Метилтрансфераза

Метилтрансферазы представляют собой большую группу ферментов, которые метилируют свои субстраты, но их можно разделить на несколько подклассов в зависимости от их структурных особенностей. Наиболее распространенным классом метилтрансфераз является класс I, все из которых содержат складку Россмана для связывания S -аденозилметионина (SAM). Метилтрансферазы класса II содержат домен SET, примером которого являются метилтрансферазы гистонов с доменом SET , и метилтрансферазы класса III, которые связаны с мембраной. [1] Метилтрансферазы также можно разделить на разные типы, использующие разные субстраты в реакциях переноса метила. Эти типы включают протеин-метилтрансферазы, ДНК/РНК-метилтрансферазы, метилтрансферазы натуральных продуктов и не-SAM-зависимые метилтрансферазы. SAM является классическим донором метила для метилтрансфераз, однако в природе встречаются примеры других доноров метила. Общий механизм переноса метила представляет собой нуклеофильную атаку, подобную S N 2 , где сера метионина служит уходящей группой, а присоединенная к ней метильная группа действует как электрофил , который переносит метильную группу на ферментный субстрат. SAM превращается в S -аденозилгомоцистеин Во время этого процесса (SAH). Разрыв связи SAM-метил и образование связи субстрат-метил происходят почти одновременно. Эти ферментативные реакции встречаются во многих путях и участвуют в генетических заболеваниях, раке и метаболических заболеваниях. Другим типом переноса метила является радикал S-аденозилметионин (SAM), который представляет собой метилирование неактивированных атомов углерода в первичных метаболитах, белках, липидах и РНК.
Функция
[ редактировать ]Генетика
[ редактировать ]Метилирование, как и другие эпигенетические модификации, влияет на транскрипцию , стабильность генов и родительский импринтинг . [2] Он напрямую влияет на структуру хроматина и может модулировать транскрипцию генов или даже полностью заглушать или активировать гены без мутации самого гена. Хотя механизмы этого генетического контроля сложны, гипо- и гиперметилирование ДНК участвует во многих заболеваниях.
Регуляция белка
[ редактировать ]Метилирование белков играет регуляторную роль в белок-белковых взаимодействиях , взаимодействиях белок-ДНК и активации белков.
Примеры: RCC1 , важный митотический белок, метилирован так, что он может взаимодействовать с центромерами хромосом. Это пример регуляции белок-белкового взаимодействия, поскольку метилирование регулирует присоединение RCC1 к белкам-гистонам H2A и H2B . Взаимодействие RCC1-хроматин также является примером взаимодействия белок-ДНК, поскольку другой домен RCC1 напрямую взаимодействует с ДНК, когда этот белок метилируется. Когда RCC1 не метилирован, делящиеся клетки имеют несколько полюсов веретена и обычно не могут выжить.
p53 метилирует лизин, чтобы регулировать его активацию и взаимодействие с другими белками в ответ на повреждение ДНК. Это пример регуляции белок-белковых взаимодействий и активации белков. p53 — известный супрессор опухоли , который активирует восстановления ДНК пути , инициирует апоптоз и приостанавливает клеточный цикл . В целом, он реагирует на мутации в ДНК, сигнализируя клетке исправить их или инициировать клеточную смерть, чтобы эти мутации не могли способствовать развитию рака.
NF-κB (белок, участвующий в воспалении) является известной мишенью метилирования метилтрансферазы SETD6 , которая выключает передачу сигналов NF-κB путем ингибирования одной из ее субъединиц, RelA . Это снижает активацию транскрипции и воспалительную реакцию , делая метилирование NF-κB регуляторным процессом, посредством которого передача сигналов через этот путь клетками снижается. [3]
Природные продукты метилтрансфераз вносят разнообразный вклад в метаболические пути, включая доступность кофакторов, сигнальных молекул и метаболитов. Это регулирует различные клеточные пути, контролируя активность белка.
Типы
[ редактировать ]Гистоновые метилтрансферазы
[ редактировать ]Гистоновые метилтрансферазы имеют решающее значение для генетической регуляции на эпигенетическом уровне. Они модифицируют в основном лизин по ε-азоту и аргинин -гуанидиниевую группу на хвостах гистонов. Лизинметилтрансферазы и аргининметилтрансферазы представляют собой уникальные классы ферментов, но оба связывают SAM в качестве донора метила для своих гистоновых субстратов. Аминокислоты лизина могут быть модифицированы одной, двумя или тремя метильными группами, а аминокислоты аргинина могут быть модифицированы одной или двумя метильными группами. Это увеличивает силу положительного заряда и гидрофобность остатков , позволяя другим белкам распознавать метильные метки. Эффект этой модификации зависит от местоположения модификации на хвосте гистона и других модификаций гистонов вокруг него. Местоположение модификаций может частично определяться последовательностью ДНК, а также малыми некодирующими РНК и метилированием самой ДНК. Чаще всего у позвоночных метилируется гистон H3 или H4. Вокруг модификации может происходить как повышенная, так и пониженная транскрипция генов. Повысился транскрипция является результатом снижения конденсации хроматина , тогда как снижение транскрипции является результатом повышенной конденсации хроматина. [4] Метильные метки на гистонах способствуют этим изменениям, служа местами для рекрутирования других белков, которые могут дополнительно модифицировать хроматин. [5]
N-концевые метилтрансферазы
[ редактировать ]N-альфа-метилтрансферазы переносят метильную группу от SAM к N-концевому азоту белковых мишеней. N-концевой метионин сначала расщепляется другим ферментом, а консенсусная последовательность X- пролин -лизин распознается метилтрансферазой. Для всех известных субстратов аминокислота X представляет собой аланин , серин или пролин. Эта реакция дает метилированный белок и SAH. Известные мишени этих метилтрансфераз у человека включают RCC-1 (регулятор белков ядерного транспорта) и белок ретинобластомы (белок-супрессор опухолей, который ингибирует чрезмерное деление клеток). Метилирование RCC-1 особенно важно при митозе , поскольку оно координирует локализацию некоторых ядерных белков в отсутствие ядерной оболочки . Когда RCC-1 не метилирован, деление клеток происходит аномально после образования дополнительных полюсов веретена . [6] Функция N-концевого метилирования белка ретинобластомы неизвестна.
ДНК/РНК-метилтрансферазы
[ редактировать ]
Метилирование ДНК, ключевой компонент генетической регуляции, происходит преимущественно на 5-углеродном элементе основания цитозина , образуя 5'-метилцитозин (см. слева). [7] Метилирование — это эпигенетическая модификация, катализируемая ферментами ДНК-метилтрансферазой , включая DNMT1, DNMT2 (переименованный в TRDMT1, чтобы отразить его функцию по метилированию тРНК, а не ДНК) и DNMT3. Эти ферменты используют S-аденозилметионин в качестве донора метила и содержат несколько высококонсервативных структурных особенностей между тремя формами; к ним относятся сайт связывания S-аденозилметионина, вицинальная пара пролин-цистеин, которая образует тиолат-анион, важный для механизма реакции, и карман связывания цитозинового субстрата. Многие особенности ДНК-метилтрансфераз высококонсервативны во многих классах жизни, от бактерий до млекопитающих. Помимо контроля экспрессии определенных генов , существует множество белковых комплексов, многие из которых имеют значение для здоровья человека, которые связываются только с сайтами узнавания метилированной ДНК . Считалось, что многие из ранних ДНК-метилтрансфераз произошли от РНК-метилтрансфераз, которые должны были быть активными в мире РНК для защиты многих видов примитивных РНК. [8] Метилирование РНК наблюдалось у разных типов РНК, а именно. мРНК , рРНК , тРНК , мяРНК , мяРНК , миРНК , тмРНК , а также виды вирусных РНК. Специфические РНК-метилтрансферазы используются клетками для маркировки их на видах РНК в соответствии с потребностями и окружающей средой, преобладающей вокруг клеток, что составляет часть области, называемой молекулярной эпигенетикой . 2'-O-метилирование , метилирование m6A , метилирование m1G, а также m5C чаще всего являются метками метилирования, наблюдаемыми в различных типах РНК.
6А представляет собой фермент, который катализирует следующие химические реакции: [9]
S-аденозил-L-метионин + ДНК-аденин S-аденозил-L-гомоцистеин + ДНК 6-метиламинопурин
m6A в основном обнаруживался у прокариот до 2015 года, а затем был идентифицирован у некоторых эукариот. Метилтрансферазы m6A метилируют аминогруппу ДНК в положении C-6 специально, чтобы не дать системе-хозяину переварить собственный геном с помощью ферментов рестрикции. [10]
m5C играет роль в регуляции транскрипции генов. Трансферазы m5C представляют собой ферменты, которые продуцируют C5-метилцитозин в ДНК в положении C-5 цитозина, и обнаружены у большинства растений и некоторых эукариот. [11]
Натуральные продукты метилтрансфераз
[ редактировать ]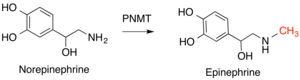
Метилтрансферазы натуральных продуктов (NPMT) представляют собой разнообразную группу ферментов, которые добавляют метильные группы к небольшим молекулам, полученным естественным путем. Как и многие метилтрансферазы, SAM используется в качестве донора метила и образуется SAH. Метильные группы добавляются к атомам S, N, O или C и классифицируются по тому, какой из этих атомов модифицирован, при этом O-метилтрансферазы представляют самый большой класс. Метилированные продукты этих реакций выполняют множество функций, включая кофакторы, пигменты, сигнальные соединения и метаболиты. NPMT могут выполнять регулирующую роль, изменяя реакционную способность и доступность этих соединений. Эти ферменты не являются высококонсервативными у разных видов, поскольку они выполняют более специфическую функцию, обеспечивая малые молекулы для специализированных путей у видов или меньших групп видов. Отражением этого разнообразия является разнообразие каталитических стратегий, включая общий кислотно-основной катализ , катализ на основе металлов близости и , а также эффекты десольватации , не требующие каталитических аминокислот. NPMT представляют собой наиболее функционально разнообразный класс метилтрансфераз. [12]

Важные примеры этого класса ферментов у людей включают фенилэтаноламин-N-метилтрансферазу (PNMT), которая превращает норадреналин в адреналин . [14] и гистамин-N-метилтрансфераза (HNMT), которая метилирует гистамин в процессе метаболизма гистамина. [15] Катехол -О -метилтрансфераза (КОМТ) разрушает класс молекул, известный как катхоламины , который включает дофамин , адреналин и норэпенеферин. [16]
Не-SAM-зависимые метилтрансферазы
[ редактировать ]Метанол , метилтетрагидрофолат , моно- , ди- и триметиламин , метантиол , метилтетрагидрометаноптерин и хлорметан — все это доноры метила, обнаруженные в биологии в качестве доноров метильных групп, обычно в ферментативных реакциях с использованием кофактора витамина B12 . [17] Эти субстраты участвуют в путях переноса метила, включая биосинтез метионина , метаногенез и ацетогенез .
Радикальные SAM-метилтрансферазы
[ редактировать ]В зависимости от различных белковых структур и механизмов катализа существует три различных типа радикальных SAM (RS) метилаз: класса A, B и C. RS-метилазы класса A лучше всего охарактеризованы из четырех ферментов и связаны как с RlmN, так и с RlmN. Сфр. RlmN повсеместно встречается у бактерий, что повышает точность трансляции, а RlmN катализирует метилирование C2 аденозина 2503 (A2503) в 23 S рРНК и C2 аденозина (A37). Cfr, с другой стороны, также катализирует метилирование C8 A2503, а также катализирует метилирование C2. [18] Класс B в настоящее время является крупнейшим классом радикальных SAM-метилаз, которые могут метилировать как sp 2-гибридные, так и sp 3-гибридизированные атомы углерода в различных наборах субстратов, в отличие от класса A, который катализирует только sp 2-гибридизированные атомы углерода. Основным отличием класса B от других является дополнительный N-концевой домен, связывающий кобаламин, который связывается с доменом RS. [19] Метилаза класса C имеет гомологичную последовательность с ферментом RS, копропорфириноген III оксидазой (HemN), который также катализирует метилирование sp 2-гибридизированных углеродных центров, но в ней отсутствуют 2 цистеина, необходимые для метилирования по механизму класса A. [18]
Клиническое значение
[ редактировать ]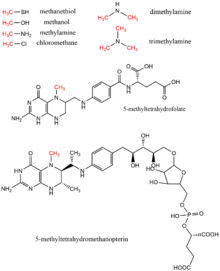
Как и любой биологический процесс, который регулирует экспрессию и/или функцию генов, аномальное метилирование ДНК связано с генетическими нарушениями, такими как ИКФ , синдром Ретта и синдром ломкой Х-хромосомы . [2] Раковые клетки обычно проявляют меньшую активность метилирования ДНК в целом, хотя часто наблюдается гиперметилирование в участках, которые неметилированы в нормальных клетках; это сверхметилирование часто действует как способ инактивации генов-супрессоров опухолей . В качестве варианта лечения было предложено ингибировать общую активность ДНК-метилтрансферазы, но было обнаружено, что ингибиторы DNMT, аналоги их цитозиновых субстратов, высокотоксичны из-за их сходства с цитозином (см. Справа); это сходство с нуклеотидом приводит к включению ингибитора в трансляцию ДНК , вызывая синтез нефункционирующей ДНК.
Метилаза, которая изменяет сайт связывания рибосомальной РНК антибиотика линезолида, вызывает перекрестную устойчивость к другим антибиотикам, действующим на рибосомальную РНК. Плазмидные векторы, способные передавать этот ген, являются причиной потенциально опасной перекрестной резистентности. [20]
Примеры ферментов метилтрансфераз, имеющих отношение к заболеванию:
- тиопуринметилтрансфераза : дефекты этого гена вызывают токсическое накопление тиопуриновых соединений, препаратов, используемых в химиотерапии и иммунодепрессантной терапии.
- метионинсинтаза : злокачественная анемия , вызванная дефицитом витамина B12 , вызвана отсутствием кофактора фермента метионинсинтазы.
Применение в открытии и разработке лекарств
[ редактировать ]Недавняя работа показала, что метилтрансферазы, участвующие в метилировании природных противораковых агентов, используют аналоги S-аденозилметионина (SAM), которые несут альтернативные алкильные группы в качестве замены метила. Разработка простой химиоферментной платформы для создания и использования дифференциально алкилированных аналогов SAM в контексте открытия и разработки лекарств известна как алкилрандомизация . [21]
Применение в лечении рака
[ редактировать ]В клетках человека было обнаружено, что m5C связан с аномальными опухолевыми клетками при раке. [22] Роль и потенциальное применение m5C включают в себя балансировку нарушенной ДНК при раке как гиперметилирования, так и гипометилирования. Эпигенетическое восстановление ДНК может быть применено путем изменения количества m5C в обоих типах раковых клеток (гиперметилирование/гипометилирование), а также окружающей среды рака, чтобы достичь эквивалентной точки для ингибирования опухолевых клеток. [23]
Примеры
[ редактировать ]Примеры включают в себя:
- Катехол-О-метилтрансфераза
- ДНК-метилтрансфераза
- Гистон метилтрансфераза
- 5-метилтетрагидрофолат-гомоцистеин метилтрансфераза
- О-метилтрансфераза
- метионинсинтаза
- корриноид-железо-серный белок
Ссылки
[ редактировать ]- ^ Кац, Дж. Э.; Длакич, М; Кларк С. (18 июля 2003 г.). «Автоматическая идентификация предполагаемых метилтрансфераз из геномных открытых рамок считывания» . Молекулярная и клеточная протеомика . 2 (8): 525–40. дои : 10.1074/mcp.M300037-MCP200 . ПМИД 12872006 .
- ^ Jump up to: а б Седлецкий, П; Зеленкевич, П. (2006). «ДНК-метилтрансферазы млекопитающих» . Акта Биохимика Полоника . 53 (2): 245–56. дои : 10.18388/abp.2006_3337 . ПМИД 16582985 .
- ^ Леви, Дэн; и др. (5 декабря 2010 г.). «Метилирование лизина субъединицы RelA NF-κB с помощью SETD6 связывает активность гистон-метилтрансферазы GLP в хроматине с тонической репрессией передачи сигналов NF-κB» . Природная иммунология . 12 (1): 29–36. дои : 10.1038/н.1968 . ПМК 3074206 . ПМИД 21131967 .
- ^ Тернер, Брайан М. (2001). Хроматин и регуляция генов: механизмы в эпигенетике . Молден, Массачусетс: Blackwell Science. ISBN 978-0865427433 .
- ^ Грир, Эрик Л.; Ши, Ян (3 апреля 2012 г.). «Метилирование гистонов: динамический показатель здоровья, болезней и наследования» . Обзоры природы Генетика . 13 (5): 343–357. дои : 10.1038/nrg3173 . ПМК 4073795 . ПМИД 22473383 .
- ^ Кларк, Пол (май 2007 г.). «Закрепление RCC1 за хвост». Природная клеточная биология . 9 (5): 485–487. дои : 10.1038/ncb0507-485 . ПМИД 17473856 . S2CID 711645 .
- ^ Лан, Дж; Хуа, С; Он, Х; Чжан, Ю (2010). «ДНК-метилтрансферазы и метилсвязывающие белки млекопитающих». Acta Biochimica et Biophysica Sinica . 42 (4): 243–52. дои : 10.1093/abbs/gmq015 . ПМИД 20383462 .
- ^ Рана, Аджай К.; Анкри, Серж (01 января 2016 г.). «Возрождение мира РНК: взгляд на появление РНК-метилтрансфераз» . Границы генетики . 7:99 . дои : 10.3389/fgene.2016.00099 . ПМЦ 4893491 . ПМИД 27375676 .
- ^ Кесслер, Кристоф; Манта, Висентиу (1 января 1990 г.). «Специфичность эндонуклеаз рестрикции и метилтрансфераз модификации ДНК - обзор (издание 3)». Джин . 92 (1): 1–240. дои : 10.1016/0378-1119(90)90486-Б . ISSN 0378-1119 . ПМИД 2172084 .
- ^ Нарва, Кеннет Э.; Ван Эттен, Джеймс Л.; Слатко, Бартон Э.; Беннер, Джек С. (25 декабря 1988 г.). «Аминокислотная последовательность эукариотической ДНК [N6-аденин]метилтрансферазы M · CviBIII имеет области сходства с прокариотическим изошизомером M · TaqI и другими ДНК [N6-аденин] метилтрансферазами». Джин . 74 (1): 253–259. дои : 10.1016/0378-1119(88)90298-3 . ISSN 0378-1119 . ПМИД 3248728 .
- ^ Посфаи, Янош; Бхагват, Ашок С.; Робертс, Ричард Дж. (25 декабря 1988 г.). «Мотивы последовательности, специфичные для цитозинметилтрансфераз». Джин . 74 (1): 261–265. дои : 10.1016/0378-1119(88)90299-5 . ISSN 0378-1119 . ПМИД 3248729 .
- ^ Лискомб, Дэвид К.; Луи, Гордон В.; Ноэль, Джозеф П. (2012). «Архитектура, механизмы и молекулярная эволюция натуральных продуктов метилтрансфераз». Отчеты о натуральных продуктах . 29 (10): 1238–50. дои : 10.1039/c2np20029e . ПМИД 22850796 .
- ^ Ашихара, Хироши; Ёкота, Такао; Крозье, Алан (2013). Биосинтез и катаболизм пуриновых алкалоидов . Достижения в ботанических исследованиях. Том. 68. стр. 111–138. дои : 10.1016/B978-0-12-408061-4.00004-3 . ISBN 9780124080614 .
- ^ «PNMT фенилэтаноламин N-метилтрансфераза» . Реестр генетических тестов NCBI . Проверено 18 февраля 2014 г.
- ^ «HNMT гистамин-N-метилтрансфераза» . Реестр генетических тестов NCBI . Проверено 18 февраля 2014 г.
- ^ «КОМТ-катехин-О-метилтрансфераза» . Реестр генетических тестов NCBI . Проверено 18 февраля 2014 г.
- ^ Рэгсдейл, SW «Катализ переноса метильных групп с участием тетрагидрофолата и B12» Витамины и гормоны , 2008.
- ^ Jump up to: а б Бауэрле, Мэтью Р.; Швальм, Эрика Л.; Букер, Сквайр Дж. (13 февраля 2015 г.). «Механистическое разнообразие радикального S-аденозилметионина (SAM)-зависимого метилирования» . Журнал биологической химии . 290 (7): 3995–4002. дои : 10.1074/jbc.R114.607044 . ISSN 0021-9258 . ПМК 4326810 . ПМИД 25477520 .
- ^ София, ГЮ; Чен, Г.; Хетцлер, Б.Г.; Рейес-Спиндола, Дж. Ф.; Миллер, штат Невада (01 марта 2001 г.). «Радикальный SAM, новое суперсемейство белков, связывающее нерешенные этапы знакомых путей биосинтеза с радикальными механизмами: функциональная характеристика с использованием новых методов анализа и визуализации информации» . Исследования нуклеиновых кислот . 29 (5): 1097–1106. дои : 10.1093/нар/29.5.1097 . ISSN 1362-4962 . ПМК 29726 . ПМИД 11222759 .
- ^ Моралес Дж., Пикасо Х.Дж., Баос Э., Кандел Ф.Дж., Арриби А., Пелаес Б., Андраде Р., де ла Торре М.А., Феререс Х., Санчес-Гарсиа М. (март 2010 г.). «Устойчивость к линезолиду опосредована геном cfr в первом сообщении о вспышке резистентного к линезолиду Staphylococcus aureus» . Клин. Заразить. Этот . 50 (6): 821–5. дои : 10.1086/650574 . ПМИД 20144045 .
- ^ Сингх, С; Чжан, Дж; Хубер, Т.Д.; Сункара, М; Херли, К; Гофф, Р.Д.; Ван, Дж; Чжан, В; Лю, С; Рор, Дж; Ван Ланен, генеральный директор; Моррис, Эй Джей; Торсон, Дж.С. (7 апреля 2014 г.). «Простые химиоферментативные стратегии синтеза и использования аналогов S-аденозил-(L)-метионина» . Angewandte Chemie International Edition на английском языке . 53 (15): 3965–9. дои : 10.1002/anie.201308272 . ПМК 4076696 . ПМИД 24616228 .
- ^ Джонс, Питер А. (1 июня 1996 г.). «Ошибки метилирования ДНК и рак» . Исследования рака . 56 (11): 2463–2467. ISSN 0008-5472 . ПМИД 8653676 .
- ^ Д, Ханахан; Ра, Вайнберг (4 марта 2011 г.). «Признаки рака: следующее поколение» . Клетка . 144 (5): 646–74. дои : 10.1016/j.cell.2011.02.013 . ПМИД 21376230 .
Дальнейшее чтение
[ редактировать ]- Метилтрансферазы Национальной медицинской библиотеки США в медицинских предметных рубриках (MeSH)
- Трехмерная структура ДНК-метилтрансферазы
- Новая метилтрансфераза: фермент, кэпирующий метилфосфат 7SK snRNA, как видно на Flintbox
- «Роль метилирования в экспрессии генов» на сайте Nature Scitable
- «Питание и депрессия: питание, метилирование и депрессия» в журнале Psychology Today
- «Метилирование ДНК. Что такое метилирование ДНК?» с сайта News-Medical.net
- «Метилирование лизина гистонов» Генетические пути с участием метилтрансфераз гистонов из технологии клеточной сигнализации