Биомолекулярная структура
Эта статья может быть запутанной или неясной для читателей . ( Февраль 2016 г. ) |
 |
 |
Биомолекулярная структура -это сложная сложенная трехмерная форма, которая образуется молекулой белка , . ДНК или РНК , и это важно для его функции Структура этих молекул может рассматриваться в любой из нескольких масштабов длины, начиная от уровня отдельных атомов до взаимосвязи между целыми субъединицами белка . Это полезное различие между масштабами часто выражается как разложение молекулярной структуры на четыре уровня: первичный, вторичный, третичный и четвертый. Формаг для этой многомасштабной организации молекулы возникает на вторичном уровне, где фундаментальными структурными элементами являются различные водородные связи молекулы . Это приводит к нескольким узнаваемым доменам структуры белка и структуры нуклеиновой кислоты , включая такие особенности вторичной структуры, как альфа-спирали и бета-листы для белков, а также петли шпильки , выпуклости и внутренние петли для нуклеиновых кислот. Термины первичной , вторичной , третичной и четвертичной структуры были введены Кадж Ульрик Линдерстростм-Ланг в своих медицинских лекциях Lane 1951 года в Стэнфордском университете .
Первичная структура
[ редактировать ]Основной структурой биополимера является точная спецификация его атомного состава и химических связей, соединяющих эти атомы (включая стереохимию ). Для типичного неразветвленного, не связанного с пересечением биополимера (например, молекулы типичного внутриклеточного белка или ДНК или РНК ), первичная структура эквивалентна определению последовательности его мономерных субъединиц, таких как аминокислоты или нуклеотиды .
Первичная структура белка сообщается, начиная с амино -N-конца до карбоксильного C-конца , в то время как первичная структура молекулы ДНК или РНК известна как последовательность нуклеиновой кислоты, о которой сообщается с 5 'конца до 3' конца . Последовательность нуклеиновой кислоты относится к точной последовательности нуклеотидов, которые составляют целую молекулу. Часто первичная структура кодирует мотивы последовательности , которые имеют функциональное значение. Некоторые примеры таких мотивов: C/D [ 1 ] и H/ACA коробки [ 2 ] из Snornas , сайта связывания LSM , обнаруженного в сплицеосомных РНК, таких как U1 , U2 , U4 , U5 , U6 , U12 и U3 , последовательность Shine-Dalgarno , [ 3 ] Козак Консенсусная последовательность [ 4 ] и терминатор РНК -полимеразы III . [ 5 ]
Вторичная структура
[ редактировать ]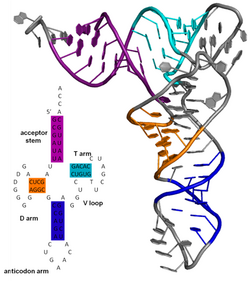
Вторичная структура белка - это паттерн водородных связей в биополимере. Они определяют общую трехмерную форму локальных сегментов биополимеров, но не описывают глобальную структуру специфических атомных положений в трехмерном пространстве, которые считаются третичной структурой . Вторичная структура формально определяется водородными связями биополимера, как наблюдается в структуре атомного разрешения. У белков вторичная структура определяется паттернами водородных связей между аминовыми группами основных целей и карбоксильными группами (боковая - майна и боковая водородные связи с гонщиками и сидехацией), где используется определение DSSP водородной связи.
Вторичная структура нуклеиновой кислоты определяется водородной связью между азотистыми основаниями.
Однако для белков водородная связь коррелирует с другими структурными особенностями, что привело к менее формальным определениям вторичной структуры. Например, спирали могут принять двойные углы магистрали в некоторых регионах заговора Рамачандран ; Таким образом, сегмент остатков с такими двугражными углами часто называют спиралью , независимо от того, имеет ли он правильные водородные связи. Было предложено много других менее формальных определений, часто применяя концепции из дифференциальной геометрии кривых, таких как кривизна и кручение . Структурные биологи, решающие новую структуру атомного разрешения, иногда назначают свою вторичную структуру по глазу и записывают свои назначения в соответствующем файле банка данных белка (PDB).
Вторичная структура молекулы нуклеиновой кислоты относится к взаимодействию спаривания основания в одной молекуле или наборе взаимодействующих молекул. Вторичная структура биологической РНК часто может быть уникально разложена на стебли и петли. Часто эти элементы или их комбинации могут быть дополнительно классифицированы, например, Tetraloops , псевдокноты и петли STEM . Существует много вторичных элементов структуры, имеющих функциональное значение для биологической РНК. Известные примеры включают в себя RH-независимые петли ствола терминатора и клеверлинт -транс-РНК (тРНК). Существует небольшая отрасль исследователей, пытающихся определить вторичную структуру молекул РНК. Подходы включают как экспериментальные , так и вычислительные методы (см. Также список программного обеспечения для прогнозирования структуры РНК ).
Третичная структура
[ редактировать ]Третичная структура белка или любой другой макромолекулы -его трехмерная структура, как определено атомными координатами. [ 6 ] Белки и нуклеиновые кислоты складываются в сложные трехмерные структуры, которые приводят к функциям молекул. Хотя такие структуры разнообразны и сложны, они часто составляют повторяющиеся, узнаваемые третичные структурные мотивы и области, которые служат молекулярными строительными блоками. биомолекулы Считается, что третичная структура в значительной степени определяется первичной структурой (ее последовательность аминокислот или нуклеотидов ).
Четвертая структура
[ редактировать ]Белковая четвертичная структура [ А ] Относится к количеству и расположению нескольких молекул белка в мультисубунском комплексе.
Для нуклеиновых кислот этот термин менее распространен, но может относиться к организации ДНК более высокого уровня в хроматине , [ 7 ] включая его взаимодействие с гистонами или взаимодействие между отдельными РНК -единицами в рибосоме [ 8 ] [ 9 ] или сплайсосома .
Определение структуры
[ редактировать ]Структурное зондирование - это процесс, с помощью которого биохимические методы используются для определения биомолекулярной структуры. [ 10 ] Этот анализ может быть использован для определения закономерностей, которые могут быть использованы для вывода молекулярной структуры, экспериментального анализа молекулярной структуры и функции, а также дальнейшего понимания развития меньших молекул для дальнейших биологических исследований. [ 11 ] Структурно-зондирующий анализ может быть проведен с помощью многих различных методов, которые включают химическое зондирование, зондирование гидроксильного радикала, аналоговое отображение нуклеотидов (NAIM) и встроенное зондирование. [ 10 ]
Структуры белка и нуклеиновой кислоты могут быть определены с использованием либо ядерной магнитно-резонансной спектроскопии ( ЯМР ), либо рентгеновской кристаллографии или одночастичной крио-электронной микроскопии ( криоем ). Первые опубликованные отчеты о ДНК ( Розалинда Франклин и Рэймонда Гослинга А-ДНК в 1953 году) в дифракционных паттернах рентгеновских лучей -и также B-ДНК-используемые анализы, основанные на преобразованиях функций Паттерсона , которые предоставили только ограниченное количество структурной информации для ориентированной волокна ДНК, выделенные из тимуса теленка . [ 12 ] [ 13 ] Альтернативный анализ был затем предложен Wilkins et al. В 1953 году для рентгеновской дифракции и рассеяния B-ДНК-дифракции и рассеяния гидратированных, бактериально-ориентированных ДНК-волокнов и головок сперматозоидов форели с точки зрения квадратов функций Бесселя . [ 14 ] Хотя форма B-ДНК «наиболее распространена в условиях, обнаруженных в клетках, [ 15 ] Это не четко определенная конформация, а семейство или нечеткий набор конформаций ДНК, которые встречаются на высоких уровнях гидратации, присутствующих в широком спектре живых клеток. [ 16 ] Их соответствующие рентгеновские дифракции и рассеяния характерны для молекулярных паракристаллов со значительной степенью расстройства (более 20%), [ 17 ] [ 18 ] и структура не поддается использованию только стандартного анализа.
Напротив, стандартный анализ, включающий только Фурье преобразования Бесселя функций [ 19 ] и ДНК- молекулярные модели все еще обычно используются для анализа рентгеновских дифракционных паттернов A-ДНК и Z-ДНК. [ 20 ]
Прогноз структуры
[ редактировать ]
Прогнозирование биомолекулярной структуры-это прогнозирование трехмерной структуры белка из его аминокислотной последовательности или нуклеиновой кислоты из ее нуклеобазы (основания). Другими словами, это предсказание вторичной и третичной структуры из ее основной структуры. Прогнозирование структуры является обратным биомолекулярного дизайна, как при рациональном дизайне , дизайне белка , дизайне нуклеиновых кислот и биомолекулярной технике .
Прогноз структуры белка является одной из наиболее важных целей, достигнутых биоинформатикой и теоретической химией . Прогнозирование структуры белка имеет большое значение в медицине (например, в дизайне лекарств ) и биотехнологии (например, при разработке новых ферментов ). Каждые два года эффективность современных методов оценивается в критической оценке эксперимента прогнозирования структуры белков ( CASP ).
Также было проведено значительное количество исследований биоинформатики, направленных на проблему прогнозирования структуры РНК. Распространенная проблема для исследователей, работающих с РНК, состоит в том, чтобы определить трехмерную структуру молекулы, которая дает только последовательность нуклеиновой кислоты. Однако в случае РНК большая часть окончательной структуры определяется вторичной структурой или внутримолекулярными взаимодействиями с пассажией основания молекулы. Это показано высокой сохранением парами оснований по разнообразным видам.
Вторичная структура молекул малых нуклеиновых кислот определяется в значительной степени сильными локальными взаимодействиями, такими как водородные связи и базовое укладку . Суммирование свободной энергии для таких взаимодействий, обычно с использованием метода ближайшего соседа , обеспечивает приближение для стабильности заданной структуры. [ 21 ] Наиболее простым способом найти самую низкую структуру свободной энергии было бы генерировать все возможные структуры и рассчитать их свободную энергию, но количество возможных структур для последовательности увеличивается в геометрической прогрессии с длиной молекулы. [ 22 ] Для более длинных молекул количество возможных вторичных структур огромно. [ 21 ]
Методы ковариации последовательности основаны на существовании набора данных, состоящего из нескольких гомологичных последовательностей РНК, с родственными, но разнородными последовательностями. Эти методы анализируют ковариацию отдельных базовых сайтов в эволюции ; с основной пары Поддержание в двух широко разделенных участках пары нуклеотидов указывает на присутствие структурно необходимой водородной связи между этими положениями. Было показано, что общая проблема прогнозирования псевдоки является NP-полным . [ 23 ]
Дизайн
[ редактировать ]Биомолекулярная конструкция может считаться обратным прогнозирования структуры. В прогнозировании структуры структура определяется из известной последовательности, тогда как в конструкции белковой или нуклеиновой кислоты генерируется последовательность, которая будет образовывать желаемую структуру.
Другие биомолекулы
[ редактировать ]Этот раздел нуждается в расширении . Вы можете помочь, добавив к этому . ( Апрель 2010 г. ) |
Другие биомолекулы, такие как полисахариды , полифенолы и липиды , также могут иметь структуру биологических последствий более высокого порядка.
Смотрите также
[ редактировать ]- Биомолекулярный
- Сравнение программного обеспечения для моделирования нуклеиновых кислот
- Структура гена
- Список программного обеспечения для прогнозирования структуры РНК
- Некодирующая РНК
Примечания
[ редактировать ]- ^ Здесь четвертичный означает « структура четвертого уровня », а не « четырехстороннее взаимодействие». Этимологически квартал правильный: четвертичный получен из латинских распределительных чисел и следует бинарным и тройным ; в то время как квартарь получен из латинских порядковых чисел и следует вторичным и третичным . Тем не менее, четверка является стандартной в биологии.
Ссылки
[ редактировать ]- ^ Самарский Д.А., Фурнье М.Дж., Сингер Р.Х., Бертран Е (июль 1998 г.). «Мотив Snorna C/D направляет нуклеолярное нацеливание, а также объединяет синтез и локализацию Snorna» . Embo Journal . 17 (13): 3747–57. doi : 10.1093/emboj/17.13.3747 . PMC 1170710 . PMID 9649444 .
- ^ Ganot P, Caizergues-Ferrer M, Kiss T (апрель 1997 г.). «Семейство коробки ACA небольших нуклеолярных RNA определяется эволюционно консервативной вторичной структурой и вездесущими элементами последовательности, необходимыми для накопления РНК» . Гены и развитие . 11 (7): 941–56. doi : 10.1101/gad.11.7.941 . PMID 9106664 .
- ^ Shine J , Dalgarno L (март 1975 г.). «Определительница специфичности цистрона в бактериальных рибосомах». Природа . 254 (5495): 34–38. Bibcode : 1975nater.254 ... 34S . doi : 10.1038/254034a0 . PMID 803646 . S2CID 4162567 .
- ^ Козак М (октябрь 1987 г.). «Анализ 5'-кодирующих последовательностей из 699 мессенджевых RNA 699 позвоночных» . Исследование нуклеиновых кислот . 15 (20): 8125–48. doi : 10.1093/nar/15.20.8125 . PMC 306349 . PMID 3313277 .
- ^ Bogenhagen DF, Brown DD (апрель 1981 г.). «Нуклеотидные последовательности в ДНК Xenopus 5S, необходимые для завершения транскрипции». Клетка . 24 (1): 261–70. doi : 10.1016/0092-8674 (81) 90522-5 . PMID 6263489 . S2CID 9982829 .
- ^ Iupac , сборник химической терминологии , 2 -е изд. («Золотая книга») (1997). Онлайн -исправленная версия: (2006–) « Третичная структура ». два : 10.1351/goldbook.t06282
- ^ Сипски М.Л., Вагнер Т.Е. (март 1977 г.). «Исследование ДНК четвертичное упорядочение с помощью круговой дихроизма спектроскопии: исследования хромосомных волокон для лошадей». Биополимеры . 16 (3): 573–82. doi : 10.1002/bip.1977.360160308 . PMID 843604 . S2CID 35930758 .
- ^ Noller HF (1984). «Структура рибосомной РНК». Ежегодный обзор биохимии . 53 : 119–62. doi : 10.1146/annurev.bi.53.070184.001003 . PMID 6206780 .
- ^ Nissen P, Ippolito JA, Ban N, Moore PB, Steitz TA (апрель 2001 г.). «Третичные взаимодействия РНК в большой рибосомальной субъединице: A-Minor Motif» . Труды Национальной академии наук Соединенных Штатов Америки . 98 (9): 4899–903. Bibcode : 2001pnas ... 98.4899n . doi : 10.1073/pnas.081082398 . PMC 33135 . PMID 11296253 .
- ^ Jump up to: а беременный Teunissen, A.W.M. (1979). РНК-структурное зондирование: биохимическая структура анализа аутоиммунных молекул РНК, связанных . С. 1–27. ISBN 978-90-901323-4-1 .
- ^ Пейс Н.Р., Томас Б.К., Woese CR (1999). Зондирующая структура, функция и история РНК путем сравнительного анализа . Cold Spring Harbor Laboratory Press. С. 113–17. ISBN 978-0-87969-589-7 .
- ^ Франклин Р.Р. , Гослинг Р.Г. (6 марта 1953 г.). «Структура натрия тимонуклеатских волокон (I. Влияние содержания воды и II. Цилиндрически симметричная функция Паттерсона)» (PDF) . Acta Crystallogr . 6 (8): 673–78. doi : 10.1107/s0365110x53001939 .
- ^ Франклин Р.Е., Гослинг Р.Г. (апрель 1953 г.). «Молекулярная конфигурация в тимонуклеате натрия». Природа . 171 (4356): 740–41. Bibcode : 1953natur.171..740f . doi : 10.1038/171740a0 . PMID 13054694 . S2CID 4268222 .
- ^ Wilkins MH, Stokes AR, Wilson HR (апрель 1953 г.). «Молекулярная структура дезоксипентозных нуклеиновых кислот». Природа . 171 (4356): 738–40. Bibcode : 1953natur.171..738W . doi : 10.1038/171738a0 . PMID 13054693 . S2CID 4280080 .
- ^ Лесли А.Г., Арнотт С., Чандрасекаран Р., Ратлифф Р.Л. (октябрь 1980). «Полиморфизм двойных спиралей ДНК». Журнал молекулярной биологии . 143 (1): 49–72. doi : 10.1016/0022-2836 (80) 90124-2 . PMID 7441761 .
- ^ Baianu, I.C. (1980). «Структурный порядок и частичное расстройство в биологических системах». Бык Математика Биол . 42 (1): 137–41. doi : 10.1007/bf02462372 . S2CID 189888972 .
- ^ Hosemann R, Bagchi RN (1962). Прямой анализ дифракции по материи . Амстердам/Нью-Йорк: Северная Голландия.
- ^ Baianu IC (1978). «Рентгеновское рассеяние от частично неупорядоченных мембранных систем». Acta Crystallogr. А 34 (5): 751–53. Bibcode : 1978accra..34..751b . doi : 10.1107/s0567739478001540 .
- ^ «Функции и дифракции Бесселя с помощью спиральных структур» . Planetphysics.org . [ Постоянная мертвая ссылка ]
- ^ «Рентгеновские дифракционные паттерны двойной спиртной дезоксирибонуклеиновой кислоты (ДНК) кристаллов» . Planetphysics.org . Архивировано из оригинала 24 июля 2009 года.
- ^ Jump up to: а беременный Мэтьюз Д.Х. (июнь 2006 г.). «Революции в предсказании вторичной структуры РНК». Журнал молекулярной биологии . 359 (3): 526–32. doi : 10.1016/j.jmb.2006.01.067 . PMID 16500677 .
- ^ Zuker M, Sankoff D (1984). «РНК вторичные структуры и их прогноз». Бык Математика Биол . 46 (4): 591–621. doi : 10.1007/bf02459506 . S2CID 189885784 .
- ^ Lyngsø RB, Pedersen CN (2000). «Прогноз РНК псевдокнот в энергетических моделях». Журнал вычислительной биологии . 7 (3–4): 409–27. Citeseerx 10.1.1.34.4044 . doi : 10.1089/106652700750050862 . PMID 11108471 .