Ассимиляция серы
Эту статью может потребовать очистки Википедии , чтобы она соответствовала стандартам качества . Конкретная проблема заключается в следующем: эта статья полна нерелевантной или неверной информации, не имеет источников и плохо написана, со множеством опечаток, указывающих не только на отсутствие навыков письма, но, вероятно, и на отсутствие базовых знаний в этой области. ( Март 2024 г. ) |
Эта статья включает список литературы , связанную литературу или внешние ссылки , но ее источники остаются неясными, поскольку в ней отсутствуют встроенные цитаты . ( Март 2024 г. ) |
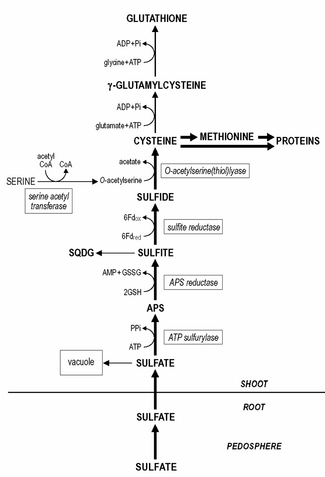
Ассимиляция серы — это процесс, посредством которого живые организмы включают серу в свои биологические молекулы. [1] У растений сульфат поглощается корнями, а затем транспирационным потоком переносится в хлоропласты, где сера восстанавливается до сульфида с помощью ряда ферментативных реакций . Кроме того, восстановленная сера включается в цистеин . [2] аминокислота, которая является предшественником многих других серосодержащих соединений. У животных ассимиляция серы происходит преимущественно с пищей, так как животные не могут производить серосодержащие соединения напрямую. Сера входит в состав аминокислот, таких как цистеин и метионин , которые используются для построения белков и других важных молекул. [2]
Поглощение сульфатов растениями
[ редактировать ]| Часть серии о |
| Биогеохимические циклы |
|---|
 |
Поглощение сульфатов сначала происходит в корнях, которые имеют высокое сродство к сере. [3] Максимальная скорость поглощения сульфата обычно достигается уже при уровне сульфата 0,1 мМ и ниже. Поглощение сульфата корнями и его транспортировка в побеги строго контролируются и, по-видимому, являются одним из основных регуляторных участков ассимиляции серы. [3]
Сульфат активно поглощается через плазматическую мембрану клеток корня и транспортируется , затем загружается в сосуды ксилемы в побег потоком транспирации . [4] Поглощение и транспорт сульфата зависят от энергии (движется градиентом протонов , генерируемым АТФазами ) посредством совместного транспорта протонов и сульфатов . [5] В побеге сульфат выгружается и транспортируется в хлоропласты, где восстанавливается. Оставшийся в растительной ткани сульфат присутствует преимущественно в вакуоли , поскольку концентрация сульфата в цитоплазме сохраняется довольно постоянной.
Различные белки-переносчики сульфата опосредуют поглощение, транспорт и внутриклеточное распределение сульфата. [6] сульфата транспортеров Семейство генов было разделено на 5 различных групп в зависимости от их клеточной и субклеточной экспрессии генов и возможного функционирования. [7] Каждая группа белков-переносчиков может экспрессироваться исключительно в корнях или побегах растения или в обоих.
- Группа 1 представляет собой «переносчики сульфатов с высоким сродством», которые участвуют в поглощении сульфата корнями.
- Группа 2 представляет собой сосудистые переносчики и представляет собой «переносчики сульфатов с низким сродством».
- Группа 3 представляет собой так называемую «листовую группу», однако о характеристиках этой группы пока мало что известно.
- Транспортеры группы 4 участвуют в оттоке сульфата из вакуолей , тогда как функция переносчиков сульфата группы 5 еще не известна и, вероятно, функционируют только как переносчики молибдатов.
Регуляция и экспрессия большинства переносчиков сульфатов контролируются состоянием питания растений серой. [8] При депривации сульфатов быстрое снижение содержания сульфатов в корнях регулярно сопровождается сильно усиленной экспрессией большинства генов-переносчиков сульфатов (до 100 раз), что сопровождается существенно увеличенной способностью поглощения сульфатов. Пока еще не до конца понятно, действуют ли сульфат и другие метаболические продукты ассимиляции серы ( О-ацетилсерин , цистеин , глутатион ) в качестве сигналов в регуляции поглощения и транспорта сульфатов или в экспрессии задействованных переносчиков сульфатов.
Сульфатредукция в растениях
[ редактировать ]Сульфатредукция преимущественно происходит в хлоропластах листьев . Здесь восстановление сульфата до сульфида происходит в три стадии. Сульфат необходимо активировать до аденозин-5'-фосфосульфата (APS) до его восстановления до сульфита .
Активация сульфата катализируется АТФ- сульфурилазой, сродство которой к сульфату довольно низкое (Km около 1 мМ), и концентрация сульфата in situ в хлоропластах, скорее всего, является одним из лимитирующих/регуляторных этапов восстановления серы. Впоследствии АПС восстанавливается до сульфита, катализируемого АПС-редуктазой, с вероятным глутатионом в качестве восстановителя .
Предполагается, что последняя реакция является одной из основных точек регуляции сульфатредукции, поскольку активность APS-редуктазы является самой низкой из ферментов пути сульфатредукции и имеет высокую скорость оборота. Сульфит с высоким сродством восстанавливается сульфитредуктазой до сульфида с помощью ферредоксина в качестве восстановителя. Оставшийся в растительной ткани сульфат переносится в вакуоль . Ремобилизация и перераспределение запасов вакуольных сульфатов происходят довольно медленно, и растения с дефицитом серы все еще могут содержать обнаруживаемые уровни сульфатов.
Синтез и функция соединений серы в растениях.
[ редактировать ]Цистеин
[ редактировать ]Сульфид включается в цистеин , катализируемый О-ацетилсерин(тиол)лиазой, с О-ацетилсерином в качестве субстрата. Синтез О-ацетилсерина катализируется серин -ацетилтрансферазой и вместе с О-ацетилсерин(тиол)лиазой он образует ферментный комплекс, называемый цистеинсинтазой .
Образование цистеина является прямым этапом взаимодействия серы ( метаболизм серы ) и ассимиляции азота в растениях. Это отличается от процесса у дрожжей, где сульфид должен сначала включиться в гомоцистеин, а затем в два этапа превратиться в цистеин.
Цистеин является донором серы для синтеза метионина , основной другой серосодержащей аминокислоты, присутствующей в растениях. Это происходит посредством пути транссульфурации и метилирования гомоцистеина .
И цистеин, и метионин являются серосодержащими аминокислотами и имеют большое значение в структуре, конформации и функциях белков и ферментов , но высокие уровни этих аминокислот также могут присутствовать в запасных белках семян. Тиоловые группы остатков цистеина в белках могут окисляться, что приводит к образованию дисульфидных мостиков с другими боковыми цепями цистеина (и образованию цистина ) и/или связыванию полипептидов .
Дисульфидные мостики ( дисульфидные связи ) вносят важный вклад в структуру белков. Тиоловые ферредоксинах группы также играют большую роль в связывании субстратов ферментов, в металлосерных кластерах в белках (например, ) и в регуляторных белках (например, тиоредоксинах ).
Глутатион
[ редактировать ]Глутатион или его гомологи, например гомоглутатион у Fabaceae ; гидроксиметилглутатион в Poaceae является основным водорастворимым небелковым тиоловым соединением, присутствующим в тканях растений, и составляет 1-2% от общего количества серы. Содержание глутатиона в растительных тканях колеблется в пределах 0,1 – 3 мМ. Цистеин является прямым предшественником синтеза глутатиона (и его гомологов). Сначала γ-глутамилцистеин синтезируется из цистеина и глутамата, катализируемый гамма-глутамилцистеинсинтетазой . Во-вторых, глутатион синтезируется из γ-глутамилцистеина и глицина (в гомологах глутатиона — β-аланина или серина ), катализируемом глутатионсинтетазой. Обе стадии синтеза глутатиона являются АТФ-зависимыми реакциями. Глутатион поддерживается в восстановленной форме с помощью НАДФН -зависимой глутатионредуктазы , а соотношение восстановленного глутатиона (GSH) к окисленному глутатиону (GSSG) обычно превышает значение 7.Глутатион выполняет различные роли в функционировании растений. В метаболизме серы он действует как восстановитель при восстановлении АПС до сульфита. Это также основная форма транспорта восстановленной серы в растениях. Уменьшение поступления серы в корни, вероятно, во многом зависит от переноса глутатиона через побеги/корни. флоэма , поскольку восстановление серы происходит преимущественно в хлоропластах. Глутатион принимает непосредственное участие в восстановлении и ассимиляции селенита в селеноцистеин . Кроме того, глутатион имеет большое значение для защиты растений от окислительного и экологического стресса, а также подавляет/удаляет образование токсичных активных форм кислорода , например, супероксида , перекиси водорода и гидроперекисей липидов . Глутатион действует как восстановитель при ферментативной детоксикации активных форм кислорода в глутатион- аскорбатном цикле и как тиоловый буфер при защите белков посредством прямой реакции с активными формами кислорода или путем образования смешанных дисульфидов. Потенциал глутатиона в качестве защитного средства связан с размером пула глутатиона, его окислительно-восстановительным состоянием (соотношением GSH/GSSG) и активностью глутатионредуктазы . Глутатион является предшественником синтеза фитохелатинов, которые синтезируются ферментативно с помощью конститутивной фитохелатинсинтазы. Число остатков γ-глутамилцистеина в фитохелатинах может колебаться от 2 до 5, иногда до 11. Несмотря на то, что фитохелатины образуют комплексы с некоторыми тяжелыми металлами, а именно. кадмий , предполагается, что эти соединения играют роль в гомеостазе тяжелых металлов и детоксикации путем буферизации цитоплазматической концентрации незаменимых тяжелых металлов. Глутатион также участвует в детоксикации ксенобиотиков , соединений, не имеющих прямой пищевой ценности или значения в метаболизме, слишком высокие уровни которых могут отрицательно повлиять на функционирование растений. Ксенобиотики могут подвергаться детоксикации в реакциях конъюгации с глутатионом, катализируемых глутатион-S-трансферазой , активность которой является конститутивной; разные ксенобиотики могут индуцировать разные изоформы фермента. Глутатион S-трансферазы имеют большое значение для детоксикации гербицидов гербицидов и повышения толерантности в сельском хозяйстве, а их индукция антидотами (« защитниками ») является решающим шагом для индукции толерантности к гербицидам у многих сельскохозяйственных культур. Предполагается, что в естественных условиях глутатион-S-трансферазы играют важную роль в детоксикации липидов. гидроперекиси , в конъюгации эндогенных метаболитов, гормонов и продуктов деградации ДНК , а также в транспорте флавоноидов .
Сульфолипиды
[ редактировать ]Сульфолипиды – серосодержащие липиды. Сульфохиновозилдиацилглицерины являются преобладающими сульфолипидами, присутствующими в растениях. В листьях ее содержание составляет до 3 – 6% от общего количества присутствующей серы. Этот сульфолипид присутствует в пластид мембранах и, вероятно, участвует в функционировании хлоропластов . Путь биосинтеза и физиологическая функция сульфохиновозилдиацилглицерина все еще изучаются. Из недавних исследований очевидно, что сульфит является вероятным предшественником серы для образования сульфохиновозной группы этого липида.
Вторичные соединения серы
[ редактировать ]Brassica Виды содержат глюкозинолаты , которые представляют собой серосодержащие вторичные соединения . Глюкозинолаты состоят из фрагмента β-тиоглюкозы, сульфированного оксима и боковой цепи. Синтез глюкозинолатов начинается с окисления исходной аминокислоты до альдоксима с последующим добавлением тиоловой группы (путем конъюгации с глутатионом) с образованием тиогидроксимата . Перенос глюкозо -сульфатной части завершает образование глюкозинолатов.
Физиологическое значение глюкозинолатов до сих пор неясно, хотя считается, что они действуют как поглотительные соединения в ситуациях избытка серы. При разрушении тканей глюкозинолаты ферментативно разлагаются мирозиназой и могут давать различные биологически активные продукты, такие как изотиоцианаты , тиоцианаты , нитрилы и оксазолидин-2-тионы. Предполагается, что глюкозинолат-мирозиназная система играет роль во взаимодействиях растение- травоядное животное и растение- патоген .
Кроме того, глюкозинолаты отвечают за вкусовые свойства Brassicaceae , а в последнее время привлекли внимание ввиду их потенциальных антиканцерогенных свойств . Виды лука содержат γ- глутамилпептиды и аллиины (S-алк(ен)илцистеинсульфоксиды). Содержание этих серосодержащих вторичных соединений сильно зависит от стадии развития растения, температуры, водообеспеченности и уровня азотного и серного питания. В луковицах их содержание может составлять до 80% органической фракции серы. Меньше известно о содержании вторичных соединений серы на стадии проростков растения.
Предполагается, что аллиины преимущественно синтезируются в листьях, откуда в дальнейшем переносятся в прикрепленные луковичные чешуи. Пути биосинтеза синтеза γ-глутамилпептидов и аллиинов до сих пор остаются неоднозначными. γ-глутамилпептиды могут образовываться из цистеина (через γ-глутамилцистеин или глутатион) и метаболизироваться в соответствующие аллиины посредством окисления и последующего гидролиза γ- глутамилтранспептидазами .
Однако нельзя исключать и другие возможные пути синтеза γ-глутамилпептидов и аллиинов. Известно, что аллиины и γ-глутамилпептиды обладают терапевтической ценностью и могут иметь потенциальную ценность в качестве фитофармацевтических средств. Аллиины и продукты их распада (например, аллицин ) являются предшественниками аромата и вкуса видов. Аромат высвобождается только тогда, когда растительные клетки разрушаются, и фермент аллииназа из вакуоли способен расщеплять аллиины, образуя широкий спектр летучих и нелетучих серосодержащих соединений. Физиологическая функция γ-глутамилпептидов и аллиинов достаточно неясна.
Усвоение серы у животных
[ редактировать ]В отличие от растений, у животных нет пути прямой ассимиляции неорганического сульфата в органические соединения. У животных основным источником серы является пищевой метионин , незаменимая аминокислота, содержащая атом серы. Метионин сначала преобразуется в S-аденозилметионин (SAM), соединение, которое участвует во многих важных биологических процессах, включая метилирование ДНК и синтез нейромедиаторов .
Затем SAM можно использовать для синтеза других важных серосодержащих соединений, таких как цистеин , таурин и глутатион . Цистеин является предшественником синтеза нескольких важных белков и пептидов, а также глутатиона, мощного антиоксиданта, защищающего клетки от окислительного стресса. Таурин участвует во множестве физиологических процессов, включая осморегуляцию , модуляцию передачи сигналов кальция и регуляцию функции митохондрий.
Ассимиация серы микроорганизмами
[ редактировать ]У бактерий и грибов путь ассимиляции серы аналогичен пути ассимиляции серы у растений, где неорганический сульфат восстанавливается до сульфида, а затем включается в цистеин и другие серосодержащие соединения.
Бактерии и грибы могут поглощать неорганический сульфат из окружающей среды через сульфатпереносчик, который регулируется наличием сульфата в среде. Попав внутрь клетки, сульфат активируется АТФ-сульфуралазой с образованием аденозин-5'-фосфосульфата (APS), который затем восстанавливается до сульфита с помощью APS-редуктазы. Сульфит далее восстанавливается до сульфида с помощью сульфитредуктазы, которая затем включается в цистеин с помощью фермента.
Цистеин после синтеза можно использовать для биосинтеза метионина и других важных биомолекул. Кроме того, микроорганизмы используют серосодержащие соединения и для различных других целей, например для синтеза антибиотиков .
Усвоение серы микроорганизмами регулируется множеством факторов внешней среды, в том числе наличием серы в среде и наличием других питательных веществ. Активность ключевых ферментов пути ассимиляции серы также регулируется путем ингибирования по принципу обратной связи со стороны последующих продуктов, аналогично регуляции, наблюдаемой у растений.
Обмен серы в растениях и загрязнение воздуха
[ редактировать ]Быстрый экономический рост, индустриализация и урбанизация связаны с сильным увеличением спроса на энергию и выбросами загрязнителей воздуха, включая диоксид серы (см. также кислотные дожди ) и сероводород растений , которые могут повлиять на метаболизм . Сернистые газы потенциально фитотоксичны , однако они также могут метаболизироваться и использоваться в качестве источника серы и даже быть полезными, если удобрение корней серой недостаточно.
Побеги растений образуют поглотитель атмосферных сернистых газов, которые могут напрямую поглощаться листвой (сухое осаждение). Поглощение диоксида серы листвой обычно находится в прямой зависимости от степени раскрытия устьиц , так как внутреннее сопротивление этому газу невелико. Сульфит хорошо растворим в апопластной воде мезофилла диссоциирует , где с образованием бисульфита и сульфита .
Сульфит может напрямую вступать в путь восстановления серы и восстанавливаться до сульфида , включающегося в цистеин, а затем в другие соединения серы. Сульфит также может быть окислен до сульфата вне- и внутриклеточно пероксидазами или неферментативно катализируемым ионами металлов или супероксидными радикалами , а затем снова восстановлен и ассимилирован. Избыточный сульфат переносится в вакуоль; Повышенный уровень сульфатов в листьях характерен для растений, подвергшихся воздействию.Поглощение сероводорода листвой, по-видимому, напрямую зависит от скорости его метаболизма в цистеин, а затем и в другие соединения серы. Имеются убедительные доказательства того, что О-ацетилсерин(тиол)лиаза непосредственно ответственна за активную фиксацию сероводорода атмосферы растениями.
Растения способны переносить серу из сульфата в поглощенную листьями атмосферную серу в качестве источника серы, и уровни 60 частей на миллиард или выше кажутся достаточными для покрытия потребности растений в сере. Существует взаимодействие между использованием серы в атмосфере и педосфере. Например, воздействие сероводорода может привести к снижению активности APS-редуктазы и подавлению поглощения сульфата.
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Карвансара, Париса Рахимзаде; Коменда, Йозеф; Коприва, Станислав (2024), «Метаболизм серы у цианобактерий» , Cyanobacteria , Elsevier, стр. 117–157, doi : 10.1016/b978-0-443-13231-5.00018-0 , ISBN 978-0-443-13231-5 , получено 15 апреля 2024 г.
- ^ Перейти обратно: а б Валлон, Оливье; Сполдинг, Мартин Х. (2009), «Метаболизм аминокислот» , Справочник по Chlamydomonas , Elsevier, стр. 115–158, doi : 10.1016/b978-0-12-370873-1.00012-5 , ISBN 978-0-12-370873-1 , получено 15 апреля 2024 г.
- ^ Перейти обратно: а б Хан, Мухаммад Сайяр; Хаас, Флориан Генрих; Аллбойе Самами, Арман; Могаддас Голами, Амин; Бауэр, Андреа; Фелленберг, Курт; Райхельт, Майкл; Хэнш, Роберт; Мендель, Ральф Р.; Мейер, Андреас Дж.; Вирц, Маркус; Черт, Рюдигер (4 июня 2010 г.). «Сульфитредуктаза определяет недавно обнаруженное узкое место для ассимиляционного восстановления сульфатов и необходима для роста и развития Arabidopsis thaliana» . Растительная клетка . 22 (4): 1216–1231. дои : 10.1105/tpc.110.074088 . ISSN 1532-298X . ПМЦ 2879758 . ПМИД 20424176 .
- ^ Гиголашвили Тамара; Коприва, Станислав (9 сентября 2014 г.). «Транспортеры в метаболизме серы растений» . Границы в науке о растениях . 5 : 442. doi : 10.3389/fpls.2014.00442 . ISSN 1664-462X . ПМК 4158793 . ПМИД 25250037 .
- ^ Хоксфорд, Малкольм Дж.; Давидиан, Жан-Клод; Гриньон, Клод (1 июня 1993 г.). «Сульфатно-протонный котранспорт в везикулах плазматической мембраны, выделенных из корней Brassica napus L.: повышенный транспорт в мембранах, изолированных от растений, лишенных серы» . Планта . 190 (3): 297–304. дои : 10.1007/BF00196957 . ISSN 1432-2048 .
- ^ Давидиан, Жан-Клод; Коприва, Станислав (март 2010 г.). «Регуляция поглощения и ассимиляции сульфатов - то же самое или не то же самое?» . Молекулярный завод . 3 (2): 314–325. дои : 10.1093/mp/ssq001 . ISSN 1674-2052 . ПМИД 20139159 .
- ^ Пуресмаэли, Фатима; Хейдари, Парвиз; Лоусон, Шанека (27 января 2023 г.). «Изучение семейства генов-переносчиков сульфатов и закономерностей его экспрессии в проростках твердой пшеницы в условиях засоления» . Гены . 14 (2): 333. doi : 10.3390/genes14020333 . ISSN 2073-4425 . ПМК 9956213 . ПМИД 36833260 .
- ^ Бюхнер, Питер; Стуивер, К. Элизабет Э.; Вестерман, Сью; Вирц, Маркус; Черт возьми, Рюдигер; Хоксфорд, Малкольм Дж.; Де Кок, Луит Дж. (1 октября 2004 г.). «Регуляция поглощения сульфатов и экспрессия генов-переносчиков сульфатов у Brassica oleracea под влиянием атмосферного H2S и педосферного сульфатного питания». Физиология растений . 136 (2): 3396–3408. дои : 10.1104/стр.104.046441. ISSN 1532-2548. PMC 523398. PMID 15377780.
- Шнуг, Э. (1998) Сера в агроэкосистемах. Kluwer Academic Publishers, Дордрехт, 221 стр., ISBN 0-7923-5123-1 .
- Гриль Д., Тауш М. и Де Кок Л.Дж. (2001) Значение глутатиона для адаптации растений к окружающей среде. Kluwer Academic Publishers, Дордрехт, ISBN 1-4020-0178-9 .
- Аброл Ю.П. и Ахмад А. (2003) Сера в растениях. Kluwer Academic Publishers, Дордрехт, ISBN 1-4020-1247-0 .
- Сайто К., Де Кок Л.Дж., Стулен И., Хоксфорд М.Дж., Шнуг Э., Сирко А. и Ренненберг Х. (2005) Транспорт и ассимиляция серы в растениях в постгеномную эпоху. Издательство Backhuys, Лейден, ISBN 90-5782-166-4 .
- Хоксфорд, М.Дж. и Де Кок, Л.Дж. (2006) Управление метаболизмом серы в растениях. Растительная клетка и окружающая среда 29: 382-395.