Целлюлаза
| Целлюлаза | |||
|---|---|---|---|
 Фермент целлюлаза, продуцируемый Thermomonospora fusca , с целлотриозой, связанной в неглубокой бороздке каталитического домена. | |||
| Идентификаторы | |||
| Номер ЕС. | 3.2.1.4 | ||
| Номер CAS. | 9012-54-8 | ||
| Базы данных | |||
| ИнтЭнк | вид IntEnz | ||
| БРЕНДА | БРЕНДА запись | ||
| Экспаси | Просмотр NiceZyme | ||
| КЕГГ | КЕГГ запись | ||
| МетаЦик | метаболический путь | ||
| ПРЯМОЙ | профиль | ||
| PDB Структуры | RCSB PDB PDBe PDBsum | ||
| Генная онтология | АмиГО / QuickGO | ||
| |||

Целлюлаза ( EC 3.2.1.4 ; систематическое название 4-β- D -глюкан-4-глюканогидролаза ) — это любой из нескольких ферментов, вырабатываемых главным образом грибами , бактериями и простейшими , которые катализируют целлюлолиз , разложение целлюлозы и некоторых родственных полисахаридов :
- Эндогидролиз (1→4)-β- D -глюкозидных связей в целлюлозе, лихенине и β- D -глюкане злаков.
Это название также используется для любой встречающейся в природе смеси или комплекса различных таких ферментов, которые действуют последовательно или синергически, разлагая целлюлозный материал.
Целлюлазы расщепляют молекулу целлюлозы на моносахариды («простые сахара»), такие как β- глюкоза , или более короткие полисахариды и олигосахариды . Расщепление целлюлозы имеет большое экономическое значение, поскольку оно делает основной компонент растений доступным для потребления и использования в химических реакциях. Специфической реакцией является гидролиз 1,4-β- D - гликозидных связей в целлюлозе, гемицеллюлозе , лихенине и β -D -глюканах злаков . Поскольку молекулы целлюлозы прочно связываются друг с другом, целлюлолиз относительно сложен по сравнению с расщеплением других полисахаридов, таких как крахмал. [2]
Большинство млекопитающих имеют очень ограниченную способность самостоятельно переваривать пищевые волокна, такие как целлюлоза. У многих травоядных животных, таких как жвачные животные , такие как крупный рогатый скот и овцы, а также ферментаторов задней кишки , таких как лошади, целлюлазы продуцируются симбиотическими бактериями. Эндогенные целлюлазы производятся некоторыми видами животных , например, некоторыми термитами , улитками, [3] [4] [5] и дождевые черви .
Недавно целлюлазы были также обнаружены в зеленых микроводорослях ( Chlamydomonas reinhardtii , Gonium pectorale и Volvox carteri ), и их каталитические домены (CD), принадлежащие к семейству GH9 , демонстрируют высочайшую гомологию последовательностей с эндогенными целлюлазами многоклеточных животных. Водорослевые целлюлазы имеют модульную структуру и состоят из предполагаемых новых, богатых цистеиномуглеводсвязывающие модули (CBM), линкеры, богатые пролином/серином (PS), в дополнение к предполагаемым Ig-подобным и неизвестным доменам у некоторых членов. Целлюлаза из Gonium pectorale состояла из двух CD, разделенных линкерами, и с C-концевым CBM. [6]
Известно несколько различных видов целлюлаз, которые различаются структурно и механически. Синонимы, производные и специфические ферменты, связанные с названием «целлюлаза», включают эндо-1,4-β -D -глюканазу (β-1,4-глюканаза, β-1,4-эндоглюкангидролаза, эндоглюканаза D, 1,4 -(1,3;1,4)-β- D- глюкан-4-глюканогидролаза), карбоксиметилцеллюлаза (CMCase), авицелаза, целлудекстриназа , целлюлаза А , целлюлозин AP , щелочная целлюлаза , целлюлаза А3 , 9,5-целлюлаза , целоксиланаза и панцеллаза SS . Ферменты, расщепляющие лигнин, иногда называют целлюлазами, но это старое использование устарело; они представляют собой ферменты, модифицирующие лигнин .
Виды и действие
[ редактировать ]Пять основных типов целлюлаз в зависимости от типа катализируемой реакции:
- Эндоцеллюлазы (EC 3.2.1.4) случайным образом расщепляют внутренние связи в аморфных участках, создавая новые концы цепей.
- Экзоцеллюлазы или целлобиогидролазы (EC 3.2.1.91) отщепляют две-четыре единицы с концов открытых цепей, продуцируемых эндоцеллюлазой, в результате чего образуются тетрасахариды . [7] или дисахариды , такие как целлобиоза . Экзоцеллюлазы подразделяются на тип I, которые действуют процессивно с восстанавливающего конца целлюлозной цепи, и тип II, которые действуют процессивно с невосстанавливающего конца.
- Целлобиазы (EC 3.2.1.21) или β-глюкозидазы гидролизуют продукт экзоцеллюлазы на отдельные моносахариды.
- Окислительные целлюлазы деполимеризуют целлюлозу посредством радикальных реакций, как, например, целлобиозодегидрогеназа (акцептор) .
- Целлюлофосфорилазы деполимеризуют целлюлозу, используя фосфаты вместо воды.
Авицелаза обладает почти исключительно экзоцеллюлазной активностью, поскольку авицел представляет собой высокомикрокристаллический субстрат.
Внутри вышеуказанных типов существуют также прогрессивные (также известные как процессивные) и непрогрессивные типы. Прогрессивная целлюлаза будет продолжать взаимодействовать с одной цепью полисахарида, непрогрессивная целлюлаза будет взаимодействовать один раз, а затем отделиться и задействовать другую цепь полисахарида.
Действие целлюлазы считается синергичным, поскольку все три класса целлюлаз могут дать гораздо больше сахара, чем добавление всех трех по отдельности. За исключением жвачных животных, большинство животных (включая человека) не производят целлюлазу в своем организме и могут лишь частично расщеплять целлюлозу посредством ферментации, что ограничивает их способность использовать энергию из волокнистого растительного материала.
Структура
[ редактировать ]Большинство грибных целлюлаз имеют двухдоменную структуру: один каталитический домен и один целлюлозосвязывающий домен, которые соединены гибким линкером. Эта структура адаптирована для работы с нерастворимым субстратом и позволяет ферменту двумерно диффундировать по поверхности, подобно гусенице. Однако существуют также целлюлазы (в основном эндоглюканазы), у которых отсутствуют домены, связывающие целлюлозу.
Как связывание субстратов, так и катализ зависят от трехмерной структуры фермента, которая возникает вследствие уровня сворачивания белка . Аминокислотная последовательность и расположение их остатков, которые встречаются в активном центре, положение, в котором связывается субстрат, могут влиять на такие факторы, как аффинность связывания лигандов, стабилизация субстратов в активном центре и катализ. Структура субстрата дополняет точную структуру активного центра фермента. Изменения положения остатков могут привести к искажению одного или нескольких из этих взаимодействий. [8] Дополнительные факторы, такие как температура, pH и ионы металлов, влияют на нековалентные взаимодействия между структурами ферментов. [9] Виды Thermotoga maritima производят целлюлазы, состоящие из 2 β-листов (белковых структур), окружающих центральную каталитическую область, которая является активным центром. [10] Фермент относится к категории эндоглюканазы, которая изнутри расщепляет β-1,4-гликозидные связи в цепях целлюлозы, способствуя дальнейшей деградации полимера. Различные виды одного семейства с T. maritima производят целлюлазы разной структуры. [10] Целлюлазы, продуцируемые видом Coprinopsis cinerea, состоят из семи белковых нитей в форме закрытого туннеля, называемого β/α-цилиндром. [11] Эти ферменты гидролизуют субстрат карбоксиметилцеллюлозу. Связывание субстрата в активном центре вызывает изменение конформации, которое приводит к деградации молекулы.
Целлюлазные комплексы
[ редактировать ]У многих бактерий целлюлазы in vivo представляют собой сложные ферментные структуры, организованные в супрамолекулярные комплексы — целлюлосомы . Они могут содержать, помимо прочего, пять различных ферментативных субъединиц, а именно эндоцеллюлазы, экзоцеллюлазы, целлобиазы, окислительные целлюлазы и целлюлозофосфорилазы, причем только экзоцеллюлазы и целлобиазы участвуют в фактическом гидролизе связи β(1→4). Количество субъединиц, составляющих целлюлосомы, также может определять скорость активности фермента. [12]
Мультидоменные целлюлазы широко распространены среди многих таксономических групп, однако целлюлазы анаэробных бактерий, обнаруженные в целлюлосомах, имеют наиболее сложную архитектуру, состоящую из разных типов модулей. Например, Clostridium cellulolyticum продуцирует 13 модульных целлюлаз GH9, содержащих различное количество и расположение каталитического домена (CD), углеводсвязывающего модуля (CBM), докерина, линкера и Ig-подобного домена. [13]
Целлюлазный комплекс Trichoderma reesei , например, включает компонент, обозначенный C1 (57 000 дальтон ), который разделяет цепи кристаллической целлюлозы, эндоглюканазы (около 52 000 дальтон), экзоглюканазы (около 61 000 дальтон) и β-глюкозидазы (76 000 дальтон). дальтоны). [14]
Многочисленные «подписные» последовательности, известные как докерины и когезины, были идентифицированы в геномах бактерий, продуцирующих целлюлосомы. В зависимости от аминокислотной последовательности и третичной структуры целлюлазы делятся на кланы и семейства. [15]
Мультимодулярные целлюлазы более эффективны, чем свободный фермент (только с ЦД) благодаря синергизму из-за непосредственной близости между ферментом и целлюлозным субстратом. CBM участвуют в связывании целлюлозы, тогда как гликозилированные линкеры обеспечивают гибкость CD для более высокой активности и защиты от протеаз, а также повышенного связывания с поверхностью целлюлозы. [6]
Механизм целлюлолиза
[ редактировать ]

Использование
[ редактировать ]Целлюлаза используется в коммерческой пищевой промышленности при производстве кофе . Осуществляет гидролиз целлюлозы при сушке бобов . Кроме того, целлюлазы широко используются в текстильной промышленности и в стиральных порошках. Их также использовали в целлюлозно-бумажной промышленности для различных целей и даже в фармацевтических целях.Целлюлаза используется при ферментации биомассы в биотопливо , хотя в настоящее время этот процесс является относительно экспериментальным. Целлюлаза используется в медицине для лечения фитобезоаров, формы целлюлозного безоара, человека обнаруженного в желудке , и она продемонстрировала эффективность в разложении полимикробных бактериальных биопленок путем гидролиза β(1-4) гликозидных связей в структурных матриксных экзополисахаридах внеклеточное полимерное вещество (ЭПС). [17] [18]
Измерение
[ редактировать ]Поскольку нативный субстрат, целлюлоза , представляет собой водонерастворимый полимер, традиционные анализы восстанавливающего сахара с использованием этого субстрата не могут быть использованы для измерения активности целлюлазы. Ученые-аналитики разработали ряд альтернативных методов.
- Метод DNSA Активность целлюлазы определяли путем инкубации 0,5 мл надосадочной жидкости с 0,5 мл 1% карбоксиметилцеллюлозы (КМЦ) в 0,05М цитратном буфере (рН 4,8) при 50°С в течение 30 минут. Реакцию останавливали добавлением 3 мл реагента динитросалициловой кислоты. Поглощение считывали при 540 нм. [19]
Вискозиметр при можно использовать для измерения снижения вязкости раствора, содержащего водорастворимое производное целлюлозы, такое как карбоксиметилцеллюлоза, инкубации с образцом целлюлазы. [20] Снижение вязкости прямо пропорционально активности целлюлазы. Хотя такие анализы очень чувствительны и специфичны для эндоцеллюлазы ( энзимы экзо -действующей целлюлазы практически не вызывают изменений вязкости), они ограничены тем фактом, что трудно определить активность в обычных единицах фермента (микромолях гидролизованного или гидролизованного субстрата). продукции, производимой в минуту).
Целлоолигосахаридные субстраты
[ редактировать ]Целлоолигосахариды с более низкой DP (DP2-6) достаточно растворимы в воде, чтобы выступать в качестве жизнеспособных субстратов для ферментов целлюлазы. [21] Однако, поскольку эти субстраты сами по себе являются « восстанавливающими сахарами », они не подходят для использования в традиционных анализах восстанавливающих сахаров, поскольку дают высокие «холостые» значения. Однако их гидролиз, опосредованный целлюлазой, можно контролировать с помощью методов ВЭЖХ или IC, чтобы получить ценную информацию о потребностях в субстрате конкретного фермента целлюлазы.
Восстановленные целлоолигосахаридные субстраты
[ редактировать ]Целлоолигосахариды можно химически восстановить под действием боргидрида натрия с образованием соответствующих сахарных спиртов . Эти соединения не реагируют при анализе восстановления сахара, но продукты их гидролиза реагируют. Это делает целлоолигосахариды, восстановленные боргидридом, ценными субстратами для анализа целлюлазы с использованием традиционных анализов восстанавливающих сахаров, таких как метод Нельсона-Симоджи. [22] [23]
Окрашенные полисахаридные субстраты
[ редактировать ]Эти субстраты можно разделить на два класса:
- Нерастворимые хромогенные субстраты: нерастворимый субстрат целлюлазы, такой как AZCL-HE-целлюлоза, поглощает воду с образованием желеобразных частиц при помещении в раствор. Этот субстрат постепенно деполимеризуется и солюбилизируется под действием целлюлазы. Реакцию останавливают добавлением щелочного раствора для остановки активности фермента и реакционную суспензию фильтруют или центрифугируют. Измеряется цвет фильтрата или надосадочной жидкости, который может быть связан с активностью фермента.
- Растворимые хромогенные субстраты: образец целлюлазы инкубируют с водорастворимым субстратом, таким как азо-CM-целлюлоза, реакцию прекращают и высокомолекулярные, частично гидролизованные фрагменты осаждают из раствора органическим растворителем, таким как этанол или метоксиэтанол. Суспензию тщательно перемешивают, центрифугируют и измеряют цвет надосадочного раствора (за счет мелких растворимых окрашенных фрагментов). С помощью стандартной кривой можно определить активность фермента.
Реагенты, связанные с ферментами
[ редактировать ]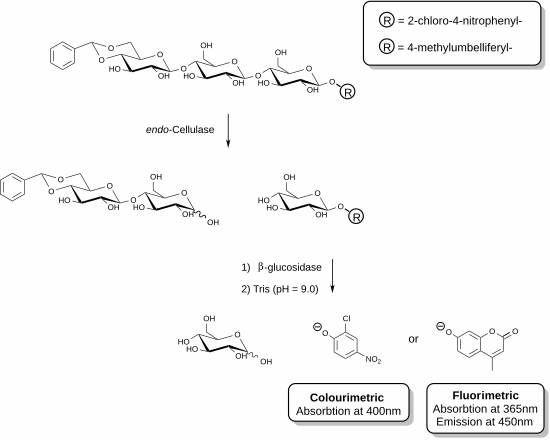
Недавно были разработаны новые реагенты, позволяющие специфически измерять эндоцеллюлазу . [25] [26] Эти методы включают использование функционализированных олигосахаридных субстратов в присутствии вспомогательного фермента. В показанном примере фермент целлюлаза способен распознавать трисахаридный фрагмент целлюлозы и расщеплять эту единицу. Вспомогательный фермент, присутствующий в смеси реагентов (β-глюкозидаза), затем гидролизует фрагмент, содержащий хромофор или флуорофор. Анализ завершается добавлением основного раствора, который останавливает ферментативную реакцию и депротонирует высвобожденное фенольное соединение с образованием фенолятных соединений. Целлюлазная активность данного образца прямо пропорциональна количеству высвободившегося фенолята, которое можно измерить с помощью спектрофотометра. Ацетальная функционализация на невосстанавливающем конце трисахаридного субстрата предотвращает действие вспомогательной β-глюкозидазы на исходный субстрат.
См. также
[ редактировать ]- Целлюлозо-1,4-бета-целлобиозидаза , эффективная целлюлаза.
- Целлюлазная единица — единица измерения активности целлюлазы.
Ссылки
[ редактировать ]- ^ PDB : 1NLR ; Сульценбахер Дж., Шарек Ф., Морозоли Р., Дюпон К., Дэвис Г.Дж. (декабрь 1997 г.). « Эндоглюканаза семейства 12 Streptomyces lividans : построение каталитического ядра, экспрессия и рентгеновская структура при разрешении 1,75 Å». Биохимия . 36 (51): 16032–9. дои : 10.1021/bi972407v . ПМИД 9440876 . ; визуализируется с помощью PyMOL
- ^ Баркалов Д.Г., Уистлер Р.Л. «Целлюлоза» . AccessScience, МакГроу-Хилл. [ постоянная мертвая ссылка ]
- ^ Бигнелл Д.Э., Ройзин Ю., Ло Н. (2011). Биология термитов: современный синтез . Дордрехт: Спрингер. ISBN 978-9048139767 .
- ^ Ватанабэ Х., Нода Х., Токуда Г., Ло Н. (июль 1998 г.). «Ген целлюлазы термитного происхождения». Природа . 394 (6691): 330–1. Бибкод : 1998Natur.394..330W . дои : 10.1038/28527 . ПМИД 9690469 . S2CID 4384555 .
- ^ Ватанабэ Х., Токуда Г. (август 2001 г.). «Животные целлюлазы». Клеточные и молекулярные науки о жизни . 58 (9): 1167–78. дои : 10.1007/PL00000931 . ПМИД 11577976 . S2CID 570164 .
- ^ Jump up to: а б Геррьеро, Хеа; сержант Кьелл; Легай, Сильвен; Хаусман, Жан-Франсуа; Коши, Анри-Мишель; Ахмад, Иршад; Сиддики, Хавар (15 июня 2018 г.). «Новые результаты сравнительного анализа целлюлаз зеленых микроводорослей in Silico» . Международный журнал молекулярных наук . 19 (6): 1782. doi : 10.3390/ijms19061782 . ISSN 1422-0067 . ПМК 6032398 . ПМИД 29914107 .
- ^ Зверлов В.В., Шанц Н., Шварц В.Х. (август 2005 г.). «Основным новым компонентом целлюлозомы Clostridium thermocellum является процессивная эндо-β-1,4-глюканаза, продуцирующая целлотетраозу» . Письма FEMS по микробиологии . 249 (2): 353–8. дои : 10.1016/j.femsle.2005.06.037 . ПМИД 16006068 .
- ^ Пейн CM, Bomble YJ, Тейлор CB, Маккейб C, Химмель ME, Кроули MF, Бекхэм GT (ноябрь 2011 г.). «Множественные функции взаимодействий ароматических углеводов в процессивной целлюлазе, исследованные с помощью молекулярного моделирования» . Журнал биологической химии . 286 (47): 41028–35. дои : 10.1074/jbc.M111.297713 . ПМК 3220501 . ПМИД 21965672 .
- ^ Ли Ю.Дж., Ким Б.К., Ли Б.Х., Джо КИ, Ли Н.К., Чанг Ч. и др. (январь 2008 г.). «Очистка и характеристика целлюлазы, продуцируемой Bacillus amyoliquefaciens DL-3, с использованием рисовой шелухи». Биоресурсные технологии . 99 (2): 378–86. doi : 10.1016/j.biortech.2006.12.013 . ПМИД 17320379 .
- ^ Jump up to: а б Ченг Ю.С., Ко Т.П., Ву Т.Х., Ма Ю., Хуан Ч.Х., Лай Х.Л. и др. (апрель 2011 г.). «Кристаллическая структура и способ связывания субстрата целлюлазы 12А из Thermotoga maritima ». Белки . 79 (4): 1193–204. дои : 10.1002/прот.22953 . ПМИД 21268113 . S2CID 23572933 .
- ^ Лю Ю, Ёсида М, Кураката Ю, Миядзаки Т, Игараси К, Самедзима М и др. (март 2010 г.). «Кристаллическая структура фермента семейства гликозидгидролаз 6, CcCel6C, целлюлазы, конститутивно продуцируемой Coprinopsis cinerea » . Журнал ФЭБС . 277 (6): 1532–42. дои : 10.1111/j.1742-4658.2010.07582.x . ПМИД 20148970 . S2CID 6338050 .
- ^ Цай С.Л., ДаСильва Н.А., Чен В. (январь 2013 г.). «Функциональное отображение сложных целлюлосом на поверхности дрожжей посредством адаптивной сборки». ACS Синтетическая биология . 2 (1): 14–21. CiteSeerX 10.1.1.701.5515 . дои : 10.1021/sb300047u . ПМИД 23656322 .
- ^ Равашоль Дж., Борн Р., Тардиф К., де Филип П., Фиеробе Х.П. (март 2014 г.). «Характеристика всех гликозидгидролаз семейства 9, синтезируемых бактерией Clostridium cellulolyticum, продуцирующей целлюлозу » . Журнал биологической химии . 289 (11): 7335–48. дои : 10.1074/jbc.M113.545046 . ПМЦ 3953250 . ПМИД 24451379 .
- ^ Worthington Biochemical Corporation (2014), Целлюлаза . Доступ: 3 июля 2014 г.
- ^ Байер Э.А., Чанзи Х., Ламед Р., Шохам Ю. (октябрь 1998 г.). «Целлюлоза, целлюлазы и целлюлосомы». Современное мнение в области структурной биологии . 8 (5): 548–57. дои : 10.1016/S0959-440X(98)80143-7 . ПМИД 9818257 .
- ^ Бхаумик, Прасенджит; Дхепе, Пареш Лакшмикант (1 января 2015 г.). «Глава 1. Преобразование биомассы в сахара» . Сахара из биомассы для нетопливных применений . Серия «Зеленая химия». Королевское химическое общество. стр. 1–53. дои : 10.1039/9781782622079-00001 . ISBN 978-1-78262-113-3 .
- ^ Флеминг Д., Рамбо К.П. (апрель 2017 г.). «Подходы к диспергированию медицинских биопленок» . Микроорганизмы . 5 (2): 15. doi : 10.3390/microorganisms5020015 . ПМК 5488086 . ПМИД 28368320 .
- ^ Флеминг Д., Чахин Л., Рамбо К. (февраль 2017 г.). «Гликозидгидролазы разрушают полимикробные бактериальные биопленки в ранах» . Антимикробные средства и химиотерапия . 61 (2): AAC.01998–16. дои : 10.1128/AAC.01998-16 . ПМЦ 5278739 . ПМИД 27872074 .
- ^ Джасани Х., Умретия Н., Дхараджия Д., Капурия М., Шах С., Патель Дж. (июнь 2016 г.). «Выделение, оптимизация и производство целлюлазы Aspergillus niger из сельскохозяйственных отходов» . Журнал чистой и прикладной микробиологии . 10 (2): 1159–66.
- ^ Умезурике Г.М. (январь 1979 г.). «Целлюлолитические ферменты Botryodiplodia theobromae Патент. Разделение и характеристика целлюлаз и β-глюкозидаз» . Биохимический журнал . 177 (1): 9–19. дои : 10.1042/bj1770009 . ПМЦ 1186335 . ПМИД 106849 .
- ^ Телке А.А., Чжуан Н., Гатге С.С., Ли Ш., Али Шах А., Хан Х. и др. (2013). «Разработка гликозидгидролазы семейства 5 (Cel5A) из некультивируемой бактерии для эффективного гидролиза целлюлозных субстратов» . ПЛОС ОДИН . 8 (6): е65727. Бибкод : 2013PLoSO...865727T . дои : 10.1371/journal.pone.0065727 . ПМЦ 3681849 . ПМИД 23785445 .
- ^ Нельсон Н. (1944). «Фотометрическая адаптация метода Сомоджи для определения глюкозы» . Ж. Биол. Хим . 153 (2): 375–80. дои : 10.1016/S0021-9258(18)71980-7 .
- ^ Смогий М. (март 1952 г.). «Заметки по определению сахара» . Журнал биологической химии . 195 (1): 19–23. дои : 10.1016/S0021-9258(19)50870-5 . ПМИД 14938350 .
- ^ Макклири Б.В. (ноябрь 1980 г.). «Новые хромогенные субстраты для анализа альфа-амилазы и (1) приводят к 4)-β-D-глюканазы». Исследование углеводов . 86 (1): 97–104. дои : 10.1016/s0008-6215(00)84584-x . ПМИД 6159974 .
- ^ Макклири Б.В., Манган Д., Дейли Р., Форт С., Айвори Р., МакКормак Н. (февраль 2014 г.). «Новые субстраты для измерения эндо-1,4-β-глюканазы (эндоцеллюлазы)». Исследование углеводов . 385 : 9–17. дои : 10.1016/j.carres.2013.12.001 . ПМИД 24398300 .
- ^ Манган Д., Макклири Б.В., Лядова А., Айвори Р., МакКормак Н. (август 2014 г.). «Количественный флуорометрический анализ для измерения эндо-1,4-β-глюканазы». Исследование углеводов . 395 : 47–51. дои : 10.1016/j.carres.2014.05.002 . ПМИД 25038461 .
Дальнейшее чтение
[ редактировать ]- Чапин Ф.С., Мэтсон П.А., Муни Х.А. (2002). Принципы экологии наземных экосистем (PDF) . Нью-Йорк: Спрингер. ISBN 978-0-387-95439-4 . Архивировано из оригинала (PDF) 5 марта 2016 г. Проверено 4 июля 2014 г.
- Руководство Merck по диагностике и терапии, глава 24
- Дека Д., Бхаргави П., Шарма А., Гоял Д., Джавед М., Гоял А. (2011). «Повышение активности целлюлазы нового штамма Bacillus subtilis путем оптимизации среды и анализа с использованием различных целлюлозных субстратов» . Ферментные исследования . 2011 : 151656. дои : 10.4061/2011/151656 . ПМК 3102325 . ПМИД 21637325 .
- Зафар М., Ахмед С., Хан М.И., Джамиль А. (май 2014 г.). «Рекомбинантная экспрессия и характеристика новой эндоглюканазы из Bacillus subtilis в Escherichia coli ». Отчеты по молекулярной биологии . 41 (5): 3295–302. дои : 10.1007/s11033-014-3192-8 . ПМИД 24493451 . S2CID 203374 .