Гликоген

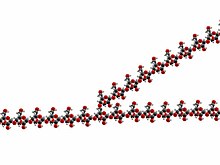

Гликоген — многоразветвленный полисахарид глюкозы , служащий у животных хранилищем энергии . [2] грибы и бактерии. [3] Это основная форма хранения глюкозы в организме человека.
Гликоген функционирует как одна из трех регулярно используемых форм энергетических резервов: креатинфосфат предназначен для очень краткосрочного хранения, гликоген предназначен для краткосрочного хранения, а запасы триглицеридов в жировой ткани (т. е. в жировых отложениях) предназначены для долгосрочного хранения. Белок, расщепленный на аминокислоты, редко используется в качестве основного источника энергии, за исключением случаев голодания и гликолитического кризиса (см. Биоэнергетические системы ) .
У человека гликоген вырабатывается и хранится в основном в клетках печени и скелетных мышц . [4] [5] В печени гликоген может составлять 5–6% сырой массы органа: печень взрослого человека массой 1,5 кг может хранить примерно 100–120 г гликогена. [4] [6] В скелетных мышцах гликоген содержится в низкой концентрации (1–2% мышечной массы): скелетные мышцы взрослого человека массой 70 кг запасают примерно 400 г гликогена. [4] Небольшие количества гликогена также обнаруживаются в других тканях и клетках, включая почки , эритроциты , [7] [8] [9] белые кровяные тельца , [10] и глиальные клетки головного мозга . [11] Матка также сохраняет гликоген во время беременности для питания эмбриона. [12]
Количество гликогена, хранящегося в организме, в основном зависит от окислительных волокон 1 типа. [13] [14] физическая подготовка, основной обмен веществ и привычки питания. [15] Различные уровни гликогена в состоянии покоя в мышцах достигаются за счет изменения количества частиц гликогена, а не за счет увеличения размера существующих частиц. [14] хотя большинство частиц гликогена в состоянии покоя меньше их теоретического максимума. [16]
присутствует около 4 граммов глюкозы ; В крови человека постоянно [4] у голодающих людей уровень глюкозы в крови поддерживается постоянным на этом уровне за счет запасов гликогена, главным образом из печени (гликоген в скелетных мышцах в основном используется в качестве непосредственного источника энергии для этих мышц, а не для поддержания физиологического уровня глюкозы в крови). ). [4] Запасы гликогена в скелетных мышцах служат хранилищем энергии для самих мышц; [4] однако распад мышечного гликогена препятствует поглощению глюкозы мышцами из крови, тем самым увеличивая количество глюкозы в крови, доступной для использования в других тканях. [4] Запасы гликогена в печени служат хранилищем глюкозы для использования во всем организме, особенно в центральной нервной системе . [4] Человеческий мозг потребляет около 60% глюкозы в крови у голодающих и малоподвижных людей. [4]
Гликоген – это аналог крахмала глюкозы , полимера , который служит хранилищем энергии в растениях . Он имеет структуру, похожую на амилопектин (компонент крахмала), но более разветвлен и компактен, чем крахмал. Оба представляют собой белые порошки в сухом состоянии. Гликоген содержится в форме гранул в цитозоле /цитоплазме многих типов клеток и играет важную роль в цикле глюкозы . Гликоген образует энергетический резерв, который можно быстро мобилизовать для удовлетворения внезапной потребности в глюкозе, но он менее компактен, чем энергетические запасы триглицеридов ( липидов ). Как таковой он также встречается в качестве резерва хранения у многих паразитических простейших. [17] [18] [19]
Структура [ править ]
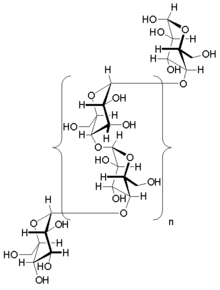
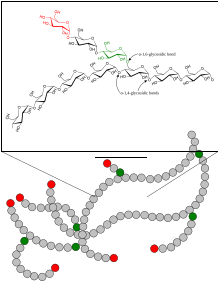
Гликоген — разветвленный биополимер, состоящий из линейных цепочек глюкозы остатков со средней длиной цепи примерно 8–12 единиц глюкозы и 2000–60 000 остатков на одну молекулу гликогена. [20] [21]
Как и амилопектин, единицы глюкозы связаны между собой линейно гликозидными связями α(1→4) от одной глюкозы к другой. Ветви связаны с цепями, от которых они отходят, гликозидными связями α(1→6) между первой глюкозой новой ветви и глюкозой в стволовой цепи. [22]
Каждый гликоген, по сути, представляет собой клубок глюкозных деревьев примерно с 12 слоями, в центре которого находится белок гликогенин , с тремя видами цепей глюкозы: A, B и C. К гликогенину прикреплена только одна C-цепь. Эта С-цепь образуется в результате самоглюкозилирования гликогенина, образуя короткую цепь праймера. Из С-цепи вырастают В-цепи, а из В-цепей разветвляются В- и А-цепи. В-цепи имеют в среднем 2 точки ветвления, тогда как А-цепи являются терминальными и, следовательно, неразветвленными. В среднем каждая цепь имеет длину 12, строго ограниченную от 11 до 15. Все А-цепи достигают сферической поверхности гликогена. [23] [24]
Гликоген в мышцах, печени и жировых клетках хранится в гидратированной форме, состоящей из трех или четырех частей воды на часть гликогена, связанных с 0,45 миллимоль (18 мг) калия на грамм гликогена. [5]
Глюкоза является осмотической молекулой и может оказывать сильное влияние на осмотическое давление в высоких концентрациях, что может привести к повреждению или гибели клеток, если она хранится в клетке без модификации. [3] Гликоген является неосмотической молекулой, поэтому его можно использовать в качестве раствора для хранения глюкозы в клетке без нарушения осмотического давления. [3]
Функции [ править ]
Печень [ править ]
Когда еда, содержащая углеводы или белок, съедается и переваривается , уровень глюкозы в крови повышается, а поджелудочная железа выделяет инсулин . Глюкоза крови из воротной вены поступает в клетки печени ( гепатоциты ). Инсулин действует на гепатоциты, стимулируя действие нескольких ферментов , включая гликогенсинтазу . Молекулы глюкозы добавляются к цепям гликогена до тех пор, пока инсулина и глюкозы остается в избытке. В этом постпрандиальном или «насыщенном» состоянии печень поглощает из крови больше глюкозы, чем высвобождает.
После того, как еда переварилась и уровень глюкозы начинает падать, секреция инсулина снижается, а синтез гликогена прекращается. Когда гликоген необходим для получения энергии , он расщепляется и снова превращается в глюкозу. Гликогенфосфорилаза является основным ферментом расщепления гликогена. В течение следующих 8–12 часов глюкоза, полученная из гликогена печени, является основным источником глюкозы в крови, используемой остальным телом в качестве топлива.
Глюкагон , еще один гормон, вырабатываемый поджелудочной железой, во многих отношениях служит контрсигналом инсулину. В ответ на уровень инсулина ниже нормы (когда уровень глюкозы в крови начинает падать ниже нормального диапазона) глюкагон секретируется в возрастающих количествах и стимулирует как гликогенолиз (распад гликогена), так и глюконеогенез (производство глюкозы из других источников). .
Мышцы [ править ]

Мышечный гликоген, по-видимому, функционирует как резерв быстродоступной фосфорилированной глюкозы в форме глюкозо-1-фосфата для мышечных клеток. Гликоген, содержащийся в клетках скелетных мышц, находится преимущественно в форме β-частиц. [25] Другие клетки, содержащие небольшие количества, также используют его локально. Поскольку в мышечных клетках отсутствует глюкозо-6-фосфатаза , которая необходима для передачи глюкозы в кровь, гликоген, который они хранят, доступен исключительно для внутреннего использования и не передается другим клеткам. В этом отличие от клеток печени, которые при необходимости легко расщепляют накопленный гликоген на глюкозу и отправляют его через кровоток в качестве топлива для других органов. [26]
Скелетным мышцам необходима АТФ (обеспечивает энергию) для сокращения и расслабления мышц , что известно как теория скользящих нитей . Скелетные мышцы преимущественно полагаются на гликогенолиз в течение первых нескольких минут при переходе от покоя к активности, а также во время высокоинтенсивной аэробной активности и любой анаэробной активности. [27] Во время анаэробной активности, такой как тяжелая атлетика и изометрические упражнения , фосфагенная система (АТФ-ПЦр) и мышечный гликоген являются единственными используемыми субстратами, поскольку они не требуют ни кислорода, ни кровотока. [27]
Различные биоэнергетические системы производят АТФ с разной скоростью, причем АТФ вырабатывается из мышечного гликогена намного быстрее, чем окисление жирных кислот. [28] Уровень интенсивности упражнений также определяет, какое количество субстрата (топлива) используется для синтеза АТФ. Мышечный гликоген может поставлять гораздо больше субстрата для синтеза АТФ, чем глюкоза в крови. Во время упражнений максимальной интенсивности мышечный гликоген может поставлять 40 ммоль глюкозы на кг сырого веса в минуту. [29] тогда как глюкоза в крови может поставлять 4-5 ммоль. [30] [4] Благодаря высокой скорости поступления и быстрому синтезу АТФ во время высокоинтенсивной аэробной активности (например, быстрой ходьбы, бега трусцой или бегом) чем выше интенсивность тренировки, тем больше мышечная клетка производит АТФ из мышечного гликогена. [31] Эта зависимость от мышечного гликогена предназначена не только для обеспечения мышц достаточным количеством АТФ во время высокоинтенсивных упражнений, но и для поддержания гомеостаза глюкозы в крови (то есть, чтобы не допустить гипогликемии из-за того, что мышцам необходимо извлечь из крови гораздо больше глюкозы, чем печень может обеспечить). [30] Дефицит мышечного гликогена приводит к мышечной усталости , известной как «удар о стену» или «бонк» (см. ниже об истощении гликогена) .
Тип структуры [ править ]
В 1999 году Мелендес и др. заявили, что структура гликогена является оптимальной в рамках конкретной модели метаболических ограничений, в которой предполагалось, что структура имеет «фрактальную» природу. [32] Однако исследования Бесфорда и др. [33] использовали эксперименты по малоугловому рассеянию рентгеновских лучей, сопровождаемые моделями теории ветвления, чтобы показать, что гликоген представляет собой случайно сверхразветвленную полимерную наночастицу. Гликоген не фрактален по своей природе. Впоследствии это было подтверждено другими людьми, которые выполнили моделирование роста частиц гликогена методом Монте-Карло и показали, что молекулярная плотность достигает максимума вблизи центра структуры наночастиц, а не на периферии (что противоречит фрактальной структуре, которая имела бы большую плотность при периферия). [34]
История [ править ]
Гликоген был открыт Клодом Бернаром . Его эксперименты показали, что в печени содержится вещество, которое может привести к восстановлению сахара под действием «фермента» в печени. К 1857 году он описал выделение вещества, которое он назвал « la matière гликоген », или «сахарообразующее вещество». Вскоре после открытия гликогена в печени М. А. Сансон обнаружил, что мышечная ткань также содержит гликоген. Эмпирическая формула гликогена ( C
66Ч
10 О
5 ) n была основана Августом Кекуле в 1858 году. [35]
Сансон, М.А. «Заметки о физиологическом образовании сахара в животноводстве». Отчеты о сессиях Академии наук 44 (1857): 1323-5.
Метаболизм [ править ]
Синтез [ править ]
Синтез гликогена, в отличие от его распада, эндергоничен — он требует затрат энергии. Энергия для синтеза гликогена поступает из уридинтрифосфата (УТФ), который реагирует с глюкозо-1-фосфатом , образуя УДФ-глюкозу , в реакции, катализируемой УТФ — глюкозо-1-фосфатуридилилтрансферазой . Гликоген синтезируется из мономеров УДФ-глюкозы первоначально белком гликогенином , имеющим два тирозиновых якоря на восстанавливающем конце гликогена, поскольку гликогенин является гомодимером. После того, как к остатку тирозина было добавлено около восьми молекул глюкозы, фермент гликогенсинтаза постепенно удлиняет гликогеновую цепь с помощью УДФ-глюкозы, добавляя α(1→4)-связанную глюкозу к невосстанавливающему концу гликогеновой цепи. [36]
Фермент разветвления гликогена катализирует перенос терминального фрагмента из шести или семи остатков глюкозы с невосстанавливающего конца на гидроксильную группу С-6 остатка глюкозы глубже внутрь молекулы гликогена. Фермент разветвления может действовать только на ветвь, имеющую по меньшей мере 11 остатков, и фермент может переносить ту же цепь глюкозы или соседние цепи глюкозы.
Разбивка [ править ]
Гликоген отщепляется от невосстанавливающих концов цепи ферментом гликогенфосфорилазой с образованием мономеров глюкозо-1-фосфата:
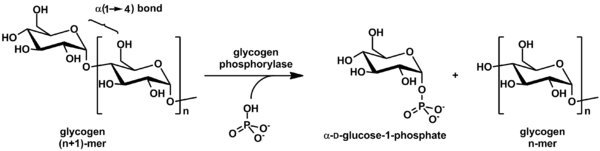
In vivo фосфоролиз протекает в направлении распада гликогена, поскольку соотношение фосфата и глюкозо-1-фосфата обычно превышает 100. [37] Глюкозо-1-фосфат затем превращается в глюкозо-6-фосфат (G6P) под действием фосфоглюкомутазы . специальный разветвляющий фермент Для удаления разветвлений α(1→6) в разветвленном гликогене и преобразования цепи в линейный полимер необходим . Произведенные мономеры G6P имеют три возможных судьбы:
- G6P может продолжать путь гликолиза и использоваться в качестве топлива.
- G6P может вступать в пентозофосфатный путь через фермент глюкозо-6-фосфатдегидрогеназу с образованием НАДФН и 5-углеродных сахаров.
- В печени и почках G6P может дефосфорилироваться обратно в глюкозу с помощью фермента глюкозо-6-фосфатазы . Это заключительный этап пути глюконеогенеза .
Клиническая значимость
Нарушения обмена гликогена [ править ]
Наиболее распространенным заболеванием, при котором метаболизм гликогена нарушается, является диабет , при котором из-за аномального количества инсулина гликоген в печени может аномально накапливаться или истощаться. Восстановление нормального метаболизма глюкозы обычно нормализует и метаболизм гликогена.
При гипогликемии, вызванной избытком инсулина, уровень гликогена в печени высок, но высокий уровень инсулина предотвращает гликогенолиз, необходимый для поддержания нормального уровня сахара в крови. Глюкагон является распространенным средством лечения этого типа гипогликемии.
Различные врожденные нарушения углеводного обмена вызваны недостатком ферментов или транспортных белков, необходимых для синтеза или распада гликогена. В совокупности они называются болезнями накопления гликогена .
истощение гликогена и выносливость на Упражнения
Спортсмены на длинные дистанции, такие как марафонцы , лыжники и велосипедисты , часто испытывают истощение гликогена, когда почти все запасы гликогена у спортсменов истощаются после длительных периодов нагрузки без достаточного потребления углеводов. Это явление называется « ударом о стену » при беге и «стук» при езде на велосипеде.
Истощение гликогена можно предотвратить тремя возможными способами:
- углеводы с максимально возможной скоростью превращения в глюкозу крови (высокий гликемический индекс Во-первых, во время физических упражнений постоянно потребляются ). Наилучший возможный результат этой стратегии — замена примерно 35% глюкозы, потребляемой при частоте сердечных сокращений выше примерно 80% от максимальной.
- Во-вторых, посредством адаптации тренировок на выносливость и специализированных режимов (например, голодание, тренировки на выносливость низкой интенсивности) организм может подготовить мышечные волокна типа I , чтобы улучшить как эффективность использования топлива, так и способность к нагрузке, чтобы увеличить процент жирных кислот, используемых в качестве топлива. [38] [39] щадящее употребление углеводов из всех источников.
- В-третьих, потребляя большое количество углеводов после истощения запасов гликогена в результате физических упражнений или диеты, организм может увеличить емкость внутримышечных запасов гликогена. [13] [40] [41] [42] Этот процесс известен как углеводная загрузка . В общем, гликемический индекс источника углеводов не имеет значения, поскольку чувствительность мышц к инсулину увеличивается в результате временного истощения гликогена. [43] [44]
Когда спортсмены после изнурительных тренировок употребляют и углеводы, и кофеин , их запасы гликогена пополняются быстрее; [45] [46] однако минимальная доза кофеина, при которой наблюдается клинически значимое влияние на восполнение запасов гликогена, не установлена. [46]
Наномедицина [ править ]
Недавно наночастицы гликогена были исследованы как потенциальные системы доставки лекарств. [47]
См. также [ править ]
Ссылки [ править ]
- ^ Макардл, Уильям Д.; Кэтч, Фрэнк И.; Кач, Виктор Л. (2006). Физиология упражнений: энергия, питание и работоспособность человека (6-е изд.). Липпинкотт Уильямс и Уилкинс. п. 12. ISBN 978-0-7817-4990-9 .
- ^ Садава, Дэвид Э.; Первс, Уильям К.; Хиллис, Дэвид М.; Орианс, Гордон Х.; Хеллер, Х. Крейг (2011). Жизнь (9-е изд.). У. Х. Фриман. ISBN 9781429254311 .
- ^ Jump up to: Перейти обратно: а б с Берг Дж. М., Тимочко Дж. Л., Гатто Г. Дж., Страйер Л. (8 апреля 2015 г.). Биохимия (8-е изд.). Нью-Йорк: WH Freeman. ISBN 9781464126109 . OCLC 913469736 .
- ^ Jump up to: Перейти обратно: а б с д и ж г час я дж Вассерман Д.Х. (январь 2009 г.). «Четыре грамма глюкозы» . Американский журнал физиологии. Эндокринология и обмен веществ . 296 (1): Е11–21. дои : 10.1152/ajpendo.90563.2008 . ПМК 2636990 . ПМИД 18840763 .
В крови человека массой 70 кг циркулирует четыре грамма глюкозы. Эта глюкоза имеет решающее значение для нормального функционирования многих типов клеток. В соответствии с важностью этих 4 г глюкозы существует сложная система контроля для поддержания постоянного уровня глюкозы в крови. Наше внимание было сосредоточено на механизмах, с помощью которых регулируется поток глюкозы из печени в кровь и из крови в скелетные мышцы. ... Мозг потребляет ~60% глюкозы в крови человека, ведущего малоподвижный образ жизни и голодающего. ...Количество глюкозы в крови сохраняется за счет резервуаров гликогена (рис. 2). У людей после абсорбции около 100 г гликогена содержится в печени и около 400 г гликогена в мышцах. Окисление углеводов работающими мышцами может увеличиться примерно в 10 раз при выполнении упражнений, и все же после 1 час уровень глюкозы в крови поддерживается на уровне ~4 г.
- ^ Jump up to: Перейти обратно: а б Крейцман С.Н., Коксон А.Ю., Саз К.Ф. (июль 1992 г.). «Запасы гликогена: иллюзии легкой потери веса, чрезмерного набора веса и искажения оценок состава тела» . Американский журнал клинического питания . 56 (1, Приложение): 292–293. дои : 10.1093/ajcn/56.1.292S . ПМИД 1615908 .
- ^ Гайтон, Артур К.; Холл, Джон Эдвард (2011). Учебник медицинской физиологии Гайтона и Холла . Нью-Йорк, Нью-Йорк: Сондерс/Эльзевир. ISBN 978-5-98657-013-6 .
- ^ Моисей С.В., Башан Н., Гутман А. (декабрь 1972 г.). «Метаболизм гликогена в нормальных эритроцитах» . Кровь . 40 (6): 836–843. дои : 10.1182/blood.V40.6.836.836 . ПМИД 5083874 .
- ^ Ингерманн Р.Л., Virgin GL (1987). «Содержание гликогена и высвобождение глюкозы из эритроцитов сипункуланового червя themiste Dyscrita» (PDF) . J Exp Biol . 129 : 141–149. дои : 10.1242/jeb.129.1.141 .
- ^ Мива I, Suzuki S (ноябрь 2002 г.). «Улучшенный количественный анализ гликогена в эритроцитах». Анналы клинической биохимии . 39 (Часть 6): 612–13. дои : 10.1258/000456302760413432 . ПМИД 12564847 .
- ^ Мюррей, Боб (апрель 2018 г.). «Основы обмена гликогена для тренеров и спортсменов» . Обзоры питания . 76 (4): 243–259. дои : 10.1093/nutrit/nuy001 . ПМК 6019055 . ПМИД 29444266 .
- ^ Оэ Ю, Баба О, Ашида Х, Накамура К.С., Хирасе Х (июнь 2016 г.). «Распределение гликогена в мозге мыши, зафиксированном в микроволновой печи, обнаруживает гетерогенные астроцитарные структуры» . Глия . 64 (9): 1532–1545. дои : 10.1002/glia.23020 . ПМК 5094520 . ПМИД 27353480 .
- ^ Кэмпбелл, Нил А.; Уильямсон, Брэд; Хейден, Робин Дж. (2006). Биология: исследование жизни . Бостон, Массачусетс: Пирсон Прентис Холл. ISBN 978-0-13-250882-7 .
- ^ Jump up to: Перейти обратно: а б Дженсен, Расмус; Ортенблад, Нильс; Стаушольм, Мария-Луиза Холлейфер; Скьербек, Метте Карина; Ларсен, Дэниел Нюквист; Хансен, Метте; Хольмберг, Ганс-Кристер; Пломгаард, Питер; Нильсен, Иоахим (1 октября 2020 г.). «Неоднородность использования субклеточного мышечного гликогена во время тренировок влияет на выносливость у мужчин» . Журнал физиологии . 598 (19): 4271–4292. дои : 10.1113/JP280247 . ISSN 1469-7793 . PMID 32686845 . S2CID 220653138 .
- ^ Jump up to: Перейти обратно: а б Дженсен, Расмус; Ортенблад, Нильс; Стаушольм, Мари-Луиза Х.; Скьербек, Метте К.; Ларсен, Дэниел Н.; Хансен, Метте; Хольмберг, Ганс-Кристер; Пломгаард, Питер; Нильсен, Иоахим (1 мая 2021 г.). «Суперкомпенсация гликогена происходит из-за увеличения количества, а не размера частиц гликогена в скелетных мышцах человека» . Экспериментальная физиология . 106 (5): 1272–1284. дои : 10.1113/EP089317 . ISSN 0958-0670 . ПМИД 33675088 . S2CID 232131416 .
- ^ Бергстрем, Йонас; Хермансен, Ларс; Хультман, Эрик; Салтин, Бенгт (октябрь 1967 г.). «Диета, мышечный гликоген и физическая работоспособность». Acta Physiologica Scandinavica . 71 (2–3): 140–150. дои : 10.1111/j.1748-1716.1967.tb03720.x . ISSN 1365-201X . ПМИД 5584523 .
- ^ Маршан, И.; Чорнейко, К.; Тарнопольский, М.; Гамильтон, С.; Ширер, Дж.; Потвин, Дж.; Грэм, TE (1 ноября 2002 г.). «Количественное определение субклеточного гликогена в покоящихся мышцах человека: размер, количество и расположение гранул» . Журнал прикладной физиологии . 93 (5): 1598–1607. doi : 10.1152/japplphysicalol.00585.2001 . ISSN 8750-7587 . ПМИД 12381743 .
- ^ Райли, Дж. Ф. (март 1955 г.). «Исследования метаболизма простейших. 5: Метаболизм паразитических жгутиконосцев Trichomonas foetus» . Биохимический журнал . 59 (3): 361–369. дои : 10.1042/bj0590361 . ПМК 1216250 . ПМИД 14363101 .
- ^ Бенчимол, Марлен; Элиас, Цезарь Антонио; де Соуза, Вандерли (декабрь 1982 г.). « Плод Tritrichomonas : Ультраструктурная локализация кальция в плазматической мембране и в гидрогеносоме». Экспериментальная паразитология . 54 (3): 277–284. дои : 10.1016/0014-4894(82)90036-4 . ISSN 0014-4894 . ПМИД 7151939 .
- ^ Мелевчик, Майкл; Мельхорн, Хайнц; аль-Курайши, Салех; Грабенштайнер, Э.; Хесс, М. (1 сентября 2008 г.). «Просвечивающее электронно-микроскопическое исследование стадий histomonas meleagridis из клональных культур». Паразитологические исследования . 103 (4): 745–750. дои : 10.1007/s00436-008-1009-1 . ISSN 0932-0113 . ПМИД 18626664 . S2CID 2331300 .
- ^ Маннерс, Дэвид Дж. (1991). «Последние достижения в нашем понимании структуры гликогена». Углеводные полимеры . 16 (1): 37–82. дои : 10.1016/0144-8617(91)90071-J . ISSN 0144-8617 .
- ^ Роннер, Питер (2018). Основы биохимии Неттера . США: Эльзевир. п. 254. ИСБН 978-1-929007-63-9 .
- ^ Берг, Джереми Марк; Тимочко, Джон Л.; Страйер, Люберт (2012). Биохимия (7-е изд.). У. Х. Фриман. п. 338 . ISBN 978-1429203142 .
- ^ Гунджа-Смит, Зинат; Маршалл, Джей-Джей; Мерсье, Кристиана; Смит, Э.Э.; Уилан, WJ (28 декабря 1970 г.). «Пересмотр модели гликогена и амилопектина Мейера-Бернфельда» . Письма ФЭБС . 12 (2): 101–104. дои : 10.1016/0014-5793(70)80573-7 . ISSN 0014-5793 . ПМИД 11945551 . S2CID 34722785 .
- ^ Роуч, Питер Дж.; Депаоли-Роуч, Анна А.; Херли, Томас Д.; Тальябраччи, Винсент С. (16 января 2012 г.). «Гликоген и его метаболизм: некоторые новые разработки и старые темы» . Биохимический журнал . 441 (3): 763–787. дои : 10.1042/BJ20111416 . ISSN 0264-6021 . ПМЦ 4945249 . ПМИД 22248338 .
- ^ Лю, QH; Ванга, З.Ы.; Тан, JW; Моу, JY; Ма, ЗВ; Дэн, Б.; Лю, З.; Ван, Л. (2022). «Бровизайн» . Углеводные полимеры . 295 . дои : 10.1016/j.carbpol.2022.119710 . ПМИД 35989025 . S2CID 249489284 . Проверено 12 мая 2023 г.
- ^ «Биосинтез гликогена; Распад гликогена» . oregonstate.edu . Архивировано из оригинала 12 мая 2021 года . Проверено 28 февраля 2018 г.
- ^ Jump up to: Перейти обратно: а б Люсия, Алехандро; Мартинуцци, Андреа; Ногалес-Гадеа, Гизела; Куинливан, Рос; Разум, Стейси; Исследовательская группа Международной ассоциации по изучению болезни накопления мышечного гликогена (декабрь 2021 г.). «Руководство по клинической практике болезней накопления гликогена V и VII (болезнь МакАрдла и болезнь Таруи) от международной исследовательской группы» . Нервно-мышечные расстройства . 31 (12): 1296–1310. дои : 10.1016/j.nmd.2021.10.006 . ISSN 1873-2364 . ПМИД 34848128 . S2CID 240123241 .
- ^ «Гормональная регуляция энергетического метаболизма – физиология Берна и Леви, 6-е изд. (2008 г.)» . Doctorlib.info . Проверено 21 октября 2023 г.
- ^ Мюррей, Боб; Розенблум, Кристина (1 апреля 2018 г.). «Основы обмена гликогена для тренеров и спортсменов» . Обзоры питания . 76 (4): 243–259. дои : 10.1093/nutrit/nuy001 . ISSN 1753-4887 . ПМК 6019055 . ПМИД 29444266 .
- ^ Jump up to: Перейти обратно: а б Брукс, Джордж А. (январь 2020 г.). «Несколько драгоценных граммов глюкозы во время тренировки» . Международный журнал молекулярных наук . 21 (16): 5733. doi : 10.3390/ijms21165733 . ISSN 1422-0067 . ПМЦ 7461129 . ПМИД 32785124 .
- ^ ван Лун, LJ; Гринхафф, Польша; Константин-Теодосиу, Д.; Сарис, Вашингтон; Вагенмейкерс, AJ (1 октября 2001 г.). «Влияние увеличения интенсивности упражнений на использование мышечного топлива у людей» . Журнал физиологии . 536 (Часть 1): 295–304. дои : 10.1111/j.1469-7793.2001.00295.x . ISSN 0022-3751 . ПМК 2278845 . ПМИД 11579177 .
- ^ Рут Мелендес; Энрике Мелендес-Эвиа; Энрик И. Канела (сентябрь 1999 г.). «Фрактальная структура гликогена: умное решение для оптимизации клеточного метаболизма». Биофизический журнал . 77 (3). 1327. дои : 10.1016/S0006-3495(99)76982-1 . hdl : 2445/122234 .
- ^ Куинн А. Бесфорд; Сяо-И Цзэн; Цзи-Мин Е; Ангус Грей-Уил (31 октября 2015 г.) [31 октября 2015 г.]. «Гликоген печени у мышей с диабетом 2 типа случайным образом разветвляется в виде увеличенных агрегатов с притупленным высвобождением глюкозы». Гликоконъюгатный журнал . 33 . 41-51. дои : 10.1007/s10719-015-9631-5 . hdl : 11343/282927 .
- ^ Пэн Чжан; Шариф С. Нада; Синлэ Тан; Бин Дэн; Митчелл А. Салливан; Роберт Г. Гилберт (8 мая 2018 г.) [8 мая 2018 г.]. «Изучение биосинтеза гликогена с помощью моделирования Монте-Карло» . Международный журнал биологических макромолекул . 116 . 264-271. doi : 10.1016/j.ijbiomac.2018.05.027 .
- ^ Янг, ФГ (22 июня 1957 г.). «Клод Бернар и открытие гликогена» . Британский медицинский журнал . 1 (5033): 1431–1437. дои : 10.1136/bmj.1.5033.1431 . JSTOR 25382898 . ЧВК 1973429 . ПМИД 13436813 .
- ^ Нельсон, Д. (2013). Ленингерские принципы биохимии (6-е изд.). WH Фриман и компания. п. 618.
- ^ Страйер, Л. (1988). Биохимия (3-е изд.). Фриман. п. 451.
- ^ «Методика тренировки выносливости. Часть 1» . 30 октября 2009 г. Архивировано из оригинала 22 июля 2018 г. Проверено 1 августа 2013 г.
- ^ «Устойчивое состояние против темповых тренировок и сжигания жира» . 2 июня 2008 г. Архивировано из оригинала 5 сентября 2017 г. Проверено 1 августа 2013 г.
- ^ Макдональд, Лайл (25 июля 2012 г.). «Обзор исследования: углубленный анализ употребления углеводов на циклической кетогенной диете» . Архивировано из оригинала 11 ноября 2020 года . Проверено 19 февраля 2017 г.
- ^ Макдональд, Лайл (1998). Кетогенная диета: полное руководство для людей, сидящих на диете, и практикующих врачей . Лайл Макдональд.
- ^ Костилл Д.Л., Бауэрс Р., Бранам Дж., Спаркс К. (декабрь 1971 г.). «Утилизация мышечного гликогена при длительных тренировках в последующие дни». J Appl Physiol . 31 (6): 834–838. дои : 10.1152/яп.1971.31.6.834 . ПМИД 5123660 .
- ^ Зорзано А., Балон Т.В., Гудман М.Н., Рудерман Н.Б. (декабрь 1986 г.). «Истощение гликогена и повышение чувствительности к инсулину и реакции мышц после тренировки». Являюсь. Дж. Физиол . 251 (6, Часть 1): E664–E669. дои : 10.1152/ajpendo.1986.251.6.E664 . ПМИД 3538900 .
- ^ Макдональд, Лайл (2003). Абсолютная диета 2.0 . Лайл Макдональд.
- ^ Педерсен, диджей; Лессард, С.Дж.; Коффи, В.Г.; и др. (июль 2008 г.). «Высокие темпы ресинтеза мышечного гликогена после изнурительных упражнений, когда углеводы усваиваются вместе с кофеином». Журнал прикладной физиологии . 105 (1): 7–13. doi : 10.1152/japplphysicalol.01121.2007 . ПМИД 18467543 .
- ^ Jump up to: Перейти обратно: а б Билен, М.; Берк, LM; Гибала, MJ; ван Лун, LJC (декабрь 2010 г.). «Стратегии питания для содействия восстановлению после тренировки». Международный журнал спортивного питания и метаболизма при физических нагрузках . 20 (6): 515–532. дои : 10.1123/ijsnem.20.6.515 . ПМИД 21116024 . S2CID 13748227 .
- ^ Куинн А. Бесфорд; Франческа Кавальери; Фрэнк Карузо (7 мая 2020 г.) [16 октября 2019 г.]. «Гликоген как строительный блок для современных биологических материалов». Продвинутые материалы . 32 (18). 1904625. Бибкод : 2020AdM....3204625B . дои : 10.1002/adma.201904625 . hdl : 11343/230737 . ПМИД 31617264 . S2CID 204738366 .
Внешние ссылки [ править ]
- «Болезнь накопления гликогена» . Болезни Макардла .
- Гликоген Национальной медицинской библиотеки США по медицинским предметным рубрикам (MeSH)
Виды углеводов |
|---|