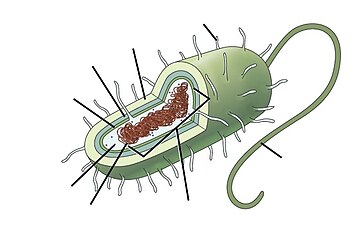
Морские прокариоты
| Часть серии обзоров по |
| Marine life |
|---|
 |
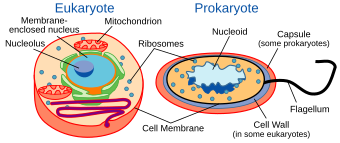
Морские прокариоты — это морские бактерии и морские археи . По среде обитания они определяются как прокариоты , обитающие в морской среде , то есть в соленой воде морей или океанов или в солоноватой воде прибрежных эстуариев . Все клеточные формы жизни можно разделить на прокариотов и эукариотов. Эукариоты — это организмы , клетки которых имеют ядро, заключенное в мембрану , тогда как прокариоты — это организмы, у которых нет ядра, заключенного в мембрану. [1] [2] [3] Трехдоменная система классификации жизни добавляет еще одно деление: прокариоты делятся на два домена жизни: микроскопические бактерии и микроскопические археи, тогда как все остальные, эукариоты, становятся третьим доменом. [4]
Прокариоты играют важную роль в экосистемах как разлагатели, перерабатывающие питательные вещества. Некоторые прокариоты являются патогенными , вызывая болезни и даже смерть растений и животных. [5] Marine prokaryotes are responsible for significant levels of the photosynthesis that occurs in the ocean, as well as significant cycling of carbon and other nutrients.[6]
Prokaryotes live throughout the biosphere. In 2018 it was estimated the total biomass of all prokaryotes on the planet was equivalent to 77 billion tonnes of carbon (77 Gt C). This is made up of 7 Gt C for archaea and 70 Gt C for bacteria. These figures can be contrasted with the estimate for the total biomass for animals on the planet, which is about 2 Gt C, and the total biomass of humans, which is 0.06 Gt C.[7] This means archaea collectively have over 100 times the collective biomass of humans, and bacteria over 1000 times.
There is no clear evidence of life on Earth during the first 600 million years of its existence. When life did arrive, it was dominated for 3,200 million years by the marine prokaryotes. More complex life, in the form of crown eukaryotes, did not appear until the Cambrian explosion a mere 500 million years ago.[8]
Evolution
[edit]−4500 — – −4000 — – −3500 — – −3000 — – −2500 — – −2000 — – −1500 — – −1000 — – −500 — – 0 — | marine prokaryotes |
| ||||||||||||||||||||||||
Marine prokaryotes have been the dominant life form for most of Earth's history, perhaps because water protected them from ionizing radiation[8] | ||||||||||||||||||||||||||
The Earth is about 4.54 billion years old.[9][10][11] The earliest undisputed evidence of life on Earth dates from at least 3.5 billion years ago,[12][13] during the Eoarchean Era after a geological crust started to solidify following the earlier molten Hadean Eon. Microbial mat fossils have been found in 3.48 billion-year-old sandstone in Western Australia.[14][15]

Past species have also left records of their evolutionary history. Fossils, along with the comparative anatomy of present-day organisms, constitute the morphological, or anatomical, record.[16] By comparing the anatomies of both modern and extinct species, paleontologists can infer the lineages of those species. However, this approach is most successful for organisms that had hard body parts, such as shells, bones or teeth. Further, as prokaryotes such as bacteria and archaea share a limited set of common morphologies, their fossils do not provide information on their ancestry.
Prokaryotes inhabited the Earth from approximately 3–4 billion years ago.[17][18] No obvious changes in morphology or cellular organisation occurred in these organisms over the next few billion years.[19] The eukaryotic cells emerged between 1.6 and 2.7 billion years ago. The next major change in cell structure came when bacteria were engulfed by eukaryotic cells, in a cooperative association called endosymbiosis.[20][21] The engulfed bacteria and the host cell then underwent coevolution, with the bacteria evolving into either mitochondria or hydrogenosomes.[22] Another engulfment of cyanobacterial-like organisms led to the formation of chloroplasts in algae and plants.[23]
The history of life was that of the unicellular prokaryotes and eukaryotes until about 610 million years ago when multicellular organisms began to appear in the oceans in the Ediacaran period.[17][26] The evolution of multicellularity occurred in multiple independent events, in organisms as diverse as sponges, brown algae, cyanobacteria, slime moulds and myxobacteria.[27] In 2016 scientists reported that, about 800 million years ago, a minor genetic change in a single molecule called GK-PID may have allowed organisms to go from a single cell organism to one of many cells.[28]
Soon after the emergence of these first multicellular organisms, a remarkable amount of biological diversity appeared over a span of about 10 million years, in an event called the Cambrian explosion. Here, the majority of types of modern animals appeared in the fossil record, as well as unique lineages that subsequently became extinct.[29] Various triggers for the Cambrian explosion have been proposed, including the accumulation of oxygen in the atmosphere from photosynthesis.[30]
Background
[edit]
The words prokaryote and eukaryote come from the Greek where pro means "before", eu means "well" or "true", and karyon means "nut", "kernel" or "nucleus".[31][32][33] So etymologically, prokaryote means "before nucleus" and eukaryote means "true nucleus".
The division of life forms between prokaryotes and eukaryotes was firmly established by the microbiologists Roger Stanier and C. B. van Niel in their 1962 paper, The concept of a bacterium.[34] One reason for this classification was so what was then often called blue-green algae (now called cyanobacteria) would cease to be classified as plants but grouped with bacteria.
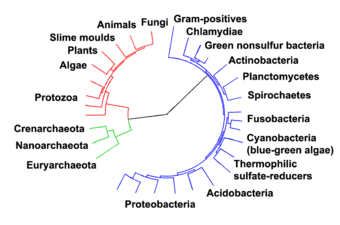
In 1990 Carl Woese et al. introduced the three-domain system.[35][36] The prokaryotes were split into two domains, the archaea and the bacteria, while the eukaryotes become a domain in their own right. The key difference from earlier classifications is the splitting of archaea from bacteria.


| External videos | |
|---|---|

The earliest evidence for life on earth comes from biogenic carbon signatures and stromatolite fossils discovered in 3.7 billion-year-old rocks.[37][38] In 2015, possible "remains of biotic life" were found in 4.1 billion-year-old rocks.[39][40] In 2017 putative evidence of possibly the oldest forms of life on Earth was reported in the form of fossilized microorganisms discovered in hydrothermal vent precipitates that may have lived as early as 4.28 billion years ago, not long after the oceans formed 4.4 billion years ago, and not long after the formation of the Earth 4.54 billion years ago.[41][42]
Microbial mats of coexisting bacteria and archaea were the dominant form of life in the early Archean Eon and many of the major steps in early evolution are thought to have taken place in this environment.[43] The evolution of photosynthesis around 3.5 Ga resulted in a buildup of its waste product oxygen in the atmosphere, leading to the great oxygenation event beginning around 2.4 Ga.[44]
The earliest evidence of eukaryotes dates from 1.85 Ga,[45][46] and while they may have been present earlier, their diversification accelerated when they started using oxygen in their metabolism. Later, around 1.7 Ga, multicellular organisms began to appear, with differentiated cells performing specialised functions.[47]

A stream of airborne microorganisms, including prokaryotes, circles the planet above weather systems but below commercial air lanes.[51] Some peripatetic microorganisms are swept up from terrestrial dust storms, but most originate from marine microorganisms in sea spray. In 2018, scientists reported that hundreds of millions of viruses and tens of millions of bacteria are deposited daily on every square meter around the planet.[52][53]
Microscopic life undersea is diverse and still poorly understood, such as for the role of viruses in marine ecosystems.[54] Most marine viruses are bacteriophages, which are harmless to plants and animals, but are essential to the regulation of saltwater and freshwater ecosystems.[55] They infect and destroy bacteria and archaea in aquatic microbial communities, and are the most important mechanism of recycling carbon in the marine environment. The organic molecules released from the dead bacterial cells stimulate fresh bacterial and algal growth.[56] Viral activity may also contribute to the biological pump, the process whereby carbon is sequestered in the deep ocean.[57]
Marine bacteria
[edit]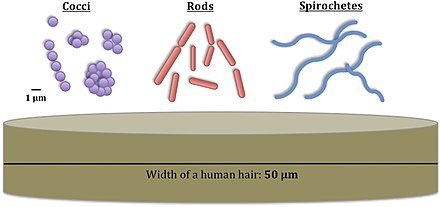



Bacteria constitute a large domain of prokaryotic microorganisms. Typically a few micrometres in length, bacteria have a number of shapes, ranging from spheres to rods and spirals. Bacteria were among the first life forms to appear on Earth, and are present in most of its habitats. Bacteria inhabit soil, water, acidic hot springs, radioactive waste,[61] and the deep portions of Earth's crust. Bacteria also live in symbiotic and parasitic relationships with plants and animals.
Once regarded as plants constituting the class Schizomycetes, bacteria are now classified as prokaryotes. Unlike cells of animals and other eukaryotes, bacterial cells do not contain a nucleus and rarely harbour membrane-bound organelles. Although the term bacteria traditionally included all prokaryotes, the scientific classification changed after the discovery in the 1990s that prokaryotes consist of two very different groups of organisms that evolved from an ancient common ancestor. These evolutionary domains are called Bacteria and Archaea.[62]
The ancestors of modern bacteria were unicellular microorganisms that were the first forms of life to appear on Earth, about 4 billion years ago. For about 3 billion years, most organisms were microscopic, and bacteria and archaea were the dominant forms of life.[63][64] Although bacterial fossils exist, such as stromatolites, their lack of distinctive morphology prevents them from being used to examine the history of bacterial evolution, or to date the time of origin of a particular bacterial species. However, gene sequences can be used to reconstruct the bacterial phylogeny, and these studies indicate that bacteria diverged first from the archaeal/eukaryotic lineage.[65] Bacteria were also involved in the second great evolutionary divergence, that of the archaea and eukaryotes. Here, eukaryotes resulted from the entering of ancient bacteria into endosymbiotic associations with the ancestors of eukaryotic cells, which were themselves possibly related to the Archaea.[21][66] This involved the engulfment by proto-eukaryotic cells of alphaproteobacterial symbionts to form either mitochondria or hydrogenosomes, which are still found in all known Eukarya. Later on, some eukaryotes that already contained mitochondria also engulfed cyanobacterial-like organisms. This led to the formation of chloroplasts in algae and plants. There are also some algae that originated from even later endosymbiotic events. Here, eukaryotes engulfed a eukaryotic algae that developed into a "second-generation" plastid.[67][68] This is known as secondary endosymbiosis.
Bacteria grow to a fixed size and then reproduce through binary fission, a form of asexual reproduction.[69] Under optimal conditions, bacteria can grow and divide extremely rapidly, and bacterial populations can double as quickly as every 9.8 minutes.[70]
Pelagibacter ubique and its relatives may be the most abundant microorganisms in the ocean, and it has been claimed that they are possibly the most abundant bacteria in the world. They make up about 25% of all microbial plankton cells, and in the summer they may account for approximately half the cells present in temperate ocean surface water. The total abundance of P. ubique and relatives is estimated to be about 2 × 1028 microbes.[71] However, it was reported in Nature in February 2013 that the bacteriophage HTVC010P, which attacks P. ubique, has been discovered and is probably the most common organism on the planet.[72][73]
Roseobacter is also one of the most abundant and versatile microorganisms in the ocean. They are diversified across different types of marine habitats, from coastal to open oceans and from sea ice to sea floor, and make up about 25% of coastal marine bacteria. Members of the Roseobacter genus play important roles in marine biogeochemical cycles and climate change, processing a significant portion of the total carbon in the marine environment. They form symbiotic relationships which allow them to degrade aromatic compounds and uptake trace metals. They are widely used in aquaculture and quorum sensing. During algal blooms, 20–30% of the prokaryotic community are Roseobacter.[74][75]
The largest known bacterium, the marine Thiomargarita namibiensis, can be visible to the naked eye and sometimes attains 0.75 mm (750 μm).[76][77]
Cyanobacteria
[edit]


Cyanobacteria were the first organisms to evolve an ability to turn sunlight into chemical energy. They form a phylum (division) of bacteria which range from unicellular to filamentous and include colonial species. They are found almost everywhere on earth: in damp soil, in both freshwater and marine environments, and even on Antarctic rocks.[79] In particular, some species occur as drifting cells floating in the ocean, and as such were amongst the first of the phytoplankton.
The first primary producers that used photosynthesis were oceanic cyanobacteria about 2.3 billion years ago.[80][81] The release of molecular oxygen by cyanobacteria as a by-product of photosynthesis induced global changes in the Earth's environment. Because oxygen was toxic to most life on Earth at the time, this led to the near-extinction of oxygen-intolerant organisms, a dramatic change which redirected the evolution of the major animal and plant species.[82]

interact with bacteria to acquire iron from dust
b. Trichodesmium can establish massive blooms in nutrient poor ocean regions with high dust deposition, partly due to their unique ability to capture dust, center it, and subsequently dissolve it.
c. Proposed dust-bound Fe acquisition pathway: Bacteria residing within the colonies produce siderophores (c-I) that react with the dust particles in the colony core and generate dissolved Fe (c-II). This dissolved Fe, complexed by siderophores, is then acquired by both Trichodesmium and its resident bacteria (c-III), resulting in a mutual benefit to both partners of the consortium.[83]
 Bloom of the filamentous cyanobacteria Trichodesmium
Bloom of the filamentous cyanobacteria Trichodesmium


 Synechococcus, a widespread marine cyanobacterium
Synechococcus, a widespread marine cyanobacterium Carboxysomes appearing as polyhedral dark structures within a species of Synechococcus
Carboxysomes appearing as polyhedral dark structures within a species of Synechococcus



The tiny (0.6 μm) marine cyanobacterium Prochlorococcus, discovered in 1986, forms today an important part of the base of the ocean food chain and accounts for much of the photosynthesis of the open ocean[85] and an estimated 20% of the oxygen in the Earth's atmosphere.[86] It is possibly the most plentiful genus on Earth: a single millilitre of surface seawater may contain 100,000 cells or more.[87]
Originally, biologists classified cyanobacteria as an algae, and referred to it as "blue-green algae". The more recent view is that cyanobacteria are bacteria, and hence are not even in the same Kingdom as algae. Most authorities exclude all prokaryotes, and hence cyanobacteria from the definition of algae.[88][89]
| External videos | |
|---|---|
Other bacteria
[edit]Other marine bacteria, apart from cyanobacteria, are ubiquitous or can play important roles in the ocean. These include the opportunistic copiotroph, Alteromonas macleodii.[90][91]
Marine archaea
[edit]
The archaea (Greek for ancient[93]) constitute a domain and kingdom of single-celled microorganisms. These microbes are prokaryotes, meaning they have no cell nucleus or any other membrane-bound organelles in their cells.
Archaea were initially classified as bacteria, but this classification is outdated.[94] Archaeal cells have unique properties separating them from the other two domains of life, Bacteria and Eukaryota. The Archaea are further divided into multiple recognized phyla. Classification is difficult because the majority have not been isolated in the laboratory and have only been detected by analysis of their nucleic acids in samples from their environment.
Bacteria and archaea are generally similar in size and shape, although a few archaea have very strange shapes, such as the flat and square-shaped cells of Haloquadratum walsbyi.[95] Despite this morphological similarity to bacteria, archaea possess genes and several metabolic pathways that are more closely related to those of eukaryotes, notably the enzymes involved in transcription and translation. Other aspects of archaeal biochemistry are unique, such as their reliance on ether lipids in their cell membranes, such as archaeols. Archaea use more energy sources than eukaryotes: these range from organic compounds, such as sugars, to ammonia, metal ions or even hydrogen gas. Salt-tolerant archaea (the Haloarchaea) use sunlight as an energy source, and other species of archaea fix carbon; however, unlike plants and cyanobacteria, no known species of archaea does both. Archaea reproduce asexually by binary fission, fragmentation, or budding; unlike bacteria and eukaryotes, no known species forms spores.
Archaea are particularly numerous in the oceans, and the archaea in plankton may be one of the most abundant groups of organisms on the planet. Archaea are a major part of Earth's life and may play roles in both the carbon cycle and the nitrogen cycle. Thermoproteota (also called Crenarchaeota or eocytes) are a phylum of archaea thought to be very abundant in marine environments and one of the main contributors to the fixation of carbon.[96]
 Eocytes may be the most abundant of marine archaea
Eocytes may be the most abundant of marine archaea Halobacteria, found in water near saturated with salt, are now recognised as archaea.
Halobacteria, found in water near saturated with salt, are now recognised as archaea. Flat, square-shaped cells of the archaea Haloquadratum walsbyi
Flat, square-shaped cells of the archaea Haloquadratum walsbyi Methanosarcina barkeri, a marine archaea that produces methane
Methanosarcina barkeri, a marine archaea that produces methane Thermophiles, such as Pyrolobus fumarii, survive well over 100 °C (212 °F).
Thermophiles, such as Pyrolobus fumarii, survive well over 100 °C (212 °F).






Nanoarchaeum equitans is a species of marine archaea discovered in 2002 in a hydrothermal vent. It is a thermophile that grows in temperatures at about 80 °C (176 °F). Nanoarchaeum appears to be an obligate symbiont on the archaeon Ignicoccus. It must stay in contact with the host organism to survive since Nanoarchaeum equitans cannot synthesize lipids but obtains them from its host. Its cells are only 400 nm in diameter, making it one of the smallest known cellular organisms, and the smallest known archaeon.[97][98]
Marine archaea have been classified as follows:[99][100][101][102][103]
- Marine Group I (MG-I or MGI): marine Nitrososphaerota with subgroups Ia (aka I.a) up to Id
- Marine Group II (MG-II): marine Euryarchaeota, order Poseidoniales[104] with subgroups IIa up to IId (IIa resembling Poseidoniaceae, IIb resembling Thalassarchaceae)
Viruses parasiting MGII are classified as magroviruses - Marine Group III (MG-III): also marine Euryarchaeota, Marine Benthic Group D[105]
- Marine Group IV (MG-IV): also marine Euryarchaeota[106]
Trophic mode
[edit]Prokaryote metabolism is classified into nutritional groups on the basis of three major criteria: the source of energy, the electron donors used, and the source of carbon used for growth.[107][108]
| Nutritional type | Source of energy | Source of carbon | Examples |
|---|---|---|---|
| Phototrophs | Sunlight | Organic compounds (photoheterotrophs) or carbon fixation (photoautotrophs) | "Cyanobacteria", Green sulfur bacteria, Chloroflexota, or Purple bacteria |
| Lithotrophs | Inorganic compounds | Organic compounds (lithoheterotrophs) or carbon fixation (lithoautotrophs) | Thermodesulfobacteriota, Hydrogenophilaceae, or Nitrospirota |
| Organotrophs | Organic compounds | Organic compounds (chemoheterotrophs) or carbon fixation (chemoautotrophs) | Bacillus, Clostridium or Enterobacteriaceae |
Marine prokaryotes have diversified greatly throughout their long existence. The metabolism of prokaryotes is far more varied than that of eukaryotes, leading to many highly distinct prokaryotic types. For example, in addition to using photosynthesis or organic compounds for energy, as eukaryotes do, marine prokaryotes may obtain energy from inorganic compounds such as hydrogen sulfide. This enables marine prokaryotes to thrive as extremophiles in harsh environments as cold as the ice surface of Antarctica, studied in cryobiology, as hot as undersea hydrothermal vents, or in high saline conditions as (halophiles).[109] Some marine prokaryotes live symbiotically in or on the bodies of other marine organisms.

- Phototrophy is a particularly significant marker that should always play a primary role in bacterial classification.[110]
- Aerobic anoxygenic phototrophic bacteria (AAPBs) are widely distributed marine plankton that may constitute over 10% of the open ocean microbial community. Marine AAPBs are classified in two marine (Erythrobacter and Roseobacter) genera. They can be particularly abundant in oligotrophic conditions where they were found to be 24% of the community.[111] These are heterotrophic organisms that use light to produce energy, but are unable to utilise carbon dioxide as their primary carbon source. Most are obligately aerobic, meaning they require oxygen to grow. Current data suggests that marine bacteria have generation times of several days, whereas new evidence exists that shows AAPB to have a much shorter generation time.[112] Coastal/shelf waters often have greater amounts of AAPBs, some as high as 13.51% AAPB%. Phytoplankton also affect AAPB%, but little research has been performed in this area.[113] They can also be abundant in various oligotrophic conditions, including the most oligotrophic regime of the world ocean.[114] They are globally distributed in the euphotic zone and represent a hitherto unrecognized component of the marine microbial community that appears to be critical to the cycling of both organic and inorganic carbon in the ocean.[115]
- Purple bacteria:
- Zetaproteobacteria: are iron-oxidizing neutrophilic chemolithoautotrophs, distributed worldwide in estuaries and marine habitats.
- Hydrogen oxidizing bacteria are facultative autotrophs that can be divided into aerobes and anaerobes. The former use hydrogen as an electron donor and oxygen as an acceptor while the latter use sulphate or nitrogen dioxide as electron acceptors.[116]
Motility
[edit]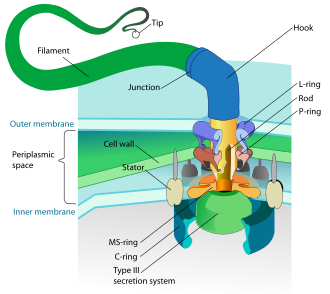
Motility is the ability of an organism to move independently, using metabolic energy.
Flagellar motility
[edit]
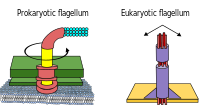
Prokaryotes, both bacteria and archaea, primarily use flagella for locomotion.
- Bacterial flagella are helical filaments, each with a rotary motor at its base which can turn clockwise or counterclockwise.[118][119][120] They provide two of several kinds of bacterial motility.[121][122]
- Archaeal flagella are called archaella, and function in much the same way as bacterial flagella. Structurally the archaellum is superficially similar to a bacterial flagellum, but it differs in many details and is considered non-homologous.[123][117]
The rotary motor model used by bacteria uses the protons of an electrochemical gradient in order to move their flagella. Torque in the flagella of bacteria is created by particles that conduct protons around the base of the flagellum. The direction of rotation of the flagella in bacteria comes from the occupancy of the proton channels along the perimeter of the flagellar motor.[124]
Some eukaryotic cells also use flagella—and they can be found in some protists and plants as well as animal cells. Eukaryotic flagella are complex cellular projections that lash back and forth, rather than in a circular motion. Prokaryotic flagella use a rotary motor, and the eukaryotic flagella use a complex sliding filament system. Eukaryotic flagella are ATP-driven, while prokaryotic flagella can be ATP-driven (archaea) or proton-driven (bacteria).[125]
| External videos | |
|---|---|
Twitching motility
[edit]Twitching motility is a form of crawling bacterial motility used to move over surfaces. Twitching is mediated by the activity of hair-like filaments called type IV pili which extend from the cell's exterior, bind to surrounding solid substrates and retract, pulling the cell forwards in a manner similar to the action of a grappling hook.[126][127][128] The name twitching motility is derived from the characteristic jerky and irregular motions of individual cells when viewed under the microscope.[129]
Gliding motility
[edit]Gliding motility is a type of translocation that is independent of propulsive structures such as flagella or pili.[130] Gliding allows microorganisms to travel along the surface of low aqueous films. The mechanisms of this motility are only partially known. The speed of gliding varies between organisms, and the reversal of direction is seemingly regulated by some sort of internal clock.[131] For example, the apicomplexans are able to travel at fast rates between 1–10 μm/s. In contrast Myxococcus xanthus bacteria glide at a rate of 5 μm/min.[132][133]
Swarming motility
[edit]Swarming motility is a rapid (2–10 μm/s) and coordinated translocation of a bacterial population across solid or semi-solid surfaces,[134] and is an example of bacterial multicellularity and swarm behaviour. Swarming motility was first reported in 1972 by Jorgen Henrichsen.[135]
Non-motile
[edit]Non-motile species lack the ability and structures that would allow them to propel themselves, under their own power, through their environment. When non-motile bacteria are cultured in a stab tube, they only grow along the stab line. If the bacteria are mobile, the line will appear diffuse and extend into the medium.[136]
Taxis: Directed motion
[edit]Magnetotaxis
[edit]

Magnetotactic bacteria orient themselves along the magnetic field lines of Earth's magnetic field.[138] This alignment is believed to aid these organisms in reaching regions of optimal oxygen concentration.[139] To perform this task, these bacteria have biomineralised organelles called magnetosomes that contain magnetic crystals. The biological phenomenon of microorganisms tending to move in response to the environment's magnetic characteristics is known as magnetotaxis. However, this term is misleading in that every other application of the term taxis involves a stimulus-response mechanism. In contrast to the magnetoreception of animals, the bacteria contain fixed magnets that force the bacteria into alignment—even dead cells are dragged into alignment, just like a compass needle.[139]
Marine environments are generally characterized by low concentrations of nutrients kept in steady or intermittent motion by currents and turbulence. Marine bacteria have developed strategies, such as swimming and using directional sensing–response systems, to migrate towards favorable places in the nutrient gradients. Magnetotactic bacteria utilize Earth's magnetic field to facilitate downward swimming into the oxic–anoxic interface, which is the most favorable place for their persistence and proliferation, in chemically stratified sediments or water columns.[140]
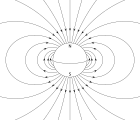
Depending on their latitude and whether the bacteria are north or south of the equator, the Earth's magnetic field has one of the two possible polarities, and a direction that points with varying angles into the ocean depths, and away from the generally more oxygen rich surface. Aerotaxis is the response by which bacteria migrate to an optimal oxygen concentration in an oxygen gradient. Various experiments have clearly shown that magnetotaxis and aerotaxis work in conjunction in magnetotactic bacteria. It has been shown that, in water droplets, one-way swimming magnetotactic bacteria can reverse their swimming direction and swim backwards under reducing conditions (less than optimal oxygen concentration), as opposed to oxic conditions (greater than optimal oxygen concentration).
Regardless of their morphology, all magnetotactic bacteria studied so far are motile by means of flagella.[141] Marine magnetotactic bacteria in particular tend to possess an elaborate flagellar apparatus which can involve up to tens of thousands of flagella. However, despite extensive research in recent years, it has yet to be established whether magnetotactic bacteria steer their flagellar motors in response to their alignment in magnetic fields.[140] Symbiosis with magnetotactic bacteria has been proposed as the explanation for magnetoreception in some marine protists.[142] Research is underway on whether a similar relationship may underlie magnetoreception in vertebrates as well.[143] The oldest unambiguous magnetofossils come from the Cretaceous chalk beds of southern England,[144] though less certain reports of magnetofossils extend to 1.9 billion years old Gunflint Chert.[145]
Gas vacuoles
[edit]| Part of a series on |
| Plankton |
|---|
 |
Some marine prokaryotes possess gas vacuoles. Gas vacuole are nanocompartments freely permeable to gas which allow marine bacteria and archaea to control their buoyancy. They take the form of spindle-shaped membrane-bound vesicles, and are found in some plankton prokaryotes, including some Cyanobacteria.[146] Positive buoyancy is needed to keep the cells in the upper reaches of the water column, so that they can continue to perform photosynthesis. Gas vacuoles are made up of a shell of protein that has a highly hydrophobic inner surface, making it impermeable to water (and stopping water vapour from condensing inside) but permeable to most gases. Because the gas vesicle is a hollow cylinder, it is liable to collapse when the surrounding pressure increases. Natural selection has fine tuned the structure of the gas vesicle to maximise its resistance to buckling, including an external strengthening protein, GvpC, rather like the green thread in a braided hosepipe. There is a simple relationship between the diameter of the gas vesicle and pressure at which it will collapse—the wider the gas vesicle the weaker it becomes. However, wider gas vesicles are more efficient, providing more buoyancy per unit of protein than narrow gas vesicles. Different species produce gas vesicle of different diameter, allowing them to colonise different depths of the water column (fast growing, highly competitive species with wide gas vesicles in the top most layers; slow growing, dark-adapted, species with strong narrow gas vesicles in the deeper layers).
The cell achieves its height in the water column by synthesising gas vesicles. As the cell rises up, it is able to increase its carbohydrate load through increased photosynthesis. Too high and the cell will suffer photobleaching and possible death, however, the carbohydrate produced during photosynthesis increases the cell's density, causing it to sink. The daily cycle of carbohydrate build-up from photosynthesis and carbohydrate catabolism during dark hours is enough to fine-tune the cell's position in the water column, bring it up toward the surface when its carbohydrate levels are low and it needs to photosynthesis, and allowing it to sink away from the harmful UV radiation when the cell's carbohydrate levels have been replenished. An extreme excess of carbohydrate causes a significant change in the internal pressure of the cell, which causes the gas vesicles to buckle and collapse and the cell to sink out.
Large vacuoles are found in three genera of filamentous sulfur bacteria, the Thioploca, Beggiatoa and Thiomargarita. The cytosol is extremely reduced in these genera and the vacuole can occupy between 40 and 98% of the cell.[147] The vacuole contains high concentrations of nitrate ions and is therefore thought to be a storage organelle.[148]
Биолюминесценция
[ редактировать ]
Биолюминесцентные бактерии — это светообразующие бактерии , которые преимущественно присутствуют в морской воде, морских отложениях, на поверхности разлагающейся рыбы и в кишечнике морских животных. Бактериальная биолюминесценция встречается не так часто, но также встречается у наземных и пресноводных бактерий. [126] Эти бактерии могут жить свободно (например, Vibrio harveyi ) или находиться в симбиозе с животными, такими как гавайский бобтейл-кальмар ( Aliivibrio fischeri ) или наземными нематодами ( Photorhabdus luminescens ). Организмы-хозяева обеспечивают этим бактериям безопасный дом и достаточное питание. Взамен хозяева используют свет, производимый бактериями, для маскировки, привлечения добычи и/или партнера. Биолюминесцентные бактерии развили симбиотические отношения с другими организмами, в которых оба участника получают почти одинаковую выгоду. [150] Другая возможная причина, по которой бактерии используют реакцию люминесценции, — это чувство кворума , способность регулировать экспрессию генов в ответ на плотность бактериальных клеток. [151]
Кальмар гавайского бобтейла живет в симбиозе с биолюминесцентными бактериями Aliivibrio fischeri, населяющими специальный световой орган в мантии кальмара. Бактерии получают от кальмара сахар и аминокислоты и, в свою очередь, скрывают силуэт кальмара, если смотреть снизу, контросвещая его, соответствуя количеству света, падающего на верхнюю часть мантии. [152] Кальмар служит модельным организмом для животно-бактериального симбиоза, и его взаимоотношения с бактериями широко изучены.
Vibrio harveyi представляет собой палочковидную, подвижную (с помощью полярных жгутиков ) биолюминесцентную бактерию, которая оптимально растет при температуре от 30 до 35 °C (от 86 до 95 °F). Его можно обнаружить в свободном плавании в тропических морских водах, комменсально в микрофлоре кишечника морских животных, а также в качестве первичного и условно-патогенного микроорганизма у ряда морских животных. [153] Считается, что это является причиной эффекта молочного моря , когда морская вода в ночное время излучает однородное голубое свечение. Некоторые сияния могут покрывать почти 6000 квадратных миль (16 000 км2). 2 ).
Микробный родопсин
[ редактировать ]
(2) он меняет свою конфигурацию, поэтому протон вылетает из клетки
(3) химический потенциал заставляет протон возвращаться в клетку
(4) таким образом генерируя энергию
(5) в форме аденозинтрифосфата . [154]
Фототрофический метаболизм основан на использовании одного из трех пигментов, преобразующих энергию: хлорофилла , бактериохлорофилла и ретиналя . Ретиналь – это хромофор, содержащийся в родопсинах . О значении хлорофилла в преобразовании световой энергии пишут уже несколько десятилетий, однако фототрофия на основе пигментов сетчатки только начинает изучаться. [155]

В 2000 году группа микробиологов под руководством Эдварда Делонга сделала важнейшее открытие в понимании морских углеродных и энергетических циклов. Они обнаружили ген у нескольких видов бактерий [157] [158] отвечает за выработку белка родопсина , ранее не встречавшегося у бактерий. Эти белки, обнаруженные в клеточных мембранах, способны преобразовывать энергию света в биохимическую энергию за счет изменения конфигурации молекулы родопсина при попадании на нее солнечного света, вызывая перекачку протона изнутри наружу и последующий приток, генерирующий энергию. [159] Археоподобные родопсины впоследствии были обнаружены среди различных таксонов, простейших, а также у бактерий и архей, хотя они редки в сложных многоклеточных организмах . [160] [161] [162]
Исследования 2019 года показывают, что эти «похищающие солнце бактерии» более распространены, чем считалось ранее, и могут изменить влияние глобального потепления на океаны. «Результаты расходятся с традиционной интерпретацией морской экологии, представленной в учебниках, которая утверждает, что почти весь солнечный свет в океане улавливается хлорофиллом в водорослях. Вместо этого бактерии, оснащенные родопсином, действуют как гибридные автомобили, приводимые в движение органическими веществами, когда они доступны, большинство бактерий — и под воздействием солнечного света, когда питательных веществ не хватает». [163] [155]
Существует астробиологическая гипотеза, называемая гипотезой пурпурной Земли , которая предполагает, что первоначальные формы жизни на Земле были основаны на сетчатке глаза, а не на хлорофилле, из-за чего Земля выглядела фиолетовой, а не зеленой. [164] [165]
Симбиоз
[ редактировать ]Некоторые морские организмы вступают в симбиоз с бактериями или архей. Черви Помпеи живут на больших глубинах у гидротермальных источников при температуре до 80 ° C (176 ° F). У них что-то вроде волосатой спины, но на самом деле эти «волосы» представляют собой колонии бактерий, таких как Nautilia profundicola , которые, как полагают, обеспечивают червю некоторую степень изоляции. Железы на спине червя выделяют слизь, которой питаются бактерии, что является формой симбиоза .
 «Волосатые» спинки червей Помпеи представляют собой колонии симбиотических бактерий.
«Волосатые» спинки червей Помпеи представляют собой колонии симбиотических бактерий. Hesiocaeca methanicola обитает на больших глубинах метанового льда и, по-видимому, выживает в симбиозе с бактериями, метаболизируют клатрат которые . [166]
Hesiocaeca methanicola обитает на больших глубинах метанового льда и, по-видимому, выживает в симбиозе с бактериями, метаболизируют клатрат которые . [166]
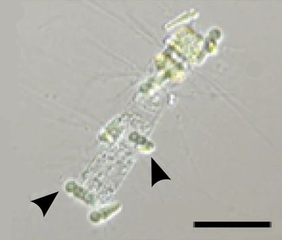 Эпифитные цианобактерии Calothrix (стрелки) в симбиозе с диатомеей Chaetoceros . Масштабная линейка 50 мкм.
Эпифитные цианобактерии Calothrix (стрелки) в симбиозе с диатомеей Chaetoceros . Масштабная линейка 50 мкм.




Эндосимбионтные бактерии — это бактерии, живущие внутри тела или клеток другого организма. Некоторые типы цианобактерий являются эндосимбионтами , и было обнаружено, что цианобактерии обладают генами, которые позволяют им подвергаться азотфиксации. [168]
Организмы обычно вступают в симбиотические отношения из-за ограниченности ресурсов в среде обитания или из-за ограничения источника пищи. Симбиотические хемосинтезирующие бактерии, обнаруженные в связи с мидиями ( Bathymodiolus ), расположенными вблизи гидротермальных источников, имеют ген, который позволяет им использовать водород в качестве источника энергии, а не серу или метан в качестве источника энергии для производства энергии. [169]
Olavius algarvensis — это червь, который обитает в прибрежных отложениях Средиземноморья и питается симбиотическими бактериями. Он живет с пятью различными видами бактерий, расположенными под его кутикулой: двумя сульфидокисляющими , двумя сульфатредуцирующими и одной спирохетой . Симбиотические бактерии также позволяют червю использовать водород и угарный газ в качестве источников энергии и метаболизировать органические соединения, такие как малат и ацетат . [170] [171]
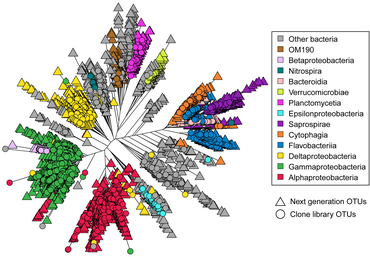
Astrangia poculata , северный звездчатый коралл, представляет собой каменистый коралл умеренного пояса , широко встречающийся вдоль восточного побережья Соединенных Штатов. Коралл может жить как с зооксантеллами (водорослевыми симбионтами), так и без них, что делает его идеальным модельным организмом для изучения взаимодействий микробного сообщества, связанных с симбиотическим состоянием. Однако возможность разработки праймеров и зондов для более специфичного воздействия на ключевые микробные группы затруднена из-за отсутствия полноразмерных последовательностей 16S рРНК , поскольку последовательности, полученные с помощью платформы Illumina, имеют недостаточную длину (около 250 пар оснований) для создания праймеры и зонды. [172] В 2019 году Голдсмит и др. продемонстрировало, что секвенирование по Сэнгеру способно воспроизводить биологически значимое разнообразие, обнаруженное с помощью более глубокого секвенирования следующего поколения , а также создавать более длинные последовательности, полезные исследовательскому сообществу для разработки зондов и праймеров (см. Диаграмму справа). [173]
Роль в морских пищевых сетях
[ редактировать ]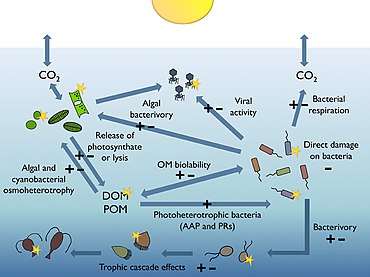
Большая часть объема Мирового океана находится во тьме. Процессы, происходящие в тонком освещенном приземном слое ( фототическом слое от поверхности до глубин от 50 до 170 метров), имеют важное значение для глобальной биосферы. Например, видимая область солнечного спектра (так называемое фотосинтетически доступное излучение или ФАР), достигающая этого освещенного солнцем слоя, обеспечивает около половины первичной продуктивности планеты и отвечает за примерно половину атмосферного кислорода, необходимого для большей части жизни. на Земле. [175] [176]
Гетеротрофный бактериопланктон является основным потребителем растворенного органического вещества (РОВ) в пелагических морских пищевых цепях , включая освещенные солнцем верхние слои океана. Их чувствительность к ультрафиолетовому излучению (УФИ), а также некоторые недавно открытые механизмы, которые бактерии развили в процессе эволюции, чтобы извлечь выгоду из фотосинтетически доступного излучения (ФАР), позволяют предположить, что естественный солнечный свет играет важную, но трудно предсказуемую роль в модуляции бактериальных биогеохимических функций в океанах. [174]
Среда обитания на поверхности океана находится на границе между атмосферой и океаном. Биопленочная среда обитания на поверхности океана является местом обитания обитающих на поверхности микроорганизмов, обычно называемых нейстоном . Этот обширный интерфейс воздух-вода находится на пересечении основных процессов воздухообмена и воды, охватывая более 70% площади мировой поверхности. Бактерии в поверхностном микрослое океана, называемые бактерионейстоном , представляют интерес из-за практических применений, таких как газообмен между воздухом и морем парниковых газов, производство климатически активных морских аэрозолей и дистанционное зондирование океана. [177] Особый интерес представляет производство и разложение поверхностно-активных веществ (поверхностно-активных материалов) посредством микробных биохимических процессов. Основными источниками поверхностно-активных веществ в открытом океане являются фитопланктон, [178] земной сток и осаждение из атмосферы. [177]

В отличие от цветения цветных водорослей, бактерии, связанные с поверхностно-активными веществами, могут быть не видны на цветных изображениях океана. Возможность обнаруживать эти «невидимые» бактерии, связанные с поверхностно-активными веществами, с помощью радара с синтезированной апертурой дает огромные преимущества в любых погодных условиях, независимо от облачности, тумана или дневного света. [177] Это особенно важно при очень сильных ветрах, поскольку именно в этих условиях происходит наиболее интенсивный газообмен между воздухом и морем и образование морских аэрозолей. Таким образом, в дополнение к цветным спутниковым изображениям спутниковые изображения SAR могут дать дополнительную информацию о глобальной картине биофизических процессов на границе океана и атмосферы, обмене парниковых газов между воздухом и морем и образовании климатически активных морских аэрозолей. [177]
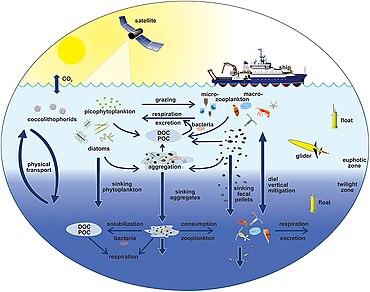
океана На диаграмме справа показаны связи между биологическим насосом и пелагической пищевой сетью, а также возможность удаленного отбора проб этих компонентов с кораблей, спутников и автономных транспортных средств. Светло-голубые воды представляют собой эвфотическую зону , а более темные голубые воды представляют собой сумеречную зону . [179]
Роль в биогеохимическом цикле
[ редактировать ]Археи перерабатывают такие элементы, как углерод , азот и сера , в различных средах обитания. [180] Археи выполняют множество этапов азотного цикла . Сюда входят как реакции, которые удаляют азот из экосистем (например, нитратное дыхание и денитрификация ), так и процессы, которые вносят азот (например, ассимиляция нитратов и фиксация азота ). [181] [182]
Недавно исследователи обнаружили участие архей в реакциях окисления аммиака . Эти реакции особенно важны в океанах. [183] [184] В цикле серы археи, которые растут за счет окисления соединений серы , выделяют этот элемент из горных пород, делая его доступным для других организмов, но археи, которые делают это, такие как Sulfolobus , производят серную кислоту в качестве побочного продукта, и рост этих организмов в заброшенных шахтах может способствовать кислотному дренажу шахт и другому экологическому ущербу. [185] В углеродном цикле метаногенные археи удаляют водород и играют важную роль в разложении органического вещества популяциями микроорганизмов, которые действуют как разлагатели в анаэробных экосистемах, таких как отложения и болота. [186]
См. также
[ редактировать ]- Методы подсчета бактериопланктона
- Биолюминесцентные бактерии
- Железоокисляющие бактерии
- Pelagibacterales – модельные организмы в теории оптимизации
- Теория оптимизации
Ссылки
[ редактировать ]- ^ Янгсон Р.М. (2006). Словарь Коллинза по биологии человека . Глазго: ХарперКоллинз. ISBN 978-0-00-722134-9 .
- ^ Нельсон Д.Л., Кокс М.М. (2005). Ленингерские принципы биохимии (4-е изд.). Нью-Йорк: WH Freeman. ISBN 978-0-7167-4339-2 .
- ^ Мартин Э.А., изд. (1983). Словарь наук о жизни Macmillan (2-е изд.). Лондон: Макмиллан Пресс. ISBN 978-0-333-34867-3 .
- ^ Фюрст Дж. А. (2010). «Помимо прокариотов и эукариотов: планктомицеты и клеточная организация» . Природное образование . 3 (9): 44.
- ^ Данные ВОЗ о смертности за 2002 г. По состоянию на 20 января 2007 г.
- ^ Университет Джорджии (10 декабря 2015 г.). «Функции микробиома глобального океана являются ключом к пониманию изменений окружающей среды» . www.sciencedaily.com . Проверено 11 декабря 2015 г.
- ^ Бар-Он Ю.М., Филлипс Р., Майло Р. (2018). «Распределение биомассы на Земле» (PDF) . Труды Национальной академии наук Соединенных Штатов Америки . 115 (25): 6506–6511. Бибкод : 1998PNAS...95.6578W . дои : 10.1073/pnas.1711842115 . ПМК 6016768 . ПМИД 29784790 .
- ^ Перейти обратно: а б Доглиони С., Пиньятти Дж., Коулман М. (2016). «Почему жизнь на поверхности Земли возникла в кембрии?» . Геонаучные границы . 7 (6): 865–873. Бибкод : 2016GeoFr...7..865D . дои : 10.1016/j.gsf.2016.02.001 . hdl : 11573/925124 .
- ^ «Возраст Земли» . Геологическая служба США . 9 июля 2007 года . Проверено 31 мая 2015 г.
- ^ Далримпл 2001 , стр. 205–221.
- ^ Манхеса Дж., Аллегре СиДжей , Дюпреа Б., Хамелен Б. (май 1980 г.). «Изотопное исследование свинца основных-ультраосновных слоистых комплексов: предположения о возрасте Земли и характеристиках примитивной мантии». Письма о Земле и планетологии . 47 (3): 370–382. Бибкод : 1980E&PSL..47..370M . дои : 10.1016/0012-821X(80)90024-2 .
- ^ Шопф Й.В. , Кудрявцев А.Б., Чая А.Д., Трипати А.Б. (5 октября 2007 г.). «Свидетельства архейской жизни: строматолиты и микроокаменелости». Докембрийские исследования . 158 (3–4): 141–155. Бибкод : 2007PreR..158..141S . doi : 10.1016/j.precamres.2007.04.009 .
- ^ Рэйвен и Джонсон 2002 , с. 68
- ^ Баумгартнер Р.Дж. и др. (2019). «Нанопористый пирит и органическое вещество в строматолитах возрастом 3,5 миллиарда лет свидетельствуют о первозданной жизни» (PDF) . Геология . 47 (11): 1039–1043. Бибкод : 2019Geo....47.1039B . дои : 10.1130/G46365.1 . S2CID 204258554 .
- ^ Самые ранние признаки жизни: ученые находят остатки микробов в древних камнях Phys.org . 26 сентября 2019 г.
- ^ Яблонски Д. (25 июня 1999 г.). «Будущее ископаемых пластинок». Наука . 284 (5423): 2114–2116. дои : 10.1126/science.284.5423.2114 . ПМИД 10381868 . S2CID 43388925 .
- ^ Перейти обратно: а б Кавалер-Смит Т. (29 июня 2006 г.). «Эволюция клеток и история Земли: застой и революция» . Философские труды Королевского общества B: Биологические науки . 361 (1470): 969–1006. дои : 10.1098/rstb.2006.1842 . ПМЦ 1578732 . ПМИД 16754610 .
- ^ Шопф JW (29 июня 2006 г.). «Ископаемые свидетельства архейской жизни» . Философские труды Королевского общества B: Биологические науки . 361 (1470): 869–885. дои : 10.1098/rstb.2006.1834 . ПМЦ 1578735 . ПМИД 16754604 .
- Альтерманн В., Казмерчак Дж. (ноябрь 2003 г.). «Архейские микроокаменелости: переоценка ранней жизни на Земле» . Исследования в области микробиологии . 154 (9): 611–617. дои : 10.1016/j.resmic.2003.08.006 . ПМИД 14596897 .
- ^ Шопф JW (19 июля 1994 г.). «Разные темпы, разные судьбы: темп и способ эволюции менялись от докембрия к фанерозою» . Труды Национальной академии наук Соединенных Штатов Америки . 91 (15): 6735–6742. Бибкод : 1994PNAS...91.6735S . дои : 10.1073/pnas.91.15.6735 . ПМЦ 44277 . ПМИД 8041691 .
- ^ Пул А.М., Пенни Д. (январь 2007 г.). «Оценка гипотез происхождения эукариот». Биоэссе . 29 (1): 74–84. doi : 10.1002/bies.20516 . ПМИД 17187354 .
- ^ Перейти обратно: а б Дьялл С.Д., Браун М.Т., Джонсон П.Дж. (9 апреля 2004 г.). «Древние вторжения: от эндосимбионтов к органеллам». Наука . 304 (5668): 253–257. Бибкод : 2004Sci...304..253D . дои : 10.1126/science.1094884 . ПМИД 15073369 . S2CID 19424594 .
- ^ Мартин В. (октябрь 2005 г.). «Недостающее звено между гидрогеносомами и митохондриями». Тенденции в микробиологии . 13 (10): 457–459. дои : 10.1016/j.tim.2005.08.005 . ПМИД 16109488 .
- ^ Ланг Б.Ф., Грей М.В., Бургер Дж. (декабрь 1999 г.). «Эволюция митохондриального генома и происхождение эукариот». Ежегодный обзор генетики . 33 : 351–397. дои : 10.1146/annurev.genet.33.1.351 . ПМИД 10690412 .
- Макфадден Дж.И. (1 декабря 1999 г.). «Эндосимбиоз и эволюция растительной клетки». Современное мнение в области биологии растений . 2 (6): 513–519. Бибкод : 1999COPB....2..513M . дои : 10.1016/S1369-5266(99)00025-4 . ПМИД 10607659 .
- ^ Обнимаю, Лаура А.; Бейкер, Бретт Дж.; Анантараман, Картик; Браун, Кристофер Т.; Пробст, Александр Дж.; Кастель, Синди Дж.; Баттерфилд, Кристина Н.; Хернсдорф, Алекс В.; Амано, Юки; Исе, Котаро; Сузуки, Йохей (11 апреля 2016 г.). «Новый взгляд на древо жизни» . Природная микробиология . 1 (5): 16048. doi : 10.1038/nmicrobiol.2016.48 . ISSN 2058-5276 . ПМИД 27572647 .
- ^ Чиккарелли Ф.Д., Доркс Т., фон Меринг С. и др. (3 марта 2006 г.). «На пути к автоматической реконструкции древа жизни с высоким разрешением». Наука . 311 (5765): 1283–1287. Бибкод : 2006Sci...311.1283C . CiteSeerX 10.1.1.381.9514 . дои : 10.1126/science.1123061 . ПМИД 16513982 . S2CID 1615592 .
- ^ Делонг Э.Ф. , Пейс Н.Р. (1 августа 2001 г.). «Экологическое разнообразие бактерий и архей». Систематическая биология . 50 (4): 470–478. CiteSeerX 10.1.1.321.8828 . дои : 10.1080/106351501750435040 . ПМИД 12116647 .
- ^ Кайзер Д. (декабрь 2001 г.). «Построение многоклеточного организма». Ежегодный обзор генетики . 35 : 103–123. дои : 10.1146/annurev.genet.35.102401.090145 . ПМИД 11700279 . S2CID 18276422 .
- ^ Циммер С (7 января 2016 г.). «Генетический переворот помог организмам перейти от одной клетки к множеству» . Нью-Йорк Таймс . Проверено 7 января 2016 г.
- ^ Валентайн Дж.В. , Яблонски Д., Эрвин Д.Х. (1 марта 1999 г.). «Ископаемые, молекулы и эмбрионы: новые взгляды на кембрийский взрыв» . Разработка . 126 (5): 851–859. дои : 10.1242/dev.126.5.851 . ПМИД 9927587 . Проверено 30 декабря 2014 г.
- ^ Оно С. (январь 1997 г.). «Причина, а также последствия кембрийского взрыва в эволюции животных». Журнал молекулярной эволюции . 44 (Приложение 1): С23–С27. Бибкод : 1997JMolE..44S..23O . дои : 10.1007/PL00000055 . ПМИД 9071008 . S2CID 21879320 .
- Валентин Дж.В., Яблонски Д. (2003). «Морфологическая макроэволюция и развитие: палеонтологическая перспектива» . Международный журнал биологии развития . 47 (7–8): 517–522. ПМИД 14756327 . Проверено 30 декабря 2014 г.
- ^ Кэмпбелл, Н. «Биология: концепции и связи». Пирсон Образование. Сан-Франциско: 2003.
- ^ Харпер, Дуглас. «прокариот» . Интернет-словарь этимологии .
- ^ Харпер, Дуглас. «эукариотический» . Интернет-словарь этимологии .
- ^ Станье Р.Ю. , Ван Нил CB (1962). «Понятие о бактерии». Архив микробиологии . 42 :17-35. дои : 10.1007/BF00425185 . ПМИД 13916221 . S2CID 29859498 .
- ^ Woese CR, Fox GE (ноябрь 1977 г.). «Филогенетическая структура прокариотического домена: первичные царства» . Труды Национальной академии наук Соединенных Штатов Америки . 74 (11): 5088–90. Бибкод : 1977PNAS...74.5088W . дои : 10.1073/pnas.74.11.5088 . ПМК 432104 . ПМИД 270744 .
- ^ Вёзе Ч.Р., Кандлер О., Уилис М.Л. (июнь 1990 г.). «На пути к естественной системе организмов: предложение по доменам архей, бактерий и эукариев» . Труды Национальной академии наук Соединенных Штатов Америки . 87 (12): 4576–9. Бибкод : 1990PNAS...87.4576W . дои : 10.1073/pnas.87.12.4576 . ПМК 54159 . ПМИД 2112744 .
- ^ Отомо, Йоко; Какегава, Такеши; Исида, Акизуми; Нагасе, Тосиро; Розинг, Миник Т. (январь 2014 г.). «Свидетельства наличия биогенного графита в метаосадочных породах раннего архея Исуа». Природа Геонауки . 7 (1): 25–28. Бибкод : 2014NatGe...7...25O . дои : 10.1038/ngeo2025 .
- ^ Натман, Аллен П.; Беннетт, Вики С.; Друг, Кларк Р.Л.; Кранендонк, Мартин Дж. Ван; Чивас, Аллан Р. (сентябрь 2016 г.). «Быстрое возникновение жизни, продемонстрированное открытием микробных структур возрастом 3700 миллионов лет» . Природа . 537 (7621): 535–538. Бибкод : 2016Natur.537..535N . дои : 10.1038/nature19355 . ПМИД 27580034 . S2CID 205250494 .
- ^ Боренштейн, Сет (19 октября 2015 г.). «Намеки на жизнь на ранней Земле, которая считалась пустынной» . Возбуждайте . Йонкерс, Нью-Йорк: Интерактивная сеть Mindspark . Ассошиэйтед Пресс . Архивировано из оригинала 23 октября 2015 года . Проверено 8 октября 2018 г.
- ^ Белл, Элизабет А.; Бенике, Патрик; Харрисон, Т. Марк; и др. (19 октября 2015 г.). «Потенциально биогенный углерод сохранился в цирконе возрастом 4,1 миллиарда лет» . Учеб. Натл. акад. наук. США . 112 (47): 14518–21. Бибкод : 2015PNAS..11214518B . дои : 10.1073/pnas.1517557112 . ПМЦ 4664351 . ПМИД 26483481 .
- ^ Додд, Мэтью С.; Папино, Доминик; Гренне, Тор; слабый, Джон Ф.; Риттнер, Мартин; Пирайно, Франко; О'Нил, Джонатан; Литтл, Криспин Т.С. (2 марта 2017 г.). «Доказательства ранней жизни в осадках старейших гидротермальных источников Земли» (PDF) . Природа . 543 (7643): 60–64. Бибкод : 2017Natur.543...60D . дои : 10.1038/nature21377 . ПМИД 28252057 . S2CID 2420384 .
- ^ Циммер, Карл (1 марта 2017 г.). «Ученые говорят, что окаменелости канадских бактерий могут быть древнейшими на Земле» . Нью-Йорк Таймс . Проверено 2 марта 2017 г.
- ^ Нисбет, Юан Г.; Фаулер, CMR (7 декабря 1999 г.). «Архейская метаболическая эволюция микробных матов» . Труды Лондонского королевского общества Б. 266 (1436): 2375–2382. дои : 10.1098/rspb.1999.0934 . ПМК 1690475 .
- ^ Анбар, Ариэль Д.; Юн Дуань; Лайонс, Тимоти В.; и др. (28 сентября 2007 г.). «Запах кислорода перед великим событием окисления?». Наука . 317 (5846): 1903–1906. Бибкод : 2007Sci...317.1903A . дои : 10.1126/science.1140325 . ПМИД 17901330 . S2CID 25260892 .
- ^ Нолл, Эндрю Х .; Жаво, Эммануэль Ж.; Хьюитт, Дэвид; Коэн, Фиби (29 июня 2006 г.). «Эукариотические организмы в протерозойских океанах» . Философские труды Королевского общества Б. 361 (1470): 1023–1038. дои : 10.1098/rstb.2006.1843 . ПМЦ 1578724 . ПМИД 16754612 .
- ^ Федонкин, Михаил А. (31 марта 2003 г.). «Происхождение Metazoa в свете протерозойской летописи окаменелостей» (PDF) . Палеонтологические исследования . 7 (1): 9–41. дои : 10.2517/prpsj.7.9 . S2CID 55178329 . Архивировано из оригинала (PDF) 26 февраля 2009 года . Проверено 2 сентября 2008 г.
- ^ Боннер, Джон Тайлер (1998). «Истоки многоклеточности». Интегративная биология . 1 (1): 27–36. doi : 10.1002/(SICI)1520-6602(1998)1:1<27::AID-INBI4>3.0.CO;2-6 .
- ^ Мэй, Р.М. (1988) «Сколько видов существует на Земле?». Science , 241 (4872): 1441–1449. дои : 10.1126/science.241.4872.1441 .
- ^ Лоси, К.Дж. и Леннон, Дж.Т. (2016) «Законы масштабирования предсказывают глобальное микробное разнообразие». Труды Национальной академии наук , 113 (21): 5970–5975. дои : 10.1073/pnas.1521291113 .
- ^ Виторино, Л.К. и Бесса, Лос-Анджелес (2018) «Микробное разнообразие: разрыв между оценками и известными». Разнообразие , 10 (2):46. два : 10.3390/d10020046 .
 Измененный текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
Измененный текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 . - ^ Живые бактерии движутся в воздушных потоках Земли Журнал Smithsonian , 11 января 2016 г.
- ^ Роббинс, Джим (13 апреля 2018 г.). «Триллионы и триллионы вирусов падают с неба каждый день» . Нью-Йорк Таймс . Проверено 14 апреля 2018 г.
- ^ Рече, Изабель; Д'Орта, Гаэтано; Младенов, Наталья; Вингет, Даниэль М; Саттл, Кертис А. (29 января 2018 г.). «Скорость осаждения вирусов и бактерий над пограничным слоем атмосферы» . Журнал ИСМЕ . 12 (4): 1154–1162. Бибкод : 2018ISMEJ..12.1154R . дои : 10.1038/s41396-017-0042-4 . ПМК 5864199 . ПМИД 29379178 .
- ^ Саттл, Калифорния (2005). «Вирусы в море». Природа . 437 (9): 356–361. Бибкод : 2005Natur.437..356S . дои : 10.1038/nature04160 . ПМИД 16163346 . S2CID 4370363 .
- ^ Шорс 2017 , с. 5
- ^ Шорс 2017 , с. 593
- ^ Саттл, Калифорния (2007). «Морские вирусы — основные игроки глобальной экосистемы». Обзоры природы Микробиология . 5 (10): 801–12. дои : 10.1038/nrmicro1750 . ПМИД 17853907 . S2CID 4658457 .
- ^ Шульц, Кестин; Смит, Мария В.; Херфорт, Лиди; Саймон, Холли М. (2018). «Невидимый мир в реке» . Границы для молодых умов . 6 . дои : 10.3389/фрим.2018.00004 . S2CID 3344238 . Измененный текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Стоекениус В. (октябрь 1981 г.). «Квадратная бактерия Уолсби: тонкая структура ортогонального прокариота» . Журнал бактериологии . 148 (1): 352–60. дои : 10.1128/JB.148.1.352-360.1981 . ПМК 216199 . ПМИД 7287626 .
- ^ Дарем, Бриндан П.; Гроте, Яна; Уиттакер, Керри А.; Бендер, Сара Дж.; Ло, Хайвэй; Грим, Шэрон Л.; Браун, Джулия М.; Кейси, Джон Р.; Дрон, Энтони; Флорес-Лейва, Леннин; Крупке, Андреас; Лурия, Екатерина М.; Мой, Арик Х.; Нигро, Оливия Д.; Патер, Сантиска; Талармин, Агата; Уир, Эмма К.; Вебер, Томас С.; Уилсон, Джесси М.; Черч, Мэтью Дж.; Делонг, Эдвард Ф.; Карл, Дэвид М.; Стюард, Григ Ф.; Эппли, Джон М.; Кирпидес, Никос К.; Шустер, Стефан; Раппе, Майкл С. (2014). «Проект последовательности генома морского альфапротеобактериального штамма HIMB11, первого культивируемого представителя уникальной линии внутри клады Roseobacter, обладающего необычно маленьким геномом» . Стандарты в геномных науках . 9 (3): 632–645. дои : 10.4056/sig.4998989 . ПМК 4148974 . ПМИД 25197450 .
- ^ Фредриксон Дж.К., Захара Дж.М., Балквилл Д.Л., Кеннеди Д., Ли С.М., Костандаритес Х.М., Дейли М.Дж., Ромин М.Ф., Брокман Ф.Дж. (2004). «Геомикробиология вадозных отложений, сильно загрязненных ядерными отходами, на полигоне в Хэнфорде, штат Вашингтон» . Прикладная и экологическая микробиология . 70 (7): 4230–41. Бибкод : 2004ApEnM..70.4230F . дои : 10.1128/АЕМ.70.7.4230-4241.2004 . ПМК 444790 . ПМИД 15240306 .
- ^ Вёзе Ч.Р., Кандлер О., Уилис М.Л. (1990). «На пути к естественной системе организмов: предложение по доменам архей, бактерий и эукариев» . Труды Национальной академии наук Соединенных Штатов Америки . 87 (12): 4576–9. Бибкод : 1990PNAS...87.4576W . дои : 10.1073/pnas.87.12.4576 . ПМК 54159 . ПМИД 2112744 .
- ^ Шопф Дж.В. (1994). «Разные темпы, разные судьбы: темп и способ эволюции менялись от докембрия к фанерозою» . Труды Национальной академии наук Соединенных Штатов Америки . 91 (15): 6735–42. Бибкод : 1994PNAS...91.6735S . дои : 10.1073/pnas.91.15.6735 . ПМЦ 44277 . ПМИД 8041691 .
- ^ Делонг Э.Ф., Пейс Н.Р. (2001). «Экологическое разнообразие бактерий и архей». Систематическая биология . 50 (4): 470–8. CiteSeerX 10.1.1.321.8828 . дои : 10.1080/106351501750435040 . ПМИД 12116647 .
- ^ Браун-младший, Дулиттл В.Ф. (1997). «Археи и переход от прокариот к эукариотам» . Обзоры микробиологии и молекулярной биологии . 61 (4): 456–502. doi : 10.1128/mmbr.61.4.456-502.1997 . ПМК 232621 . ПМИД 9409149 .
- ^ Пул А.М., Пенни Д. (2007). «Оценка гипотез происхождения эукариот». Биоэссе . 29 (1): 74–84. doi : 10.1002/bies.20516 . ПМИД 17187354 . S2CID 36026766 .
- ^ Ланг Б.Ф., Грей М.В., Бургер Дж. (1999). «Эволюция митохондриального генома и происхождение эукариот». Ежегодный обзор генетики . 33 : 351–97. дои : 10.1146/annurev.genet.33.1.351 . ПМИД 10690412 .
- ^ Макфадден Дж.И. (1999). «Эндосимбиоз и эволюция растительной клетки». Современное мнение в области биологии растений . 2 (6): 513–9. Бибкод : 1999COPB....2..513M . дои : 10.1016/S1369-5266(99)00025-4 . ПМИД 10607659 .
- ^ Кох А.Л. (2002). «Контроль бактериального клеточного цикла путем роста цитоплазмы». Критические обзоры по микробиологии . 28 (1): 61–77. дои : 10.1080/1040-840291046696 . ПМИД 12003041 . S2CID 11624182 .
- ^ Иагон Р.Г. (апрель 1962 г.). «Pseudomonas natriegens, морская бактерия со временем генерации менее 10 минут» . Журнал бактериологии . 83 (4): 736–37. дои : 10.1128/jb.83.4.736-737.1962 . ПМК 279347 . ПМИД 13888946 .
- ^ "Кандидат Pelagibacter Ubique". Европейский институт биоинформатики. Европейский институт биоинформатики, 2011. Интернет. 8 января 2012 г. http://www.ebi.ac.uk/2can/genomes/bacteria/Candidatus_Pelagibacter_ubique.html. Архивировано 1 декабря 2008 г. в Wayback Machine.
- ^ «Блошиный рынок: недавно обнаруженный вирус может оказаться самым распространенным организмом на планете» . Экономист . 16 февраля 2013 года . Проверено 16 февраля 2013 г.
- ^ Чжао, Ю.; Темпертон, Б.; Трэш, Джей Си; Швальбах, М.С.; Вергин, КЛ; Лэндри, ЗК; Эллисман, М.; Диринк, Т.; Салливан, МБ; Джованнони, SJ (2013). «Обилие вирусов SAR11 в океане». Природа . 494 (7437): 357–360. Бибкод : 2013Natur.494..357Z . дои : 10.1038/nature11921 . ПМИД 23407494 . S2CID 4348619 .
- ^ Бентзон-Тилиа М., Грам Л. (2017). «Биотехнологическое применение группы Roseobacter». Биоразведка . Темы биоразнообразия и охраны природы. Том. 16. Спрингер, Чам. стр. 137–166. дои : 10.1007/978-3-319-47935-4_7 . ISBN 978-3-319-47933-0 .
- ^ Браузер таксономии NCBI: Национальный центр биотехнологической информации Roseobacter . Доступ: 8 мая 2020 г.
- ^ «Самая большая бактерия: ученый обнаруживает новую бактериальную форму жизни у африканского побережья» , Институт морской микробиологии Макса Планка , 8 апреля 1999 г., заархивировано из оригинала 20 января 2010 г.
- ^ Список названий прокариот, имеющих место в номенклатуре - Род Thiomargarita
- ^ Изменения концентрации кислорода в нашем океане могут нарушить фундаментальные биологические циклы Phys.org , 25 ноября 2019 г.
- ^ Уолш П.Дж., Смит С., Флеминг Л., Соло-Габриэль Х., Гервик У.Х., ред. (2 сентября 2011 г.). «Цианобактерии и цианобактериальные токсины» . Океаны и здоровье человека: морские риски и средства правовой защиты . Академическая пресса. стр. 271–296. ISBN 978-0-08-087782-2 .
- ^ «Повышение уровня кислорода — журнал астробиологии» . Журнал «Астробиология» . 30 июля 2003 г. Проверено 6 апреля 2016 г.
- ^ Фланнери, DT; Р. М. Уолтер (2012). «Архейские тафтинговые микробные маты и Великое событие окисления: новый взгляд на древнюю проблему». Австралийский журнал наук о Земле . 59 (1): 1–11. Бибкод : 2012AuJES..59....1F . дои : 10.1080/08120099.2011.607849 . S2CID 53618061 .
- ^ Ротшильд, Линн (сентябрь 2003 г.). «Понять эволюционные механизмы и экологические пределы жизни» . НАСА. Архивировано из оригинала 29 марта 2012 года . Проверено 13 июля 2009 г.
- ^ Басу, Субхаджит; Гледхилл, Марта; Де Бир, Дирк; Прабху Матондкар, генеральный директор; Шакед, Йела (2019). «Колонии морских цианобактерий Trichodesmium взаимодействуют с ассоциированными бактериями, приобретая железо из пыли» . Коммуникационная биология . 2 : 284. doi : 10.1038/s42003-019-0534-z . ПМК 6677733 . ПМИД 31396564 . Измененный текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Патрик Дж. Килинг (2004). «Разнообразие и история эволюции пластид и их хозяев» . Американский журнал ботаники . 91 (10): 1481–1493. дои : 10.3732/ajb.91.10.1481 . ПМИД 21652304 .
- ^ Надис С. (декабрь 2003 г.). «Клетки, управляющие морями» (PDF) . Научный американец . 289 (6): 52–3. Бибкод : 2003SciAm.289f..52N . doi : 10.1038/scientificamerican1203-52 . ПМИД 14631732 . Архивировано из оригинала (PDF) 19 апреля 2014 года . Проверено 2 июня 2019 г.
- ^ «Самый важный микроб, о котором вы никогда не слышали» . npr.org .
- ^ Фломбаум, П.; Гальегос, Дж.Л.; Гордилло, РА; Ринкон, Дж.; Забала, LL; Цзяо, Н.; Карл, DM; Ли, WKW; Ломас, Миссури; Венециано, Д.; Вера, CS; Вругт, Дж.А.; Мартини, AC (2013). «Настоящее и будущее глобальное распространение морских цианобактерий Prochromococcus и Synechococcus» . Труды Национальной академии наук . 110 (24): 9824–9829. Бибкод : 2013PNAS..110.9824F . дои : 10.1073/pnas.1307701110 . ПМЦ 3683724 . ПМИД 23703908 .
- ^ Нэборс, Мюррей В. (2004). Введение в ботанику . Сан-Франциско, Калифорния: ISBN Pearson Education, Inc. 978-0-8053-4416-5 .
- ^ Аллаби, М., изд. (1992). «Водоросли». Краткий словарь ботаники . Оксфорд: Издательство Оксфордского университета.
- ^ Иварс-Мартинес, Елена; д'Аурия, Джузеппе; Родригес-Валера, Франциско; Санчес-Порро, Кристина; Вентоза, Антонио; Джойнт, ИАН; Мюлинг, Мартин (2008). «Биогеография вездесущей морской бактерии Alteromonas macleodiide, определенная с помощью анализа многолокусных последовательностей». Молекулярная экология . 17 (18): 4092–4106. Бибкод : 2008MolEc..17.4092I . дои : 10.1111/j.1365-294x.2008.03883.x . ПМИД 19238708 . S2CID 38830049 .
- ^ Лопес-Перес, Марио; Гонзага, Айтор; Мартин-Куадрадо, Ана-Белен; Онищенко, Ольга; Гавидель, Акбар; Гай, Рохит; Родригес-Валера, Франциско (2012). «Геномы поверхностных изолятов Alteromonas macleodii: жизнь широко распространенного морского оппортунистического копиотрофа» . Научные отчеты . 2 : 696. Бибкод : 2012NatSR...2E.696L . дои : 10.1038/srep00696 . ПМЦ 3458243 . ПМИД 23019517 .
- ^ Банг С., Шмитц Р.А. (2015). «Археи, связанные с человеческими поверхностями: не следует недооценивать» . Обзоры микробиологии FEMS . 39 (5): 631–48. дои : 10.1093/femsre/fuv010 . ПМИД 25907112 .
- ^ Archaea Интернет-словарь этимологии . Проверено 17 августа 2016 г.
- ^ Пейс НР (май 2006 г.). «Время перемен» . Природа . 441 (7091): 289. Бибкод : 2006Natur.441..289P . дои : 10.1038/441289a . ПМИД 16710401 . S2CID 4431143 .
- ^ Стокениус В. (1 октября 1981 г.). «Квадратная бактерия Уолсби: тонкая структура ортогонального прокариота» . Журнал бактериологии . 148 (1): 352–60. дои : 10.1128/JB.148.1.352-360.1981 . ПМК 216199 . ПМИД 7287626 .
- ^ Мэдиган М., Мартинко Дж., ред. (2005). Брок Биология микроорганизмов (11-е изд.). Прентис Холл. ISBN 978-0-13-144329-7 .
- ^ Хубер, Харальд; и др. (2002). «Новый тип архей, представленный наноразмерным гипертермофильным симбионтом». Природа . 417 (6884): 63–67. Бибкод : 2002Natur.417...63H . дои : 10.1038/417063а . ПМИД 11986665 . S2CID 4395094 .
- ^ Брошье, Селин; Грибальдо, С; Живанович, Ю; Конфалоньери, Ф; и др. (2005). «Наноархеи: представители нового типа архей или быстро развивающейся эвриархейной линии, родственной Thermococcales?» . Геномная биология . 6 (5): Р42. дои : 10.1186/gb-2005-6-5-r42 . ПМК 1175954 . ПМИД 15892870 .
- ^ Орельяна, Луис Х.; Бен Фрэнсис, Т.; Крюгер, Карен; Тилинг, Ханно; Мюллер, Мари-Каролина; Фукс, Бернхард М.; Константинидис, Константинос Т.; Аманн, Рудольф И. (2019). «Диференциация ниш среди ежегодно повторяющихся прибрежных морских групп II эвриархеот» . Журнал ISME . 13 (12): 3024–3036. Бибкод : 2019ISMEJ..13.3024O . дои : 10.1038/s41396-019-0491-z . ПМК 6864105 . PMID 31447484 .
- ^ См. особенно рис. 4 в Нишимура, Ёсуке; Хонда, Такаши; Омаэ, Кимихо, Саймон; Ямамото, Кейго; Хингам, Ёсихико; Ёсида, Такаши (2017). в океане» . mSphere Вирусные 2 2 ( Сусуму ; геномы окружающей среды проливают новый свет на взаимодействие вируса и . « ) хозяина ПМИД 28261669 .
- ^ Философ, Алон; Ютин, Наталья; Флорес-Урибе, Хосе; Шарон, Итай; Кунин Евгений Владимирович; Бежа, Одед (2017). «Новые многочисленные океанические вирусы некультивируемых морских эвриархей II группы» . Современная биология . 27 (9): 1362–1368. Бибкод : 2017CBio...27.1362P . дои : 10.1016/j.cub.2017.03.052 . ПМЦ 5434244 . ПМИД 28457865 .
- ^ Ся, Сяоминь; Го, Ван; Лю, Хунбин (2017). «Вариации состава и разнообразия архей Тихого океана в масштабах бассейна» . Границы микробиологии . 8 : 2057. doi : 10.3389/fmicb.2017.02057 . ПМК 5660102 . ПМИД 29109713 .
- ^ Мартин-Куадрадо, Ана-Белен; Гарсиа-Эредиа, Инмакулада; Молто, Айтор Гонзага; Лопес-Убеда, Ребека; Каймс, Николь; Лопес-Гарсия, «Очищение»; Морейра, Дэвид; Родригес-Валера, Франциско (2015). «Новый класс морских эвриархей группы II из Средиземноморского глубокого максимума хлорофилла» . Журнал ISME . 9 (7): 1619–1634. Бибкод : 2015ISMEJ...9.1619M . дои : 10.1038/ismej.2014.249 . ПМЦ 4478702 . ПМИД 25535935 .
- ^ NCBI: Кандидат Посейдониалес (орден)
- ^ NCBI: Группа морской пехоты III
- ^ NCBI: Группа морской пехоты IV
- ^ Зиллиг В. (декабрь 1991 г.). «Сравнительная биохимия архей и бактерий». Текущее мнение в области генетики и развития . 1 (4): 544–51. дои : 10.1016/S0959-437X(05)80206-0 . ПМИД 1822288 .
- ^ Слончевски Дж.Л., Фостер Дж.В. Микробиология: развивающаяся наука (3-е изд.). WW Нортон и компания. стр. 491–44.
- ^ Хоган CM (2010). «Экстремофил» . В Моноссон Э, Кливленд С (ред.). Энциклопедия Земли . Национальный совет науки и окружающей среды.
- ^ Юрков В.В.; Битти, Джей Ти (1998). «Аэробные аноксигенные фототрофные бактерии» . Обзоры микробиологии и молекулярной биологии . 62 (3): 695–724. дои : 10.1128/MMBR.62.3.695-724.1998 . ПМК 98932 . ПМИД 9729607 .
- ^ Лами, Р.; Коттрелл, Монтана; Рас, Дж.; Уллоа, О.; Оберностерер, И.; Клаустр, Х.; Кирчман, Д.Л.; Лебарон, П. (2007). «Высокая численность аэробных аноксигенных фотосинтетических бактерий в южной части Тихого океана» . Прикладная и экологическая микробиология . 73 (13): 4198–205. Бибкод : 2007ApEnM..73.4198L . дои : 10.1128/АЕМ.02652-06 . ЧВК 1932784 . ПМИД 17496136 .
- ^ Еженедельник наук о жизни. (2012). Бактерии; Отчеты Национального исследовательского совета Испании (CSIC) описывают последние достижения в области бактерий. ISSN 1552-2466 . Р.4582.
- ^ Цзяо, Няньчжи; Чжан, Яо; Цзэн, Юнхуэй; Хун, Нин; Лю, Рулонг; Чен, Фэн; Ван, Пинксянь (2007). «Четкая картина распределения численности и разнообразия аэробных аноксигенных фототрофных бактерий в мировом океане». Экологическая микробиология . 9 (12): 3091–3099. Бибкод : 2007EnvMi...9.3091J . дои : 10.1111/j.1462-2920.2007.01419.x . ПМИД 17991036 .
- ^ Лами, Рафаэль; Коттрелл, Мэтью Т.; Рас, ЖозеФин; Уллоа, Освальдо; Оберностерер, Ингрид; Клаустр, Эрве; Кирчман, Дэвид Л.; Лебарон, Филипп (2007). «Высокая численность аэробных аноксигенных фотосинтетических бактерий в южной части Тихого океана» . Прикладная и экологическая микробиология . 73 (13): 4198–4205. Бибкод : 2007ApEnM..73.4198L . дои : 10.1128/АЕМ.02652-06 . ЧВК 1932784 . ПМИД 17496136 .
- ^ Кольбер, З.С. (2001). «Вклад аэробных фотогетеротрофных бактерий в углеродный цикл в океане». Наука . 292 (5526): 2492–2495. дои : 10.1126/science.1059707 . ПМИД 11431568 . S2CID 1970984 .
- ^ Араньо М., Шлегель Х.Г. (1981). «Водородокисляющие бактерии». В Старр М.П., Столп Х., Трюпер Х.Г., Баловс А., Шлегель Х.Г. (ред.). Прокариоты . Берлин, Гейдельберг: Springer. стр. 865–893. дои : 10.1007/978-3-662-13187-9_70 . ISBN 978-3-662-13187-9 .
- ^ Перейти обратно: а б Альберс С.В., Джаррелл К.Ф. (27 января 2015 г.). «Архееллум: как плавают археи» . Границы микробиологии . 6:23 . дои : 10.3389/fmicb.2015.00023 . ПМЦ 4307647 . ПМИД 25699024 .
- ^ Сильверман М., Саймон М. (май 1974 г.). «Вращение жгутиков и механизм подвижности бактерий». Природа . 249 (452): 73–4. Бибкод : 1974Natur.249...73S . дои : 10.1038/249073a0 . ПМИД 4598030 . S2CID 10370084 .
- ^ Мейстер Г.Л., Берг Х.К. (1987). «Быстрое вращение пучков жгутиков у плавающих бактерий». Природа . 325 (6105): 637–640. Бибкод : 1987Natur.325..637L . дои : 10.1038/325637a0 . S2CID 4242129 .
- ^ Берг Х.К., Андерсон Р.А. (октябрь 1973 г.). «Бактерии плавают, вращая свои жгутиковые нити». Природа . 245 (5425): 380–2. Бибкод : 1973Natur.245..380B . дои : 10.1038/245380a0 . ПМИД 4593496 . S2CID 4173914 .
- ^ Ян Т.Л., Бови ЕС (1965). «Передвижение и локомоция микроорганизмов». Ежегодный обзор микробиологии . 19 :21–58. дои : 10.1146/annurev.mi.19.100165.000321 . ПМИД 5318439 .
- ^ Харши РМ (2003). «Подвижность бактерий на поверхности: много путей к общей цели». Ежегодный обзор микробиологии . 57 : 249–73. дои : 10.1146/annurev.micro.57.030502.091014 . ПМИД 14527279 .
- ^ Джаррелл К. (2009). «Археальные жгутики и пили». Пили и жгутики: текущие исследования и будущие тенденции . Кайстер Академик Пресс. ISBN 978-1-904455-48-6 .
- ^ Брэди, Ричард М. (1993). «Крутящий момент и переключение бактериального жгутикового двигателя. Электростатическая модель» . Биофизический журнал . 64 (4): 961–973. Бибкод : 1993BpJ....64..961B . дои : 10.1016/S0006-3495(93)81462-0 . ПМЦ 1262414 . ПМИД 7684268 .
- ^ Штрайф С., Штаудингер В.Ф., Марван В., Остерхельт Д. (2008). «Вращение жгутиков у архей Halobacterium salinarum зависит от АТФ». Журнал молекулярной биологии . 384 (1): 1–8. дои : 10.1016/j.jmb.2008.08.057 . ПМИД 18786541 .
- ^ Перейти обратно: а б Скеркер, Дж. М.; Берг, ХК (5 июня 2001 г.). «Прямое наблюдение за расширением и втягиванием пилей IV типа» . Труды Национальной академии наук Соединенных Штатов Америки . 98 (12): 6901–6904. Бибкод : 2001PNAS...98.6901S . дои : 10.1073/pnas.121171698 . ISSN 0027-8424 . ПМК 34450 . ПМИД 11381130 .
- ^ Мэттик, Джон С. (2002). «Пили IV типа и подергивания моторики». Ежегодный обзор микробиологии . 56 : 289–314. дои : 10.1146/annurev.micro.56.012302.160938 . ISSN 0066-4227 . ПМИД 12142488 .
- ^ Мерц, AJ; Итак, М.; Шитц, член парламента (7 сентября 2000 г.). «Втягивание пилуса приводит к подергивающейся подвижности бактерий». Природа . 407 (6800): 98–102. Бибкод : 2000Natur.407...98M . дои : 10.1038/35024105 . ISSN 0028-0836 . ПМИД 10993081 . S2CID 4425775 .
- ^ Хенриксен, Дж. (декабрь 1972 г.). «Бактериальная поверхностная транслокация: обзор и классификация» . Бактериологические обзоры . 36 (4): 478–503. дои : 10.1128/BR.36.4.478-503.1972 . ISSN 0005-3678 . ПМК 408329 . ПМИД 4631369 .
- ^ Нан, Бэйян (февраль 2017 г.). «Скользящая подвижность бактерий: разработка консенсусной модели» . Современная биология . 27 (4): Р154–Р156. Бибкод : 2017CBio...27.R154N . дои : 10.1016/j.cub.2016.12.035 . ПМИД 28222296 .
- ^ Нан, Бэйян; Макбрайд, Марк Дж.; Чен, Цзин; Зусман, Дэвид Р.; Остер, Джордж (февраль 2014 г.). «Бактерии, скользящие по спиральным дорожкам» . Современная биология . 24 (4): 169–174. Бибкод : 2014CBio...24.R169N . дои : 10.1016/j.cub.2013.12.034 . ПМЦ 3964879 . ПМИД 24556443 .
- ^ Сибли, Л.Дэвид; Хоканссон, Себастьян; Каррутерс, Верн Б. (1 января 1998 г.). «Скользящая подвижность: эффективный механизм проникновения в клетки» . Современная биология . 8 (1): С12–Р14. Бибкод : 1998CBio....8..R12S . дои : 10.1016/S0960-9822(98)70008-9 . ПМИД 9427622 . S2CID 17555804 .
- ^ Сибли, LDI (октябрь 2010 г.). «Как апикомплексные паразиты проникают в клетки и выходят из них» . Curr Opin Biotechnol . 21 (5): 592–8. дои : 10.1016/j.copbio.2010.05.009 . ПМЦ 2947570 . ПМИД 20580218 .
- ^ Харши, Расика М. (1 января 2003 г.). «Подвижность бактерий на поверхности: множество путей к общей цели». Ежегодный обзор микробиологии . 57 (1): 249–73. дои : 10.1146/annurev.micro.57.030502.091014 . ПМИД 14527279 .
- ^ Хенрихсен, Дж (1972). «Бактериальная поверхностная транслокация: обзор и классификация» . Бактериологические обзоры . 36 (4): 478–503. дои : 10.1128/BR.36.4.478-503.1972 . ПМК 408329 . ПМИД 4631369 .
- ^ «Лабораторное руководство BIOL 230: Неподвижные бактерии в подвижной среде» . факультет.ccbcmd.edu . Архивировано из оригинала 15 апреля 2017 года . Проверено 31 мая 2020 г.
- ^ Посфаи М., Лефевр К., Трубицын Д., Базылински Д.А. и Франкель Р. (2013) «Филогенетическое значение состава и кристаллической морфологии минералов магнитосом». Границы микробиологии , 4 : 344. два : 10.3389/fmicb.2013.00344 . Измененный текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 3.0 .
- ^ Линь, Вэй; Чжан, Вэньси; Чжао, Сян; Робертс, Эндрю; Патерсон, Грейг; Базылински, Деннис; Пан, Юнсинь (март 2018 г.). «Геномная экспансия магнитотаксисных бактерий показывает раннее общее происхождение магнитотаксиса с эволюцией, специфичной для каждой линии» . Журнал ISME . 12 (6): 1508–1519. Бибкод : 2018ISMEJ..12.1508L . дои : 10.1038/s41396-018-0098-9 . ПМЦ 5955933 . ПМИД 29581530 .
- ^ Перейти обратно: а б Дюсенбери, Дэвид Б. (2009). Жизнь на микроуровне: неожиданная физика маленького размера . Кембридж, Массачусетс: Издательство Гарвардского университета. стр. 100–101. ISBN 978-0-674-03116-6 .
- ^ Перейти обратно: а б Чжан, У.Дж. и Ву, Л.Ф., 2020. Жгутики и плавательное поведение морских магнитотактических бактерий. Биомолекулы, 10(3), стр.460. два : 10.3390/biom10030460 . Измененный текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Джоглер, К.; Ваннер, Г.; Колинько С.; Ниблер, М.; Аманн, Р.; Петерсен, Н.; Кубе, М.; Рейнхардт, Р.; Шулер, Д. (2010). «Консервация генов и структур протеобактериальных магнитосом у некультивируемых представителей глубоко разветвленного типа Nitrospira» . Труды Национальной академии наук . 108 (3): 1134–1139. Бибкод : 2011PNAS..108.1134J . дои : 10.1073/pnas.1012694108 . ПМК 3024689 . ПМИД 21191098 .
- ^ Монтей, Кэролайн Л.; Валленет, Дэвид; Менги, Николас; Бензерара, Карим; Барб, Валери; Футо, Стефани; Круо, Коринн; Флориани, Магали; Виолье, Эрик; Адрианчик, Жеральдин; Леонхардт, Натали (июль 2019 г.). «Эктосимбиотические бактерии в основе магниторецепции у морских простейших» . Природная микробиология . 4 (7): 1088–1095. дои : 10.1038/s41564-019-0432-7 . ISSN 2058-5276 . ПМК 6697534 . ПМИД 31036911 .
- ^ Натан, Эвиатар; Фитак, Роберт Роджерс; Вербер, Юваль; Вортман, Йони (28 сентября 2020 г.). «Симбиотическое магнитное зондирование: сбор доказательств и не только» . Философские труды Королевского общества B: Биологические науки . 375 (1808): 20190595. doi : 10.1098/rstb.2019.0595 . ПМЦ 7435164 . ПМИД 32772668 .
- ^ Копп, Р.Э. и Киршвинк, Дж.Л. (2007). «Идентификация и биогеохимическая интерпретация ископаемых магнитотактических бактерий» (PDF) . Обзоры наук о Земле . 86 (1–4): 42–61. Бибкод : 2008ESRv...86...42K . doi : 10.1016/j.earscirev.2007.08.001 .
- ^ Чанг, С.Р. и Дж.Л. Киршвинк (1989). «Магнитофоссилии, намагниченность отложений и эволюция биоминерализации магнетита». Ежегодный обзор наук о Земле и планетах . 17 : 169–195. Бибкод : 1989AREPS..17..169C . doi : 10.1146/annurev.ea.17.050189.001125 .
- ^ Уолсби А.Е. (1969). «Проницаемость газовакуольных мембран сине-зеленых водорослей для газа». Труды Лондонского королевского общества. Серия Б, Биологические науки . 173 (1031): 235–255. Бибкод : 1969RSPSB.173..235W . дои : 10.1098/rspb.1969.0049 . JSTOR 75817 . ОСЛК 479422015 . S2CID 95321956 .
- ^ Каланетра К.М., Хьюстон С.Л., Нельсон, округ Колумбия (декабрь 2004 г.). «Новые прикрепленные сероокисляющие бактерии в неглубоких гидротермальных источниках обладают вакуолями, не участвующими в накоплении нитратов в дыхательных путях» . Прикладная и экологическая микробиология . 70 (12): 7487–96. Бибкод : 2004ApEnM..70.7487K . дои : 10.1128/АЕМ.70.12.7487-7496.2004 . ПМЦ 535177 . ПМИД 15574952 .
- ^ Шульц-Фогт Х.Н. (2006). «Вакуоли». Включения у прокариот . Монографии по микробиологии. Том. 1. С. 295–298. дои : 10.1007/3-540-33774-1_10 . ISBN 978-3-540-26205-3 .
- ^ Монтанчес, Ичасо; Огаяр, Эликсабет; Плагаро, Андер Эрнандес; Эстев-Кодина, Анна; Гомес-Гарридо, Джессика; Орруньо, Майте; Арана, Инес; Кабердин, Владимир Робертович (2019). «Анализ адаптации Vibrio harveyi в микрокосмах морской воды при повышенной температуре дает представление о предполагаемых механизмах его сохранения и распространения во время глобального потепления» . Научные отчеты . 9 (1): 289. Бибкод : 2019НатСР...9..289М . дои : 10.1038/s41598-018-36483-0 . ПМК 6343004 . ПМИД 30670759 . S2CID 58950215 .
- ^ Макфолл-Нгай, Маргарет; Хит-Хекман, Элизабет AC; Джилетт, Амани А.; Пейер, Сюзанна М.; Харви, Элизабет А. (2012). «Тайные языки совместно развившихся симбиозов: выводы из симбиоза сколопов Euprymna и Vibrio fischeri» . Семинары по иммунологии . 24 (1): 3–8. дои : 10.1016/j.smim.2011.11.006 . ПМК 3288948 . ПМИД 22154556 .
- ^ Уотерс, Кристофер М.; Басслер, Бонни Л. (7 октября 2005 г.). «ЗОНДИРОВАНИЕ КВОРУМА: Межклеточная связь у бактерий». Ежегодный обзор клеточной биологии и биологии развития . 21 (1): 319–346. doi : 10.1146/annurev.cellbio.21.012704.131001 . ПМИД 16212498 .
- ^ Янг, Р.; Ропер, К. (1976). «Биолюминесцентное затенение у средневодных животных: данные по живым кальмарам». Наука . 191 (4231): 1046–1048. Бибкод : 1976Sci...191.1046Y . дои : 10.1126/science.1251214 . ПМИД 1251214 .
- ^ Оуэнс, Ли; Бусико-Сальседо, Нанси (2006). «Vibrio harveyi: Симпатичные проблемы в раю (глава 19)». В Томпсоне, Фабиано; Остин, Брайан; Качели, Джин (ред.). Биология вибрионов . АСМ Пресс.
- ^ Делонг, EF; Бежа, О. (2010). «Светоуправляемый протонный насос протеородопсин повышает выживаемость бактерий в трудные времена» . ПЛОС Биология . 8 (4): e1000359. дои : 10.1371/journal.pbio.1000359 . ПМК 2860490 . ПМИД 20436957 . е1000359.
- ^ Перейти обратно: а б Гомес-Консарнау, Л.; Рэйвен, Дж.А.; Левин, Нью-Мексико; Каттер, Л.С.; Ван, Д.; Сигерс, Б.; Аристеги, Ж.; Фурман, Дж. А.; Газоль, Дж. М.; Саньюдо-Вильгельми, SA (2019). «Микробные родопсины вносят основной вклад в солнечную энергию, улавливаемую в море» . Достижения науки . 5 (8): eaaw8855. Бибкод : 2019SciA....5.8855G . doi : 10.1126/sciadv.aaw8855 . ПМЦ 6685716 . ПМИД 31457093 .
- ^ Орен, Аарон (2002). «Молекулярная экология чрезвычайно галофильных архей и бактерий» . ФЭМС Микробиология Экология . 39 (1): 1–7. дои : 10.1111/j.1574-6941.2002.tb00900.x . ПМИД 19709178 .
- ^ Бежа, О.; Аравинд, Л.; Кунин Е.В.; Сузуки, Монтана; Хадд, А.; Нгуен, LP; Йованович, С.Б.; Гейтс, КМ; Фельдман, РА; Спудич, Дж.Л.; Спудич, Э.Н. (2000). «Бактериальный родопсин: свидетельства нового типа фототрофии в море». Наука . 289 (5486): 1902–1906. Бибкод : 2000Sci...289.1902B . дои : 10.1126/science.289.5486.1902 . ПМИД 10988064 . S2CID 1461255 .
- ^ «Интервью с коллегами: Эд Делонг» . Американская академия микробиологии . Архивировано из оригинала 7 августа 2016 года . Проверено 2 июля 2016 г.
- ^ Бактерии с батарейками , Popular Science, январь 2001 г., стр. 55.
- ^ Бежа, О.; Аравинд, Л.; Кунин Е.В.; Сузуки, Монтана; Хадд, А.; Нгуен, LP; Йованович, С.Б.; Гейтс, КМ; Фельдман, РА; Спудич, Дж.Л.; Спудич, Э.Н. (2000). «Бактериальный родопсин: свидетельства нового типа фототрофии в море». Наука . 289 (5486): 1902–1906. Бибкод : 2000Sci...289.1902B . дои : 10.1126/science.289.5486.1902 . ПМИД 10988064 .
- ^ Бёф, Доминик; Аудик, Стефан; Брилье-Геген, Лорейн; Кэрон, Кристоф; Жантон, Кристиан (2015). «MicRhoDE: тщательно подобранная база данных для анализа разнообразия и эволюции микробного родопсина» . База данных . 2015 : bav080. дои : 10.1093/база данных/bav080 . ПМК 4539915 . ПМИД 26286928 .
- ^ Яво, Хирому; Кандори, Хидеки; Коидзуми, Амане (5 июня 2015 г.). Оптогенетика: светочувствительные белки и их применение . Спрингер. стр. 3–4. ISBN 978-4-431-55516-2 . Проверено 30 сентября 2015 г.
- ^ Крошечный морской микроб может сыграть большую роль в изменении климата Университет Южной Калифорнии , Пресс-центр, 8 августа 2019 г.
- ^ ДасСарма, Шиладитья; Швитерман, Эдвард В. (11 октября 2018 г.). «Ранняя эволюция фиолетовых пигментов сетчатки на Земле и значение биосигнатур экзопланеты». Международный журнал астробиологии . 20 (3): 241–250. arXiv : 1810.05150 . дои : 10.1017/S1473550418000423 . ISSN 1473-5504 . S2CID 119341330 .
- ^ Спаркс, Уильям Б.; ДасСарма, С.; Рид, Индиана (декабрь 2006 г.). «Эволюционная конкуренция между примитивными фотосинтетическими системами: существование ранней фиолетовой Земли?». Тезисы докладов о заседании Американского астрономического общества . 38 : 901. Бибкод : 2006AAS...209.0605S .
- ^ Дэйн Коноп (29 июля 1997 г.). «Ученые обнаружили метановых ледяных червей на дне Мексиканского залива» . Национальное управление океанических и атмосферных исследований . Архивировано из оригинала 9 июня 2010 года . Проверено 22 января 2010 г.
- ^ Шарп, Коти Х.; Пратте, Зои А.; Кервин, Эллисон Х.; Ротьян, Рэнди Д.; Стюарт, Фрэнк Дж. (2017). «Структурой микробиома коралла Astrangia poculata умеренного пояса управляет сезон, а не состояние симбионта» . Микробиом . 5 (1): 120. дои : 10.1186/s40168-017-0329-8 . ПМК 5603060 . ПМИД 28915923 .
- ^ Лема, К.А., Уиллис, Б.Л. и Борн, Д.Г. (2012) «Кораллы образуют характерные ассоциации с симбиотическими азотфиксирующими бактериями». Прикладная и экологическая микробиология , 78 (9): 3136-3144. дои : 10.1128/AEM.07800-11 .
- ^ Петерсен, Джиллиан М.; Фрэнк У. Зелински; Томас Пейп; Ричард Зейферт; Кристина Морару; и др. (11 августа 2011 г.). «Водород является источником энергии для симбиозов гидротермальных источников». Природа . 476 (7359): 176–180. Бибкод : 2011Natur.476..176P . дои : 10.1038/nature10325 . ПМИД 21833083 . S2CID 25578 .

- ^ Кляйнер, Мануэль; Вентрап, Сесилия; Лотт, Кристиан; Тилинг, Ханно; Ветцель, Силке; Янг, Жак; Чанг, Юн-Хуан; Шах, Манеш; ВерБеркмоэс, Натан К. (8 мая 2012 г.). «Метапротеомика бескишечного морского червя и его симбиотического микробного сообщества открывает необычные пути использования углерода и энергии» . Труды Национальной академии наук Соединенных Штатов Америки . 109 (19): E1173–E1182. дои : 10.1073/pnas.1121198109 . ПМЦ 3358896 . ПМИД 22517752 .
- ^ Войке, Таня; Тилинг, Ханно; Иванова Наталья Н.; Хантеманн, Марсель; Рихтер, Майкл; Глекнер, Фрэнк Оливер; Боффелли, Дарио; Андерсон, Иэн Дж.; Барри, Керри В. (26 октября 2006 г.). «Понимание симбиоза посредством метагеномного анализа микробного консорциума» (PDF) . Природа . 443 (7114): 950–955. Бибкод : 2006Natur.443..950W . дои : 10.1038/nature05192 . ПМИД 16980956 . S2CID 140106758 .
- ^ Ученые Геологической службы США публикуют давно читаемые последовательности микробиома кораллов умеренного пояса, предоставляя сообществу ресурсы для разработки зондов и праймеров , Геологическая служба США , 6 марта 2019 г. Измененный текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ б. Голдсмит, Дон; а. Пратте, Зоя; а. Келлог, Кристина; е. Снадер, Сара; час Шарп, Коти (2019). «Стабильность микробиома коралла Astrangia poculata умеренного пояса отражается в различных методологиях секвенирования» . АИМС Микробиология . 5 (1): 62–76. дои : 10.3934/микробиол.2019.1.62 . ПМК 6646935 . ПМИД 31384703 . Измененный текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Перейти обратно: а б Руис-Гонсалес К., Симо Р., Соммаруга Р. и Газоль Дж. М. (2013) «Вдали от темноты: обзор влияния солнечной радиации на активность гетеротрофного бактериопланктона». Границы микробиологии , 4 : 131. два : 10.3389/fmicb.2013.00131 . Измененный текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 3.0 .
- ^ Уокер, JCG (1980). Круговорот кислорода в природной среде и биогеохимические циклы. Берлин: Springer-Verlag.
- ^ Лонгхерст А., Сатиендранат С., Платт Т. и Каверхилл К. (1995). Оценка глобальной первичной продукции в океане по данным спутникового радиометра. Дж. Планктон Рез. 17, 1245–1271.
- ^ Перейти обратно: а б с д и Курата Н., Велла К., Гамильтон Б., Шивджи М., Соловьев А., Мэтт С., Тартар А. и Перри В. (2016) «Бактерии, связанные с поверхностно-активными веществами, в ближайшем будущем». -поверхностный слой океана». Научные отчеты , 6 (1): 1–8. дои : 10.1038/srep19123 . Измененный текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Хутич В., Чосович Б., Марченко Э., Бихари Н. и Кршинич Ф. (1981) «Производство поверхностно-активных веществ морским фитопланктоном». Морская химия , 10 (6): 505–520. дои : 10.1016/0304-4203(81)90004-9 .
- ^ Перейти обратно: а б Сигел, Дэвид А.; Бюсселер, Кен О.; Беренфельд, Майкл Дж.; Бенитес-Нельсон, Клаудия Р.; Босс, Эммануэль; Бжезинский, Марк А.; Бурд, Адриан; Карлсон, Крейг А.; д'Асаро, Эрик А.; Дони, Скотт С.; Перри, Мэри Дж.; Стэнли, Рэйчел HR; Стейнберг, Дебора К. (2016). «Прогнозирование экспорта и судьбы чистой первичной продукции мирового океана: научный план экспорта» . Границы морской науки . 3 . дои : 10.3389/fmars.2016.00022 . hdl : 1912/8770 . Измененный текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Лю X, Пань Дж, Лю Ю, Ли М, Гу Дж.Д. (октябрь 2018 г.). «Разнообразие и распространение архей в глобальных эстуарных экосистемах». Наука об общей окружающей среде . 637–638: 349–358. Бибкод : 2018ScTEn.637..349L . doi : 10.1016/j.scitotenv.2018.05.016 . ПМИД 29753224 . S2CID 21697690 .
- ^ Кабельо П., Ролдан, доктор медицинских наук, Морено-Вивиан С. (ноябрь 2004 г.). «Редукция нитратов и круговорот азота у архей» . Микробиология . 150 (Часть 11): 3527–46. дои : 10.1099/mic.0.27303-0 . ПМИД 15528644 .
- ^ Мехта, член парламента, Баросс Дж.А. (декабрь 2006 г.). «Фиксация азота при 92 градусах Цельсия археями гидротермальных источников». Наука . 314 (5806): 1783–86. Бибкод : 2006Sci...314.1783M . дои : 10.1126/science.1134772 . ПМИД 17170307 . S2CID 84362603 .
- ^ Фрэнсис К.А., Беман Дж.М., Кайперс М.М. (май 2007 г.). «Новые процессы и участники круговорота азота: микробная экология анаэробного и архейного окисления аммиака» . Журнал ISME . 1 (1): 19–27. Бибкод : 2007ISMEJ...1...19F . дои : 10.1038/ismej.2007.8 . ПМИД 18043610 .
- ^ Кулен М.Дж., Аббас Б., ван Блейсвейк Дж., Хопманс Э.К., Кайперс М.М., Уэйкхэм С.Г., Синнингхе Дамсте Дж.С. и др. (апрель 2007 г.). «Предполагаемая кренархеота, окисляющая аммиак, в субкислородных водах Черного моря: экологическое исследование всего бассейна с использованием 16S рибосомных и функциональных генов и мембранных липидов». Экологическая микробиология . 9 (4): 1001–16. Бибкод : 2007EnvMi...9.1001C . дои : 10.1111/j.1462-2920.2006.01227.x . HDL : 1912/2034 . ПМИД 17359272 .
- ^ Бейкер Б.Дж., Банфилд Дж.Ф. (май 2003 г.). «Микробные сообщества в кислых шахтных дренажах» . ФЭМС Микробиология Экология . 44 (2): 139–52. Бибкод : 2003FEMME..44..139B . дои : 10.1016/S0168-6496(03)00028-X . ПМИД 19719632 .
- ^ Шимель Дж. (август 2004 г.). «Игра весов в метановом цикле: от микробной экологии до земного шара» . Труды Национальной академии наук Соединенных Штатов Америки . 101 (34): 12400–01. Бибкод : 2004PNAS..10112400S . дои : 10.1073/pnas.0405075101 . ПМК 515073 . ПМИД 15314221 .
Библиография
[ редактировать ]- Далримпл, Дж. Брент (2001). «Возраст Земли в двадцатом веке: проблема (в основном) решена». Специальные публикации Лондонского геологического общества . 190 (1): 205–221. Бибкод : 2001GSLSP.190..205D . дои : 10.1144/ГСЛ.СП.2001.190.01.14 . S2CID 130092094 .
- Рэйвен, Питер Гамильтон; Джонсон, Джордж Брукс (2002). Биология . Макгроу-Хилл Образование. ISBN 978-0-07-112261-0 .
- Шорс, Тери (2017). Понимание вирусов . Издательство Джонс и Бартлетт. ISBN 978-1284025927 .
