Хеликс (брюхоногие моллюски)
| спираль Временной диапазон: | |
|---|---|
 | |
| Helix поматия | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Моллюска |
| Сорт: | Брюхоногие моллюски |
| Подкласс: | Гетеробранхия |
| Заказ: | Стиломматофора |
| Семья: | Хелициды |
| Подсемейство: | Хелицины |
| Племя: | Хеличини |
| Род: | спираль Линней , 1758 г. |
| Типовой вид | |
| Helix поматия | |
| Синонимы | |
см. текст | |
Helix — род , дышащих воздухом, крупных наземных улиток обитающих в западной Палеарктике и характеризующихся шаровидной раковиной. [1] [2]
Это типовой род семейства Helicidae и один из родов животных, описанных Карлом Линнеем. [3] на заре зоологической номенклатуры .
Представители этого рода впервые появились в летописи окаменелостей в миоцене . [4]
К хорошо известным видам относятся Helix pomatia (римская улитка, бургундская улитка или съедобная улитка) и Helix lucorum (турецкая улитка). Cornu aspersum (садовая улитка), хотя внешне похожа и долгое время классифицировалась как член Helix (как « Helix aspersa »), не имеет близкого родства с Helix. [5] [6] принадлежит к другому племени Helicinae и . [7]
Таксономия
[ редактировать ]В » Линнея 10 - м издании « Системы природы , которое положило начало зоологической номенклатуре , родовое название Helix использовалось для множества наземных (например, Zonites algirus ), пресноводных (например, Lymnaea stagnalis ) и морских (например, Fossarus ambiguus ) видов. брюхоногие моллюски . Более поздние авторы ограничили использование этого названия видами стиломматофоранов с уплощенной до шаровидной раковиной, включая зонитиды и другие группы. В течение 1800-х годов у Helix было описано несколько тысяч видов в Европе и за рубежом . [8] [9] К началу 1900-х годов род был разделен на множество отдельных родов, в результате чего остались только виды, тесно связанные с его типовым видом Helix pomatia в роду . Однако из-за ранее широкой концепции рода Helix является частью исходной комбинации ( базионима ) многих названий брюхоногих моллюсков, и в Helix до сих пор описано множество номинальных таксонов , родовое положение которых остается нерешенным ( taxa inquirenda ), [10] хотя они явно не относятся ни к одному из видов Helix в его нынешнем смысле.
С 2000-х годов Helix стал объектом обширных молекулярно-филогенетических исследований и таксономических пересмотров. [1] [11] [2] [12] [13] Это привело к исключению нескольких видов, в первую очередь садовой улитки , и включению других ( H. ceratina , H. nicaeensis ). Мальтзанелла , долгое время считавшаяся подродом Helix , также была формально исключена из рода, [14] но это сестринская группа Helix .
В настоящее время признаны два подрода: [2] [15]
- Хеликс (Helix) Линней , 1758 г.
- Хеликс (Пеласга), Гессен , 1908 г.
- Спираль ( Aegaeohelix ) Корабек и Хаусдорф, 2023 г.
Описание
[ редактировать ]Helix включает крупные виды наземных улиток с диаметром раковины 2–6 см. Раковина имеет форму шаровидной или конической формы, с пятью более темными полосами, которые могут уменьшаться или сливаться вместе. Шаровидная оболочка отличает Helix от большинства родственных родов (трибы Helicini ), за исключением Maltzanella и Lindholmia . Поверхность имеет структуру из мелких поперечных ребер, развитых в разной степени, а также могут иметься очень мелкие спиральные бороздки. Скорлупа никогда не бывает молотковой. Цвет стопы варьируется. Он может быть серым, коричневым, черным или розовым; задняя часть стопы у некоторых видов темная.
Раковины видов Helix правосторонние. Левистральные особи очень редки, но изредка встречаются (например, H. pomatia , [16] Х. Фессалика [17] и H. lutescens [18] ).
Символы половой системы использовались для определения рода и его подродов. В отличие от Корню , половой член Helix содержит два сосочка с центральным отверстием. По-видимому, существует тенденция к укорочению дивертикулов копулятивной сумки и эппифаллуса, но по этим признакам наблюдается перекрытие с родственными родами. Слизистые железы, примыкающие к дротиковому мешку, обычно сильно разветвлены.
Распределение
[ редактировать ]Helix — западнопалеарктический род. Видовое разнообразие сосредоточено на Балканах и в Анатолии , наибольшее филогенетическое разнообразие наблюдается в Греции . Естественные западные пределы распространения проходят через материковую Францию ( Helix pomatia ), Корсику ( Helix ceratina ) и Алжир ( Helix melanostoma ). На севере естественное распространение H. pomatia достигает центральной Германии и южных окраин Северо-Европейской равнины . Самые южные виды обитают в Северной Африке ( H. melanostoma , H. pronuba ) и южном Леванте ( H. engaddensis ). Восточные пределы достигаются в Западном Иране и Иракском Курдистане ( H. salomonica ) и на Кавказе ( H. lucorum ); H. thessalica проникает через Украину , по крайней мере, до западной границы России . [1] [2]
Генетика
[ редактировать ]Размер гаплоидного генома оценивался примерно в 4 Гб ( значение C 4 пг) с содержанием GC ~ 42%. [19] [20] но неясно, какой вид изучался из-за несоответствия указанного вида и происхождения образцов. Гаплоидное — число хромосом 27 (исследованные виды — H. lucorum , H. buchii , H. pomatia , H. gussoneana и H. straminea ). [21] [22] [23] [24] У H. pomatia все хромосомы имеют срединные или субмедиальные центромеры . [25] Небольшие нештатные хромосомы были обнаружены у H. pomatia из Англии . [25]
митохондриальный геном H. pomatia (длиной около 14 070 п.н.). Доступен [26] [27]
Половая система
[ редактировать ]
Строение половой системы во многом соответствует таковому у других Helicidae . Его анатомия и функции подробно изучены у H. pomatia . [29] [30] [31]
Как и все стиломматофоры, Helix улитки — гермафродиты . Сперматозоиды и яйцеклетки производятся в общей гонаде — овотестисе (гермафродитной железе), которая встроена в гепатопанкреас (пищеварительную железу) вблизи вершины скорлупы. Гаметы транспортируются через гермафродитный проток (овотестисный проток) к комплексу оплодотворительной сумки-сперматеки (карфур), расположенному в основании белковой железы. [32] В этом органе чужеродная сперма (сперма другого человека) хранится в семенных мешочках (receptacula seminis), а оплодотворение яйцеклетки происходит в сумке для оплодотворения. Белковая железа обеспечивает питание развивающихся оплодотворенных яиц, и ее размер сильно варьируется в зависимости от стадии репродуктивного цикла. Собственная сперма и оплодотворенные яйцеклетки улитки транспортируются специализированными участками семявывода (спермовидная канавка и матка), которые дистально разделяются на мужскую ( семявыводящий проток ) и женскую (свободный яйцевод) части половой системы. [32]
Гениталии самца состоят из трубки, служащей образованию сперматофора и передаче его в женские части партнера по спариванию. Пенис – самая дистальная и мускулистая часть. Сперматофор формируется в эпифаллусе (между семявыносящим протоком и половым членом) и жгутике (продолжении эпифаллуса проксимальнее отверстия семявыносящего протока); последний образует хвост сперматофора. Во время совокупления пенис выворачивается (как рукав, вывернутый наизнанку). [29] и при этом вводится во влагалище. Втягивающая мышца прикрепляется к эпифаллусу и втягивает мужские гениталии после совокупления.
Женская часть состоит из влагалища (иногда называемого копулятивным каналом). [33] ) и копулятивную сумку (гаметолитическую железу) с ее ножкой и обычно дивертикулом ножки. Влагалище служит для транспортировки чужеродных сперматофоров и яйцеклеток. Бурса прикрепляется к влагалищу тонкой ножкой (обозначая границу между влагалищем и свободным яйцеводом). Стебель в большинстве случаев несет дивертикул - слепую трубку, в которую вмещается передняя часть сперматофора, если он присутствует. Предполагается, что дивертикул является остатком семенного протока, который первоначально транспортировал чужеродную сперму в сумку для оплодотворения. [34] Сперма покидает хвост сперматофора и мигрирует в яйцевод, а затем в сумку для оплодотворения; подавляющее большинство сперматозоидов не выходит таким путем и переваривается в бурсе. [31] [32] У Helix наблюдается тенденция к уменьшению дивертикула, у некоторых видов он может отсутствовать. [2]
Аппарат-дротик, хотя и расположен во влагалище, функционально также является частью мужских гениталий. Он состоит из одного мышечного стреловидного мешка и двух слизистых желез (пальцевых или добавочных желез) по бокам. Слизистые железы у основания разветвлены; количество ветвей варьируется в зависимости от вида Helix . Одиночный дротик арагонитовый , прямой или слабоизогнутый, с четырьмя лопастями (лопастями) по длине и короной у основания. [35] [5] отсутствует дротик У Helix salomonica . [2] Слизистые железы вырабатывают слизь, которая покрывает дротик во время выстрела и тем самым впрыскивается в тело партнера, где вызывает укорочение дивертикула и перистальтические движения ножки бурсы, которые помогают чужеродным сперматозоидам избежать лизиса в бурсе. [33]
Вся половая система формируется из единого трубчатого впячивания эктодермы . В процессе развития вдоль внутреннего пространства трубы образуются складки, которые в конечном итоге полностью отделяют сначала семявыносящий проток и половой член, а затем и копулятивную сумку. [36] Мужская и женская части открываются в общее предсердие и генитальную пору, расположенную вентрально позади правого зрительного щупальца.
редкая тератологическая зарегистрирована Из Германии особь с парными мужскими гениталиями (пенис, эпифаллус, жгутик) . [37]
Воспроизведение
[ редактировать ]
Аспекты размножения изучались в основном у H. pomatia с ограниченной информацией о других видах.
было описано несколько раз Брачное поведение H. pomatia . [29] [38] На начальном этапе две улитки поднимают ноги, прижимают подошвы друг к другу и касаются щупалец и ротового аппарата друг друга. Это занимает 15–30 мин. Некоторое время спустя происходит стрельба дротиками, хотя многие спаривания происходят без любовного дротика использования . Непосредственно перед выстрелом слизистые железы выделяют беловатый секрет, содержащий гормоны, способствующие образованию соединения, улучшающего сохранность чужеродных сперматозоидов у принимающего человека. [39] Затем, снова после паузы, наступает совокупление, которому обычно предшествуют несколько неудачных попыток, при которых взаимное введение пенисов не достигается и половые органы частично втягиваются обратно в тело. Наконец, обе особи одновременно вставляют пенисы в женское отверстие друг друга. В пределах ок. Через 4–7 мин формируется и перемещается сперматофор, после чего улитка отрывается и втягивает вывернутые половые органы. Однако полное получение сперматофора занимает еще 2–3 часа, в течение которых улитки остаются частично втянутыми и малоподвижными. [40] [41]
Сообщалось, что во время совокупления у H. pomatia обычно переносится только один сперматофор , поэтому при каждом спаривании одно животное действует как самец, а одно — как самка. Согласно этому отчету, в основном яйца откладывают старшие улитки, а молодые функционируют как самцы. [40]
У H. pomatia спаривание происходит преимущественно с мая по июнь, но часто продолжается более спорадически до осени. [30] [42] Однако, поскольку активность зависит от климатических условий, сроки спаривания и откладки яиц у некоторых других видов различаются. [43]
У H. pomatia улитки обычно совокупляются с несколькими самками. [30] [42] Полученная чужеродная сперма может храниться более года до оплодотворения. [31] Яйца откладываются в камеру, выкопанную родителем в почве, через 4–6 дней после спаривания. [40] Яйца образуются только по мере строительства гнезда. [44] Как и у других легочных , яйца богаты галактогеном, вырабатываемым белковой железой. Яичная скорлупа частично минерализована, с кристаллами карбоната кальция в гибкой оболочке. [32] Размер сцепления в литературе указан в пределах 3–93. [45] Вылупление происходит примерно через 25–26 (диапазон 18–31). [42] ) дней после откладки яиц, но улитки остаются в гнезде еще 8–10 дней. [40] Особь может отложить более одной кладки за сезон. [42] У других видов размер кладки может быть другим. Средний размер кладки H. lucorum аналогичен таковому у H. pomatia . [46] в то время как диапазон, указанный для H. albescens (меньший размер тела, чем у H. pomatia , с более крупными яйцами), составляет всего 7–22 яйца на кладку. [45]
Морфология сперматозоидов соответствует основному образцу, известному для « легочных ». Митохондрии сливаются и образуют сплошную оболочку вокруг жгутика . Большую часть митохондриальных производных представляет собой белковая паракристаллическая структура, в которой имеется канал, заполненный гликогеном . Канал проходит спирально вдоль жгутика, образуя так называемую гликогеновую спираль. имеется только одна слабо закрученная спираль гликогена В Helix . [47] [48]
История жизни
[ редактировать ]У H. pomatia половая зрелость достигается после 2–4 (–6) зимовок, т. е. в возрасте 2–4 (–6) лет, но она различается как в зависимости от локалитета, так и между улитками из одной кладки. [40] [49] [50] Взрослые улитки перестают расти и вокруг отверстия образуют утолщенную губу. Однако иногда улитки спариваются уже незадолго до образования губы. Продолжительность жизни H. pomatia в дикой природе может достигать 30 лет. [50] Однако в дикой природе они в основном живут всего 5–9 лет. [49] [50] Возраст особей H. pomatia можно оценить, подсчитав перерывы в росте на раковине и количество слоев, отложившихся на губе края апертуры. [49] [51] [50]
Данные по другим видам очень ограничены. Helix lucorum в Греции достигает зрелости в 3 года. [46] Мелкий вид H. ceratina достигает зрелости через 1,5–2 года и живет 4–5 лет. [52]
Продолжительность жизненного цикла зависит от условий окружающей среды. Время от вылупления до первой откладки яиц может быть сокращено у H. pomatia всего до 12–13 месяцев при оптимальных условиях содержания в неволе, главным образом за счет пропуска спячки. [42]
Экология и поведение
[ редактировать ]Виды Helix обитают в самых разных средах обитания и в самых разных климатических режимах. Некоторые виды встречаются исключительно в открытых известняковых скалистых местах обитания ( H. secernenda ), тогда как другие переносят кислую почву (например, H. pelagonesica ) или обитают преимущественно в лесах (например, H. thessalica ). Представители рода встречаются от тропических лесов умеренного пояса ( H. buchii ) до полузасушливых регионов ( Helix pronuba ). [53] [1]
Пищевые предпочтения для большинства видов неизвестны. Helix pomatia питается живыми растениями. [42] [49] а также мертвые растительные остатки. [54] Наблюдения показали предпочтение некоторых конкретных видов растений. [49] и избегание других. [42] Крапива Urtica dioica является предпочтительным пищевым растением, особенно у молодых особей. [54]
Активность H. pomatia происходит преимущественно ночью, особенно у молоди. [42] У H. pomatia наблюдалось самонаведение . Улитки рассеиваются в течение сезона, но к концу сезона, как правило, возвращаются в места спячки. [55] [49] [56] В течение сезона у них может быть территория, где они проживают и откуда они совершают экскурсии к известным местам нагула и, при необходимости, пытаются найти новые. [56] Также было замечено, что улитки могут мигрировать в определенные места для откладки яиц. [49] и агрегат для спаривания. [57] Улитки способны найти дорогу обратно на расстоянии десятков метров. [55] Активное расселение в течение жизни может привести к смещению более чем на 200 м. [56]
Некоторые виды (например, H. pomatia , H. salomonica ) образуют толстую, кальцинированную эпифрагму , которая закрывает отверстие раковины во время спячки или летнего периода . [30] [43] За эпифрагмой внутри раковины следует несколько дополнительных оболочек из засохшей слизи. [58] Когда животное выходит из состояния покоя, оно сбрасывает известковую надфрагму задней частью стопы. [ нужна ссылка ] Спячка и зимовка проходят в почве, куда улитки зарываются ногой. [43] [30] Виды Helix различаются по тому, где они проводят более короткие периоды бездействия: некоторые склонны оставаться на поверхности почвы, некоторые прячутся в почве (в частности, виды подрода Pelasga [43] ), а некоторые виды часто забираются на растительность (стволы деревьев, кустарники). [ нужна ссылка ]
Спячка или зимовка могут занять значительную часть года. Например, H. salomonica в Ширнаке , Турция, в течение года наблюдений проводила в среднем 85 дней в спячке и 165 дней в состоянии покоя. [43]
Биотические взаимодействия
[ редактировать ]Хищники и паразиты
[ редактировать ]Несколько видов птиц охотятся на виды Helix . Это наблюдалось у Phasianus colchicus , Burhinus oedicemus , Coracias garrulus , Lanius excubitor , врановых и, возможно, у других птиц. [59] [60] Млекопитающие охотятся на Хеликс тоже . Известные хищники включают ежа , крота , землеройку , грызунов ( Rattus , Apodemus ) и кабана. [61] [60] поедает Медленный червь моллюсков, в том числе и Helix . [62] Улитки Helix также подвергаются нападению различных жуков и мух . Жуки-хищники относятся к семействам Carabidae и Lampyridae . [49] [50] [63] Хищничество со стороны птиц, мелких млекопитающих и жуков в основном поражает молодь. [49] Личинки мух нескольких семейств нападают на улиток Helix и могут убить даже взрослых особей ( Phoridae , Muscidae , Sarcophagidae , Sciomyzidae ). [64]
Хорошо известным факультативным паразитом наземных улиток, в том числе Helix , является нематода Phasmarhabiditis hermaphrodita . [65] Паразитический клещ Riccardoella limacum встречается у видов Helix . [66] [67]
Кинетопластид pomatia Cryptobia helicis обитает в копулятричной сумке Helix . [31] Инфузория Tetrahymena limacis также была обнаружена из Helix . [68]
Бактериальные заболевания брюхоногих моллюсков, включая Helix, известны, но эта область недостаточно изучена. [69]
Влияние на другие виды и окружающую среду
[ редактировать ]Некоторые виды пчел строят свои гнезда внутри пустых Helix раковин (например, Rhodanthidium semptemdentatum ). [ нужна ссылка ]
Использование человеком
[ редактировать ]
Некоторые виды, прежде всего H. pomatia и H. lucorum , собираются для потребления человеком . [ нужна ссылка ] Кулинарное использование насчитывает несколько тысячелетий и было подтверждено для нескольких видов всего ареала рода.
Остатки мезолитических раковин, датированные 9370 ± 80 и 8110 ± 90 некалиброванными C-14 годами назад и свидетельствующие о сборе, были задокументированы для H. pomatella в Абруццо , Италия. [70] Helix salomonica потреблялась в Загросе в больших количествах во время докерамического неолита , имеются свидетельства потребления примерно с 12 000 г. до н.э. [71] [72] В Северной Африке остатки раковин каспийской культуры, содержащие большое количество H. melanostoma . были обнаружены [73]
Древние римляне собирали улиток в пищу и даже держали их в загонах, как описано Марком Теренцием Варроном (в De Re Rustica и повторено Плинием Старшим ). Считается, что римская улитка H. pomatia была завезена в Англию римлянами. [74]
Сохранение
[ редактировать ]Большинство видов, включенных в Красный список МСОП, относятся к категории «вызывающие наименьшее опасение». [75] Один из видов, H. ceratina , находится под угрозой исчезновения , и известное в настоящее время распространение ограничено очень небольшой территорией вблизи аэропорта Аяччо . [76] [52]
В прошлом сбор дикой H. pomatia в пищу приводил к опасениям чрезмерной эксплуатации и введению законодательной защиты в ряде стран. [77]
Список существующих видов
[ редактировать ]| Научное название | Красный список МСОП статус | Распределение | Картина |
|---|---|---|---|
| Белая спираль Россмасслер, 1839 г. | ЛК МСОП | Азербайджан, Армения, Грузия, Россия, Украина [1] |  |
| Хеликс анктостома Мартенс, 1874 г. | Турция (Нур-Даги и окрестности), Сирия (северо-запад) | ||
| Хеликс Анке Корабек и Хаусдорф, 2023 г. | Турция (северо-западная Анатолия) [15] | ||
| Helix antiochiensis Кобельт, 1896 г. | Сирия |  | |
| Хеликс Асемнис Бургинья, 1860 г. | ЛК МСОП | 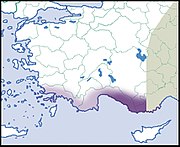 |  |
| Helix Borealis Муссон, 1859 г. | ДД МСОП | Греция |  |
| Хеликс бучии (Дюбуа де Монпере, 1840 г.) | Турция (северо-восток), Грузия, Армения (север) |  | |
| Хеликс Калабрика Вестерлунд, 1876 г. | |||
| Хеликс цератина Шаттлворт, 1843 г. | ЧР МСОП |  |  |
| Спираль окружена ОФ Мюллер, 1774 г. | ЛК МСОП | Сирия, Турция (вероятно, завезена частично), Кипр (вероятно завезена), Греция ( восточная часть Эгейского моря , вероятно завезена), Хорватия (введена), Словения (введена), Италия (введена) [1] [78] |  |
| Спальни Хеликс Кобельт, 1898 г. | ЛК МСОП |  |  |
| Helix engaddensis Бургинья, 1852 г. | Израиль, Западный Берег, Иордания, Ливан [79] |  | |
| Helix Fathallae Вот, 1901 год. | |||
| Хеликс фигурка Россмасслер, 1839 г. | ЛК МСОП |  |  |
| Хеликс годециана Кобельт, 1878 г. | В МСОП | ||
| Helix gussoneana Л. Пфайффер, 1848 г. | |||
| Хеликс Казуиниана Паллари, 1939 год. | |||
| Спираль связана ОФ Мюллер, 1774 г. | ДД МСОП |  |  |
| Helix lucorum Линней, 1758 г. |  |  | |
| Хеликс лютесценс Россмасслер, 1837 г. | ЛК МСОП |  |  |
| Хеликс меланостома Драпарно, 1801 г. | Тунис, Алжир, Франция (введена), Испания (введена) |  | |
| Хеликс миллети Кобельт, 1906 г. |  | ||
| Helix nicaeensis А. Ферюссак, 1821 г. | Турция |  | |
| Ядро спирали Муссон, 1854 г. | ЛК МСОП | Ливан, Сирия, Кипр, Турция (вероятно, частично завезена) [1] | |
| Хеликс пачья Бургинья, 1860 г. | |||
| Хеликс патетика Муссон, 1854 г. | |||
| Helix pelagonesica (Ролл, 1898 г.) | |||
| Хеликс филибинская Россмасслер, 1839 г. | ЛК МСОП | Греция, Северная Македония, Болгария (юго-запад), Албания (у озера Преспа) |  |
| Хеликс помачелла Муссон, 1854 г. | ЛК МСОП |  | |
| Хеликс помателла Кобельт, 1876 г. | |||
| Helix поматия Линней, 1758 г. | ЛК МСОП |  | |
| Хеликс невеста Вестерлунд, 1879 г. | Египет, Ливия, Тунис (юг), Греция (введены Крит и другие острова) |  | |
| Хеликс Саломоника Вот, 1899 год. | Турция (юго-восток), Иран (запад), Ирак (Курдистан) | ||
| Helix schlaeflii Муссон, 1859 г. | Албания (центральная и южная), Греция, Северная Македония (Галичица) [2] [1] | ||
| Спираль, которую нужно отделить Россмасслер, 1847 г. | ЛК МСОП | Хорватия, Босния и Герцеговина, Черногория, Албания (север) [2] [1] |  |
| Хеликс страминеа Разбойники, 1825 г. | ЛК МСОП | Албания, Северная Македония (восточная), Италия |  |
| Хеликс Фессалика О. Беттгер, 1886 г. | ЛК МСОП |  | |
| Хеликс Валентини Кобельт, 1891 г. | В МСОП | | |
| Бишопс Хеликс (Кобельт, 1898 г.) | ЛК МСОП | Сербия, Черногория |  |
Ископаемая запись
[ редактировать ]несколько вымерших таксонов Helix Описано :
- Helix jasonis Mayer, 1856 (Украина: Севастополь. Миоцен: Тортон [80] )
- Helix pseudoligata Sinzov, 1897 (Украина. Миоцен: сармат. [80] )
- Helix kadolskyi Neubauer & Harzhauser, 2023 ( номен. ноябрь для Helix toulai Kojumdgieva, 1969 ) (Болгария: Балчик. Миоцен: средний тортон [81] )
- Helix barbeyana Де Стефани в книге Де Стефани и др., 1891 г.
- Helix krejcii Wenz в Krejci-Graf & Wenz, 1926 г.
- Хеликс мразечи Севастос, 1922 г.
- Helix sublutescens Wenz в Крейчи и Венце, 1926 г.
- Helix maeotica Steklov, 1966 (Россия: Чечня: р. Десны. Миоцен: мэот. [82] =? Тортониан [83] )
- Helix varnensis Toula, 1892 (Болгария: Варна. Миоцен: Сармат)
- Helix lucorum supralevantina Wenz, 1942 (Румыния. Плиоцен).
Некоторые современные виды известны из четвертичных отложений. Наиболее изученным видом в этом отношении является H. pomatia , окаменелости которого были использованы для документирования самых ранних послеледниковых находок в Центральной Европе . [84] Самая ранняя запись в Чехии была датирована непосредственно радиоуглеродом 10 120–9 690 лет назад (но, вероятно, на несколько сотен лет моложе); [85] окаменелости предположительно старше 9 402–9 027 лет назад [86] или 9 403–9 003 п.н. [87] были найдены в Баден-Вюртемберге, Германия. Такие записи документируют скорость, с которой виды Helix могут расширять свои ареалы естественным путем.
Летопись окаменелостей четвертичных наземных улиток в более южных частях Европы скудна, но некоторые записи о Helix все же существуют. Фигурка Helix, датируемая примерно 16 000 лет назад, была обнаружена на греческом острове Антикитера . [88] Helix Borealis, Сообщалось о раковинах датируемых 8 000–27 000 лет назад, с другого острова, Гавдос . [89]
Другие записи взяты из археологических источников.
Филогения
[ редактировать ]Филогенетические взаимоотношения Helix внутренние взаимоотношения внутри рода изучены пока лишь с использованием частичных последовательностей митохондриальных с родственными родами, а также генов и ядерной кластера генов рРНК . [90] [1]
Показанная кладограмма основана на филогенетическом анализе данных митохондриальных последовательностей. [91] [11] [78] [1]
| Мальцанелла | |||||||||||||||||||||||||||||||
| спираль |
| ||||||||||||||||||||||||||||||
Синонимы
[ редактировать ]Следующие таксоны уровня рода считаются синонимами Helix :
- Каллунея Скаддер, 1882 г.
- Кохлеа да Коста, 1778 г.
- Коэнатория Хелд, 1838 г.
- Кунула Паллари , 1936 год.
- Глишрус С. Штудер, 1820 г.
- Helicites W. Martin, 1809 (Установлено для окаменелостей Helix , чтобы отличить их от современных представителей этого таксона. Недействительно, доступно только для целей Принципа омонимии (Статья 20))
- Геликогена А. Ферюссак, 1821 г.
- Мегастома Скаддер, 1882 г.
- Нагелеа П. Гессен, 1918 год.
- Пахифалл П. Гессен, 1918 г.
- Пентатения А. Шмидта, 1855 г. (младший объективный синоним)
- Фисоспира Беттгер , 1914 г.
- Поматия Бек, 1837 г.
- Поматиана Фагот, 1903 г.
- Поматиэлла Паллари, 1909 г.
- Псевдофигулина П. Гессен, 1917 г.
- Рододерма П. Гессен, 1918 г.
- Тахеопсис Беттгер, 1909 г.
- Таммузия Паллари, 1939 год.
- Тирренский П. Гессен, 1918 г.
Ссылки
[ редактировать ]- ^ Перейти обратно: а б с д и ж г час я дж к Корабек, Ондржей; Юржичкова, Люси; Петрусек, Адам (январь 2022 г.). «Разнообразие племени наземных улиток Helicini (Gastropoda: Stylommatophora: Helicidae): где мы находимся после 20 лет секвенирования митохондриальных маркеров?» . Разнообразие . 14 (1): 24. дои : 10.3390/d14010024 . ISSN 1424-2818 .
- ^ Перейти обратно: а б с д и ж г час Нойберт, Эйке (2014). «Пересмотр Helix Linnaeus, 1758 в ареале его распространения в восточном Средиземноморье и перевод Helix godetiana Kobelt, 1878 на Maltzanella Hesse, 1917 (Gastropoda, Pulmonata, Helicidae)». Вклад в естествознание . 26 : 1–200.
- ^ Линней, К. (1758). Система природы по трем царствам природы, по классам, отрядам, родам, видам, с признаками, различиями, синонимами, местами. Том I. Издание десятое, реформированное . Стокгольм п. 768
- ^ Харцхаузер, Матиас; Нойбауэр, Томас А. (1 июня 2021 г.). «Обзор фаун наземных улиток европейского кайнозоя – состав, разнообразие и круговорот» . Обзоры наук о Земле . 217 : 103610. Бибкод : 2021ESRv..21703610H . doi : 10.1016/j.earscirev.2021.103610 . ISSN 0012-8252 . S2CID 233835790 .
- ^ Перейти обратно: а б Коэн, Йорис М; Шуленбург, Хинрих (2005). «Стрельба дротиками: коэволюция и контрадаптация улиток-гермафродитов» . Эволюционная биология BMC . 5 (1): 25. дои : 10.1186/1471-2148-5-25 . ПМЦ 1080126 . ПМИД 15799778 .
- ^ Джусти, Ф.; Манганелли, Г.; Шембри, П.Дж. (1995). Неморские моллюски Мальтийских островов . Турин: Региональный музей естественных наук.
- ^ Найбер, Марко Т; Корабек, Ондржей; Глаубрехт, Матиас; Хаусдорф, Бернхард (11 апреля 2022 г.). «Неправильно истолкованная дизъюнкция: филогенетические отношения североафриканской наземной улитки Gyrostomella (Gastropoda: Stylommatophora: Helicidae)» . Зоологический журнал Линнеевского общества . 194 (4): 1236–1251. doi : 10.1093/zoolinnean/zlab059 . ISSN 0024-4082 .
- ^ Вестерлунд, Калифорния, 1889. Фауна внутренних конхилий, обитающих в Палеарктическом регионе (Европа, Кавказ, Сибирь, Туран, Персия, Курдистан, Армения, Месопотамия, Малая Азия, Сирия, Аравия, Египет, Триполи, Тунис, Алжир и Марокко) . II. Генерал Хеликс. - С. 1-473, 1-31, 1-8. Берлин. (Фридлендер).
- ^ Пфайффер, Л. и Клессин, С. 1881. Номенклатура живых улиток, содержащая названия всех известных сегодня родов и видов этого семейства, упорядоченных по природному родству. - стр. 1-617 Касселли. (Фишер).
- ^ Билер Р., Буше П., Гофас С., Маршалл Б., Розенберг Г., Ла Перна Р., Нойбауэр Т.А., Сартори А.Ф., Шнайдер С., Вос С., Тер Поортен Дж.Дж., Тейлор Дж., Дейкстра Х., Финн Дж., Банк Р., Нойберт Э., Морецсон Ф., Фабер М., Хуарт Р., Пиктон Б., Гарсиа-Альварес О., ред. (2023). « Геликс Линней, 1758 год» . МоллюскаБаза . Всемирный регистр морских видов . Проверено 29 апреля 2023 г.
- ^ Перейти обратно: а б Фиорентино, В.; Манганелли, Г.; Джусти, Ф.; Кетмайер, В. (1 мая 2016 г.). «Недавнее расширение и реликтовое выживание: филогеография рода наземных улиток Helix (Mollusca, Gastropoda) с юга на север Европы» . Молекулярная филогенетика и эволюция . 98 : 358–372. Бибкод : 2016МОЛПЭ..98..358Ф . дои : 10.1016/j.ympev.2016.02.017 . ISSN 1055-7903 . ПМИД 26926944 .
- ^ Корабек, Ондржей; Петрусек, Адам; Нойберт, Эйке; Юржичкова, Люси (2015). «Молекулярная филогения рода Helix (Pulmonata: Helicidae)» . Зоологика Скрипта . 44 (3): 263–280. дои : 10.1111/zsc.12101 . S2CID 84673484 .
- ^ Манганелли, Джузеппе; Саломоне, Никола; Джусти, Фолько (2005). «Молекулярный подход к филогенетическим взаимоотношениям западнопалеарктических Helicoidea (Gastropoda: Stylommatophora)» . Биологический журнал Линнеевского общества . 85 (4): 501–512. дои : 10.1111/j.1095-8312.2005.00514.x .
- ^ Нойберт, Эйке; Банк, Рууд А. (2008). «Заметки о видах Caucasotachea C. Boettger 1909 и Lindholmia P. Hesse 1919, с аннотациями к Helicidae (Gastropoda: Stylommatophora: Helicidae)». Архив Моллюскенконде . 135 (1): 101–132. дои : 10.1127/arch.moll/0003-9284/135/101-132 .
- ^ Перейти обратно: а б Корабек, Ондржей; Хаусдорф, Бернхард (13 декабря 2023 г.). «Раскрытие двойной Helix escherichi, с описанием Helix ankae sp. nov. и обсуждением Maltzanella и подродов Helix (Gastropoda: Helicidae)» . Archiv für Molluskenkunde Международный журнал малакологии . 152 (2): 239–256. дои : 10.1127/arch.moll/152/239-256 . ISSN 1869-0963 .
- ^ Блошик, Ежи; Калиновский, Томаш; Ксёншкевич, Зофия; Шибяк, Кристина (4 марта 2015 г.). «Новые данные о левых и лестничных раковинах римской улитки Helix pomatia Linnaeus, 1758 в Польше» . Фолиа Малакологическая . 23 (1): 47–50. дои : 10.12657/folmal.023.004 .
- ^ Корабек, Ондржей; Юржичкова, Люси; Петрусек, Адам (2016). «Полосатая улитка – новый вид для фауны Европы и Чехии» (PDF) . Живой . 2016 (1): 31–34.
- ^ Коралевска-Батура, Э. (1997). «Свернутый влево образец Helix lutescens Rossmässler, 1837 (Gastropoda: Stylommatophora: Helicidae)». Малакологические трактаты . 18 (2): 233–239.
- ^ Виноградов, Александр Е (1 февраля 2000 г.). «Большие геномы для пионеров суши моллюсков» . Геном . 43 (1): 211–212. дои : 10.1139/g99-063 . ISSN 0831-2796 . ПМИД 10701134 .
- ^ «Геномы на дереве» . goat.genomehubs.org . Проверено 11 мая 2023 г.
- ^ Бахтадзе Н.Г.; Чакветадзе, Нидерланды; Мумладзе, ЖЖ; Бахтадзе, Г.И.; Цхадая, Э.А. (2016). «Кариологические данные наземных моллюсков (Mollusca: Gastropoda: Pulmonata) Грузии» (PDF) . Труды Института зоологии (Государственный университет Ильи, Тбилиси) . 25 : 23–27.
- ^ Перро, Жан-Луи; Перро, Макс (1937). «Хромосомная формула Helix pomatia» . Швейцарский журнал зоологии . 44 : 203–209.
- ^ Петраччиоли, Аньезе; Кровато, Паоло; Гуарино, Фабио Мария; Меццасалма, Марчелло; Сегодня Гаэтано; Пикариелло, Орфей; Майо, Никола (2021). «Хромосомное разнообразие и эволюция у Helicoide a (Gastropoda: Stylommatophora): синтез оригинальных и литературных данных» . Животные . 11 (9): 2551. дои : 10.3390/ani11092551 . ISSN 2076-2615 . ПМЦ 8470273 . ПМИД 34573517 .
- ^ Петраччиоли, А.; Ниеро, И.; Каранденте, Ф.; Кровато, П.; де ВИКО, Г.; Сегодня Г.; Пикариелло, OLA; Тарди, Э.; Вильетти, С.; Гуарино, FM; Майо, Н. (1 января 2021 г.). «Helix straminea Briganti, 1825 в Италии (Gastropoda: Pulmonata): таксономическая история, морфология, биология, распространение и филогения» . Европейский зоологический журнал . 88 (1): 390–416. дои : 10.1080/24750263.2021.1892217 . S2CID 234221660 .
- ^ Перейти обратно: а б Эванс, HJ (1960). «Сверхкомплектные хромосомы в диких популяциях улитки Helix pomatia L». Наследственность . 15 (2): 129–138. дои : 10.1038/hdy.1960.70 . ISSN 1365-2540 . S2CID 36907929 .
- ^ Корабек, Ондржей; Петрусек, Адам; Ровацос, Михаил (5 марта 2019 г.). «Полный митогеном Helix pomatia и базальная филогения Helicinae (Gastropoda, Stylommatophora, Helicidae)» . ZooKeys (827): 19–30. Бибкод : 2019ZooK..827...19K . дои : 10.3897/zookeys.827.33057 . ISSN 1313-2970 . ПМК 6472302 . ПМИД 31114424 .
- ^ Гроенберг, DSJ; Дуйм, Э. (2 января 2019 г.). «Полный митогеном римской улитки Helix pomatia Linnaeus 1758 (Stylommatophora: Helicidae)» . Митохондриальная ДНК Часть B. 4 (1): 1494–1495. дои : 10.1080/23802359.2019.1601512 . ISSN 2380-2359 .
- ^ Коэн, Йорис М; Шуленбург, Хинрих (2005). «Стрельба дротиками: коэволюция и контрадаптация улиток-гермафродитов» . Эволюционная биология BMC . 5 (1): 25. дои : 10.1186/1471-2148-5-25 . ПМЦ 1080126 . ПМИД 15799778 .
- ^ Перейти обратно: а б с Майзенхаймер, Йоханнес (1907). «Биология, морфология и физиология процесса спаривания и яйцекладки Helix pomatia» . Зоологические ежегодники. Кафедра систематики, географии и биологии животных . 25 : 461–502, пл. 16-18.
- ^ Перейти обратно: а б с д и Майзенхаймер, Йоханнес (1912). Римская улитка Helix pomatia L. Leipzig: Издательство Dr. Вернер Клинхардт.
- ^ Перейти обратно: а б с д Линд, Ганс (1973). «Функциональное значение сперматофора и судьба сперматозоидов в половых путях Helix pomatia (Gastropoda: Stylommatophora)» . Журнал зоологии . 169 (1): 39–64. дои : 10.1111/j.1469-7998.1973.tb04652.x . ISSN 0952-8369 .
- ^ Перейти обратно: а б с д Томпа, А.С. (1984). «Наземные улитки (Стиломматофора)». В Томпе, AS; Вердонк, Нью-Хэмпшир; ван ден Биггелаар, JAM (ред.). Моллюска. Том 7. Репродукция . Лондон: Академическая пресса. стр. 47–140. ISBN 0-12-751407-4 .
- ^ Перейти обратно: а б Лоди, Моника; Коэн, Йорис М. (2016). «О специфичности действия продуктов добавочных желез, переносимых любовным дротиком наземных улиток» . Эволюционная биология BMC . 16 (1): 104. Бибкод : 2016BMCEE..16..104L . дои : 10.1186/s12862-016-0672-6 . ISSN 1471-2148 . ПМК 4866404 . ПМИД 27178200 .
- ^ Хохпёхлер, Ф.; Котбауэр, Х. (1979). «Триаулы у гелицидов (Gastropoda) О филогенетическом значении дивертикула копулятивной сумки» . Журнал зоологической систематики и эволюционных исследований . 17 (4): 281–285. дои : 10.1111/j.1439-0469.1979.tb00711.x .
- ^ Хант, С. (1979). «Строение и состав дротика любовного (гипсобелума) Helix pomatia» . Ткань и клетка . 11 (1): 51–61. дои : 10.1016/0040-8166(79)90005-3 . ПМИД 451995 .
- ^ Хохпёхлер, Франц (1979). «Сравнительные исследования развития полового аппарата Stylommatophora (Gastropoda)». Зоологический вестник . 6/5 : 289–306.
- ^ Эшворт, Дж. Х. (1907). «Особь Helix pomatia с парными мужскими органами» . Труды Королевского общества Эдинбурга . 27 : 312–331, 1 табл. дои : 10.1017/S037016460001751X .
- ^ Джеппесен, Лейф Лау (1976). «Контроль брачного поведения Helix pomatia L. (Gastropoda: Pulmonata)» . Поведение животных . 24 (2): 275–290. дои : 10.1016/S0003-3472(76)80036-X . S2CID 53188212 .
- ^ Лоди, Моника; Коэн, Йорис М. (25 августа 2015 г.). «Любовные дротики наземных улиток: интеграция физиологии, морфологии и поведения» . Журнал исследований моллюсков : eyv046. дои : 10.1093/mollus/eyv046 . ISSN 0260-1230 .
- ^ Перейти обратно: а б с д и Килиас, Рудольф (1985). Римская улитка (на немецком языке). Лютерштадт Виттенберг: А. Цимсен.
- ^ Дункан, CJ (1975). «Репродукция». Во Фреттере, Вера; Пик, Дж. (ред.). Легочные. Том 1. Функциональная анатомия и физиология . Лондон: Академическая пресса. стр. 309–365. ISBN 0-12-267501-0 .
- ^ Перейти обратно: а б с д и ж г час Тишлер, Вольфганг (1973). «К биологии и экологии римской улитки ( Helix pomatia )». Фаунистически-экологические связи . 4 : 283-298.
- ^ Перейти обратно: а б с д и Экин, Ихсан (18 июня 2024 г.). «Helix salomonica Nägele, 1899 (Gastropoda: Helicidae): годовые циклы активности в естественной среде обитания на юго-востоке Турции» . Исследования моллюсков : 1–13. дои : 10.1080/13235818.2024.2364372 . ISSN 1323-5818 .
- ^ Перро, Ж.-Л. (1938). «Гнездостроение и яйцекладка Helix pomatia» . Швейцарский журнал зоологии . 45 : 221–235.
- ^ Перейти обратно: а б Крамаренко С.С. (2013). «Анализ репродуктивных особенностей легочных моллюсков: мини-обзор». Рутеница . 23 : 115–125.
- ^ Перейти обратно: а б Стайку, А.; Лазариду-Димитриаду, М.; Фармакис, Н. (1988). «Аспекты жизненного цикла, динамики численности, роста и вторичной продукции съедобной улитки Helix lucorum Linnaeus, 1758 (Gastropoda, Pulmonata) в Греции» . Журнал исследований моллюсков . 54 (2): 139–155. дои : 10.1093/моллюс/54.2.139 . ISSN 0260-1230 .
- ^ Хили, Дж. М.; Джеймисон, фоновая музыка (1989). «Ультраструктурное исследование сперматозоидов Helix aspersa и Helix pomatia (Gastropoda, Pulmonata)» . Журнал исследований моллюсков . 55 (3): 389–404. дои : 10.1093/моллюс/55.3.389 . ISSN 0260-1230 .
- ^ Томпсон, Т.Э. (1973). «Эутинейран и другие сперматозоиды моллюсков» . Малакология . 14 : 167–206.
- ^ Перейти обратно: а б с д и ж г час я Поллард, Э. (1975). «Аспекты экологии Helix pomatia L.». Журнал экологии животных . 44 (1): 305–329. Бибкод : 1975JAnEc..44..305P . дои : 10.2307/3865 . JSTOR 3865 .
- ^ Перейти обратно: а б с д и Фолкнер, Герхард (1984). «Проект Баварской римской улитки (Исследования Helix pomatia L.)». Объявления Немецкого малакозоологического общества . 37 : 182-197.
- ^ Поллард, Э.; Кук, А.С.; Уэлч, Дж. М. (1977). «Использование особенностей раковины в определении возраста молодых и взрослых римских улиток Helix pomatia ». Журнал зоологии, Лондон . 183 (2): 269–279. дои : 10.1111/j.1469-7998.1977.tb04186.x .
- ^ Перейти обратно: а б Камю, Луиза; Поли, Педро; Деложер, Мишель-Жан; Дреано, Стефан; Кучера, Ксавье; Натали, Кристина; Гиллер, Энни (22 мая 2023 г.). «Неожиданное и пространственно структурированное генетическое разнообразие реликтовой популяции находящейся под угрозой исчезновения корсиканской наземной улитки Tyrrenaria ceratina» . Сохраняющая генетика . 24 (5): 661–672. Бибкод : 2023ConG...24..661C . дои : 10.1007/s10592-023-01535-0 . ISSN 1566-0621 . S2CID 258855822 .
- ^ Корабек, Ондржей (2014). «Улитки – автостопщик не только по Средиземноморью» (PDF) . Живой . 2014 (4): 171–175.
- ^ Перейти обратно: а б Тлусте, Клаудия; Биркхофер, Клаус (14 апреля 2023 г.). «Римская улитка (Gastropoda: Helicidae) не является универсальным травоядным животным, но в еде предпочитает Urtica dioica и растительный мусор» . Журнал естественной истории . 57 (13–16): 758–770. дои : 10.1080/00222933.2023.2203335 . ISSN 0022-2933 .
- ^ Перейти обратно: а б Эдельштам, Карл; Палмер, Карина (1950). «Поведение брюхоногих моллюсков» . Ойкос . 2 (2): 259–270. Бибкод : 1950Ойкос...2..259Е . дои : 10.2307/3564796 . JSTOR 3564796 – через JSTOR.
- ^ Перейти обратно: а б с Линд, Ганс (1990). «Стратегии пространственного поведения Helix pomatia ». Этология . 86 (1): 1–18. Бибкод : 1990Этол..86....1Л . дои : 10.1111/j.1439-0310.1990.tb00414.x .
- ^ Ярчуска, Беньямин; Ярчушкова Данкова, Люсия (2014). «Комментарии Helix к агрегационному поведению римской улитки ( pomatia )» (PDF) . Малакологика Богемословаца . 13 : 114–115. дои : 10.5817/MaB2014-13-114 .
- ^ Schileyko, A. A. (1978). Mollyuski, Tom III, vyp. 6. Nazemnye mollyuski nadsemeystva Helicoidea . Fauna SSSR. Leningrad: Nauka.
- ^ Миенис, Хенк (1993). «Какие птицы едят виноградных улиток?» (PDF) . Заочный журнал Голландской малакологической ассоциации . 273 : 92–93.
- ^ Перейти обратно: а б Аллен, Джон А. (2004). «Птицы - млекопитающие, хищники наземных брюхоногих моллюсков». В Баркере, GM (ред.). Естественные враги наземных моллюсков . Кембридж (США): Издательство CABI. стр. 1–36. ISBN 0-85199-319-2 .
- ^ Шли, Лоран; Ропер, Тимоти Дж. (2003). «Диета дикого кабана Sus scrofa в Западной Европе, с особым упором на потребление сельскохозяйственных культур: Диета дикого кабана» . Обзор млекопитающих . 33 (1): 43–56. дои : 10.1046/j.1365-2907.2003.00010.x .
- ^ Лапота-Феррейра, Яра Люсия (2004). «Рептилии-хищники наземных брюхоногих моллюсков». В Баркере, GM (ред.). Естественные враги наземных моллюсков . Кембридж (США): Издательство CABI. стр. 427–482. ISBN 0-85199-319-2 .
- ^ Симондсон, Уильям О.К. (2004). «Жестокрылые (Carabidae, Staphylinidae, Lampyridae, Drilidae и Silphidae) как хищники наземных брюхоногих моллюсков». В Баркере, GM (ред.). Естественные враги наземных моллюсков . Кембридж (США): Издательство CABI. стр. 37–84. ISBN 0-85199-319-2 .
- ^ Купленд, Джеймс Б.; Баркер, Гэри Б. (2004). «Двукрылые как хищники и паразитоиды наземных брюхоногих моллюсков с упором на Phoridae, Calliphoridae, Sarcophagidae, Muscidae и Fanniidae». В Баркере, GM (ред.). Естественные враги наземных моллюсков . Кембридж (США): Издательство CABI. стр. 85–158. ISBN 0-85199-319-2 .
- ^ Моран, Серж; Уилсон, Майкл Дж.; Глен, Дэвид М. (2004). «Нематоды (Нематоды), паразитирующие на наземных брюхоногих моллюсках». В Баркере, GM (ред.). Естественные враги наземных моллюсков . Кембридж (США): Издательство CABI. стр. 525–558. ISBN 0-85199-319-2 .
- ^ Терк, Фрэнк А.; Филлипс, Стелла-Марис (1946). «Монография слизняка - Riccardoella limacum (Schrank.)» . Труды Лондонского зоологического общества . 115 (3–4): 448–472. дои : 10.1111/j.1096-3642.1946.tb00102.x . ISSN 0370-2774 .
- ^ Фейн, Александр (2004). «Клещи (Acari), паразитирующие и хищные на наземных брюхоногих моллюсках». В Баркере, GM (ред.). Естественные враги наземных моллюсков . Кембридж (США): Издательство CABI. стр. 505–524. ISBN 0-85199-319-2 .
- ^ Ван Ас, Джо Г.; Бассон, Линда (2004). «Цилиофоры (Ciliophora) паразиты наземных брюхоногих моллюсков». В Баркере, GM (ред.). Естественные враги наземных моллюсков . Кембридж (США): Издательство CABI. стр. 559–578. ISBN 0-85199-319-2 .
- ^ Раут, Шриманта К. (2004). «Бактериальные и немикробные заболевания наземных брюхоногих моллюсков». В Баркере, GM (ред.). Естественные враги наземных моллюсков . Кембридж (США): Издательство CABI. стр. 599–611. ISBN 0-85199-319-2 .
- ^ Бисенья, Август; Д'Анджело, Эмануэла; Джой, Патрисия; Мусси, Маргарита (2011). «Сбор и приготовление наземных улиток в Абруццо: доисторическое прошлое и настоящее». В июле Франческа; Стоппьелло, Алессандра Ассунта; Бьяджетти, Стефано (ред.). Материалы 4-й Национальной конференции по этноархеологии, Рим, 17-19 мая 2006 г. (на итальянском языке). Оксфорд: Археопресс. стр. 95–101. ISBN 978-1-4073-0797-8 .
- ^ Иверсен, Ингрид (2015). «Моллюски Бестансура, Иракский Курдистан: пример эксплуатации Helix salomonica в раннем голоцене в Загросе» . Экологическая археология . 20 (3): 239–250. дои : 10.1179/1749631415Y.0000000023 . ISSN 1461-4103 .
- ^ Рид, Чарльз А. (1962). «Улитки на склоне персидского холма. Экология — Предыстория — Гастрономия» . Постилла . 66 . Йельский музей Пибоди: 1–20.
- ^ Любелл, Дэвид; Хасан, Фекри А.; Готье, Ахиллес; Балле, Жан-Луи (5 марта 1976 г.). «Капсийские эскарготьеры: междисциплинарное исследование, проливающее свет на экологию и образ жизни голоцена в Северной Африке» . Наука . 191 (4230): 910–920. дои : 10.1126/science.191.4230.910 . ISSN 0036-8075 .
- ^ Кеннард, А.С.; Вудворд, BB; Блэкмор, HP (1901). «Постплиоценовый неморской моллюск юга Англии» . Труды Ассоциации геологов . 17 (5): 213–260. дои : 10.1016/s0016-7878(01)80060-2 . ISSN 0016-7878 .
- ^ «Красный список видов, находящихся под угрозой исчезновения» МСОП . Проверено 11 мая 2023 г.
- ^ Буше, П.; Рипкен, Т.; Рекорбет, Р. (1998). «Сохранение узкого ареала средиземноморского острова, Helix ceratina с Корсики». Журнал конхологии . Специальное издание № 2: 205–208.
- ^ Уэлч, Джоан М.; Поллард, Э. (1 сентября 1975 г.). «Эксплуатация Helix pomatia L.». Биологическая консервация . 8 (2): 155–160. Бибкод : 1975BCCons...8..155W . дои : 10.1016/0006-3207(75)90040-3 . ISSN 0006-3207 .
- ^ Перейти обратно: а б Корабек, Ондржей; Косова, Тереза; Долейш, Петр; Петрусек, Адам; Нойберт, Эйке; Юржичкова, Люси (29 ноября 2021 г.). «Географическая изоляция и распространение наземных улиток с помощью человека: средиземноморская история Helix Borealis и ее родственников (Gastropoda: Stylommatophora: Helicidae)» . Зоологический журнал Линнеевского общества . 193 (4): 1310–1335. doi : 10.1093/zoolinnean/zlaa186 . ISSN 0024-4082 .
- ^ Хеллер, Джозеф (1984). «Наземные улитки из южного Ливана». Журнал конхологии . 31 : 331–336.
- ^ Перейти обратно: а б Венц, Вильгельм (1923). Каталог окаменелостей I: Animalia. Парс 18: Gastropoda extramarina tertiaria. II . Берлин: В. Джанк. стр. 706–708.
- ^ Нойбауэр, Томас А.; Харцхаузер, Матиас (13 декабря 2023 г.). «Омонимия и почти-омонимия у двух европейских кайнозойских видов наземных улиток (Gastropoda: Helicoidea) с описанием нового рода» . Archiv für Molluskenkunde Международный журнал малакологии . 152 (2): 153–158. дои : 10.1127/arch.moll/152/153-158 .
- ^ Steklov, A. A. (1966). Nazemnȳe mollyuski neogena Predkavkaz'ya i ikh stratigraficheskoe znachenie . Moskva: Nauka.
- ^ Белуженко Е.В. (2014). «Неоген-эоплейстоценовые ракушечники Северного Кавказа и Предкавказья» . Стратиграфия и геологическая корреляция . 22 (4): 406–425. Бибкод : 2014SGC....22..406B . дои : 10.1134/S0869593814040029 . ISSN 0869-5938 .
- ^ Корабек, Ондржей; Петрусек, Адам; Юржичкова, Люси (1 января 2018 г.). «Ледниковые рефугиумы и послеледниковое распространение культовой крупной европейской наземной улитки Helix pomatia (Pulmonata: Helicidae)» . Биологический журнал Линнеевского общества . 123 (1): 218–234. дои : 10.1093/biolinnean/blx135 . ISSN 0024-4066 .
- ^ Корабек, Ондржей; Адамцова, Тереза; Прочков, Малгожата; Петрусек, Адам; Хаусдорф, Бернхард; Юржичкова, Люси (2023). «В обоих направлениях: Экспансия европейских наземных улиток на север и юг от ледниковых рефугиумов» . Журнал биогеографии . 50 (4): 654–668. Бибкод : 2023JBiog..50..654K . дои : 10.1111/jbi.14531 . ISSN 0305-0270 . S2CID 253909222 .
- ^ Реле, Вольфганг (1983). «Моллюски из раскопок Хельга-Абри близ Шельклингена с примечанием об открытии некоторых мезолитических декоративных улиток». Лист археологической переписки (на немецком языке). 13 :29–36.
- ^ Реле, Вольфганг (1987). «Фауна моллюсков из раскопок Фельсшталле недалеко от Мюлена, город Эхинген, Альб-Донау-Крайс». Исследования и отчеты по предыстории и ранней истории Баден-Вюртемберга . 23 :269-274.
- ^ Гиттенбергер, Эдмунд; Хороший друг, Гленн А. (1993). «Наземные улитки из последнего ледникового максимума на Андикитире, южная Греция, и их палеоклиматические последствия» . Журнал четвертичной науки . 8 (2): 109–116. Бибкод : 1993JQS.....8..109G . дои : 10.1002/jqs.3390080203 .
- ^ Вельтер-Шультес, Франциско (1998). «Наземные улитки греческого острова Гавдос, самого южного острова Европы». Труды по малакозоологии . 12 :1-120.
- ^ Найбер, Марко Т.; Хаусдорф, Бернхард (2015). «Молекулярная филогения выявляет полифилию улиток рода Cepaea (Gastropoda: Helicidae)» . Молекулярная филогенетика и эволюция . 93 : 143–149. Бибкод : 2015МОЛПЭ..93..143Н . дои : 10.1016/j.ympev.2015.07.022 . ПМИД 26256642 .
- ^ Корабек, Ондржей; Петрусек, Адам; Нойберт, Эйке; Юржичкова, Люси (2015). «Молекулярная филогения рода Helix (Pulmonata: Helicidae)» . Зоологика Скрипта . 44 (3): 263–280. дои : 10.1111/zsc.12101 . S2CID 84673484 .
| Базы данных органов управления : Национальные |
|---|