секвенированию генома картофеля (июль 2011 г.). «Секвенирование генома и анализ клубнеплодов картофеля» . Природа . 475 (7355): 189–195. дои : 10.1038/nature10158 . ПМИД 21743474 .
- ↑ Перейти обратно: Перейти обратно: а б Ганн, Би; Бодуэн, Люк; Олсен, Кеннет М. (2011). «Независимое происхождение культивируемого кокоса (Cocos nucifera L.) в тропиках Старого Света» . ПЛОС ОДИН . 6 (6): e21143. Бибкод : 2011PLoSO...621143G . дои : 10.1371/journal.pone.0021143 . ПМК 3120816 . ПМИД 21731660 .
- ↑ Перейти обратно: Перейти обратно: а б Зедер, Мелинда; Эмшвиллер, Ева; Смит, Брюс Д.; Брэдли, Дэниел Г. (март 2006 г.). «Документирование одомашнивания: пересечение генетики и археологии» . Тенденции в генетике . 22 (3): 139–55. дои : 10.1016/j.tig.2006.01.007 . ПМИД 16458995 . Проверено 28 ноября 2011 г.
- ^ Матч, Лесли А.; Янг, Дж. Питер В. (2004). «Разнообразие и специфичность Rhizobium leguminosarum biovar viciae на дикорастущих и культивируемых бобовых» . Молекулярная экология . 13 (8): 2435–2444. Бибкод : 2004MolEc..13.2435M . дои : 10.1111/j.1365-294X.2004.02259.x . ISSN 1365-294X . ПМИД 15245415 . S2CID 1123490 .
- ^ Кирс, Э. Тоби; Хаттон, Марк Г.; Денисон, Р. Форд (22 декабря 2007 г.). «Человеческий отбор и ослабление защиты бобовых от неэффективных ризобий» . Труды Королевского общества B: Биологические науки . 274 (1629): 3119–3126. дои : 10.1098/rspb.2007.1187 . ПМК 2293947 . ПМИД 17939985 .
- ↑ Перейти обратно: Перейти обратно: а б с Абдельфаттах, Ахмед; Тэк, Эйко Дж. М.; Вассерман, Биргит; Лю, Цзя; Берг, Габриэле; и др. (2021). «Доказательства коэволюции хозяина и микробиома яблока» . Новый фитолог . 234 (6): 2088–2100. дои : 10.1111/nph.17820 . ISSN 1469-8137 . ПМЦ 9299473 . ПМИД 34823272 . S2CID 244661193 .
- ^ Коулман-Дерр, Девин; Дегаренн, Дамарис; Фонсека-Гарсия, Читлали; Гросс, Стивен; Клингенпил, Скотт; и др. (2016). «Растительный компартмент и биогеография влияют на состав микробиома культивируемых и местных видов агавы» . Новый фитолог . 209 (2): 798–811. дои : 10.1111/nph.13697 . ISSN 1469-8137 . ПМК 5057366 . ПМИД 26467257 .
- ^ Буффо, Мари-Лара; Пуарье, Мари-Андре; Мюллер, Дэниел; и др. (2014). «Корневой микробиом связан с эволюцией растений-хозяев кукурузы и других злаков» . Экологическая микробиология . 16 (9): 2804–2814. Бибкод : 2014EnvMi..16.2804B . дои : 10.1111/1462-2920.12442 . ISSN 1462-2920 . ПМИД 24588973 .
- ^ Абдуллаева, Юлдузхон; Амбика Манираджан, Биной; Хонермейер, Бернд; и др. (1 июля 2021 г.). «Одомашнивание влияет на состав, разнообразие и совместное возникновение микробиоты семян зерновых» . Журнал перспективных исследований . 31 : 75–86. дои : 10.1016/j.jare.2020.12.008 . ISSN 2090-1232 . ПМК 8240117 . ПМИД 34194833 .
- ^ Фавела, Алонсо; О. Бон, Мартин; Д. Кент, Анджела (август 2021 г.). «Хронопоследовательность зародышевой плазмы кукурузы показывает, что история селекции сельскохозяйственных культур влияет на пополнение микробиома ризосферы» . Журнал ISME . 15 (8): 2454–2464. Бибкод : 2021ISMEJ..15.2454F . дои : 10.1038/s41396-021-00923-z . ISSN 1751-7370 . ПМЦ 8319409 . ПМИД 33692487 .
- ^ « Agaricus bisporus : пуговица» . MushroomExpert.com . Проверено 25 марта 2017 г.
- ^ Легра, Жан-Люк; Мердиноглу, Дидье; Корнюэ, Жан-Мари; Карст, Фрэнсис (2007). «Хлеб, пиво и вино: разнообразие Saccharomyces cerevisiae отражает историю человечества». Молекулярная экология . 16 (10): 2091–2102. Бибкод : 2007MolEc..16.2091L . дои : 10.1111/j.1365-294X.2007.03266.x . ПМИД 17498234 . S2CID 13157807 .
- ^ «Работа Pfizer над пенициллином для Второй мировой войны становится национальной исторической вехой в области химии» . Американское химическое общество . 12 июня 2008 г.
- ^ Берри, Р.Дж. (1969). «Генетические последствия одомашнивания животных». В Уко, Питер Дж.; Димблби, Г.В. (ред.). Одомашнивание и эксплуатация растений и животных . Чикаго: Олдин. стр. 207–217.
- ^ Даймонд 2005 , стр. 198, 213.
- ^ Кальдараро, Никколо Лео (2012). «Эволюционные аспекты предотвращения болезней: роль болезней в развитии сложного общества» . Серия рабочих документов SSRN . дои : 10.2139/ssrn.2001098 . ISSN 1556-5068 . S2CID 87639702 .
- ^ Бойден, Стивен Викерс (1992). «Книги ES&T». Экологические науки и технологии . 8 (приложение 173): 665. Бибкод : 1992EnST...26..665. . дои : 10.1021/es00028a604 .
- ^ Букчин, Мюррей (2022). Философия социальной экологии (3-е изд.). АК Пресс. стр. 85–87. ISBN 9781849354400 .
- ^ Ниберт, Дэвид (2013). Угнетение животных и насилие над людьми: осквернение дома, капитализм и глобальный конфликт . Издательство Колумбийского университета . стр. 1–5. ISBN 9780231151894 .
- ^ Бойвин, Николь Л.; Зедер, Мелинда А.; Фуллер, Дориан К.; и др. (2016). «Экологические последствия строительства человеческой ниши: изучение долгосрочного антропогенного формирования глобального распределения видов» . Труды Национальной академии наук . 113 (23): 6388–6396. Бибкод : 2016PNAS..113.6388B . дои : 10.1073/pnas.1525200113 . ПМЦ 4988612 . ПМИД 27274046 .
- ^ Флинт-Гарсия, Шерри А. (4 сентября 2013 г.). «Генетика и последствия одомашнивания сельскохозяйственных культур» . Журнал сельскохозяйственной и пищевой химии . 61 (35): 8267–8276. дои : 10.1021/jf305511d . ISSN 0021-8561 . ПМИД 23718780 .
- ↑ Перейти обратно: Перейти обратно: а б Браун, Теренс А. (апрель 2019 г.). «Является ли проблема приручения мифом?» . Природные растения . 5 (4): 337–338. дои : 10.1038/s41477-019-0404-1 . ПМИД 30962526 . S2CID 102353100 .
- ↑ Перейти обратно: Перейти обратно: а б с Аллаби, Робин Г.; Уэр, Розелин Л.; Кистлер, Логан (январь 2019 г.). «Переоценка проблем приручения на основе археогеномных данных» . Эволюционные приложения . 12 (1): 29–37. Бибкод : 2019EvApp..12...29A . дои : 10.1111/eva.12680 . ПМК 6304682 . ПМИД 30622633 .
- ^ Шеперд, Лара Д.; Ланге, Питер Дж. де; Кокс, Саймон; Макленачан, Патрисия А.; Роскруге, Ник Р.; Локхарт, Питер Дж. (24 марта 2016 г.). «Свидетельства сильного узкого места при одомашнивании недавно выращенного эндемичного новозеландского корнеплода Arthropodium cirratum (Asparagaceae)» . ПЛОС ОДИН . 11 (3): e0152455. Бибкод : 2016PLoSO..1152455S . дои : 10.1371/journal.pone.0152455 . ПМЦ 4806853 . ПМИД 27011209 .
- ^ Кассон, Мэтью Т.; Викерт, Кристен Л.; Стаудер, Кэмерон М.; Масиас, Энджи М.; Бергер, Мэтью С.; Симмонс, Д. Раберн; Коротко, Дилан П.Г.; ДеВалланс, Дэвид Б.; Хулкр, Иржи (октябрь 2016 г.). «Мутуализм с агрессивным разлагающим древесину Flavodon ambrosius (Polyporales) способствует расширению ниши и созданию общественной социальной структуры у жуков Ambrosiophilus ambrosia» . Грибная экология . 23 : 86–96. дои : 10.1016/j.funeco.2016.07.002 .
- ^ Рейнджер, Кристофер М.; Бидерманн, Питер Х.В.; Пхунтумарт, Випапорн; Белигала, Гаятри У.; Гош, Сатьяки; Палмквист, Дебра Э.; Мюллер, Роберт; Барнетт, Дженни; Шульц, Питер Б.; Рединг, Майкл Э.; Бенц, Дж. Филипп (24 апреля 2018 г.). «Отбор симбионтов с помощью алкоголя приносит пользу жукам-амброзиям, выращивающим грибы» . Труды Национальной академии наук . 115 (17): 4447–4452. Бибкод : 2018PNAS..115.4447R . дои : 10.1073/pnas.1716852115 . ПМЦ 5924889 . ПМИД 29632193 .
- ^ Хюлькр, Иржи; Стелински, Лукаш Л. (31 января 2017 г.). «Симбиоз амброзии: от эволюционной экологии к практическому управлению» . Ежегодный обзор энтомологии . 62 : 285–303. doi : 10.1146/annurev-ento-031616-035105 . ПМИД 27860522 .
- ^ Шик, Джонатан З.; Гомес, Эрнесто Б.; Коой, Пепейн В.; Сантос, Хуан К.; Всисло, Уильям Т.; Бумсма, Якобус Дж. (6 сентября 2016 г.). «Питание опосредует выражение конфликта между сортом и фермером у муравья, выращивающего грибы» . Труды Национальной академии наук . 113 (36): 10121–10126. дои : 10.1073/pnas.1606128113 . ISSN 0027-8424 . ПМК 5018747 . ПМИД 27551065 .
- ^ Токуда, Г.; Ватанабэ, Х.; Мацумото, Т.; Нода, Х. (1997). «Переваривание целлюлозы у высших термитов, питающихся древесиной, Nasutitermes takasagoensis (Shiraki): распределение целлюлаз и свойства эндо-бета-1,4-глюканазы» . Зоологическая наука . 14 (1): 83–93. дои : 10.2108/zsj.14.83 . ПМИД 9200983 . S2CID 2877588 .
- ^ Икеда-Оцубо, В.; Брюн, А. (2009). «Совместное видообразование жгутиконосцев кишечника термитов и их бактериальных эндосимбионтов: виды Trichonympha и Candidatus Endomicrobium trichonymphae». Молекулярная экология . 18 (2): 332–342. Бибкод : 2009MolEc..18..332I . дои : 10.1111/j.1365-294X.2008.04029.x . ПМИД 19192183 . S2CID 28048145 .
- ^ Слейтор, М. (1992). «Переваривание целлюлозы у термитов и тараканов: какую роль играют симбионты?». Сравнительная биохимия и физиология Б . 103 (4): 775–784. дои : 10.1016/0305-0491(92)90194-В .
Источники [ править ]
- Бэннинг, Эдвард Б. (2002). «Акерамический неолит». В Перегрине, Питер Н.; Эмбер, Мелвин (ред.). Энциклопедия предыстории, том 8: Южная и Юго-Западная Азия . Kluwer Academic / Plenum. Издательство
- Даймонд, Джаред (2005) [1997]. Оружие, микробы и сталь: Краткая история каждого человека за последние 13 000 лет . Лондон: Чатто и Виндус . ISBN 9780099302780 .
- Хэйр, Брайан ; Вудс, Ванесса (август 2020 г.). «Выживание самых дружелюбных: естественный отбор по гиперсоциальным признакам позволил высшим видам Земли превзойти неандертальцев и других конкурентов». Научный американец . Том. 323, нет. 2. С. 58–63.
- Хайден, Брайан (2003). «Были ли роскошные продукты первыми одомашненными? Этноархеологические перспективы Юго-Восточной Азии». Мировая археология . 34 (3): 458–469. дои : 10.1080/0043824021000026459a . S2CID 162526285 .
- Марчиняк, Аркадиуш (2005). Помещение животных в эпоху неолита: социальная зооархеология доисторических земледельческих сообществ . Лондон: UCL Press . ISBN 9781844720927 .
- Зохари, Дэниел ; Хопф, Мария; Вайс, Эхуд (2012). Одомашнивание растений в Старом Свете (4-е изд.). Оксфорд: Издательство Оксфордского университета . doi : 10.1093/acprof:osobl/9780199549061.001.0001 . ISBN 9780199549061 .
Внешние ссылки [ править ]
- Инвентаризация диких родственников сельскохозяйственных культур и анализ пробелов : надежный источник информации о том, где и что сохранять ex-situ для генофондов сельскохозяйственных культур, имеющих глобальное значение.
- Обсуждение приручения животных с Джаредом Даймондом
- Первоначальное одомашнивание Cucurbita pepo в Америке 10 000 лет назад
- Схема приручения крупного рогатого скота. Архивировано 19 декабря 2010 г. в Wayback Machine.
- Основная тема «одомашнивание»: бесплатные полнотекстовые статьи (более 100 плюс обзоры) в Национальной медицинской библиотеке.