5-метилцитозин
 | |
| Имена | |
|---|---|
| Предпочтительное название ИЮПАК 4-Амино-5-метилпиримидин-2(1H ) -он | |
| Идентификаторы | |
3D model ( JSmol ) | |
| 3DMeet | |
| 120387 | |
| ЧЭБИ | |
| ХимическийПаук | |
| Информационная карта ECHA | 100.008.236 |
| Номер ЕС |
|
| КЕГГ | |
| МеШ | 5-метилцитозин |
ПабХим CID | |
| НЕКОТОРЫЙ | |
Панель управления CompTox ( EPA ) | |
| Характеристики | |
| C5H7N3C5H7N3O | |
| Молярная масса | 125.131 g·mol −1 |
| Опасности | |
| СГС Маркировка : | |
 | |
| Предупреждение | |
| Х317 , Х319 | |
| P261 , P264 , P272 , P280 , P302+P352 , P305+P351+P338 , P321 , P333+P313 , P337+P313 , P363 , P501 | |
Если не указано иное, данные приведены для материалов в стандартном состоянии (при 25 °C [77 °F], 100 кПа). | |
5-Метилцитозин представляет собой метилированную форму ДНК основания цитозина (C), которая регулирует транскрипцию генов и выполняет ряд других биологических функций. [1] Когда цитозин метилируется, ДНК сохраняет ту же последовательность, но экспрессия метилированных генов может быть изменена (изучение этого является частью области эпигенетики ). 5-Метилцитозин включен в состав нуклеозида 5-метилцитидина .
В 5-метилцитозине метильная группа присоединена к 5-му атому 6-атомного кольца, считая против часовой стрелки от NH-связанного азота в положении «шесть часов». Эта метильная группа отличает 5-метилцитозин от цитозина.
Открытие
[ редактировать ]Пытаясь выделить бактериальный токсин, вызывающий туберкулез новую нуклеиновую кислоту , названную туберкулиновой кислотой , В. Г. Руппель в 1898 году выделил из туберкулезной палочки . [2] Нуклеиновая кислота оказалась необычной тем, что содержала помимо тимина , гуанина и цитозина , метилированный нуклеотид. В 1925 году Джонсон и Когхилл успешно обнаружили незначительное количество метилированного производного цитозина как продукта гидролиза туберкулиновой кислоты серной кислотой . [3] [4] Этот отчет подвергся резкой критике, поскольку их идентификация была основана исключительно на оптических свойствах кристаллического пикрата , а другим ученым не удалось воспроизвести тот же результат. [5] Но его существование было окончательно доказано в 1948 году, когда Хочкисс выделил нуклеиновые кислоты ДНК из теленка тимуса с помощью бумажной хроматографии , с помощью которой он обнаружил уникальный метилированный цитозин, совершенно отличный от обычных цитозина и урацила . [6] Спустя семь десятилетий выяснилось, что это также общая черта для разных молекул РНК , хотя точная роль неясна. [7]
В естественных условиях
[ редактировать ]Функция этого химического вещества значительно различается у разных видов: [8]
- У бактерий 5-метилцитозин можно обнаружить в различных местах, и он часто используется в качестве маркера для защиты ДНК от разрезания нативными чувствительными к метилированию рестриктазами .
- У растений 5-метилцитозин встречается в последовательностях CpG , CpHpG и CpHpH (где H = A, C или T).
- У грибов и животных 5-метилцитозин преимущественно встречается в динуклеотидах CpG . Большинство эукариот метилировано 70–80% цитозинов CpG метилируют лишь небольшой процент этих сайтов, но у позвоночных . В клетках млекопитающих кластеры CpG на 5'-концах генов называются CpG-островками. [9] 1% всей ДНК млекопитающих составляет 5 мК. [10]
В то время как спонтанное дезаминирование цитозина образует урацил , который распознается и удаляется ферментами репарации ДНК, дезаминирование 5-метилцитозина образует тимин . Это преобразование основания ДНК из цитозина (C) в тимин (T) может привести к переходной мутации . [11] Кроме того, активное ферментативное дезаминирование цитозина или 5-метилцитозина с помощью цитозиндезаминаз семейства APOBEC может иметь благотворное влияние на различные клеточные процессы, а также на эволюцию организма. [12] последствия дезаминирования 5-гидроксиметилцитозина С другой стороны, остаются менее изученными.
В пробирке
[ редактировать ]Группа NH 2 может быть удалена (дезаминирование) из 5-метилцитозина с образованием тимина с использованием таких реагентов, как азотистая кислота ; цитозин дезаминирует до урацила (U) в аналогичных условиях. [ нужна ссылка ]
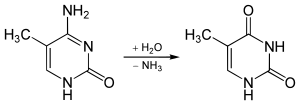
5-метилцитозин устойчив к дезаминированию при обработке бисульфитом , который дезаминирует остатки цитозина. Это свойство часто используется для анализа закономерностей метилирования цитозина ДНК с помощью бисульфитного секвенирования . [13]
Добавление и регулирование с помощью DNMT (эукариоты)
[ редактировать ]Метки 5mC помещаются на геномную ДНК с помощью ДНК-метилтрансфераз (DNMT). У человека имеется 5 DNMT: DNMT1, DNMT2, DNMT3A, DNMT3B и DNMT3L, а в водорослях и грибах присутствуют еще 3 (DNMT4, DNMT5 и DNMT6). [14] DNMT1 содержит последовательность, нацеленную на фокусы репликации (RFTS) и домен CXXC, которые катализируют добавление меток 5mC. RFTS направляет DNMT1 к локусам репликации ДНК, чтобы помочь поддерживать 5mC на дочерних цепях во время репликации ДНК, тогда как CXXC содержит домен цинкового пальца для de novo . добавления метилирования к ДНК [15] Было обнаружено, что DNMT1 является преобладающей ДНК-метилтрансферазой во всех тканях человека. [16] В первую очередь DNMT3A и DNMT3B отвечают за метилирование de novo , а DNMT1 сохраняет отметку 5mC после репликации. [1] DNMT могут взаимодействовать друг с другом, увеличивая способность метилирования. Например, 2 DNMT3L может образовывать комплекс с 2 DNMT3A для улучшения взаимодействия с ДНК, способствуя метилированию. [17] Изменения в экспрессии DNMT приводят к аберрантному метилированию. Сверхэкспрессия приводит к усилению метилирования, тогда как нарушение работы фермента снижает уровень метилирования. [16]

Механизм присоединения следующий: сначала остаток цистеина в мотиве PCQ DNMT создает нуклеофильную атаку по углероду 6 на нуклеотид цитозина, который должен быть метилирован. Затем S-аденозилметионин отдает метильную группу углероду 5. Основание фермента DNMT депротонирует остаточный водород на углероде 5, восстанавливая двойную связь между атомами углерода 5 и 6 в кольце, образуя пару оснований 5-метилцитозин. [15]
Деметилирование
[ редактировать ]После того как цитозин метилируется до 5mC, его можно вернуть в исходное состояние с помощью нескольких механизмов. Пассивное деметилирование ДНК путем разведения метку постепенно устраняет за счет репликации из-за отсутствия поддержки со стороны DNMT. При активном деметилировании ДНК серия окислений превращает ее в 5-гидроксиметилцитозин (5hmC), 5-формилцитозин (5fC) и 5-карбоксилцитозин (5caC), а последние два в конечном итоге вырезаются тимин-ДНК-гликозилазой (TDG), а затем путем эксцизионной репарации оснований (BER) для восстановления цитозина. [1] Нокаут TDG привел к двукратному увеличению 5fC без каких-либо статистически значимых изменений до уровней 5hmC, что указывает на то, что 5mC должен быть итеративно окислен по крайней мере дважды перед его полным деметилированием. [18] Окисление происходит посредством диоксигеназ семейства TET (Ten-eleven translocation) ( ферментов TET ), которые могут превращать 5mC, 5hmC и 5fC в их окисленные формы. Однако наибольшее предпочтение фермент имеет к 5mC, а начальная скорость реакции превращений 5hmC и 5fC с помощью ТЕТ2 в 4,9–7,6 раза медленнее. [19] ТЕТ требует Fe (II) в качестве кофактора, а также кислорода и α-кетоглутарата (α-KG) в качестве субстратов, причем последний субстрат образуется из изоцитрата с помощью фермента изоцитратдегидрогеназы (IDH). [20] Однако рак может производить 2-гидроксиглутарат (2HG), который конкурирует с α-KG, снижая активность ТЕТ и, в свою очередь, уменьшая превращение 5mC в 5hmC. [21]
Роль у человека
[ редактировать ]При раке
[ редактировать ]При раке ДНК может стать как чрезмерно метилированной, называемой гиперметилированием , так и недостаточно метилированной, называемой гипометилированием. [22] Островки CpG, перекрывающие промоторы генов, метилируются de novo, что приводит к аберрантной инактивации генов, обычно связанных с ингибированием роста опухолей (пример гиперметилирования). [23] Сравнивая опухоль и нормальную ткань, в первой наблюдались повышенные уровни метилтрансфераз DNMT1, DNMT3A и, главным образом, DNMT3B, все из которых связаны с аномальными уровнями 5mC при раке. [16] Повторяющиеся последовательности в геноме, включая сателлитную ДНК, Alu и длинные вкрапления элементов (LINE), часто обнаруживаются гипометилированными при раке, что приводит к экспрессии этих обычно молчащих генов, а их уровни часто являются важными маркерами прогрессирования опухоли. [22] Была выдвинута гипотеза, что существует связь между гиперметилированием и гипометилированием; Повышенная активность ДНК-метилтрансфераз, которые вызывают аномальное метилирование 5mC de novo, может быть компенсирована удалением метилирования, типом эпигенетической репарации. Однако удаление метилирования неэффективно, что приводит к чрезмерному гипометилированию всего генома. Возможно и обратное; Чрезмерная экспрессия гипометилирования может быть подавлена за счет полногеномного гиперметилирования. [22] Отличительные способности рака, вероятно, приобретаются посредством эпигенетических изменений, которые изменяют 5mC как в раковых клетках, так и в окружающей опухоль-ассоциированной строме в микроокружении опухоли. [24] противораковый препарат Цисплатин реагирует с 5mC. Сообщалось, что [25]
Как биомаркер старения
[ редактировать ]«Эпигенетический возраст» относится к связи между хронологическим возрастом и уровнем метилирования ДНК в геноме. [26] Сочетание уровней метилирования ДНК в определенных наборах CpG, называемых «часовыми CpG», с алгоритмами, которые регрессируют типичные уровни коллективного общегеномного метилирования в заданном хронологическом возрасте, позволяет прогнозировать эпигенетический возраст. В молодости (0–20 лет) изменения в метилировании ДНК происходят быстрее по мере развития и роста, а в старшем возрасте эти изменения начинают замедляться. Существует множество эпигенетических оценок возраста. Часы Хорвата измеряют набор из 353 CpG, состоящий из нескольких тканей, половина из которых положительно коррелирует с возрастом, а другая половина отрицательно, для оценки эпигенетического возраста. [27] Часы Ханнума используют образцы крови взрослых для расчета возраста на основе ортогональной основы из 71 CpG. [28] Часы Левина, известные как DNAm PhenoAge, зависят от 513 CpG и превосходят другие оценки возраста в прогнозировании смертности и продолжительности жизни, но при этом демонстрируют смещение в отношении тканей, не относящихся к крови. [29] Имеются сообщения об оценщиках возраста со состоянием метилирования только одного CpG в гене ELOVL2. [30] Оценка возраста позволяет прогнозировать продолжительность жизни на основе ожиданий возрастных состояний, которым могут подвергаться люди, на основе их маркеров метилирования 5mC. [ нужна ссылка ]
Ссылки
[ редактировать ]- ^ Jump up to: а б с У С, Чжан Ю (30 мая 2017 г.). «ТЕТ-опосредованное активное деметилирование ДНК: механизм, функции и не только». Обзоры природы Генетика . 18 (9): 517–534. дои : 10.1038/nrg.2017.33 . ISSN 1471-0056 . ПМИД 28555658 . S2CID 3393814 .
- ^ Мэтьюз АП (2012). Физиологическая химия . Компания Уильямс и Уилкинс/. п. 167. ИСБН 978-1130145373 .
- ^ Джонсон Т.Б., Когхилл Р.Д. (1925). «Открытие 5-метилцитозина в туберкулиновой кислоте, нуклеиновой кислоте туберкулезной палочки ». J Am Chem Soc . 47 (11): 2838–2844. дои : 10.1021/ja01688a030 .
- ^ Грожан Х (2009). Нуклеиновые кислоты не скучны. Длинные полимеры, состоящие всего из четырех типов нуклеотидов: экскурсия . Ландес Бионаука.
- ^ Вишер Э., Заменгоф С., Чаргафф Э. (1949). «Микробные нуклеиновые кислоты: дезоксипентозные нуклеиновые кислоты птичьих туберкулезных палочек и дрожжей» . J Биол Хим . 177 (1): 429–438. дои : 10.1016/S0021-9258(18)57100-3 . ПМИД 18107446 .
- ^ Гочкис Р.Д. (1948). «Количественное разделение пуринов, пиримидинов и нуклеозидов методом бумажной хроматографии» . J Биол Хим . 175 (1): 315–332. дои : 10.1016/S0021-9258(18)57261-6 . ПМИД 18873306 .
- ^ Сквайрс Дж.Э., Патель Х.Р., Нуш М., Сиббритт Т., Хамфрис Д.Т., Паркер Б.Дж., Сутер СМ, Прейсс Т. (2012). «Широкое распространение 5-метилцитозина в кодирующей и некодирующей РНК человека» . Нуклеиновые кислоты Рез . 40 (11): 5023–5033. дои : 10.1093/nar/gks144 . ПМК 3367185 . ПМИД 22344696 .
- ^ Колот В., Россиньоль Дж.Л. (1999). «Метилирование ДНК эукариот как эволюционный механизм». Биоэссе . 21 (5): 402–411. doi : 10.1002/(SICI)1521-1878(199905)21:5<402::AID-BIES7>3.0.CO;2-B . ПМИД 10376011 . S2CID 10784130 .
- ^ Берд АП (май 1986 г.). «Островки, богатые CpG, и функция метилирования ДНК». Природа . 321 (6067): 209–213. Бибкод : 1986Natur.321..209B . дои : 10.1038/321209a0 . ISSN 0028-0836 . ПМИД 2423876 . S2CID 4236677 .
- ^ Эрлих М., Ван Р.Ю. (19 июня 1981 г.). «5-Метилцитозин в эукариотической ДНК». Наука . 212 (4501): 1350–1357. Бибкод : 1981Sci...212.1350E . дои : 10.1126/science.6262918 . ISSN 0036-8075 . ПМИД 6262918 .
- ^ Сасса А., Канемару Ю., Камосита Н., Хонма М., Ясуи М. (2016). «Мутагенные последствия изменений цитозина, специфически встроенных в геном человека» . Гены и окружающая среда . 38 (1): 17. Бибкод : 2016GeneE..38...17S . дои : 10.1186/s41021-016-0045-9 . ПМК 5007816 . ПМИД 27588157 .
- ^ Чахван Р., Вонтакал С.Н., Роа С. (2010). «Перекресток между генетической и эпигенетической информацией посредством дезаминирования цитозина». Тенденции в генетике . 26 (10): 443–448. дои : 10.1016/j.tig.2010.07.005 . ПМИД 20800313 .
- ^ Кларк С.Дж., Харрисон Дж., Пол К.Л., Фроммер М. (1994). «Высокочувствительное картирование метилированных цитозинов» . Нуклеиновые кислоты Рез . 22 (15): 2990–2997. дои : 10.1093/нар/22.15.2990 . ПМК 310266 . ПМИД 8065911 .
- ^ Понгер Л., Ли WH (1 апреля 2005 г.). «Эволюционная диверсификация ДНК-метилтрансфераз в геномах эукариот» . Молекулярная биология и эволюция . 22 (4): 1119–1128. дои : 10.1093/molbev/msi098 . ISSN 0737-4038 . ПМИД 15689527 .
- ^ Jump up to: а б Лико Ф (февраль 2018 г.). «Семейство ДНК-метилтрансфераз: универсальный набор инструментов для эпигенетической регуляции». Обзоры природы Генетика . 19 (2): 81–92. дои : 10.1038/nrg.2017.80 . ISSN 1471-0064 . ПМИД 29033456 . S2CID 23370418 .
- ^ Jump up to: а б с Робертсон К.Д., Узволги Э., Лян Г., Талмадж С., Сумеги Дж., Гонсалес Ф.А., Джонс П.А. (01.06.1999). «ДНК-метилтрансферазы человека (DNMT) 1, 3a и 3b: координируют экспрессию мРНК в нормальных тканях и сверхэкспрессию в опухолях» . Исследования нуклеиновых кислот . 27 (11): 2291–2298. дои : 10.1093/нар/27.11.2291 . ISSN 0305-1048 . ПМК 148793 . ПМИД 10325416 .
- ^ Цзя Д., Юрковска Р.З., Чжан X, Елч А., Ченг X (сентябрь 2007 г.). «Структура Dnmt3a, связанного с Dnmt3L, предполагает модель метилирования ДНК de novo» . Природа . 449 (7159): 248–251. Бибкод : 2007Natur.449..248J . дои : 10.1038/nature06146 . ISSN 1476-4687 . ПМК 2712830 . ПМИД 17713477 .
- ^ Сун CX, Шулвак К.Э., Дай Q, Фу Ю, Мао С.К., Линь Л, Стрит С, Ли Ю, Пойдевин М, Ву Х, Гао Дж (25 апреля 2013 г.). «Полногеномное профилирование 5-формилцитозина раскрывает его роль в эпигенетическом прайминге» . Клетка . 153 (3): 678–691. дои : 10.1016/j.cell.2013.04.001 . ISSN 1097-4172 . ПМЦ 3657391 . ПМИД 23602153 .
- ^ Ито С., Шен Л., Дай Ц., Ву С.К., Коллинз Л.Б., Свенберг Дж.А., Хэ С., Чжан Ю. (02.09.2011). «Тет-белки могут превращать 5-метилцитозин в 5-формилцитозин и 5-карбоксилцитозин» . Наука . 333 (6047): 1300–1303. Бибкод : 2011Sci...333.1300I . дои : 10.1126/science.1210597 . ISSN 0036-8075 . ПМЦ 3495246 . ПМИД 21778364 .
- ^ Лу X, Чжао Б.С., Хэ С (12 февраля 2015 г.). «Белки семейства ТЕТ: окислительная активность, взаимодействующие молекулы и функции при заболеваниях» . Химические обзоры . 115 (6): 2225–2239. дои : 10.1021/cr500470n . ISSN 0009-2665 . ПМЦ 4784441 . ПМИД 25675246 .
- ^ Сюй В, Ян Х, Лю И, Ян И, Ван П, Ким Ш, Ито С, Ян С, Ван П, Сяо МТ, Лю Лкс (18 января 2011 г.). «Онкометаболит 2-гидроксиглутарат является конкурентным ингибитором α-кетоглутарат-зависимых диоксигеназ» . Раковая клетка . 19 (1): 17–30. дои : 10.1016/j.ccr.2010.12.014 . ISSN 1535-6108 . ПМЦ 3229304 . ПМИД 21251613 .
- ^ Jump up to: а б с Эрлих М (01 декабря 2009 г.). «Гипометилирование ДНК в раковых клетках» . Эпигеномика . 1 (2): 239–259. дои : 10.2217/эпи.09.33 . ISSN 1750-1911 . ПМК 2873040 . ПМИД 20495664 .
- ^ Джонс, Пенсильвания (1 июня 1996 г.). «Ошибки метилирования ДНК и рак» . Исследования рака . 56 (11): 2463–2467. ISSN 0008-5472 . ПМИД 8653676 .
- ^ Ханахан Д., Вайнберг Р.А. (4 марта 2011 г.). «Признаки рака: следующее поколение» . Клетка . 144 (5): 646–674. дои : 10.1016/j.cell.2011.02.013 . ISSN 0092-8674 . ПМИД 21376230 .
- ^ Менке А., Дубини Р.К., Майер П., Рово П., Дауманн Л. (23 октября 2020 г.). «Образование аддуктов цисплатина с эпигенетически релевантным нуклеиновым основанием 5-метилцитозином» . Европейский журнал неорганической химии . 2021 : 30–36. дои : 10.1002/ejic.202000898 . ISSN 1434-1948 .
- ^ Хорват С., Радж К. (июнь 2018 г.). «Биомаркеры, основанные на метилировании ДНК, и теория старения эпигенетических часов». Обзоры природы Генетика . 19 (6): 371–384. дои : 10.1038/s41576-018-0004-3 . ISSN 1471-0064 . ПМИД 29643443 . S2CID 4709691 .
- ^ Хорват С (10 декабря 2013 г.). «Возраст метилирования ДНК тканей и типов клеток человека» . Геномная биология . 14 (10): 3156. doi : 10.1186/gb-2013-14-10-r115 . ISSN 1474-760X . ПМК 4015143 . ПМИД 24138928 . (Ошибка: два : 10.1186/s13059-015-0649-6 , PMID 25968125 , Часы втягивания )
- ^ Ханнум Дж., Гинни Дж., Чжао Л., Чжан Л., Хьюз Дж., Садда С., Клотцле Б., Бибикова М., Фан Дж.Б., Гао Ю., Деконде Р. (24 января 2013 г.). «Профили метилирования всего генома раскрывают количественные данные о темпах старения человека» . Молекулярная клетка . 49 (2): 359–367. дои : 10.1016/j.molcel.2012.10.016 . ISSN 1097-2765 . ПМК 3780611 . ПМИД 23177740 .
- ^ Левин М.Э., Лу А.Т., Куах А., Чен Б.Х., Ассимес Т.Л., Бандинелли С., Хоу Л., Баккарелли А.А., Стюарт Дж.Д., Ли Ю., Уитсел Е.А. (17 апреля 2018 г.). «Эпигенетический биомаркер старения для продолжительности жизни и здоровья» . Старение (Олбани, штат Нью-Йорк) . 10 (4): 573–591. дои : 10.18632/aging.101414 . ISSN 1945-4589 . ПМК 5940111 . ПМИД 29676998 .
- ^ Гараньани П., Бакалини М.Г., Пираццини С., Гори Д., Джулиани С., Мари Д., Блазио А.М., Джентилини Д., Витале Г., Коллино С., Резци С. (2012). «Метилирование гена ELOVL2 как новый эпигенетический маркер возраста». Стареющая клетка . 11 (6): 1132–1134. дои : 10.1111/acel.12005 . hdl : 11585/128353 . ISSN 1474-9726 . ПМИД 23061750 . S2CID 8775590 .
Литература
[ редактировать ]- Гриффитс, Энтони Дж. Ф. (1999). Введение в генетический анализ . Сан-Франциско: WH Freeman. Глава 15: Генная мутация. ISBN 0-7167-3520-2 . ( доступно онлайн США в Национальном центре биотехнологической информации )