Гемицеллюлоза
Гемицеллюлоза ( матриксных (также известная как полиоза ) — один из ряда гетерополимеров полисахаридов), таких как арабиноксиланы , присутствующих наряду с целлюлозой почти во всех наземных растений клеточных стенках . [1] Целлюлоза кристаллическая, прочная и устойчивая к гидролизу . Гемицеллюлозы разветвлены, короче целлюлозы по длине, а также склонны к кристаллизации. [2] Они могут быть гидролизованы разбавленной кислотой или основанием, а также множеством ферментов гемицеллюлазы .
Состав
[ редактировать ]Известны различные виды гемицеллюлоз. Важные примеры включают ксилан , глюкуроноксилан , арабиноксилан , глюкоманнан и ксилоглюкан .
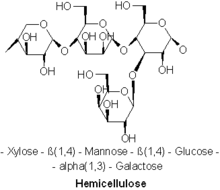
Гемицеллюлозы — это полисахариды , часто связанные с целлюлозой , но имеющие особый состав и структуру. В то время как целлюлоза получается исключительно из глюкозы , гемицеллюлозы состоят из различных сахаров и могут включать пятиуглеродные сахара ксилозу и арабинозу , шестиуглеродные сахара глюкозу, маннозу и галактозу и шестиуглеродный дезоксисахар рамнозу . Гемицеллюлозы содержат большую часть D- пентозных сахаров, а также иногда небольшое количество L-сахаров. Ксилоза в большинстве случаев представляет собой сахарный мономер, присутствующий в наибольшем количестве, хотя в хвойных породах сахар может быть наиболее распространенным. В гемицеллюлозе могут находиться не только обычные сахара, но и их подкисленные формы, например глюкуроновая кислота и галактуроновая кислота . [3] [4]
Структурное сравнение с целлюлозой
[ редактировать ]В отличие от целлюлозы гемицеллюлозы состоят из более коротких цепочек – 500–3000 сахарных единиц. Напротив, каждый полимер целлюлозы содержит 7 000–15 000 молекул глюкозы. [5] Кроме того, гемицеллюлозы могут быть разветвленными полимерами , а целлюлоза – неразветвленной. Гемицеллюлозы встроены в клеточные стенки растений, иногда в виде цепочек, образующих « землю » — они связываются с пектином целлюлозы, образуя сеть поперечно-сшитых волокон. [ нужна ссылка ]

На основании структурных различий, таких как основные связи и боковые группы, а также других факторов, таких как численность и распределение в растениях, гемицеллюлозы можно разделить на четыре группы: [4] 1) ксиланы, 2) маннаны ; 3) β-глюканы со смешанной связью ; 4) ксилоглюканы.
Ксиланы
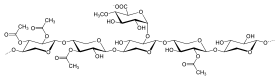
[ редактировать ]Ксиланы обычно состоят из основной цепи из β-(1→4)-связанных остатков ксилозы и могут быть далее разделены на гомоксиланы и гетероксиланы. Гомоксиланы имеют основу из остатков D-ксилопиранозы, связанных β(1→4) гликозидными связями. Гомоксиланы выполняют в основном структурные функции. Гетероксиланы, такие как глюкуроноксиланы, глюкуроноксиланы и сложные гетероксиланы, имеют основу из D-ксилопиранозы и короткие углеводные разветвления. Например, глюкуроноксилан имеет замену на α-(1→2)-связанные глюкуронозильные и 4-О-метилглюкуронозильные остатки. Арабиноксиланы и глюкуроноарабиноксиланы содержат остатки арабинозы, прикрепленные к основной цепи. [6]
Маннаны
[ редактировать ]Гемицеллюлозу маннанового типа можно разделить на два типа в зависимости от различия их основных цепей: галактоманнаны и глюкоманнаны. Галактоманнаны имеют только β-(1→4) связанные остатки D-маннопиранозы в линейных цепях. Глюкоманнаны состоят как из β-(1→4)-связанных остатков D-маннопиранозы, так и из β-(1→4)-связанных остатков D-глюкопиранозы в основных цепях. Что касается боковых цепей, остатки D-галактопиранозы имеют тенденцию быть 6-связанными с обоими типами в виде одиночных боковых цепей с различным количеством. [1]
β-глюканы со смешанной связью
[ редактировать ]Конформация глюкановых цепей со смешанными связями обычно содержит блоки β-(1→4) D-глюкопиранозы, разделенные одиночной β-(1→3) D-глюкопиранозой. Население β-(1→4) и β-(1→3) составляет около 70% и 30%. Эти глюканы в основном состоят из целлотриозильных (C 18 H 32 O 16 ) и целлотраозильных (C 24 H 42 O 21 ) сегментов в случайном порядке. Некоторые исследования показывают молярное соотношение целлотриозил/целлотраозил для овса (2,1–2,4), ячменя (2,8–3,3) и пшеницы (4,2–4,5). [1] [5]
Ксилоглюканы
[ редактировать ]Ксилоглюканы имеют основную цепь, аналогичную целлюлозе, с остатками α-D-ксилопиранозы в положении 6. Чтобы лучше описать различные боковые цепи, для каждого типа боковой цепи используется однобуквенное обозначение. G -- неразветвленный остаток Glc; X -- α-d-Ксил-(1→6)-Glc. L -- β-Гал, S -- α-1-Араф, F-- α-1-Фук. Это наиболее распространенные боковые цепи. [5]
Два наиболее распространенных типа ксилоглюканов в клеточных стенках растений идентифицируются как XXXG и XXGG. [1]
Биосинтез
[ редактировать ]клетки Гемицеллюлозы синтезируются из сахарных нуклеотидов в аппарате Гольджи . [8] Их синтез объясняют две модели: 1) «двухкомпонентная модель», в которой модификация происходит в двух трансмембранных белках, и 2) «однокомпонентная модель», в которой модификация происходит только в одном трансмембранном белке. После синтеза гемицеллюлозы транспортируются к плазматической мембране через пузырьки Гольджи.
Каждый вид гемицеллюлозы биосинтезируется специализированными ферментами. [8] [9]
Основы цепи маннана синтезируются с помощью семейства белков A, подобных целлюлозо-синтазе (CSLA), и, возможно, ферментов семейства D, подобных синтазы целлюлозы (CSLD). [8] [9] Маннансинтаза, особый фермент CSLA, отвечает за добавление маннозных единиц к основной цепи. [8] [9] Боковые цепи галактозы некоторых маннанов присоединяются галактоманнангалактозилтрансферазой. [8] [9] Ацетилирование маннанов опосредуется маннан-О-ацетилтрансферазой, однако этот фермент окончательно не идентифицирован. [9]
Синтез основной цепи ксилоглюкана опосредован целлюлозо-синтазоподобными белками семейства C (CSLC), в частности глюкансинтазой , которая добавляет звенья глюкозы к цепи. [8] [9] Синтез остова ксилоглюкана также каким-то образом опосредован ксилозилтрансферазой , но этот механизм отделен от его трансферазной функции и остается неясным. [9] Однако ксилозилтрансфераза в своей трансферазной функции используется для добавления ксилозы к боковой цепи. [8] [9] Другие ферменты, используемые для синтеза боковой цепи ксилоглюкана, включают галактозилтрансферазу (которая отвечает за добавление галактозы, две разные формы которой используются), фукозилтрансферазу (которая отвечает за добавление фукозы) и ацетилтрансферазу (которая отвечает за добавление фукозы). для ацетилирования). [8] [9]
Синтез основной цепи ксилана, в отличие от синтеза других гемицеллюлоз, не опосредован какими-либо белками, подобными целлюлозо-синтазе. [9] Вместо этого за синтез основной цепи отвечает ксилансинтаза, способствуя добавлению ксилозы. [9] Идентифицировано несколько генов ксилансинтаз. [9] Для добавления и модификации звеньев боковой цепи ксилана используются несколько других ферментов, в том числе глюкуронозилтрансфераза (которая добавляет звенья глюкуроновой кислоты), ксилозилтрансфераза (которая добавляет дополнительные звенья ксилозы), арабинозилтрансфераза (которая добавляет арабинозу), метилтрансфераза (отвечает за метилирование ) и ацетилтрансфераза] (отвечает за ацетилирование). [9] Учитывая, что глюкан со смешанными связями представляет собой неразветвленный гомополимер глюкозы, синтез боковой цепи отсутствует, а только присоединение глюкозы к основной цепи по двум связям: β1-3 и β1-4. [9] Синтез основной цепи опосредуется ферментами семейств белков F и H, подобных целлюлозо-синтазе (CSLF и CSLH), в частности глюкансинтазой. [8] [9] Было идентифицировано несколько форм глюкансинтазы из CSLF и CSLH. [8] [9] Все они отвечают за добавление глюкозы к основной цепи и все способны производить связи как β1-3, так и β1-4, однако неизвестно, какой вклад каждый конкретный фермент вносит в распределение β1-3 и β1-4. связи. [8] [9]
Приложения
[ редактировать ]В процессе сульфитной целлюлозы гемицеллюлоза в значительной степени гидролизуется кислым варочным щелоком и попадает в коричневый щелок, где сбраживаемые гексозные сахара (около 2%) могут быть использованы для производства этанола . Этот процесс в первую очередь применялся к коричневым щелокам сульфита кальция. [10]
Арабиногалактаны могут использоваться в качестве эмульгаторов , стабилизаторов и связующих в соответствии с Федеральным законом о пищевых продуктах, лекарствах и косметике . Арабиногалактаны также можно использовать в качестве связующего вещества в подсластителях . [11]
- Ксилан
Пленки на основе ксилана обладают низкой кислородопроницаемостью и поэтому представляют потенциальный интерес в качестве упаковки для чувствительных к кислороду продуктов. [12]
Агар используется при приготовлении желе и пудингов. Это также питательная среда с другими питательными веществами для микроорганизмов . [13]

Курдлан можно использовать в качестве заменителя жира для производства диетических продуктов, сохраняя при этом вкус и ощущение во рту настоящих жиросодержащих продуктов. [13]
b-глюканы играют важную роль в пищевых добавках, в то время как b-глюканы также перспективны в вопросах, связанных со здоровьем, особенно в иммунных реакциях и лечении рака. [14]
Ксантан вместе с другими полисахаридами может образовывать гели с высокой вязкостью раствора , которые можно использовать в нефтяной промышленности для загущения бурового раствора. В пищевой промышленности ксантан используется в таких продуктах, как заправки и соусы. [15]
Альгинат играет важную роль в разработке антимикробного текстиля благодаря своим характеристикам экологичности и высокому уровню индустриализации в качестве устойчивого биополимера . [16]
Естественные функции
[ редактировать ]
Будучи полисахаридным соединением в стенках растительных клеток, подобным целлюлозе, гемицеллюлоза помогает целлюлозе укреплять стенки растительных клеток. [6] Гемицеллюлоза взаимодействует с целлюлозой , обеспечивая поперечное сшивание целлюлозы микрофибрилл : гемицеллюлоза будет искать пустоты в клеточной стенке во время ее формирования и обеспечивать поддержку вокруг фибрилл целлюлозы , чтобы придать клеточной стенке максимально возможную прочность, которую она может обеспечить. [6] Гемицеллюлоза доминирует в средней пластинке растительной клетки, в отличие от целлюлозы , которая в основном находится во вторичных слоях. Это позволяет гемицеллюлозе обеспечивать промежуточную поддержку целлюлозы на внешних слоях растительной клетки. В некоторых клеточных стенках гемицеллюлоза также взаимодействует с лигнином , обеспечивая структурную поддержку тканей большего количества сосудистых растений. [3] [17]
Добыча
[ редактировать ]Существует много способов получения гемицеллюлозы; все они основаны на методах экстракции из деревьев лиственных или хвойных пород, измельченных на более мелкие образцы. В лиственных породах основным экстрактом гемицеллюлозы является глюкуронокслян (ацетилированные ксиланы), а в хвойных породах содержится галактоглюкоманнан. [18] [19] Перед экстракцией древесину обычно необходимо измельчить в щепу разного размера в зависимости от используемого реактора. После этого используется процесс экстракции горячей водой, также известный как автогидролиз или гидротермальная обработка, с добавлением кислот и оснований для изменения размера выхода и свойств. [18] [19] Основным преимуществом экстракции горячей водой является то, что она предлагает метод, при котором единственным необходимым химическим веществом является вода, что делает его экологически чистым и дешевым. [20]
Целью обработки горячей водой является удаление как можно большего количества гемицеллюлозы из древесины. Это делается посредством гидролиза гемицеллюлозы для получения более мелких олигомеров и ксилозы. Ксилоза при обезвоживании превращается в фурфурол. [21] Когда ксилоза и фурфурол [ проверьте орфографию ] Целью добавления кислотных катализаторов, таких как муравьиная кислота, является увеличение перехода полисахарида в моносахариды. Также было показано, что этот катализатор также использует эффект растворителя для облегчения реакции. [21]
Один из методов предварительной обработки — пропитка древесины разбавленными кислотами (концентрацией около 4%). Это превращает гемицеллюлозу в моносахариды. Предварительная обработка основаниями (например, гидроксидом натрия или калия) разрушает структуру лигнина. [19] Это меняет структуру с кристаллической на аморфную. Еще одним методом является гидротермальная предварительная обработка. [ нужны дальнейшие объяснения ] Это дает такие преимущества, как отсутствие необходимости в токсичных или коррозийных растворителях, специальных реакторах и дополнительных затратах на утилизацию опасных химикатов. [18]
Процесс экстракции горячей водой осуществляется в реакторах периодического действия, полунепрерывных реакторах или суспензионных реакторах непрерывного действия. Для реакторов периодического и полунепрерывного действия образцы древесины могут использоваться в таких условиях, как щепа или гранулы, в то время как шламовый реактор должен содержать частицы размером от 200 до 300 микрометров. [19] Хотя размер частиц уменьшается, выход продукции также снижается. [22] Это связано с увеличением клетчатки. [ нужна ссылка ]
Процесс с использованием горячей воды осуществляется при температуре от 160 до 240 градусов Цельсия, чтобы поддерживать жидкую фазу. Это делается при температуре выше нормальной температуры кипения воды, чтобы увеличить солюбилизацию гемицеллюлозы и деполимеризацию полисахаридов. [21] Этот процесс может занять от нескольких минут до нескольких часов в зависимости от температуры и pH системы. [19] Более высокие температуры в сочетании с более длительным временем экстракции приводят к более высоким выходам. Максимальный выход достигается при pH 3,5. [18] Если ниже, выход экстракции экспоненциально снижается. Для контроля pH обычно добавляют бикарбонат натрия. [18] Бикарбонат натрия ингибирует автолиз ацетильных групп, а также ингибирует гликозильные связи. В зависимости от температуры и времени гемицеллюлоза может далее превращаться в олигомеры, мономеры и лигнин. [18]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а б с д Шеллер Г.В., Ульвсков П. Гемицеллюлозы. // Годовой ред. Растительная биол. 2010;61:263-89. doi: 10.1146/annurev-arplant-042809-112315 .
- ^ Смит, Питер Дж.; Карри, Томас М.; Ян, Чон-Йе; Барнс, Уильям Дж.; Зиглер, Саманта Дж.; Миттал, Ашутош; Моремен, Келли В.; Йорк, Уильям С.; Бомбл, Янник Дж.; Пенья, Мария Дж.; Урбанович, Брианна Р. (13 июля 2022 г.). «Ферментативный синтез микрочастиц ксилана с настраиваемой морфологией» . ACS Материалы Au . 2 (4): 440–452. doi : 10.1021/acsmaterialsau.2c00006 . ISSN 2694-2461 . ПМЦ 9284610 . ПМИД 35856073 .
- ^ Jump up to: а б Эбрингер, Анна; Громадкова, Зденка; Хайнце, Томас (2005), Хайнце, Томас (редактор), «Гемицеллюлоза», Полисахариды I: структура, характеристика и использование , Достижения в области науки о полимерах, Springer, стр. 1–67, номер домена : 10.1007/b136816 , ISBN. 978-3-540-31583-4
- ^ Jump up to: а б Хайнце, Томас (2005). Полисахариды I: строение, характеристика и применение . Берлин: Шпрингер. ISBN 978-3-540-31583-4 . OCLC 262681325 .
- ^ Jump up to: а б с Гибсон ЖЖ (2013). «Иерархическая структура и механика растительного сырья» . Журнал интерфейса Королевского общества . 9 (76): 2749–2766. дои : 10.1098/rsif.2012.0341 . ПМЦ 3479918 . ПМИД 22874093 .
- ^ Jump up to: а б с Шеллер, Хенрик Вибе; Ульвсков, Петр (2 июня 2010 г.). «Гемицеллюлозы». Ежегодный обзор биологии растений . 61 (1): 263–289. doi : 10.1146/annurev-arplant-042809-112315 . ISSN 1543-5008 . ПМИД 20192742 .
- ^ Нимц, Хорст Х; Шмитт, Уве; Шваб, Эккарт; Виттманн, Отто; Вольф, Франц (15 июня 2000 г.), «Древесина», Wiley-VCH Verlag GmbH & Co. KGaA (редактор), Энциклопедия промышленной химии Ульмана , Wiley-VCH Verlag GmbH & Co. КГаА, стр. 101-1. a28_305, номер домена : 10.1002/14356007.a28_305 , ISBN 978-3-527-30673-2
- ^ Jump up to: а б с д и ж г час я дж к Чжу, Сяоюй; Синь, Сяорань; Гу, Ин (2019), Коэн, Ефрем; Мерцендорфер, Ганс (ред.), «Синтез целлюлозы и гемицеллюлозы и их регуляция в растительных клетках», Матрицы внеклеточных биополимеров на основе сахара , том. 12, Springer International Publishing, стр. 303–353, номер номера : 10.1007/978-3-030-12919-4_7 , ISBN. 978-3-030-12918-7 , S2CID 198238237
- ^ Jump up to: а б с д и ж г час я дж к л м н тот п д Поли, Маркус; Гилле, Саша; Лю, Лифэн; Мансури, Насим; де Соуза, Амансио; Шультинк, Алекс; Сюн, Гуанъянь (2013). «Биосинтез гемицеллюлозы». Планта . 238 (4): 627–642. дои : 10.1007/s00425-013-1921-1 . ISSN 0032-0935 . ПМИД 23801299 . S2CID 17501948 .
- ^ «Сульфитный процесс — обзор | Темы ScienceDirect» . www.sciencedirect.com . Проверено 8 сентября 2022 г.
- ^ Уистлер, РЛ (1993). Гемицеллюлозы. В «Промышленных резинках» (стр. 295–308). Эльзевир. https://doi.org/10.1016/b978-0-08-092654-4.50015-2
- ^ Грёндаль, М., и Гатенхольм, П. (2007). Кислородные барьерные пленки на основе ксиланов, выделенных из биомассы. В серии симпозиумов ACS (стр. 137–152). Американское химическое общество. https://doi.org/10.1021/bk-2007-0954.ch009
- ^ Jump up to: а б Спиридон И. и Попа В.И. (2008). Гемицеллюлозы: основные источники, свойства и применение. В книге «Мономеры, полимеры и композиты из возобновляемых ресурсов» (стр. 289–304). Эльзевир. https://doi.org/10.1016/b978-0-08-045316-3.00013-2
- ^ Ветвицка В., Ваннуччи Л., Сима П. и Рихтер Дж. (2019). Бета-глюкан: добавка или лекарство? От лаборатории к клиническим испытаниям. Молекулы, 24(7), 1251. https://doi.org/10.3390/molecules24071251.
- ^ Наваррете, Р.К., Хаймс, Р.Э. и Сехолт, Дж.М. (2000). Применение ксантановой камеди для контроля потери жидкости и связанного с этим повреждения пласта. Конференция SPE по добыче нефти и газа в Пермском бассейне. Конференция SPE по добыче нефти и газа в Пермском бассейне. https://doi.org/10.2118/59535-ms
- ^ Ли Дж., Хэ Дж. и Хуан Ю. (2017). Роль альгината в антибактериальной отделке текстиля. Международный журнал биологических макромолекул, 94, 466–473. https://doi.org/10.1016/j.ijbiomac.2016.10.054
- ^ Плаза, Меришель; Тернер, Шарлотта (2015). «Экстракция биоактивных веществ горячей водой под давлением» . Тенденции в аналитической химии . 71 : 39–54. дои : 10.1016/j.trac.2015.02.022 .
- ^ Jump up to: а б с д и ж Галлина, Джанлука; Кабеса, Альваро; Гренман, Хенрик; Бьязи, Пьердоменико; Гарсиа-Серна, Хуан; Салми, Тапио (01 марта 2018 г.). «Экстракция гемицеллюлозы предварительной обработкой горячей водой под давлением при 160 ° C для 10 различных пород древесины: выход и молекулярная масса» . Журнал сверхкритических жидкостей . Фракционирование биомассы докритической и сверхкритической водой. 133 : 716–725. дои : 10.1016/j.supflu.2017.10.001 . ISSN 0896-8446 .
- ^ Jump up to: а б с д и Ли З, Цинь М, Сюй С и Чен X (2013). «Горячая экстракция гемицеллюлозы из осиновой щепы разной крупности :: БиоРесурсы» . bioresources.cnr.ncsu.edu . Проверено 24 апреля 2020 г.
{{cite web}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Джанлука Галлинаа Альваро Кабесаа Хенрик Гренманbc Пьердоменико Биасибд Хуан Гарсиа-Сернаа Тапио Салми (март 2018 г.). «Экстракция гемицеллюлозы предварительной обработкой горячей водой под давлением при 160 ° C для 10 различных пород древесины: выход и молекулярная масса» (PDF) . Журнал сверхкритических жидкостей . 133 (Часть 2): 716–725. дои : 10.1016/j.supflu.2017.10.001 . Проверено 7 октября 2017 г.
- ^ Jump up to: а б с Тунч, М. Сефик; ван Хайнинген, Адриан Р.П. (17 сентября 2008 г.). «Экстракция гемицеллюлозы из смешанной древесины южных лиственных пород водой при 150 ° C: влияние времени». Исследования в области промышленной и инженерной химии . 47 (18): 7031–7037. дои : 10.1021/ie8007105 . ISSN 0888-5885 .
- ^ Айрилмис Н., Квон Дж., Хан Т. (октябрь 2017 г.). «Влияние размера древесной щепы на экстракцию гемицеллюлозы и технологические свойства ДСП» . Турецкий журнал сельского и лесного хозяйства . 41 : 331–337. дои : 10.3906/tar-1704-63 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка )
Внешние ссылки
[ редактировать ]- Структура и свойства гемицеллюлозы / Урок химии древесины Дэвида Ванга
Виды углеводов |
|---|