Микробная петля
Эта статья нуждается в дополнительных цитатах для проверки . ( март 2020 г. ) |
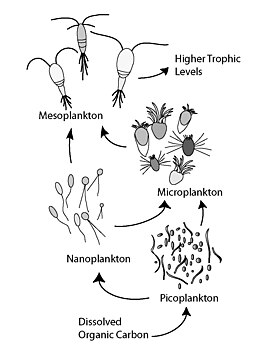
Микробная петля описывает трофический путь, при котором в водных системах растворенный органический углерод (DOC) возвращается на более высокие трофические уровни посредством его включения в бактериальную биомассу, а затем соединяется с классической пищевой цепью, образованной фитопланктоном - зоопланктоном - нектоном . В почвенных системах микробная петля относится к почвенному углероду . Термин «микробная петля» был придуман Фаруком Азамом , Томом Фенчелом и др. [1] в 1983 году, чтобы включить роль бактерий в круговороте углерода и питательных веществ в морской среде.
В общем, растворенный органический углерод (РОУ) попадает в окружающую среду океана в результате бактериального лизиса , утечки или выделения фиксированного углерода из фитопланктона (например, слизистого экзополимера диатомовых водорослей ), внезапного старения клеток , небрежного питания зоопланктона, выделения отходов. продукты водных животных, а также расщепление или растворение органических частиц наземных растений и почв. [2] Бактерии в микробной петле разлагают эти частицы детрита, чтобы использовать это богатое энергией вещество для роста. Поскольку более 95% органического вещества в морских экосистемах состоит из полимерных высокомолекулярных ( ВМВ) соединений (например, белков, полисахаридов, липидов), лишь небольшая часть общего количества растворенного органического вещества (РОВ) легко утилизируется большинством морских организмов. организмы, находящиеся на более высоких трофических уровнях. Это означает, что растворенный органический углерод недоступен непосредственно большинству морских организмов; Морские бактерии вносят этот органический углерод в пищевую сеть, в результате чего дополнительная энергия становится доступной для более высоких трофических уровней. Недавно термин « микробная пищевая сеть » был заменен термином «микробная петля».
История
[ редактировать ]| Часть серии о |
| Углеродный цикл |
|---|
 |
До открытия микробной петли классическое представление о морских пищевых сетях представляло собой линейную цепочку от фитопланктона к нектону . В целом морские бактерии не считались значительными потребителями органического вещества (включая углерод), хотя их существование было известно. Однако взгляд на морскую пелагическую пищевую сеть был оспорен в 1970-х и 1980-х годах Помроем и Азамом, которые предложили альтернативный путь потока углерода от бактерий к простейшим и многоклеточным организмам . [3] [1]
Ранние работы в области морской экологии, изучавшие роль бактерий в океанической среде, пришли к выводу, что их роль очень минимальна. Традиционные методы подсчета бактерий (например, культивирование на чашках с агаром ) дали лишь небольшое количество бактерий, которые были намного меньшими, чем их истинная численность в морской воде. Развитие технологий подсчета бактерий привело к пониманию значительной важности морских бактерий в океанической среде.
В 1970-х годах альтернативный метод прямого микроскопического счета был разработан Франциско и др. (1973) и Хобби и др. (1977). Бактериальные клетки подсчитывали с помощью эпифлуоресцентного микроскопа , производя так называемый « прямой подсчет акридинового оранжевого » (AODC). Это привело к переоценке высокой концентрации бактерий в морской воде, которая оказалась больше, чем ожидалось (обычно порядка 1 миллиона на миллилитр). Кроме того, разработка «анализа бактериальной продуктивности» показала, что большая часть (т.е. 50%) чистой первичной продукции (ЧПП) перерабатывается морскими бактериями.
В 1974 году Ларри Помрой опубликовал в журнале BioScience статью под названием «Пищевая сеть океана: меняющаяся парадигма», в которой была подчеркнута ключевая роль микробов в продуктивности океана. [3] В начале 1980-х годов Азам и группа ведущих ученых-океанистов опубликовали обобщение своего обсуждения в журнале Marine Ecology Progress Series под названием «Экологическая роль микробов водного столба в море». Термин «микробная петля» был введен в этой статье, в которой отмечалось, что потребляющие бактерии протисты принадлежали к тому же размерному классу, что и фитопланктон, и, вероятно, были важным компонентом рациона планктонных ракообразных . [1]
Данные, накопленные с тех пор, показали, что некоторые из этих бактериоядных простейших (например, инфузории ) на самом деле являются выборочной добычей этих копепод . В 1986 году , Роберт Дж. Олсон и другие сотрудники обнаружили прохлорококк , который встречается в большом количестве в олиготрофных районах океана Салли В. Чисхолм (хотя ранее было несколько записей об очень мелких цианобактериях, содержащих хлорофилл b в океан [4] [5] Прохлорококк был открыт в 1986 году. [6] ). [7] Благодаря этому открытию исследователи наблюдали изменение роли морских бактерий в градиенте питательных веществ от эвтрофных к олиготрофным областям океана.
Факторы, контролирующие микробную петлю
[ редактировать ]| Часть серии о |
| Биогеохимические циклы |
|---|
 |
Эффективность микробной петли определяется плотностью морских бактерий внутри нее. [8] Стало ясно, что плотность бактерий в основном контролируется пастбищной деятельностью мелких простейших и различных таксономических групп жгутиконосцев. Кроме того, вирусная инфекция вызывает бактериальный лизис, в результате которого содержимое клеток высвобождается обратно в пул растворенных органических веществ (РОВ), снижая общую эффективность микробного цикла. Смертность от вирусной инфекции имеет почти такую же величину, как и от выпаса простейших. Однако по сравнению с выеданием простейших эффект вирусного лизиса может сильно отличаться, поскольку лизис очень специфичен для каждой морской бактерии. И простейшие, и вирусная инфекция уравновешивают основную часть роста бактерий. Кроме того, микробная петля доминирует в олиготрофных водах, а не в эвтрофных районах - там преобладает классическая пищевая цепь планктона, обусловленная частым притоком свежих минеральных питательных веществ (например, весеннее цветение в водах умеренного пояса, районы апвеллинга ). Величину эффективности микробной петли можно определить путем измерения бактериального включения радиоактивно меченных субстратов (таких как тритированный тимидин или лейцин).
В морских экосистемах
[ редактировать ]Микробная петля имеет особое значение для повышения эффективности морской пищевой сети за счет использования растворенного органического вещества (РОВ), которое обычно недоступно для большинства морских организмов. В этом смысле этот процесс способствует переработке органических веществ и питательных веществ и опосредует передачу энергии выше термоклина . Более 30% растворенного органического углерода (РОУ), содержащегося в бактериях, вдыхается и выделяется в виде углекислого газа . Другим основным эффектом микробной петли в толще воды является то, что она ускоряет минерализацию за счет регенерации продукции в средах с ограниченным количеством питательных веществ (например, в олиготрофных водах). В целом, вся микробная петля в некоторой степени обычно в пять-десять раз превышает массу всех многоклеточных морских организмов в морской экосистеме. Морские бактерии являются основой пищевой сети в большинстве океанических сред, и они улучшают трофическую эффективность как морских пищевых сетей, так и важных водных процессов (таких как продуктивность рыболовства и количество углерода, экспортируемого на дно океана). Таким образом, микробный цикл вместе с первичной продукцией контролирует продуктивность морских систем океана.
Многие планктонные бактерии подвижны, используют жгутик для размножения и хемотакс для обнаружения, перемещения и прикрепления к точечному источнику растворенного органического вещества (РОВ), где быстрорастущие клетки переваривают всю частицу или ее часть. Накопление в течение всего нескольких минут на таких патчах можно наблюдать непосредственно. Следовательно, толщу воды можно в некоторой степени рассматривать как пространственно организованное место небольшого масштаба, а не как полностью смешанную систему. Образование этого пятна влияет на биологически опосредованный перенос вещества и энергии в микробной петле.
В настоящее время микробная петля считается более расширенной. [9] Химические соединения типичных бактерий (такие как ДНК, липиды, сахара и т. д.) и аналогичные значения соотношения C:N на частицу обнаруживаются в микрочастицах, образовавшихся абиотически. Микрочастицы являются потенциально привлекательным источником пищи для бактериоядного планктона. Если это так, микробная петля может быть расширена за счет прямого переноса растворенного органического вещества (РОВ) через образование абиотических микрочастиц на более высокие трофические уровни. Это имеет экологическое значение по двум причинам. Во-первых, это происходит без потери углерода и делает органическое вещество более доступным для фаготрофных организмов, а не только для гетеротрофных бактерий. Более того, абиотическая трансформация в расширенном микробном цикле зависит только от температуры и способности РОВ к агрегации, тогда как биотическая трансформация зависит от его биологической доступности. [9]
В наземных экосистемах
[ редактировать ]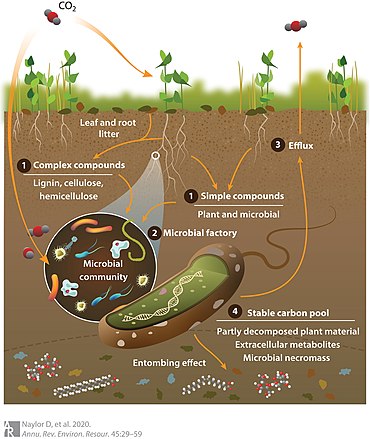
Почвенные экосистемы очень сложны и подвержены различным возмущениям ландшафтного масштаба, которые определяют, удерживается ли почвенный углерод или выбрасывается в атмосферу. [11] Конечная судьба органического углерода почвы является функцией совместной деятельности растений и подземных организмов, включая почвенные микробы. Хотя известно, что почвенные микроорганизмы поддерживают множество биогеохимических функций, связанных с круговоротом углерода, [12] подавляющее большинство почвенного микробиома остается некультивируемым и выполняет в основном загадочные функции. [13] На сегодняшний день каталогизирована лишь незначительная часть почвенной микробной жизни, хотя новые почвенные микробы [13] и вирусы обнаруживаются все чаще. [14] Этот недостаток знаний приводит к неопределенности вклада почвенных микроорганизмов в круговорот органического углерода в почве и препятствует построению точных прогностических моделей глобального потока углерода в условиях изменения климата. [15] [10]
Отсутствие информации о метаболическом потенциале микробиома почвы делает особенно сложным точное объяснение изменений в микробной активности, которые происходят в ответ на изменение окружающей среды. Например, поступления углерода растительного происхождения могут привести к тому, что микробная активность разложит существующий органический углерод в почве со скоростью, превышающей ожидания модели, что приведет к ошибке в моделях прогнозирования потоков углерода. [16] [10]
Чтобы объяснить это, была разработана концептуальная модель, известная как микробный углеродный насос, показанная на диаграмме справа, чтобы определить, как почвенные микроорганизмы трансформируют и стабилизируют органическое вещество почвы. [17] Как показано на схеме, углекислый газ в атмосфере фиксируется растениями (или автотрофными микроорганизмами) и добавляется в почву посредством таких процессов, как (1) корневая экссудация низкомолекулярных простых соединений углерода или отложение листового и корневого опада, приводящее к накоплению сложных растительных полисахаридов. (2) Благодаря этим процессам углерод становится биодоступным для микробной метаболической «фабрики» и впоследствии либо (3) выдыхается в атмосферу, либо (4) попадает в стабильный пул углерода в виде микробной некромассы. Точный баланс оттока углерода и устойчивости является функцией нескольких факторов, включая состав надземного растительного сообщества и профили корневого экссудата, переменные окружающей среды и коллективные микробные фенотипы (т. е. метафеном). [18] [10]
В этой модели метаболическая активность микробов по обмену углерода разделена на две категории: модификация ex vivo, подразумевающая трансформацию углерода растительного происхождения внеклеточными ферментами, и оборот in vivo, когда внутриклеточный углерод используется в обмене микробной биомассы или откладывается в виде мертвых микробов. биомасса, называемая некромассой. Контрастное воздействие катаболической активности, которая высвобождает органический углерод почвы в виде углекислого газа (CO 2 ), и анаболических путей, которые производят стабильные углеродные соединения, контролируют чистую скорость удержания углерода. В частности, микробная секвестрация углерода представляет собой недостаточно представленный аспект потока углерода в почве, который пытается решить модель микробного углеродного насоса. [17] Связанная с этим область неопределенности заключается в том, как тип углерода растительного происхождения увеличивает микробное накопление органического углерода в почве или, альтернативно, ускоряет разложение органического углерода в почве. [19] Например, опавшие листья и опад хвои служат источниками углерода для роста микробов в лесных почвах, но химический состав и pH подстилки варьируются в зависимости от типа растительности (например, между корневой и листовой подстилкой). [20] или между подстилкой лиственных и хвойных лесов (14)]. В свою очередь, эти биохимические различия влияют на уровень органического углерода в почве посредством изменения динамики разложения. [21] Кроме того, увеличение разнообразия растительных сообществ увеличивает скорость ризодепозиции, стимулируя микробную активность и накопление органического углерода в почве. [22] хотя почвы в конечном итоге достигают точки насыщения, после которой они не могут хранить дополнительный углерод. [23] [10]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а б с Азам, Фарук ; Фенчел, Том ; Филд, Дж. Г.; Грей, Дж. С.; Мейер-Рейл, Луизиана; Вингстад, Ф. (1983). «Экологическая роль микробов водной толщи моря» . Серия «Прогресс в области морской экологии» . 10 : 257–263. Бибкод : 1983MEPS...10..257A . дои : 10.3354/meps010257 .
- ^ Ван ден Меерше, Карел; Мидделбург, Джек Дж.; Соэтарт, Карлайн; ван Рейсвейк, Питер; Бошкер, Хенрикус Т.С.; Хейп, Карло Х.Р. (2004). «Сопряжение углерода и азота и взаимодействие водорослей и бактерий во время экспериментального цветения: моделирование эксперимента с индикатором a13C» . Лимнология и океанография . 49 (3): 862–878. Стартовый код : 2004LimOc..49..862V . дои : 10.4319/lo.2004.49.3.0862 . hdl : 1854/LU-434810 . ISSN 0024-3590 .
- ^ Jump up to: а б Помрой, Лоуренс Р. (1974). «Пищевая сеть океана, меняющаяся парадигма». Бионаука . 24 (9): 499–504. дои : 10.2307/1296885 . ISSN 0006-3568 . JSTOR 1296885 .
- ^ Джонсон, PW; Зибурт, Дж. М. (1979). «Хроококкоидные цианобактерии в море: повсеместная и разнообразная фототрофная биомасса». Лимнология и океанография . 24 (5): 928–935. Бибкод : 1979LimOc..24..928J . дои : 10.4319/lo.1979.24.5.0928 .
- ^ Гискес, WWC; Краай, GW (1983). «Неизвестные производные хлорофилла А в Северном море и тропической части Атлантического океана, выявленные с помощью ВЭЖХ-анализа» . Лимнология и океанография . 28 (4): 757–766. Бибкод : 1983LimOc..28..757G . дои : 10.4319/lo.1983.28.4.0757 .
- ^ Чисхолм, Юго-Запад; Олсон, Р.Дж.; Зеттлер, ER; Уотербери, Дж.; Герике, Р.; Вельшмайер, Н. (1988). «Новый свободноживущий прохлорофит встречается при высоких концентрациях клеток в эвфотической зоне океана». Природа . 334 (6180): 340–343. Бибкод : 1988Natur.334..340C . дои : 10.1038/334340a0 . S2CID 4373102 .
- ^ Чисхолм, Салли В .; Франкель, Шейла Л.; Герике, Ральф; Олсон, Роберт Дж.; Паленик, Брайан; Уотербери, Джон Б.; Вест-Джонсруд, Лиза; Зеттлер, Эрик Р. (1992). «Prochromococcus marinus nov. gen. nov. sp.: оксифототрофный морской прокариот, содержащий дивинилхлорофилл a и b». Архив микробиологии . 157 (3): 297–300. дои : 10.1007/bf00245165 . ISSN 0302-8933 . S2CID 32682912 .
- ^ Тейлор, АХ; Джойнт, Я (1990). «Установившийся анализ« микробной петли »в стратифицированных системах» . Серия «Прогресс в области морской экологии» . 59 . Межисследовательский научный центр: 1–17. Бибкод : 1990MEPS...59....1T . дои : 10.3354/meps059001 . ISSN 0171-8630 .
- ^ Jump up to: а б Кернер, Мартин; Хоэнберг, Хайнц; Эртль, Зигмунд; Рекерманн, Маркус; Спитци, Алехандро (2003). «Самоорганизация растворенного органического вещества в мицеллоподобные микрочастицы в речной воде». Природа . 422 (6928). ООО «Спрингер Сайенс энд Бизнес Медиа»: 150–154. Бибкод : 2003Natur.422..150K . дои : 10.1038/nature01469 . ISSN 0028-0836 . ПМИД 12634782 . S2CID 4380194 .
- ^ Jump up to: а б с д и Нейлор, Дэн; Сэдлер, Натали; Бхаттачарджи, Арунима; Грэм, Эмили Б.; Андертон, Кристофер Р.; МакКлюр, Райан; Липтон, Мэри; Хофмокель, Кирстен С.; Янссон, Джанет К. (2020). «Почвенные микробиомы в условиях изменения климата и последствия для круговорота углерода» . Ежегодный обзор окружающей среды и ресурсов . 45 : 29–59. doi : 10.1146/annurev-environ-012320-082720 .
 Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 . - ^ Янссон, Джанет К.; Хофмокель, Кирстен С. (2020). «Почвенные микробиомы и изменение климата» . Обзоры природы Микробиология . 18 (1): 35–46. дои : 10.1038/s41579-019-0265-7 . ОСТИ 1615017 . ПМИД 31586158 . S2CID 203658781 .
- ^ Томпсон, Люк Р.; и др. (2017). «Общий каталог раскрывает многомасштабное микробное разнообразие Земли» . Природа . 551 (7681): 457–463. Бибкод : 2017Natur.551..457T . дои : 10.1038/nature24621 . ПМК 6192678 . ПМИД 29088705 .
- ^ Jump up to: а б Гоэл, Рита; Кумар, Винай; Суял, Дип Чандра; Нараян; Сони, Равиндра (2018). «На пути к некультивируемым микробам для устойчивого сельскохозяйственного производства». Роль ризосферных микробов в почве . стр. 107–123. дои : 10.1007/978-981-10-8402-7_4 . ISBN 978-981-10-8401-0 .
- ^ Шульц, Фредерик; Альтейо, Лорен; Гудо, Даниэль; Райан, Элизабет М.; Ю, Фейцяо Б.; Мальмстрем, Рекс Р.; Бланшар, Джеффри; Войке, Таня (2018). «Скрытое разнообразие вирусов почвенных гигантов» . Природные коммуникации . 9 (1): 4881. Бибкод : 2018NatCo...9.4881S . дои : 10.1038/s41467-018-07335-2 . ПМК 6243002 . ПМИД 30451857 .
- ^ Цзэн, Н.; Ёсикава, К.; Уивер, Эй Джей; Штрассманн, К.; Шнур, Р.; Шницлер, К.-Г.; Рокнер, Э.; Рейк, К.; Рейнер, П.; Раддац, Т.; Мэтьюз, HD; Линдси, К.; Норр, В.; Кавамия, М.; Като, Т.; Йоос, Ф.; Джонс, К.; Джон, Дж.; Бала, Г.; Фунг, И.; Эби, М.; Дони, С.; Кадуле, П.; Бровкин В.; фон Бло, В.; Бопп, Л.; Беттс, Р.; Кокс, П.; Фридлингштейн, П. (2006). «Анализ обратной связи между климатом и углеродным циклом: результаты взаимного сравнения моделей C4MIP» . Журнал климата . 19 (14): 3337–3353. Бибкод : 2006JCli...19.3337F . дои : 10.1175/JCLI3800.1 . hdl : 10036/68733 .
- ^ Кузяков, Яков ; Хорват, Уильям Р.; Дородников Максим; Благодатская, Евгения (2019). «Обзор и синтез воздействия повышенного содержания CO2 в атмосфере на почвенные процессы: никаких изменений в водоемах, но увеличение потоков и ускорение циклов». Биология и биохимия почвы . 128 : 66–78. doi : 10.1016/j.soilbio.2018.10.005 .
- ^ Jump up to: а б Лян, Чао; Шимель, Джошуа П.; Ястроу, Джули Д. (2017). «Важность анаболизма в микробном контроле над хранением углерода в почве». Природная микробиология . 2 (8): 17105. doi : 10.1038/nmicrobiol.2017.105 . ПМИД 28741607 . S2CID 9992380 .
- ^ Бонковски, Майкл (2004). «Простейшие и рост растений: новый взгляд на микробную петлю в почве» . Новый фитолог . 162 (3): 617–631. дои : 10.1111/j.1469-8137.2004.01066.x . ПМИД 33873756 .
- ^ Сулман, Бенджамин Н.; Филлипс, Ричард П.; Оиси, А. Кристофер; Шевлякова, Елена; Пакала, Стивен В. (2014). «Оборот, управляемый микробами, компенсирует накопление углерода в почве за счет минералов при повышенном уровне CO2». Природа Изменение климата . 4 (12): 1099–1102. Бибкод : 2014NatCC...4.1099S . дои : 10.1038/nclimate2436 .
- ^ Хармон, Марк Э.; Сильвер, Уинди Л.; Фаст, Бекки; Чен, Хуа; Берк, Ингрид К.; Партон, Уильям Дж.; Харт, Стивен С.; Карри, Уильям С. (2009). «Долгосрочные закономерности потери массы при разложении листового и мелкого корневого опада: межсайтовое сравнение». Биология глобальных изменений . 15 (5): 1320–1338. Бибкод : 2009GCBio..15.1320H . дои : 10.1111/j.1365-2486.2008.01837.x . hdl : 2027.42/74496 .
- ^ Куоллс, Роберт (2016). «Многолетние (13 лет) скорости разложения органического вещества лесной подстилки на парных хвойных и лиственных водоразделах с контрастными температурными режимами» . Леса . 7 (12): 231. дои : 10.3390/f7100231 .
- ^ Ланге, Маркус; Эйзенхауэр, Нико; Сьерра, Карлос А.; Бесслер, Хольгер; Энгельс, Кристоф; Гриффитс, Роберт И.; Мелладо-Васкес, Перла Г.; Малик, Ашиш А.; Рой, Жак; Шой, Стефан; Штайнбайс, Сибилла; Томсон, Брюс С.; Трамбор, Сьюзен Э .; Гляйкснер, Герд (2015). «Разнообразие растений увеличивает микробную активность почвы и накопление углерода в почве» (PDF) . Природные коммуникации . 6 : 6707. Бибкод : 2015NatCo...6.6707L . дои : 10.1038/ncomms7707 . ПМИД 25848862 .
- ^ Стюарт, Кэтрин Э.; Паустиан, Кейт; Конант, Ричард Т.; Планте, Ален Ф.; Шесть, Йохан (2007). «Насыщение почвы углеродом: концепция, данные и оценка». Биогеохимия . 86 : 19–31. дои : 10.1007/s10533-007-9140-0 . S2CID 97153551 .
Библиография
[ редактировать ]- Фенчел, Т. (1988) Морские планктонные пищевые цепи. Ежегодный обзор экологии и систематики
- Фенчел, Т. (2008) Микробная петля – 25 лет спустя. Журнал экспериментальной морской биологии и экологии
- Фурман Дж. А., Азам Ф. (1982) Включение тимидина как мера образования гетеротрофного бактериопланктона в морских поверхностных водах. Морская биология
- Кернер М., Хоэнберг Х., Эртл С., Рекерманнк М., Спитци А. (2003) Самоорганизация растворенного органического вещества в мицеллоподобные микрочастицы в речной воде. Природа
- Кирчман Д., Сигда Дж., Капусцински Р., Митчелл Р. (1982) Статистический анализ метода прямого подсчета для подсчета бактерий. Прикладная и экологическая микробиология
- Мейнхард С., Азам Ф. (1989) Содержание белка и скорость синтеза белка планктонных морских бактерий. Серия «Прогресс в области морской экологии»
- Мюнстер, В.У. (1985) Исследования структуры, распределения и динамики различных органических субстратов в РОВ озера Плюсзее. Гидробиология
- Помрой, Л.Р., Уильямс, П.Дж.леБ., Азам, Ф. и Хобби, Дж.Э. (2007) «Микробная петля». Океанография , 20 (2): 28–33. дои : 10.4319/lo.2004.49.3.0862 .
- Стодереггер К., Херндл Г.Дж. (1998) Производство и выпуск бактериального капсульного материала и его последующее использование морским бактериопланктоном. Лимнология и океанография

